")
Back to Journals » Cancer Management and Research » Volume 12
Regulatory Roles of Tumor Necrosis Factor-α-Induced Protein 8 Like-Protein 2 in Inflammation, Immunity and Cancers: A Review
Authors Gu Z , Cui X, Sun P, Wang X
Received 25 September 2020
Accepted for publication 15 November 2020
Published 14 December 2020 Volume 2020:12 Pages 12735—12746
DOI https://doi.org/10.2147/CMAR.S283877
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eileen O'Reilly
Zhengzhong Gu,1 Xiaohan Cui,2 Pengda Sun,1 Xudong Wang1
1Department of Gastrointestinal Nutrition and Hernia Surgery, The Second Hospital of Jilin University, Changchun, Jilin Province, People’s Republic of China; 2Nanjing Medical University, Nanjing, Jiangsu Province, People’s Republic of China
Correspondence: Xudong Wang
Department of Gastrointestinal Nutrition and Hernia Surgery, The Second Hospital of Jilin University, No. 218, Ziqiang Street, Changchun 130041 Jilin Province, People’s Republic of China
Tel +86 13514406378
Email [email protected]
Abstract: Tumor necrosis factor-alpha (TNF-α)-induced protein 8 (TNFAIP8/TIPE) family, including TNFAIP8 (TIPE), TNFAIP8 like-protein 1 (TNFAIP8L1/TIPE1), TNFAIP8 like-protein 2 (TNFAIP8L2/TIPE2), and TNFAIP8 like-protein 3 (TNFAIP8L3/TIPE3), plays a vital role in regulating inflammatory responses, immune homeostasis, and cancer development. Over the last decade, studies have shown that TIPE2 protein is differentially expressed in diverse cells and tissues. The dysregulation of TIPE2 protein can lead to dysregulation of inflammatory responses and immune homeostasis, and change the basic characteristics of cancers. In consideration of the immeasurable values of TIPE2 in diagnosis, treatment, and prognosis of various human diseases, this review will focus on the expression pattern, structure, and regulatory roles of TIPE2 in inflammation, immunity, and cancers.
Keywords: TIPE2, inflammation, immune homeostasis, tumor, tumorigenesis, metastasis
Introduction
It was reported that chronic inflammation contributes to tumor pathogenesis.1 Immune cells can respond to inflammatory stimulations, secreting tumor necrosis factor-alpha (TNF-α).2 TNF-α can bind to tumor necrosis factor receptor type 1 (TNFR1) and tumor necrosis factor receptor type 2 (TNFR2), activating nuclear factor-κB (NF-κB) signaling pathway and inducing the expression of tumor necrosis factor-α-induced protein 8 (TNFAIP8/TIPE) family proteins, including TNFAIP8 (TIPE), TNFAIP8 like-protein 1 (TNFAIP8L1/TIPE1), TNFAIP8 like-protein 2 (TNFAIP8L2/TIPE2), and TNFAIP8 like-protein 3 (TNFAIP8L3/TIPE3).3 All these four proteins consist of a death effector domain (DED), except for this, there is no significant sequence similarity with other proteins.4–7 TIPE family proteins are highly similar in structure, with approximately 54% homology and 75% amino acid sequence similarities. All four members comprise a homologous domain, which includes several α-helices and a highly conserved hydrophobic cavity.8 Nevertheless, all members exhibit significantly differential expressions and seem to play diverse roles in different biological activities among different cells and tissues. Previous studies suggest that TIPE and TIPE3 proteins promote cell viability and induce drug resistance which ultimately facilitate the development and progression of cancers,6,9 and meanwhile, TIPE is also a risk factor of bacterial infection and TIPE3 serves as a translocator of lipid second messengers.6,10 In contrast, TIPE1 and TIPE2 are associated with cell apoptosis and antitumorigenesis.11,12 Particularly, TIPE2 also acts as a regulator in inflammatory responses and immune homeostasis.7
Although Sun et al7 initially introduced TIPE2 and its regulatory role in maintaining immune homeostasis in 2008, the newfound functions and mechanisms of TIPE2 in inflammation, immunity, and tumors have not been comprehensively summarized. In consideration of the new vital findings of TIPE2 in regulating inflammation, immunity and cancer, it is of great value to learn more about it. This current review aimed to summarize the expression pattern, structure, and regulatory functions of TIPE2 in inflammation, immunity and cancers based on the latest literature.
TIPE Proteins: TNFAIP8, TIPE1, and TIPE3 and Functions in Human Diseases
TNF-α is a cellular signaling regulator, which modulates inflammatory response.13 TNF-α binds to TNFR1 and TNFR2, activating NF-κB and inducing the expression of TIPE proteins. All four TIPE proteins mostly exist in cytoplasm, and TIPE2 is also reported to localize in nucleus.3 The amino acid sequence comparison of TIPE proteins showed that C-terminal residues are significantly conserved and N-terminal residues are highly varied. The brief functions of TNFAIP8, TIPE1, and TIPE3 in human diseases are discussed in the following sections.
TNFAIP8
TNFAIP8 was found in head and neck squamous cell carcinoma (HNSCC) cell lines and is the first identified TIPE family protein.14 TNFAIP8 is expressed in most human tissues. Expression analysis of TNFAIP8 showed that it is mainly expressed in bone marrow, immune system, gastrointestinal tract, lung and adipose tissues.15 TNFAIP8 also exists in epididymis, seminal vesicles, testis, and prostate, and meanwhile, it is expressed in fallopian tube, cervix, and endometrium in women.15
TNFAIP8 inhibits cell apoptosis and facilitates cell viability. TNFAIP8 overexpression in MDA-MB-435 cells of breast cancer enhanced the migration of tumor cells via upregulation of vascular endothelial growth factor receptor-2 (VEGFR-2), matrix metalloproteinase 1 (MMX-1), and MMX-9.16 In non-small-cell lung cancer (NSCLC), increased expression of TNFAIP8 was observed in tumor tissues, and TNFAIP8 promoted tumor cell proliferation and cisplatin resistance through murine double minute 2 (MDM2)/p53 pathway.17 Moreover, TNFAIP8 reduced the phosphorylation of large tumor suppressor gene 1 (LATS1), increasing the expression of Yes-associated protein (YAP), ultimately enhancing the proliferation and invasion of NSCLC cells.18 TNFAIP8 also acted as a tumorigenic gene in hepatocellular carcinoma (HCC) via LATS1-YAP signaling pathway.19 TNFAIP8 overexpression resulted in decreased fatty acid oxidation genes’ expression and increased several tumor genes’ expression, such as nuclear factor of activated T-cells 5 (NFAT5), metastasis-associated lung adenocarcinoma transcript 1 (MALAT1), forkhead box protein A1 (FOXA1), and kirsten rat sarcoma viral oncogene (KRAS).9 In addition to cancer, Porturas et al20 also revealed the biological role of TNFAIP8 in infection. TNFAIP8 regulated Listeria monocytogenes infection via modulating activity of Ras-related C3 botulinum toxin substrate 1 (Rac1).20
TIPE1
TIPE1 was first identified in 2011 when specific antibody was produced. TIPE1 is expressed in various cell types, including hepatocytes, intestinal epithelial cells, muscle tissues, neurons, and germ cells, but is totally absent in mature B lymphocytes and T lymphocytes.21
TIPE1 mostly plays an antineoplastic role in most cancers. The interaction between TIPE1 and Rac1 promotes tumor cell apoptosis via inhibiting C-Jun N-terminal kinase (JNK) and p65 activity in primary liver cancer. Moreover, the expression of TIPE1 was negatively associated with lymph node metastasis of primary liver cancer.11 In mice models of NSCLC, TIPE1 inhibited tumor growth and facilitated cell apoptosis, indicating that TIPE1 could serve as a tumor suppressor of NSCLC.22 TIPE1 was capable of inhibiting epithelial-mesenchymal transition (EMT) via modulating Wnt/β-catenin signaling and downregulating MMP-2 and MMP-9 expression, which ultimately suppressed the proliferation and migration of gastric cancer.23 Additionally, in breast cancer cells, TIPE1 suppressed the proliferation, invasion, metastasis, and EMT via downregulating the phosphorylation of extracellular signal-regulated protein kinase (ERK).24 In osteosarcoma, TIPE1 was found to suppress the activity of monocyte chemotactic protein-1 (MCP-1) to reduce macrophage infiltration, thus inhibiting the proliferation of osteosarcoma cells[38].25 In hematological diseases, TIPE1 was observed to promote cell apoptosis and inhibit tumor growth in RAW264.7 cells by upregulating pro-apoptotic members of the B cell lymphoma/leukemia-2 (Bcl-2) family.26 However, TIPE1 was also observed to enhance the tumorigenicity of cervical cancer cells and promote cancer progression via inhibiting p53 acetylation, suggesting that TIPE1 serves as an oncogene in cervical cancer.27 Generally, TIPE1 induces cell apoptosis and inhibits tumorigenesis and progression in most cancers.
