")
Back to Journals » Infection and Drug Resistance » Volume 15
Pyrazinamide Resistance and pncA Mutation Profiles in Multidrug Resistant Mycobacterium Tuberculosis
Authors Shi D, Zhou Q , Xu S, Zhu Y, Li H , Xu Y
Received 6 April 2022
Accepted for publication 13 July 2022
Published 30 August 2022 Volume 2022:15 Pages 4985—4994
DOI https://doi.org/10.2147/IDR.S368444
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Dawei Shi,1,* Qiulong Zhou,2,* Sihong Xu,1 Yumei Zhu,3 Hui Li,4 Ye Xu2
1National Institutes for Food and Drug Control, Institute of Pathogen Biology at the Chinese Academy of Medical Sciences & Peking Union Medical College, Beijing, People’s Republic of China; 2Engineering Research Centre of Molecular Diagnostics of the Ministry of Education, State Key Laboratory of Cellular Stress Biology, State Key Laboratory of Molecular Vaccinology and Molecular Diagnostics, School of Life Sciences, Xiamen University, Xiamen, People’s Republic of China; 3Shenzhen Center for Chronic Disease Control, Shenzhen, People’s Republic of China; 4Tuberculosis Reference Laboratory, Henan Provincial Centers for Disease Control and Prevention, Zhengzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hui Li, Tuberculosis Reference Laboratory, Henan Provincial Centers for Disease Control and Prevention, Zhengzhou, People’s Republic of China, Tel/Fax +86 371-68089049, Email [email protected] Ye Xu, Engineering Research Centre of Molecular Diagnostics of the Ministry of Education, State Key Laboratory of Cellular Stress Biology, State Key Laboratory of Molecular Vaccinology and Molecular Diagnostics, School of Life Sciences, Xiamen University, Xiamen, People’s Republic of China, Tel +86 592 2187992, Fax +86 592 2187363, Email [email protected]
Purpose: Pyrazinamide (PZA) is a critical component of standardized chemotherapy for tuberculosis (TB) and is recommended for the treatment of multidrug-resistant (MDR) TB. We aimed to characterize mutations in pncA of M. tuberculosis and evaluate their diagnostic accuracy for PZA susceptibility in China. We also combined genotypic methods with phenotypic susceptibility testing and pyrazinamidase (PZAse) activity to confirm PZA-resistant M. tuberculosis isolates.
Results: An evaluation of 82 MDR M. tuberculosis strains revealed that 28.0% (23/82) were phenotypically resistant to 100 mg/L PZA and 15.9% (13/82) showed resistance to 300 mg/L PZA. Mutations in pncA were detected at 33 unique sites, and the majority were point mutations. No evident mutation hotspots or mutations affecting multiple amino acids were found, but the association between pncA mutations and PZA resistance was significant under 100 and 300 mg/L. The sensitivity of pncA mutation detection for predicting PZA susceptibility was 82.6% (19/23), and the specificity was 61.0% (36/59), based on 100 mg/L PZA, whereas the sensitivity was 84.6% (11/13) and the specificity was 55.1% (38/69), based on 300 mg/L PZA. All mutations identified in the highly PZA-resistant (300 mg/L) strains had an 80% loss relative to PZAse activity. No evident PZAse activity loss was observed in one synonymous mutation strain and the loss exceed 60% in all other strains.
Conclusion: The association between pncA mutation and PZA resistance was significant. Relatively, the molecular method have shown better reliability than the phenotypic method for the detection of PZA resistance. This provides a theoretical basis for the clinical diagnosis of drug-resistant TB.