TIPE3
TIPE3 is mainly expressed in secretory epithelium and serves as a carcinogenic molecule.6 TIPE3 is capable of shuttling phosphatidylinositol bisphosphate (PIP2) and phosphatidylinositol trisphosphate (PIP3) to the plasma membrane which can enhance phosphatidylinositol-3-kinase (PI3K)-mediated signaling transduction, thus promoting the occurrence and development of cancers.6 The increased expression of TIPE3 was identified in colon cancer, NSCLC, breast cancer, esophageal cancer, gastric cancer, and malignant glioma.28 Knockout of TIPE3 can inhibit tumor proliferation, while its overexpression can enhance tumor aggressiveness.28 TIPE3 was found to activate PI3K/protein kinase B (AKT) signaling pathway to facilitate tumor cell proliferation and migration in gastric cancer.29 TIPE3 also promoted cell metastasis by activating AKT and NF-κB signals in breast cancer.30 In addition, TIPE3 promoted the progression of malignant glioma via inhibiting p38 phosphorylation.31 However, it was also reported that TIPE3 inhibited the proliferation and invasion of nasopharyngeal carcinoma cells.32
By introducing the brief biological functions of three of the TIPE family proteins, we have already laid the foundation to summarize the expression pattern, structure, and regulatory functions of TIPE2 in inflammation, immunity, and cancers based on the latest literature.
TIPE2 Expression Pattern
It is known that TIPE2 gene is located on chromosome 1(1q21.2–1q21.3) and its protein was first identified in autoimmune encephalomyelitis in 2008.7 TIPE2 protein was reported to mainly exist in cytoplasm. The expression analysis of TIPE2 suggested that it followed a tissue-specific expression pattern. TIPE2 was mainly expressed in lymphoid and myeloid tissues, and constitutively expressed in immune cells.7 The subsequent studies of TIPE2 expression showed that it was primarily expressed in T lymphocytes, but not B lymphocytes.33 Surprisingly, TIPE2 was also detected in endocrine and reproductive cells of mice, suggesting its regulatory roles in endocrine and reproductive systems.34,35 Besides, different from the expression pattern in mice, the expression of human TIPE2 was typically observed in non-hematopoietic cells.33,35,36 However, high expression of TIPE2 was only found in macrophage-derived cells and few cancer cells, such as renal cell carcinoma (RCC) and skin squamous cell carcinoma (SSCC), while in most cancer cells, the expression of TIPE2 was low or undetectable, including bladder cancer, breast cancer, gastric cancer, NSCLC, prostate cancer and rectal cancer, indicating that it was primarily expressed in mononuclear cells and epithelial-derived secretory cells.34
TIPE2 Structure
As shown by the crystal structure of TIPE2, a highly conserved hydrophobic cavity is located in the center, which is considered to be the binding site for cofactors like phospholipid or lipid second messengers.37 Interestingly, these cofactors share similarities in binding pattern, which is exposing inositol head group and inserting lipid tail into the hydrophobic cavity.38 Moreover, TIPE2 also consists of six cylindrical antiparallel α-helices, which encircle the conserved hydrophobic cavity. Of these α-helices, α5-helix is divided by Pro153 into two segments, α5a and α5b37 (Figure 1). However, the high-resolution structure of TIPE2 exhibits a particular and uncharacterized fold, implying that TIPE2 has a unique topological structure which is quite different from the DED structure of other members of TIPE family.37 It was demonstrated that there are 184 amino acids in TIPE2 structure, which is much more than those in ordinary DED (approximately 90 amino acids). Furthermore, the N-to-C topological structure of TIPE2 is perfectly consistent with the C-to-N topological structure of ordinary DED. Therefore, the topological structure of TIPE2 appears to be a mirror diagram of ordinary DED.37,39
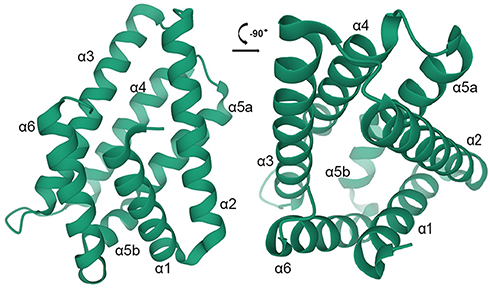 |
Figure 1 The structure of TIPE2 is shown in two vertical views. The six a-helices are colored in green. |
Upstream Regulating Factors of TIPE2
TIPE2 is the intermediary molecule in the inhibition of cell apoptosis mediated by microRNA-21, while microRNA-21 is the direct target of NF-κB. TIPE2 expression is regulated by microRNA-21 via editing of the coding region. In activated T lymphocytes and macrophages, expression of microRNA-21 is highly upregulated, while TIPE2 expression is downregulated. Compared with microRNA-21 deficiency, T cells with TIPE2 deficiency are clearly insensitive to apoptosis. Therefore, it is obvious that microRNA-21 is the upstream regulator of TIPE240 (Figure 2). Moreover, TIPE2 was significantly upregulated in human HeLa cells transfected with OAS/RNase L-expressing VACV recombinants, signifying that RNase L was a transcriptional regulator of TIPE2.41 Furthermore, it was proven that activating protein-1 (AP-1) is an important transcriptional regulator of porcine TIPE2. Due to the conspicuous sequence similarity of TIPE2 between porcine and human, it is supposed that AP-1 may also be a regulatory factor in the transcription of human TIPE2.41
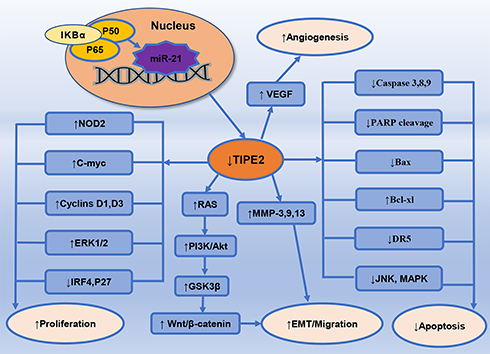 |
Figure 2 Downregulation of TIPE2 heightens activating EMT, proliferation, angiogenesis, migration, invasion, and metastasis of malignant cells, ultimately facilitating tumorigenesis and progression of cancers. ↑- upregulate; ↓- downregulate. |
Downstream Effect Factors of TIPE2
The negative immunomodulatory factor TIPE2 could reduce the activation and expression of transforming growth factor (TGF)-β-activated kinase 1 (Tak1) through blocking the Tak1-TAB1-TAB2 complex formation.42 The activation of caspase-8 induced by TIPE2 resulted in the reduced levels of AP-1 and NF-κB, which could inhibit caspase-1 activation and consequently promote Fas-induced cell apoptosis.43 TIPE2 also inhibited mitogen-activated protein kinase (MAPK) and NF-κB pathways via inhibiting nucleotide binding oligomerization domain-2 (NOD2), leading to the downregulation of NOD2-mediated inflammatory signals44 (Figure 2). In addition, TIPE2 was found to antagonize JNK, NF-κB, and p38MAPK pathways via hampering nuclear translocation of c-Fos, c-Jun and NF-κB and reduced the degradation of NF-κB inhibitor alpha (IKBα)7 (Figure 2). Regulatory T cells (Tregs) with TIPE2 deficiency were observed to secrete lower levels of cell surface markers like cytotoxic T-lymphocyte-associated protein-4 (CTLA-4) and forkhead box protein 3 (Foxp3), and anti-inflammatory cytokines like interleukin (IL)-10 and transforming growth factor (TGF)-β, leading to IL-2 dysregulation and enhanced NF-κB activation.45,46 In TIPE2-deficient myeloid cells, polyinosinic-polycytidylic acid (Poly(I:C))-mediated dsRNA signaling pathway was overactive, while Poly(I:C) was capable of downregulating TIPE2 expression mediated by downstream cytokines.47 Furthermore, TIPE2 was found to regulate Rac1-signal transducer and activator of transcription 3 (STAT3) and ERK1/2 signaling pathways and downregulate the expression levels of cyclin D1 and cyclin D3 in mice model of injury-induced restenosis disease48 (Figure 2).