Keywords: MDR, PZA, pncA, DST, Beijing genotype, enzymatic activity
Introduction
Tuberculosis (TB), a respiratory disease caused by Mycobacterium tuberculosis infection, is one of the top 10 health problems worldwide. The World Health Organization (WHO) reported about 9.9 million new cases of TB worldwide in 2020, of which 2.59% exhibited a multidrug-resistant phenotype (MDR-TB). MDR-TB continues to be a significant threat to public health.1,2
Pyrazinamide (PZA) is an important first-line anti-TB drug known for its unique sterilizing activity and reduction of TB therapy time.3–5 Good penetration of PZA into the lung tissue of patients with MDR-TB has been shown to produce a significant correlation between tissue PZA concentrations and necrosis. Tissue penetration of pyrazinamide into the diseased lung and the favorable acidic environment of most chronic tuberculous lesions, which promotes the bactericidal and sterilizing activity of pyrazinamide.6 In drug-susceptible TB, the addition of PZA to the rifampin/isoniazid regimen has facilitated the shortening of the treatment period from 9 to 6 months.7,8 A recent study showed that introducing a molecular drug susceptibility test (DST) to PZA successfully improved treatment outcomes, increasing the relapse-free success rate to 82.4% in PZA-susceptible cases and shortening the regimen period to 12 months without any additional agents. In addition, clinical trials to evaluate short-course chemotherapy regimens in MDR-TB are ongoing, including STREAM, Simplici TB, MDR-END, and TB-TRUST trials. All these regimens include PZA.10–16 The potential synergistic activities between PZA and important second-line as well as novel drugs (such as bedaquiline, delamanid, and pretomanid) highlight its importance in both current and future MDR-TB treatment strategies.
The sterilizing activity of PZA is activated following its catalysis by pyrazinamidase (PZAse), and PZA susceptibility evaluations depend on the acidity of the culture medium. Currently, growth-based DSTs using the MGIT 960 PZA kit are widely regarded as the gold standard for evaluating M. tuberculosis drug susceptibility to PZA. However, there are limitations to the current PZA resistance tests. The long incubation period for M. tuberculosis strains means phenotypic DSTs may take months to complete, resulting in significant diagnostic delays. Moreover, PZA phenotypic DSTs are rarely performed in routine clinical settings as the test is time-consuming and technically demanding. The low-pH medium and stringent inoculum size of these assays often lead to inconsistent and non-reproducible results that deter clinicians from relying on these tests for guidance.
Molecular testing for PZA susceptibility is based on the detection of mutations in the pncA gene. The M. tuberculosis pncA gene encodes a functional PZAse, the key enzyme in PZA activation, and mutations in this gene may reduce the activity of PZAse and thus increase M. tuberculosis resistance to PZA.17 Several studies have shown that 72–98% of PZA resistance is due to pncA mutations, which are highly diverse and scattered across both the open reading frames and upstream regulatory regions of this gene.18,19 Similarly, some studies have found a correlation between mutations in the pncA gene and phenotypic PZA susceptibility, but the performance of these molecular diagnostic assays is not as good as that for isoniazid (INH) and rifampicin (RIF).19,20
This study aimed to characterize the mutation pattern and mutant types in the pncA gene of MDR-TB strains from Henan, China using direct sequencing and subsequently comparing these results to those of the MGIT 960 PZA susceptibility assay and confirming these results with a PZAse activity test. We also evaluated the agreement between the genotypic and phenotypic evaluations of PZA susceptibility in this MDR M. tuberculosis population dominated by Beijing-genotype strains. Our findings will help to further develop molecular PZA detection methods.
Materials and Methods
M. tuberculosis Clinical Strains
From January 2017 to March 2018, a population of 95 MDR M. tuberculosis clinical strains were collected via active screening of clinical M. tuberculosis isolates from patients in Henan Province. The isolation, culture, and species identification of these clinical strains were conducted in the Tuberculosis Reference Laboratory (TRL) at Henan Provincial Centers for Disease Control and Prevention (Henan CDC), China. This study was approved by the Institutional Review Board of the Institute of Pathogen Biology at the Chinese Academy of Medical Sciences & Peking Union Medical College. Written informed consent was obtained from all patients before bacterial isolation.