Apart from what was clarified previously, TIPE2 also decreased the phosphorylation of PI3K and Akt by inhibiting the activation of c-myc, cyclin D1 and β-catenin, which led to inhibition of PI3K/Akt and Wnt/β-catenin signaling pathways49,50 (Figure 2). TIPE2 also participated in downregulating PI3K/Akt/glycogen synthase kinase 3β (GSK3β)-mediated β-catenin signaling via inhibiting the phosphorylation of Akt, which resulted in increased phosphorylation of GSK3β and eventually led to decreased nuclear translocation of β-catenin51 (Figure 2). In arthritic fibroblast-like synoviocytes, death receptor 5 (DR5) expression was upregulated by TIPE2, which could activate caspase, suppress NF-κB, and ultimately result in apoptosis of synoviocytes52 (Figure 2). Besides, it was proven that TIPE2 upregulated pro-apoptotic proteins’ expression such as Bcl-2 associated X (Bax), caspase-3, and caspase-9, facilitated the cleavage of poly ADP ribose polymerase (PARP), and downregulated the expression levels of anti-apoptotic proteins such as Akt, Bcl-xl and ERK1/253 (Figure 2). Moreover, it was demonstrated that TIPE2 could downregulate vascular endothelial growth factor (VEGF), suggesting its function of attenuating angiogenesis54 (Figure 2). Additionally, TIPE2 also played a role in the activation of interferon regulatory factor 4 (IRF4) signaling pathway to increase p27 expression and subsequently inhibited cell proliferation55(Figure 2).
Functions of TIPE2 in Inflammation and Immunity
TIPE2 is a crucial regulatory factor of inflammatory responses and immune homeostasis, which can suppress T cell receptor (TCR) and Toll-like receptor (TLR).7 Decreased expression of TIPE2 was associated with lethal inflammatory conditions in mice and autoimmune diseases in humans, suggesting its indispensable role in maintaining immune homeostasis.34 Knocking out TIPE2 in mice led to inflammation in multiple organs, splenomegaly, and even death.7 Besides, the loss of TIPE2 was also associated with higher serum levels of proinflammatory factors, such as IL-6, IL-17, IL-21, and TNF-α, and lower serum levels of anti-inflammatory factors, like IL-10 and TGF-β, resulting in enhanced proliferation and differentiation of T lymphocytes thus inducing inflammatory cells’ aggregation, overactive responses, and inflammatory diseases.4 On the contrary, TIPE2 selective expression was relevant in preventing hyperreactivity and maintaining immune homeostasis.56 In the differentiation of dendritic cells, TIPE2 activated PI3K-PKCδ-MAPK signaling pathway to increase the expression of CD80CD86mRNA, a marker of dendritic cells’ maturity, indicating that TIPE2 could enhance immune responses under homeostatic state by suppressing peripheral immune tolerance.45
In the mice model of clinical chronic kidney allograft rejection, TIPE2 was found to exhibit a high transcription level in peripheral blood and kidney biopsy samples, and significantly reduced immunological rejections, suggesting that TIPE2 could be a diagnostic biomarker in monitoring chronic kidney allograft rejection.57 A similar result was also observed in corneal allograft rejection and acute cardiac allograft rejection, implying the immunoregulatory role of TIPE2 in graft rejection.58,59 In rheumatoid arthritis mice models, TIPE2 was found to inhibit Rac activation and IRF3 phosphorylation, and thus significantly reduced proinflammatory cytokines’ expression in synovial fibroblasts stimulated by lipopolysaccharide; these foundings could help in designing novel strategies for the prevention and treatment of rheumatoid arthritis.60 Adding to this, DR5-caspase-NF-κB signaling pathway mediated by TIPE2 was also involved in anti-inflammatory responses in rheumatoid arthritis.52 Moreover, it was demonstrated that significantly amplified phosphorylation of JNK, p38, and IκBα was observed in TIPE2-deficient macrophages, and TIPE2 was found to participate in modulating L-arginase metabolism from nitric oxide to urea to suppress inflammatory responses.61 Inpatients with asthma and systemic lupus erythematosus, TIPE2 mRNA expression in peripheral blood mononuclear cells (PBMC) was notably decreased, meaning that TIPE2 serves as an anti-inflammatory regulator to reduce inflammation intensity.62,63 In chronic hepatitis B virus infection, the expression of TIPE2 was reduced and negatively correlated with the serum levels of virus load and hepatitis markers.64 The reduced TIPE2 expression was also identified in PBMC from patients with primary biliary cirrhosis, which enhanced monocytes’ sensitivity to TLR ligands.65
In the mice model of myocardial ischemia/reperfusion injury, it was proven that TIPE2 suppressed the activation of NOD2 and downstream factors, MAPK and NF-κB, inversely regulating NOD2-mediated inflammatory responses.44 Furthermore, TIPE2 was remarkably expressed in CD4+CD25+ Tregs of Bal b/c nude mice, and when TIPE2 was silenced by small interfering RNA (siRNA) or completely knocked out, CD4+CD25+ Tregs were capable of elevating T-cell proliferation and differentiation, revealing that TIPE2 was associated with the immunosuppressive function of CD4+CD25+ Tregs.66 TIPE2 also functioned as a suppressor of AP-1 and NF-κB via binding to and activating caspase-8, which could promote Fas-induced cell apoptosis.43 Stimulated by oxidized low density lipoprotein (OX-LDL), macrophages with TIPE2 deficiency exhibited amplified JNK, NF-κB, and p38MAPK signals, elevated the expression level of proinflammatory cytokines, and enhanced inflammatory responses.67 Consistent with this new finding, in the LDLR (-/-) mice with TIPE2 deficiency on a high-fat diet, the atherosclerosis formation was apparently aggravated and OX-LDL was found to downregulate the transcription of TIPE2 mRNA.67 In the mice model of injury-induced restenosis, TIPE2 overexpression was found to reduce activity of macrophages and impede proliferation and differentiation of vascular smooth muscle cells by blocking G 1/S phase transition via Rac1-STAT3 and ERK1/2 signaling pathways, consequently inhibiting vascular neointima and atherosclerosis formation.48,68 TIPE2 blocked the activation and nuclear translocation of STAT3 in a Rac1-dependent manner.48 These results prove TIPE2 to be an inhibitor of atherosclerosis and that it may serve as a therapeutic target for treating certain kinds of diseases.48,67,68 Interestingly, atorvastatin was shown to increase TIPE2 expression mediated by lipopolysaccharide in RAW264.7 cells, resulting in decreased expression of downstream inflammatory mediators, including nitric oxide synthase and NF-κB.69
Apart from these functions, TIPE2 also played a part in preventing stroke. When blocking the middle cerebral artery of mice with TIPE2 deficiency, the infarction volume, neurological dysfunction, and inflammatory cells’ infiltration in ischemic hemisphere were significantly exacerbated.70 Another study demonstrated that TIPE2 mRNA expression in survivors was much higher than that in dead, showing a prominent odds ratio on 3-month mortality.71 These findings implied that TIPE2 regulated inflammatory responses of stroke and showed an important neuroprotective effect on brain cells, suggesting its potential as a diagnostic and prognostic biomarker for acute ischemic stroke.70,71 Moreover, TIPE2 was also capable of modulating inflammation intensity by regulating macrophage polarization via suppressing mammalian target of rapamycin complex1 (mTORC1) activation.72 Interestingly, TIPE2 null mice were susceptible to pseudomonas aeruginosa (PA) infection and showed serious keratitis. Mechanistically, decreased inflammatory cell infiltration and NF-κB signaling were found to participate in TIPE2-mediated immunoregulation.73
Roles of TIPE2 in Cancers
TIPE2 can antagonize the oncogene Ras.12 It was capable of preventing Ras from forming a complex via binding to domains of RalGDS proteins, ultimately reducing the activation of downstream effectors Akt and Ral and maintaining dynamic balance between cell survival and apoptosis.12 Complete knockout of TIPE2 contributed to increased Akt and Ral activation, dysregulation of exocyst complex formation, enhanced cell proliferation, and reduced cell apoptosis. Conversely, elevated expression of TIPE2 promoted cell apoptosis and significantly prevented Ras-induced tumorigenesis, proving that it might be a potential suppressor and drug target for neoplastic diseases.12
Compared with the control group, TIPE2 was prominently upregulated, while myxoma resistance protein 1 (MX1) was downregulated in RCC cells and tissues74 (Table 1). Moreover, TIPE2 was negatively correlated with MX1 expression level and positively with Tumor Node Metastasis (TNM) stage, indicating its tumorigenic role in RCC pathogenesis74 (Table 1). A similar result was also observed in colon cancer samples and elevated expression of TIPE2 was positively associated with lymphatic metastasis and Dukes stage. TIPE2 inhibited caspase-8 activity and regulated TLR4-mediated inflammatory effects to promote the progression of colon cancer cells, suggesting that TIPE2 could be a new target for clinical colon cancer treatment43 (Table 1). TIPE2 expression level in rectal cancer tissues was also much higher than that in adjacent normal tissues. TIPE2 suppressed proliferation, growth, migration, and invasion of rectal adenocarcinoma cells via inhibiting Wnt/β-catenin and TGF-β/Smad2/3 signaling pathways, indicating it might be a potential target in rectal adenocarcinoma treatment75 (Table 1). In the study of papillary thyroid carcinoma (PTC), TIPE2 overexpression was observed in tumor samples, and inhibited viability, proliferation, and invasion of PTC cells. Furthermore, TIPE2 attenuated tumor invasiveness via inhibition of Rac1, resulting in reduced MMP-9 and uPA expression, indicating its crucial role in predicting tumor aggressiveness of PTC76 (Table 1). In Non-Hodgkin’s lymphoma (NHL), the increased expression of TIPE2 was identified in both peripheral T cell lymphoma and diffuse large B-cell lymphoma (DLBCL), and TIPE2 expression in DLBCL was stronger than that in T lymphoma. Besides, among DLBCL, TIPE2 expression in germinal center of B-cell (GCB) type was much stronger than that in non-GCB type, indicating that TIPE2 may serve as a prognostic predictor of better survival for DLBCL77 (Table 1). Furthermore, TIPE2 was also observed to modulate the crosstalk between SSCC and tumor-associated macrophages (TAMs). The deficiency of TIPE2 in TAMs was capable of abolishing the phenotypic modification of TAMs exerted by SSCC cells when co-cultured together. Additionally, higher expression of TIPE2 in TAMs was relevant to a worse 5-year overall survival, highlighting TIPE2 as a promising predictor of prognosis and a new therapeutic target for SSCC78 (Table 1).
 |
Table 1 TIPE2 Protein Expression, Function and Signaling Molecules in Cancers |
Contrary to the expression profile of previously mentioned cancers, the expression level of TIPE2 in patients with primary hepatocellular carcinoma (HCC) was significantly weak or undetectable, which showed a negative correlation with tumor migration and invasion. In HCC cell lines, TIPE2 deficiency activated metastasis-associated PI3K/AKT cascade and Rac1 signaling pathways, and then enhanced F-actin polymerization, and increased the secretion of MMP-9 and urokinase-type plasminogen activator (uPA), which ultimately facilitated tumor proliferation and migration.79,80 Correspondingly, TIPE2 overexpression significantly eliminated the effects of lipopolysaccharide on TNF-α expression and abrogated the effects of TNF-α on the upregulation of MMP-3/MMP-13, activation of ERK1/2 and NF-κB, ultimately suppressing TNF-α-induced primary HCC metastasis via inhibition of MMP-3/MMP-13, ERK1/2 and NF-κB signaling cascades81 (Table 1). In addition, the non-structural protein NS5A which is encoded by hepatitis C virus (HCV) was involved in the degradation of TIPE2, resulting in genomic DNA instability and HCV-induced hepatocellular carcinogenesis.82 TIPE2 might be a new diagnostic and therapeutic target for primary hepatocellular carcinoma.12,79–82
Downregulation of TIPE2 was also identified in glioma cells and tissues. Increased TIPE2 expression was found to inhibit proliferation, migration, and EMT of glioma cells through decreasing the levels of β-catenin, c-myc, and cyclin D1 in hypoxia-induced Wnt/β-catenin pathway49 (Table 1). Low expression of TIPE2 was also identified in prostate cancer, and TIPE2 selective overexpression in prostate cancer cell lines was capable of attenuating tumorigenesis, migration, invasion, and EMT via inhibiting PI3K/Akt pathway. TIPE2 might function as a promising therapeutic target for prostate cancer50 (Table 1). The decreased expression of TIPE2 was also identified in NSCLC and selective overexpression of TIPE2 was found to attenuate lymph node metastasis and clinical stage.83 TIPE2 suppressed cell colony formation and tumor angiogenesis through reducing Rac1 activation, downstream factors F-actin polymerization and VEGF expression, which consequently inhibited angiogenesis, invasion, and metastasis of NSCLC cells.84 It was also reported that TIPE2 reduced cisplatin resistance by inducing cell autophagy via mTOR signaling pathway in NSCLC and thus improved the prognosis of NSCLC patients85 (Table 1). Interestingly, a similar result was also identified in osteosarcoma. TIPE2 significantly blocked multidrug resistance1 (MDR1) promoter from transcribing via suppressing Tak1-NF-κB and AP-1 signaling pathways, which greatly reduced MDR1 transcription and improved the sensitivity of osteosarcoma cells to cisplatin86 (Table 1). In accordance with findin50 in NSCLC, TIPE2 was also downregulated in esophageal squamous cell carcinoma (ESCC). It was demonstrated that TIPE2 suppressed tumorigenesis and progression of ESCC through inhibiting Wnt/β-catenin signaling pathway87 (Table 1). The newest clinical result of oral tongue squamous cell carcinoma (OTSCC) showed that TIPE2 also had low expression in OTSCC and enforced overexpression of it affected biological behavior in vitro and suppressed tumor growth in vivo via negatively regulating Foxp3+ Treg cells88 (Table 1).