Drug Susceptibility Testing (DST)
Drug susceptibility testing was performed with WHO-recommended proportion method. DSTs for INH and RIF were conducted by the TRL at the Henan CDC. They used the Löwenstein–Jensen proportion method recommended by the WHO/IUATLD at the following critical drug concentrations: INH, 0.2 mg/L; RIF, 40.0 mg/L.21 PZA resistance of these bacilli was evaluated using the BACTEC MGIT 960 automatic method, according to the manufacturer’s instructions (Becton Dickinson, Sparks, MD, USA). The MGIT 960 medium is a modified Middlebrook 7H9 broth with a reduced pH of 5.9. According to the BACTEC MGIT 960 PZA instructions, the McFerland turbidity of the strain was adjusted to 1, then a total of 500 µL of each inoculum was added to the PZA-containing tube, and a 1:10 dilution of the inoculum was added to another tube without PZA as the growth control. We evaluated two concentrations of PZA (100 mg/L and 300 mg/L) to define the phenotypic drug resistance of each strain; 100 mg/L is the concentration recommended by the manufacturer and is the common concentration used in routine clinical practice.
Genomic DNA Extraction and Detection of pncA Mutations
The crude genomic DNA of each strain was extracted using the boiling method,22 and the entire pncA coding region was amplified by PCR. PCR was conducted according to the protocols for Phusion® Hot Start High-Fidelity DNA Polymerase (Finnzymes, Espoo, Finland) with some modifications. The reaction mixture contained 10 μL HF buffer, 1.5 μL DMSO, 2 μL dNTP mix (10 nM), 0.5 μL of each primer (final concentration: 50 pmol/L), 0.5 μL Phusion® High-Fidelity DNA Polymerase, and 3 μL crude genomic DNA. The PCR cycling conditions were set up as follows: initial denaturation at 98 °C for 30s; 35 cycles of 98 °C for 10s, 63 °C for 30s, 72 °C for 1 min; and then final extension at 72 °C for 7 min. PCR products (763 bp) were then submitted to Sinogenomax Co. Ltd (Beijing, China) for sequencing. The primers used for amplification were pncA-1: 5′-CGAACGTATGGTGGACGTAT-3′ (position −18 to position +2 of pncA ORF) and pncA-2: 5′-CCGATGAAGGTGTCGTAGAA-3′ (downstream 165 bp to 184 bp of the stop codon of pncA ORF); the same primers were also used in the subsequent sequencing of the PCR products.
Sequence Alignment and Statistical Analysis
Sequence data were assembled using by Seqman pro (version11.2, DNAstar Lasergene, Madison, WI, USA), and mutations were then evaluated by comparing these sequences to the pncA gene (NC_000962.2) from M. tuberculosis H37Rv, the M. tuberculosis reference strain. The frequency calculations and association analyses were performed using SPSS for windows (version 26.0; SPSS Inc. Armonk, NY, USA) and a P-value < 0.05 was considered significant.
MIRU-VNTR Genotyping and Strain Identification
MIRU-VNTR genotyping was performed as previously described.23 Briefly, this method relies on the evaluation of seven VNTR loci (VNTR-7) known for their high discriminatory power in Chinese M. tuberculosis strains.24 Samples with more than one band in the polymerase chain reaction (PCR) products on any VNTR locus were treated as mixed strains and excluded from the evaluation. Most of these strains were identified as the Beijing genotype, and this classification was confirmed using Deletion-targeted Multiplex PCR (DTM-PCR) as previously reported.25
Expression and Purification
PZAse for each mutation type containing a carboxy-terminal His-tag was expressed in Escherichia coli and purified by affinity chromatography on a Ni-nitrilotriacetic acid column (GE Healthcare, Uppsala, Sweden) as described previously.26,27 Briefly, the pncA gene was amplified, using PCR with specific primers. DNA of an M. tuberculosis isolate cloned into a pET28a expression vector to produce a His-tag prokaryotic expression plasmid. After verified by sequencing, this fusion protein was expressed in E.coli BL21 cells. After that, sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was used to analysis the purity and molecular weight of the proteins, and we have measured protein concentration according to the BCA method.28
PZAse Enzymatic Activity
PZAse activity was then evaluated using the standard Wayne test.29,30 Briefly, PZA (100 µg/mL) was hydrolyzed using 100 ng/µL recombinant PZAse, and pyrazinoic acid (POA) production was quantified following the addition of 20% FeNH4(SO4)2, and the reaction was stopped with 0.1 M glycine–HCl (pH 3.4) after 1 min. The absorbance was then measured at 450 nm and the OD was adjusted to produce a standard curve of known concentrations. Each recombinant PZAse was then evaluated at least three times and their enzymatic activity was estimated using the amount of POA produced in the 1-min reaction divided by the total amount of enzyme in each reaction. This method effectively assessed PZAse activity.