Compared with normal gastric mucous cells, TIPE2 expression in gastric cancer samples was decreased. Selective expression of TIPE2 in gastric cancer cell lines was found to upregulate activity of N-Ras and p27 via IRF4 signaling pathway, which led to attenuating cell proliferation and growth.55,89 Additionally, due to the upregulation of p27, TIPE2 suppressed the progression of gastritis to gastric cancer.55,89 It was identified that EMT played an indispensable role in tumorigenesis and progression of gastric cancer. In the in-vitro experiments, the expression level of EMT biomarkers like Snail1 and Snail2/Slug was suppressed by TIPE2, and moreover, TIPE2 attenuated invasiveness of gastric cancer cells via downregulating β-catenin signaling through inhibition of AKT and activation of GSK3β, ultimately reversing EMT process and inhibiting the invasion, migration, and metastasis of gastric cancer cells.51,90 Furthermore, attenuation of Akt and ERK1/2 signaling mediated by TIPE2 was also involved in the apoptosis of gastric cancer cells53 (Table 1). The functions of TIPE2 in breast cancer have already been confirmed. TIPE2 expression in breast cancer cells and tissues was much lower when compared with normal ones, and similar to its role in gastric cancer, enforced expression of TIPE2 distinctly impededtumor growth, proliferation, migration, invasion, and EMT in breast cancer cells, like MDA-MB-231 cells.91,92 Mechanistically, limited expression of β-catenin, c-myc, cyclin D1 and decreased phosphorylation of p38 and Akt mediated by TIPE2 were involved in the suppression of occurrence and development of breast cancer cells.91,92 Moreover, it was defined that TIPE2 also induced CD8+ T cells and natural killer (NK) cells to secrete more cytokines, like interferon-γ (IFN-γ) and TNF-α, which in turn enhanced CD8+ T cells and NK cells’ cytotoxicity and antitumor immune responses in spleen and tumor microenvironment, ultimately inhibiting the development and metastasis of breast cancer cells93 (Table 1). In endometrial cancer cells, TIPE2 bound with β-catenin and decreased its nuclear translocation, suppressing EMT and tumorigenesis of endometrial cancer cells94 (Table 1). The expression level of TIPE2 was extremely weak or undetectable in bladder, cervical and ovarian cancers, but the mechanisms remain unknown currently34,35,95 (Table 1).
Conclusions and Perspectives
The expression pattern and biological functions of TIPE2 have been ceaselessly explored in the last decade. In addition to regulating inflammation and maintaining immune homeostasis, TIPE2 also acts as an indispensable suppressor in most cancers. This present review summarized the expression pattern, structure, and regulatory functions of TIPE2 in inflammation, immunity, and cancers based on the latest literature. The differential expression and unique functions of TIPE2 indicate that it is a potential biomarker for diagnosis and prognosis as well as a promising drug target for treatment of TIPE2-associated cancers. However, how to conveniently apply TIPE2 to clinical diagnosis and prognosis remains unknown. We put forward an assumption that the accurate serum expression quantity of TIPE2 in each specific cancer must be detected and the particular diagnostic threshold and reference range of each specific cancer should be reasonably formulated. Although drug therapy targeting TIPE2 seems to be beneficial and is promising to prolong the survival time of cancer patients, the emerging issue, whether targeted therapy is secure and whether it will bring about severe side effects, deserves more consideration and needs more clinical trials to verify it.
Gaining insight into the expression pattern, structure, and functions of TIPE2 is of great significance for the prevention and treatment of various human diseases, especially malignant tumors. As far as what has been clarified about TIPE2, there is still a lack of comprehensive and precise cognition of it, much work is needed to uncover the underlying mysteries of this novel protein.
Abbreviations
Akt, protein kinase B; AP-1, activating protein-1; Bax, Bcl-2 associated X; Bcl-2, B cell lymphoma/leukemia-2; CTLA-4, cytotoxic T-lymphocyte-associated protein-4; DED, death effector domain; DR5, death receptor 5; EMT, epithelial-mesenchymal transition; ERK, extracellular signal-regulated kinase; ESCC, esophageal squamous cell carcinoma; Foxp3, forkhead box protein3; GSK3β, glycogen synthase kinase 3β; HCC, hepatocellular carcinoma; HCV, hepatitis C virus; IFN-γ, interferon-γ; IKBα, NF-κB inhibitor alpha; IL, interleukin; IRF4, interferon regulatory factor 4; JNK, c-Jun N-terminal kinase; KRAS, kirsten rat sarcoma viral oncogene; LATS1, large tumor suppressor gene 1; MAPK, mitogen-activated protein kinases; MALAT1, metastasis-associated lung adenocarcinoma transcript 1; MCP-1, monocyte chemotactic protein-1; MDR1, multidrug resistance1; MMP, matrix metalloproteinase; mTOR, mammalian Target of Rapamycin; mTORC1, mammalian target of rapamycin complex 1; MX1, myxoma resistance protein 1; NFAT5, nuclear factor of activated T-cells 5; NF-κB, nuclear factor-κB; NHL, Non-Hodgkin’s lymphoma; NOD2, oligomerization domain-2; NSCLC, non-small-cell lung cancer; OTSCC, oral tongue squamous cell carcinoma; OX-LDL, oxidized low density lipoprotein; PA, pseudomonas aeruginosa; PARP, poly ADP ribose polymerase; PBMC, peripheral blood mononuclear cells; PI3K, phosphatidylinositol 3-kinase; Poly(I:C), polyinosinic-polycytidylic acid; PTC, papillary thyroid carcinoma; RCC, renal cell carcinoma; Rac1, Ras-related C3 botulinum toxin substrate 1; siRNA, small interfering RNA; SSCC, sksquamous cell carcinoma; STAT3, signal transducer and activator of transcription 3; Tak1, TGF-β-activated kinase 1; TAMs, tumor-associated macrophages; TCR, T cell receptor; TGF-β, transforming growth factor-β; TIPE2, tumor necrosis factor-α-induced protein 8 like 2; TLR, Toll-like receptor; TNF-α, tumor necrosis factor alpha; Tregs, regulatory T cells; TNFAIP8/TIPE, tumor necrosis factor-α-induced protein 8; TNM, tumor node metastasis; uPA, urokinase plasminogen activator; VEGF, vascular endothelial growth factor; YAP, Yes-associated protein.
Data Sharing Statement
Not applicable.
Ethics Approval and Informed Consent
This study was approved by the Academic Committee of The Second Hospital of Jilin University and was conducted in accordance with the principles expressed in the Helsinki Declaration. All datasets were obtained from published literature, so it can be confirmed that written informed consent was obtained.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. The manuscript was approved by all authors for publication.
Disclosure
All authors declare no conflicts of interest for this work.
References
1. Nistico P, Ciliberto G. Biological mechanisms linked to inflammation in cancer: discovery of tumor microenvironment-related biomarkers and their clinical application in solid tumors. Int J Biol Markers. 2020;35(1_suppl):8–11. doi:10.1177/1724600820906155
2. Mantovani A, Garlanda C, Allavena P. Molecular pathways and targets in cancer-related inflammation. Ann Med. 2010;42(3):161–170. doi:10.3109/07853890903405753
3. Zhang M, Wang J, Jia L, et al. Transmembrane TNF-alpha promotes activation-induced cell death by forward and reverse signaling. Oncotarget. 2017;8(38):63799–63812. doi:10.18632/oncotarget.19124
4. Lou Y, Liu S. The TIPE (TNFAIP8) family in inflammation, immunity, and cancer. Mol Immunol. 2011;49(1–2):4–7. doi:10.1016/j.molimm.2011.08.006
5. Kumar D, Whiteside TL, Kasid U. Identification of a novel tumor necrosis factor-alpha-inducible gene, SCC-S2, containing the consensus sequence of a death effector domain of fas-associated death domain-like interleukin- 1beta-converting enzyme-inhibitory protein. J Biol Chem. 2000;275(4):2973–2978. doi:10.1074/jbc.275.4.2973
6. Fayngerts SA, Wu J, Oxley CL, et al. TIPE3 is the transfer protein of lipid second messengers that promote cancer. Cancer Cell. 2014;26(4):465–478. doi:10.1016/j.ccr.2014.07.025
7. Sun HH, Gong S, Carmody RJ, et al. TIPE2, a negative regulator of innate and adaptive immunity that maintains immune homeostasis. Cell. 2008;133(3):415–426. doi:10.1016/j.cell.2008.03.026
8. Niture S, Dong XL, Arthur E, et al. Oncogenic role of tumor necrosis factor alpha-induced protein 8 (TNFAIP8). Cells. 2019;8(1):16.
9. Day TF, Mewani RR, Starr J, et al. Transcriptome and proteome analyses of TNFAIP8 knockdown cancer cells reveal new insights into molecular determinants of cell survival and tumor progression. Methods Mol Biol. 2017;1513(2):83–100.
10. Ahn S-H, Deshmukh H, Johnson N, et al. Two genes on A/J chromosome 18 are associated with susceptibility to staphylococcus aureus infection by combined microarray and QTL analyses. PLoS Pathog. 2010;6(9):e1001088. doi:10.1371/journal.ppat.1001088
11. Zhang Z, Liang X, Gao L, et al. TIPE1 induces apoptosis by negatively regulating Rac1 activation in hepatocellular carcinoma cells. Oncogene. 2015;34(20):2566–2574. doi:10.1038/onc.2014.208
12. Gus-Brautbar Y, Johnson D, Zhang L, et al. The anti-inflammatory TIPE2 is an inhibitor of the oncogenic Ras. Molecular Cell. 2012;45(5):610–618. doi:10.1016/j.molcel.2012.01.006
13. Bradley JR. TNF-mediated inflammatory disease. J Pathol. 2008;214(2):149–160. doi:10.1002/path.2287
14. Patel S, Wang FH, Whiteside TL, Kasid U. Identification of seven differentially displayed transcripts in human primary and matched metastatic head and neck squamous cell carcinoma cell lines: implications in metastasis and/or radiation response. Oral Oncol. 1997;33(4):197–203.
15. Uhlen M, Fagerberg L, Hallstrom BM, et al. Tissue-based map of the human proteome. Science. 2015;347(6220):1260419.
16. Zhang C, Chakravarty D, Sakabe I, et al. Role of SCC-S2 in experimental metastasis and modulation of VEGFR-2, MMP-1, and MMP-9 expression. Mol Ther. 2006;13(5):947–955. doi:10.1016/j.ymthe.2005.11.020
17. Xing Y, Liu Y, Liu T, et al. TNFAIP8 promotes the proliferation and cisplatin chemoresistance of non-small cell lung cancer through MDM2/p53 pathway. Cell Commun Signal. 2018;16(1):43. doi:10.1186/s12964-018-0254-x
18. Han Y, Tang Z, Zhao Y, Li Q, Wang E. TNFAIP8 regulates Hippo pathway through interacting with LATS1 to promote cell proliferation and invasion in lung cancer. Mol Carcinogenesis. 2018;57(2):159–166. doi:10.1002/mc.22740
19. Dong Q, Fu L, Zhao Y, Xie C, Li Q, Wang E. TNFAIP8 interacts with LATS1 and promotes aggressiveness through regulation of Hippo pathway in hepatocellular carcinoma. Oncotarget. 2017;8(9):15689–15703. doi:10.18632/oncotarget.14938
20. Porturas TP, Sun H, Buchlis G, et al. Crucial roles of TNFAIP8 protein in regulating apoptosis and listeria infection. J Immunol. 2015;194(12):5743–5750. doi:10.4049/jimmunol.1401987
21. Cui J, Zhang G, Hao C, et al. The expression of TIPE1 in murine tissues and human cell lines. Mol Immunol. 2011;48(12–13):1548–1555. doi:10.1016/j.molimm.2011.04.023
22. Wu X, Ma Y, Cheng J, et al. TIPE1 function as a prognosis predictor and negative regulator of lung cancer. Oncotarget. 2017;8(45):78496–78506. doi:10.18632/oncotarget.19655
23. Liu WW, Chen Y, Xie H, et al. TIPE1 suppresses invasion and migration through down-regulating Wnt/beta-catenin pathway in gastric cancer. J Cell Mol Med. 2018;22(2):1103–1117.
24. Qiu S, Hu W, Ma Q, Zhao Y, Li L, Ding Y. TIPE1 suppresses the invasion and migration of breast cancer cells and inhibits epithelial-to-mesenchymal transition primarily via the ERK signaling pathway. Acta Biochimica Et Biophysica Sinica. 2019;51(10):1008–1015. doi:10.1093/abbs/gmz099
25. Chen P, Zhou J, Li J, Zhang Q, Zuo Q. TIPE1 suppresses osteosarcoma tumor growth by regulating macrophage infiltration. Clin Transl Oncol. 2019;21(3):334–341. doi:10.1007/s12094-018-1927-z
26. Wang YN, Liu Y, Hu CF, Ni XY, Huang XB. Tumor necrosis factor -induced protein 8-like 1 promotes apoptosis by regulating B-cell leukemia/lymphoma-2 family proteins in RAW264.7 cells. Oncol Letters. 2016;12(5):3506–3512. doi:10.3892/ol.2016.5090
27. Zhao PQ, Pang XM, Jiang J, et al. TIPE1 promotes cervical cancer progression by repression of p53 acetylation and is associated with poor cervical cancer outcome. Carcinogenesis. 2019;40(4):592–599. doi:10.1093/carcin/bgy163
28. Cui J, Hao C, Zhang W, et al. Identical expression profiling of human and murine TIPE3 protein reveals links to its functions. J Histochem Cytochem. 2015;63(3):206–216. doi:10.1369/0022155414564871
29. Gao JF, Zhang H, Lv J, Fan YY, Feng D, Song L. Effects of the long and short isoforms of TIPE3 on the growth and metastasis of gastric cancer. Biomed Pharmacother. 2020;124(4):109853. doi:10.1016/j.biopha.2020.109853
30. Lian K, Ma C, Hao C, et al. TIPE3 protein promotes breast cancer metastasis through activating AKT and NF-κB signaling pathways. Oncotarget. 2017;8(30):48889–48904. doi:10.18632/oncotarget.16522
31. Yuan F, Liu B, Xu Y, et al. TIPE3 is a regulator of cell apoptosis in glioblastoma. Cancer Lett. 2019;45(446):1–14. doi:10.1016/j.canlet.2018.12.019
32. Ren XY, Wen X, Li YQ, et al. TIPE3 hypermethylation correlates with worse prognosis and promotes tumor progression in nasopharyngeal carcinoma. J Exp Clin Cancer Res. 2018;37(1):227–238.
33. Li Z, Jia W, Niu J, Zhang L. Understanding the roles of negative immune regulator TIPE2 in different diseases and tumourigenesis. Histol Histopathol. 2018;33(9):919–928.
34. Zhang G, Hao C, Lou Y, et al. Tissue-specific expression of TIPE2 provides insights into its function. Mol Immunol. 2010;47(15):2435–2442. doi:10.1016/j.molimm.2010.06.016
35. Zhang L, Shi Y, Wang Y, et al. The unique expression profile of human TIPE2 suggests new functions beyond its role in immune regulation. Mol Immunol. 2011;48(9–10):1209–1215. doi:10.1016/j.molimm.2011.03.001
36. Goldsmith JR, Chen YH. Regulation of inflammation and tumorigenesis by the TIPE family of phospholipid transfer proteins. Cell Mol Immunol. 2017;14(6):482–487. doi:10.1038/cmi.2017.4
37. Zhang X, Wang J, Fan C, et al. Crystal structure of TIPE2 provides insights into immune homeostasis. Nat Struct Mol Biol. 2009;16(1):89–90. doi:10.1038/nsmb.1522
38. Antony P, Baby B, Vijayan R. Molecular insights into the binding of phosphoinositides to the TH domain region of TIPE proteins. J Mol Model. 2016;22(11):272–280. doi:10.1007/s00894-016-3141-7
39. Fei X, Sheng Z, Yao Y. Update in immune regulatory pathway of tumor necrosis factor-α-induced protein 8-like 2. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue. 2019;31(6):772–776.