Results
Patient Demographics and MDR Strain Genotyping
A total of 13 of the original 95 clinical isolates (13.7%) were excluded from this study as they were found to be mixed samples, as demonstrated by the presence of more than one band in any of the seven VNTR loci used for MIRU-VNTR genotyping, or they produced an unreadable nesting sequence signal for pncA. This left 82 MDR isolates which were then subjected to PZA susceptibility testing and DTM-PCR. There were no significant differences in the age, gender, or treatment history of the patients in the present study (Table 1) who were treated with different doses of PZA. The DTM-PCR indicated that 96% (79/82) of these MDR isolates belonged to the Beijing family of M. tuberculosis strains, and there was no significance between the PZA susceptibility profiles of our strains and those representing the Beijing family of MDR isolates (Table 1).
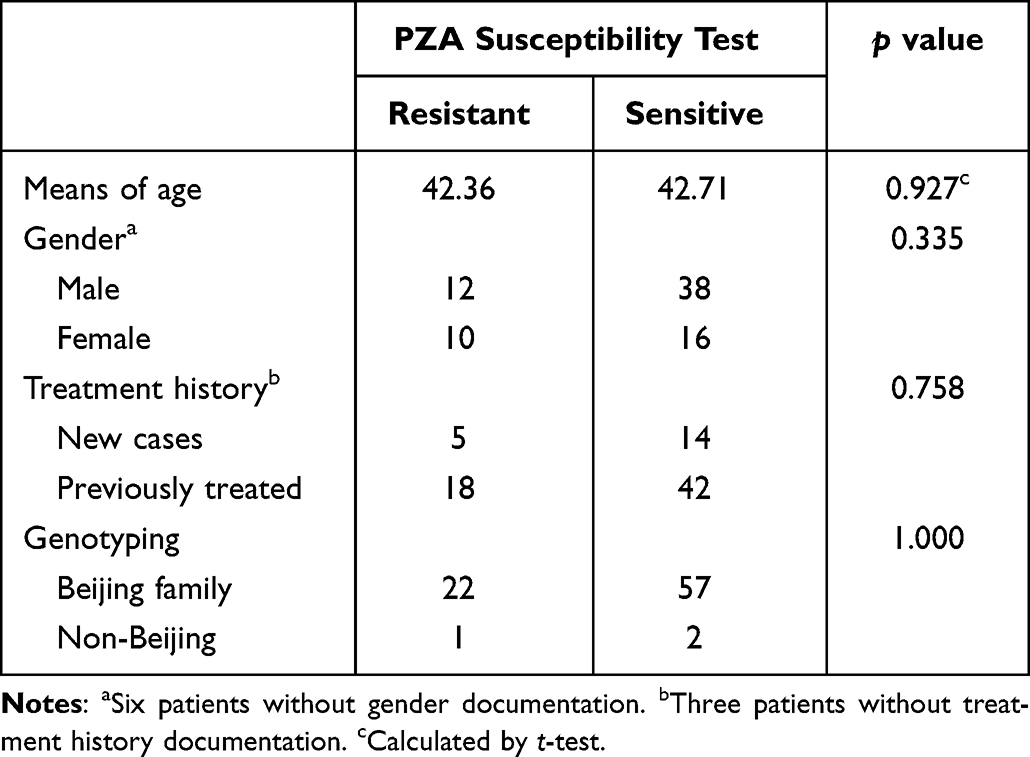 |
Table 1 Demographic Characteristics of Patients and Genotyping of Clinical Strains Resistant and Susceptible to PZA |
PZA Resistance and pncA Mutation Profiles
The results of the MGIT 960 assays revealed that 28.0% (23/82) of our MDR strains were phenotypically resistant to PZA when evaluated using the standard 100 mg/L concentration and that 15.9% (13/82) were resistant to 300 mg/L PZA. More than half (56.5%, 13/23) of the PZA-resistant MDR strains demonstrated continued PZA resistance even at very high concentrations (300 mg/L).