40. Ruan Q, Wang P, Wang T, et al. MicroRNA-21 regulates T-cell apoptosis by directly targeting the tumor suppressor gene Tipe2. Cell Death Dis. 2014;5(2):8.
41. Domingo-Gil E, González JM, Esteban M. Identification of cellular genes induced in human cells after activation of the OAS/RNaseL pathway by vaccinia virus recombinants expressing these antiviral enzymes. J Interferon Cytokine Res. 2010;30(3):171–188. doi:10.1089/jir.2009.0037
42. Oho M, Nakano R, Nakayama R, et al. TIPE2 (Tumor Necrosis Factor α-induced Protein 8-like 2) is a novel negative regulator of TAK1 signal. J Biol Chem. 2016;291(43):22650–22660. doi:10.1074/jbc.M116.733451
43. Li X-M, Su J-R, Yan S-P, Cheng Z-L, Yang -T-T, Zhu Q. A novel inflammatory regulator TIPE2 inhibits TLR4-mediated development of colon cancer via caspase-8. Cancer Biomarkers. 2014;14(4):233–240.
44. Zhang H, Zhu T, Liu W, et al. TIPE2 acts as a negative regulator linking NOD2 and inflammatory responses in myocardial ischemia/reperfusion injury. J Mol Med. 2015;93(9):1033–1043. doi:10.1007/s00109-015-1288-9
45. Liu R, Liu C, Liu C, Fan T, Geng W, Ruan Q. TIPE2 in dendritic cells inhibits the induction of pTregs in the gut mucosa. Biochem Biophys Res Commun. 2019;509(4):911–917. doi:10.1016/j.bbrc.2019.01.008
46. Luan -Y-Y, Yao Y-M, Zhang L, et al. Expression of tumor necrosis factor-alpha induced protein 8 like-2 contributes to the immunosuppressive property of CD4(+)CD25(+) regulatory T cells in mice. Mol Immunol. 2011;49(1–2):219–226.
47. Jiang J, Wang S, Fang J, et al. Stable silencing of TIPE2 reduced the Poly I:C‑induced apoptosis in THP‑1 cells. Mol Med Rep. 2017;16(5):6313–6319. doi:10.3892/mmr.2017.7364
48. Zhang G, Zhao L, Wang Y, et al. TIPE2 protein prevents injury-induced restenosis in mice. Biochim Biophys Acta. 2015;1852(8):1574–1584. doi:10.1016/j.bbadis.2015.04.018
49. Liu ZJ, Liu HL, Zhou HC, Wang GC. TIPE2 inhibits hypoxia-induced Wnt/β-catenin pathway activation and EMT in glioma cells. Oncol Res. 2016;24(4):255–261.
50. Lu Q, Liu Z, Li Z, et al. TIPE2 overexpression suppresses the proliferation, migration, and invasion in prostate cancer cells by inhibiting PI3K/Akt signaling pathway. Oncol Res. 2016;24(5):305–313. doi:10.3727/096504016X14666990347437
51. Wu J, Zhang H, Xu C, et al. TIPE2 functions as a metastasis suppressor via negatively regulating β-catenin through activating GSK3β in gastric cancer. Int J Oncol. 2016;48(1):199–206. doi:10.3892/ijo.2015.3224
52. Shi C, Zhang S, Hong S, et al. The pro-apoptotic effects of TIPE2 on AA rat fibroblast-like synoviocytes via regulation of the DR5-caspase-NF-κB pathway in vitro. Onco Targets Ther. 2016;10(9):993–1000.
53. Zhu Y, Tao M, Wu J, et al. Adenovirus-directed expression of TIPE2 suppresses gastric cancer growth via induction of apoptosis and inhibition of AKT and ERK1/2 signaling. Cancer Gene Ther. 2016;23(4):98–106. doi:10.1038/cgt.2016.6
54. Suo LG, Cui YY, Bai Y, Qin XJ. Anti-inflammatory TIPE2 inhibits angiogenic VEGF in retinal pigment epithelium. Mol Immunol. 2016;73(9):46–52. doi:10.1016/j.molimm.2016.03.013
55. Peng Y, Zhao Q, Zhang H, et al. TIPE2, a negative regulator of TLR signaling, regulates p27 through IRF4-induced signaling. Oncol Rep. 2016;35(4):2480–2486. doi:10.3892/or.2016.4562
56. Luan YY, Yao YM, Sheng ZY. The tumor necrosis factor-alpha-induced protein 8 family in immune homeostasis and inflammatory cancer diseases. J Biol Regul Homeost Agents. 2013;27(3):611–619.
57. Jia L, Gui B, Tian P, et al. TIPE2, a novel biomarker for clinical chronic kidney allograft rejection. Artif Organs. 2013;37(2):221–225. doi:10.1111/j.1525-1594.2012.01527.x
58. Wang Q, Wei C, Ma L, et al. Inflammatory cytokine TNF-α promotes corneal endothelium apoptosis via upregulating TIPE2 transcription during corneal graft rejection. Graefes Arch Clin Exp Ophthalmol. 2018;256(4):709–715. doi:10.1007/s00417-018-3913-0
59. Zhao Y, Wang Y, Zhu MS, et al. Expression pattern of tumor necrosis factor-α-induced Protein 8-Like 2 in acute rejection of cardiac allograft. Transplant Proc. 2018;50(1):293–298. doi:10.1016/j.transproceed.2017.11.020
60. Shi C, Wang Y, Zhuang G, Qi Z, Li Y, Yin P. Tumor necrosis factor‑α‑induced protein‑8 like 2 regulates lipopolysaccharide‑induced rat rheumatoid arthritis immune responses and is associated with Rac activation and interferon regulatory factor 3 phosphorylation. Mol Med Rep. 2017;16(5):6346–6353. doi:10.3892/mmr.2017.7311
61. Lou Y, Zhang G, Geng M, Zhang W, Cui J, Liu S. TIPE2 negatively regulates inflammation by switching arginine metabolism from nitric oxide synthase to arginase. PLoS One. 2014;9(5):e96508. doi:10.1371/journal.pone.0096508
62. Li F, Zhu X, Yang Y, Huang L, Xu J. TIPE2 alleviates systemic lupus erythematosus through regulating macrophage polarization. Cell Physiol Biochem. 2016;38(1):330–339. doi:10.1159/000438633
63. Zhao JW, Shi G, Ping JD, Ming L. Study on the negative regulation of TIPE2 from peripheral blood mononuclear cells on tissue factor in patients with bronchial asthma. Zhonghua Yi Xue Za Zhi. 2018;98(36):2889–2893.
64. Liu Y, Jin J, Ji J, Gao XM, Fan YC. Tumor necrosis factor-α-induced protein 8-like 2 mRNA in peripheral blood mononuclear cells is associated with the disease progression of chronic hepatitis B virus infection. Virol J. 2019;16(1):120. doi:10.1186/s12985-019-1224-7
65. Qin B, Wei T, Wang L, et al. Decreased expression of TIPE2 contributes to the hyperreactivity of monocyte to Toll-like receptor ligands in primary biliary cirrhosis. Journal of Gastroenterology and Hepatology. 2016;31(6):1177–1183. doi:10.1111/jgh.13251
66. Luan YY, Yao YM, Zhang L, et al. Expression of tumor necrosis factor-α induced protein 8 like-2 contributes to the immunosuppressive property of CD4(+)CD25(+) regulatory T cells in mice. Mol Immunol. 2011;49(1–2):219–226.
67. Lou Y, Liu S, Zhang C, et al. Enhanced atherosclerosis in TIPE2-deficient mice is associated with increased macrophage responses to oxidized low-density lipoprotein. J Immunol. 2013;191(9):4849–4857. doi:10.4049/jimmunol.1300053
68. Zhang G, Zhang W, Lou Y, et al. TIPE2 deficiency accelerates neointima formation by downregulating smooth muscle cell differentiation. Cell Cycle. 2013;12(3):501–510. doi:10.4161/cc.23325
69. Liu MW, Su MX, Zhang W, Wang L, CY Q. Atorvastatin increases lipopolysaccharide-induced expression of tumour necrosis factor-alpha-induced protein 8-like 2 in RAW264.7 cells. Exp Ther Med. 2014;8(8):219–228. doi:10.3892/etm.2014.1722
70. Zhang Y, Wei X, Liu L, et al. TIPE2, a novel regulator of immunity, protects against experimental stroke. J Biol Chem. 2012;287(39):32546–32555. doi:10.1074/jbc.M112.348755
71. Zhang YY, Huang NN, Zhao YX, et al. Elevated tumor necrosis factor-a-induced Protein 8-like 2 mRNA from peripheral blood mononuclear cells in patients with acute ischemic stroke. Int J Med Sci. 2018;15(14):1713–1722. doi:10.7150/ijms.27817
72. Jiang Y, Li Q, Zhang Y, Gao Y, Jiang L, Chen Z. TIPE2 governs macrophage polarization via negative regulation of mTORC1. Mol Med Rep. 2018;17(1):952–960.
73. Wang Q, Ma L, Liu T, et al. TIPE2 suppresses pseudomonas aeruginosa keratitis by inhibiting NF-κB signaling and the infiltration of inflammatory cells. J Infect Dis. 2019;220(6):1008–1018. doi:10.1093/infdis/jiz246
74. Zhang Z, Qi H, Hou S, Jin X. TIPE2 mRNA overexpression correlates with TNM staging in renal cell carcinoma tissues. Oncol Letters. 2013;6(2):571–575. doi:10.3892/ol.2013.1388
75. Wu DD, Liu SY, Gao YR, et al. Tumour necrosis factor-α-induced protein 8-like 2 is a novel regulator of proliferation, migration, and invasion in human rectal adenocarcinoma cells. J Cell Mol Med. 2019;23(3):1698–1713. doi:10.1111/jcmm.14065
76. Jia W, Li Z, Chen J, et al. TIPE2 acts as a biomarker for tumor aggressiveness and suppresses cell invasiveness in papillary thyroid cancer (PTC). Cell Biosci. 2018;8(8):49. doi:10.1186/s13578-018-0247-x
77. Hao C, Zhang N, Geng M, et al. Clinical significance of TIPE2 protein upregulation in non-hodgkin’s lymphoma. J Histochem Cytochem. 2016;64(9):556–564. doi:10.1369/0022155416662262
78. Li X. TIPE2 regulates tumor-associated macrophages in skin squamous cell carcinoma. Tumour Biol. 2016;37(4):5585–5590. doi:10.1007/s13277-015-4388-9
79. Cao X, Zhang L, Shi Y, et al. Human tumor necrosis factor (TNF)-alpha-induced protein 8-like 2 suppresses hepatocellular carcinoma metastasis through inhibiting Rac1. Mol Cancer. 2013;12(1):149. doi:10.1186/1476-4598-12-149
80. Wang L, Chen C, Feng S, Tian J. TIPE‑2 suppresses growth and aggressiveness of hepatocellular carcinoma cells through downregulation of the phosphoinositide 3‑kinase/AKT signaling pathway. Mol Med Rep. 2018;17(5):7017–7026.
81. Zhang YH, Yan HQ, Wang F, et al. TIPE2 inhibits TNF-α-induced hepatocellular carcinoma cell metastasis via Erk1/2 downregulation and NF-κB activation. Int J Oncol. 2015;46(1):254–264. doi:10.3892/ijo.2014.2725
82. Wang Y, Jiang Y, Zhou J, et al. Hepatitis C virus promotes hepatocellular carcinogenesis by targeting TIPE2, a new regulator of DNA damage response. Tumour Biol. 2016;37(11):15265–15274. doi:10.1007/s13277-016-5409-z
83. Li Y, Li X, Liu G, et al. Downregulated TIPE2 is associated with poor prognosis and promotes cell proliferation in non-small cell lung cancer. Biochem Biophys Res Commun. 2015;457(1):43–49. doi:10.1016/j.bbrc.2014.12.080
84. Li Z, Guo C, Liu X, et al. TIPE2 suppresses angiogenesis and non-small cell lung cancer (NSCLC) invasiveness via inhibiting Rac1 activation and VEGF expression. Oncotarget. 2016;7(38):62224–62239. doi:10.18632/oncotarget.11406
85. Guo H, Ren H, Li J, et al. TIPE2 suppressed cisplatin resistance by inducing autophagy via mTOR signalling pathway. Exp Mol Pathol. 2020;113(9):104367. doi:10.1016/j.yexmp.2020.104367
86. Zhao P, Wang S, Jiang J, et al. TIPE2 sensitizes osteosarcoma cells to cis-platin by down-regulating MDR1 via the TAK1- NF-κB and - AP-1 pathways. Mol Immunol. 2018;101:471–478. doi:10.1016/j.molimm.2018.08.010
87. Zhu L, Zhang X, Fu X, et al. TIPE2 suppresses progression and tumorigenesis of esophageal carcinoma via inhibition of the Wnt/β-catenin pathway. J Transl Med. 2018;16(1):7. doi:10.1186/s12967-018-1383-0
88. Zhao LL. TIPE2 suppresses progression and tumorigenesis of the oral tongue squamous cell carcinoma by regulating FoxP3(+) regulatory T cells. J Bioenerg Biomembr. 2020;52(4):279–289. doi:10.1007/s10863-020-09840-w
89. Zhao Q, Zhao M, Dong T, et al. Tumor necrosis factor-alpha-induced Protein-8 Like-2 (TIPE2) upregulates p27 to decrease gastic cancer cell proliferation. J Cell Biochem. 2015;116(6):1121–1129. doi:10.1002/jcb.25068
90. Yin H, Huang X, Tao M, et al. Adenovirus-mediated TIPE2 overexpression inhibits gastric cancer metastasis via reversal of epithelial-mesenchymal transition. Cancer Gene Ther. 2017;24(4):180–188. doi:10.1038/cgt.2017.3
91. Wang K, Ren Y, Liu Y, Zhang J, He JJ. Tumor Necrosis Factor (TNF)-α-Induced Protein 8-like-2 (TIPE2) inhibits proliferation and tumorigenesis in breast cancer cells. Oncol Res. 2017;25(1):55–63. doi:10.3727/096504016X14719078133320
92. Zhang Z, Liu L, Liu C, Cao S, Zhu Y, Mei Q. TIPE2 suppresses the tumorigenesis, growth and metastasis of breast cancer via inhibition of the AKT and p38 signaling pathways. Oncol Rep. 2016;36(6):3311–3316. doi:10.3892/or.2016.5192
93. Zhang Z, Liu L, Cao S, Zhu Y, Mei Q. Gene delivery of TIPE2 inhibits breast cancer development and metastasis via CD8(+) T and NK cell-mediated antitumor responses. Mol Immunol. 2017;85(9):230–237. doi:10.1016/j.molimm.2017.03.007
94. Liu Y, Wang X, Wan L, et al. TIPE2 inhibits the migration and invasion of endometrial cells by targeting β-catenin to reverse epithelial-mesenchymal transition. Hum Reprod. 2020;35(6):1377–1390. doi:10.1093/humrep/deaa062
95. Liang D, Xu W, Zhang Q, Tao BB. Study on the effect of Integrin αVβ6 on proliferation and apoptosis of cervical cancer cells. Eur Rev Med Pharmacol Sci. 2017;21(12):2811–2815.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.