Our evaluations identified 36 mutation patterns in the pncA genes of our MDR clinical strains (Table 2), which occurred across 33 unique mutation sites (nucleotide positions). Of these, 29 were missense mutations, one was a nonsense mutation, three were deletions, two were insertions, and one was a synonymous mutation. A total of 40 strains were found to carry single mutations and only two strains encoded two point mutations each. Interestingly, both double point mutations occurred within the neighboring nucleotide positions of one codon, one was a TG→CC transition at nucleotide positions 104–105, and the other was a CG→GC transition at nucleotides 161 and 162. In addition, we noted a fairly even distribution of these mutations across the coding region of pncA with no evident mutational hotspots. Frequency evaluations revealed that the insertion at position 418 (codon 140) of pncA had the highest mutation frequency, being identified in four strains, while the other mutations occurred in no more than two individual strains. In addition, we identified a single large-fragment deletion in one strain that resulted in a 435-bp deletion from nucleotide 72 to 506 (Figure S1). Eleven types of pncA mutations identified in this study were also reported in the WHO TB GUIDE,50 and the DST results were congruous except for one mutation types (56 T→C).
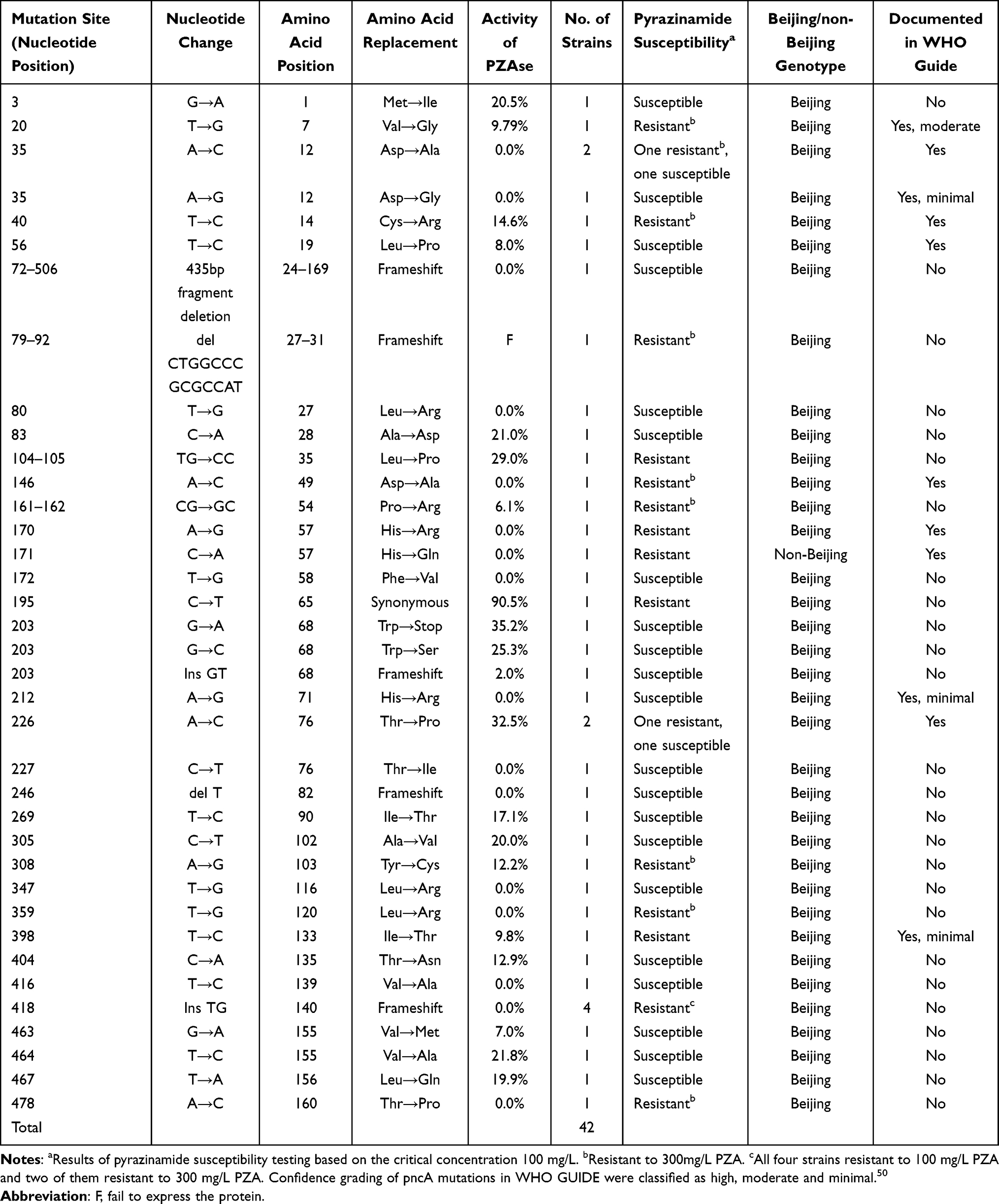 |
Table 2 Characteristics of pncA Mutation, PZA Susceptibility and Genotyping in pncA Mutated MDR Strains |
Concordance Between pncA Mutation and PZA Resistance
The concordance analyses for each pncA mutation and PZA resistance are shown in Table 3. We also noted that molecular evaluations could predict sensitivity to 100 mg/L PZA at a sensitivity of 82.6% (19/23) and a specificity of 61.0% (36/59). In addition, of the 42 MDR strains that encoded pncA mutations, 19 were shown to be PZA-resistant by the MGIT 960 method, and more than half (54.8%, 23/42) of these pncA-mutant MDR strains were shown to be PZA-sensitive. Despite this, the association between pncA mutation and PZA resistance was found to be significant (P = 0.001).
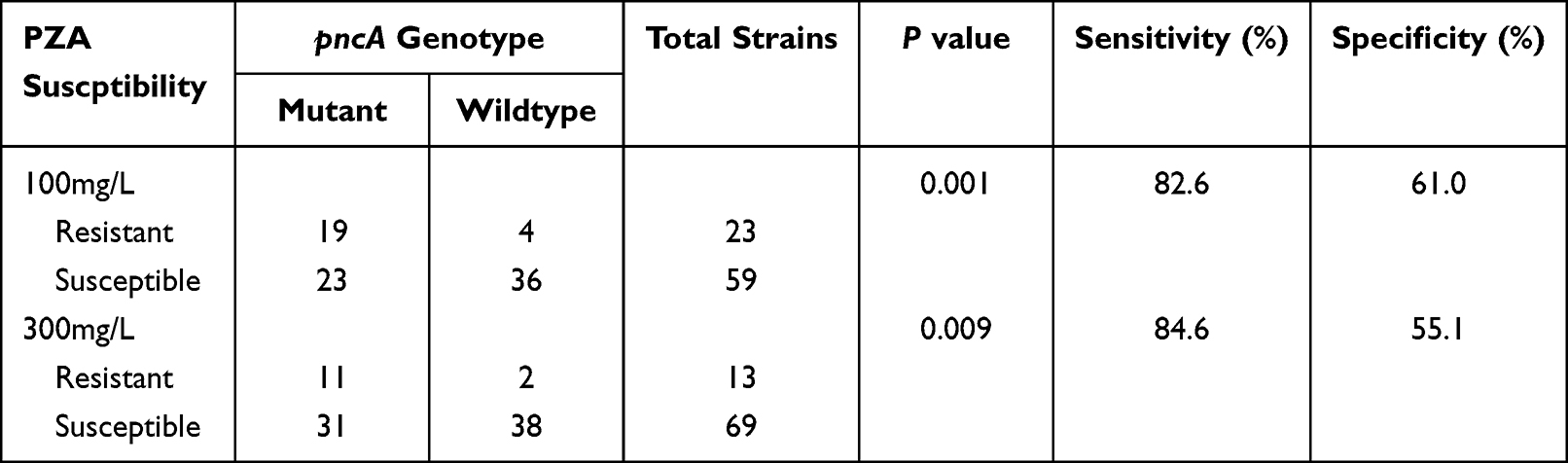 |
Table 3 Concordance of pncA Mutation Detection and PZA Susceptibility by MGIT 960 |
Evaluations of these strains at 300 mg/L PZA increased the sensitivity of the molecular identification assays to 84.6% (11/13), but the specificity was further reduced to only 55.1% (38/69). In addition, 11 of the MDR strains with pncA mutations were shown to be resistant to this increased concentration of PZA when evaluated using the MGIT 960 method, and 73.8% (31/42) of the pncA-mutant MDR strains were shown to be PZA-sensitive at 300 mg/L. The association between pncA mutation and increased PZA resistance was also significant (P = 0.009). However, it is worth noting that there were two extremely resistant strains that included no pncA mutations.
Concordance Between PZAse Enzymatic Activity and PZA Resistance
As previously mentioned, of the 23 phenotypically PZA-resistant strains, 19 were shown to have mutations in their pncA gene. We then identified 14 different mutation types in 18 strains that induced a significant decrease in PZAse activity (less than 40%). However, one strain, which encoded a 195 C→T mutation, synonymously maintained 90.5% of its relative PZAse activity. In addition, all nine mutations identified in the 13 highly PZA (300 mg/L)-resistant strains reduced the relative PZAse activity to 20% or less (Figure 1). Moreover, we identified 23 mutations in the pncA genes of the 23 phenotypically PZA-susceptible strains, with all of these mutations producing a significant decrease in PZAse activity (< 40%), and 56.5% (13/23) of these mutations even reduced the relative PZAse activity to less than 20%. It is worth noting that we failed to measure the relative enzymatic activity of the “79–92 del 14 bp” deletion owing to its early termination.
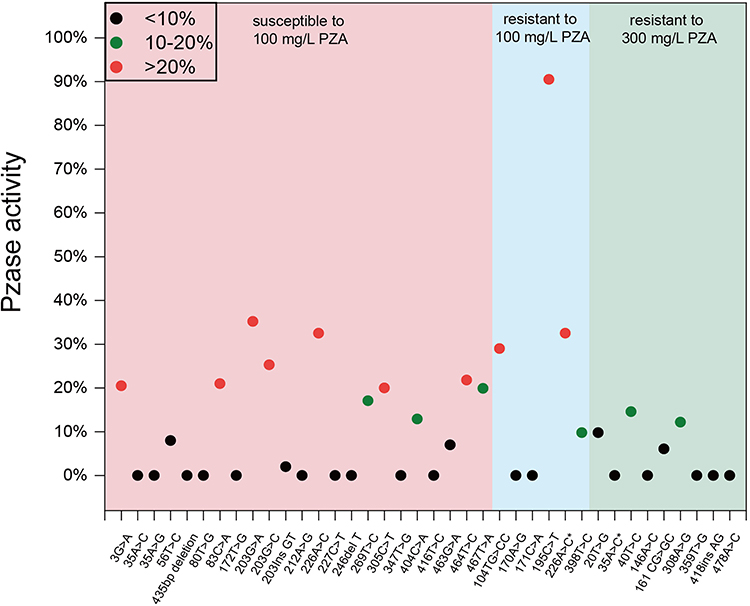 |
Figure 1 Pyrazinamidase (PZAse) activity of strains. Red: susceptible to pyrazinamide (PZA) at 100 mg/L; Blue: resistant to PZA at 100 mg/L; Green: resistant to PZA at 300 mg/L. *Mutation type with different phenotype. |
Discussion
PZA was first initiated as one of the four first-line drugs for intensive anti-TB treatment in the 1980s, making it a relative newcomer to TB chemotherapy when compared with INH and RIF.29 However, the recent emergence of both MDR- and extensively drug-resistant-TB has considerably narrowed the range of active therapeutics for the clinical management of TB. One recent study reported that a shorter regimen, in which the injectable agent is replaced by bedaquiline, is highly preferred for patients with resistance to fluoroquinolones.1 However, the all-oral bedaquiline-containing regimen still includes six agents, and most patients cannot afford bedaquiline because of its high price, which continues to hamper its application. Fortunately, recent research has shown that PZA molecular susceptibility testing could improve the treatment outcomes of MDR-TB without using new drugs in PZA-susceptible patients. This is further supported by the fact that recent results suggest that the current regimen could be shortened to 12 months with comparable success rates to those associated with the WHO-recommended regimen.9 This emphasizes the importance of PZA in the treatment of drug-resistant TB patients and shows that effective management of MDR-TB is not solely reliant on the development of novel medicines but may also benefit from the application of traditionally effective modalities as determined by DST evaluation.30–33
However, unlike the prevalence of other first-line drug resistance, the data around PZA resistance in clinical MDR M. tuberculosis strains remain limited. The divergent prevalence of PZA resistance in MDR M. tuberculosis clinical strains has been reported in several studies conducted across all six WHO-defined regions.34–37 A recent systemic review reported a pooled summarized prevalence estimate for PZA resistance of approximately 60.5% (95% CI 52.3–68.6%) in MDR-TB patients and 41.3% (95% CI 29.0–53.7%) in TB patients at high-risk of MDR-TB. Thus, the PZA resistance rate (28.0%) in our study is much lower than this mean value20 and may be the result of inaccuracies in PZA DST evaluations, or it may simply be a reflection of the less consistent use of PZA for MDR-TB treatment in Henan, China. However, if calculated from the accuracy of the molecular method, excluding the silence mutation, the mutation rate of the samples reached 50%(41/82), which is comparable to the previous studies.
There are many reports describing a wide variety of pncA mutations. The majority of these mutations can be classified as missense mutations, with a smaller proportion being described as short insertions or deletions, nonsense mutations in the coding region, or mutations in the putative promoter.38 All of these classes of mutations were found in our study, and we also identified a relatively rare large-fragment deletion in this gene. Although large-fragment genetic alterations are rarely reported, they often result in truncation.39 In the present study, we found that the majority of our mutations could be described as single point mutations, resulting in only single amino acid substitutions. We also noted that there were no mutational hotspots in this evaluation.
Given the difficulties in creating an accurate phenotypic PZA susceptibility test, it is only natural that evaluations of the concordance between the phenotypic and genotypic assays for PZA susceptibility have become more common.40–42 We used a routine critical concentration of PZA (100 mg/L) to complete the first round of sensitivity evaluations and then compared this with the predictive outcomes from our pncA mutation assays. These comparisons showed that our mutational evaluations had a concurrence of 82.6%, which is slightly higher than the values reported by studies in Thailand (75%)45 and Taiwan (80.6%)48 but is lower than that of the South African (91%) evaluations.47 The specificity of our study was only 61.0% at the recommended cutoff of 100 mg/L, which is much lower than that of some previous studies that reported specificities of more than 90%.44,46,47 However, compared to the mutation types documented in the TB GUIDE that were also identified in our study, the results showed a high degree of consistency.
Some of our sequencing and DST results were shown to be slightly contradictory, including the observation that our “large-fragment deletion at 435 bp” mutation, which should induce a code shift and present with PZA resistance, was shown to be susceptible in the DST assays. Similarly, one strain with a T deletion at position 246 and a strain with a GT insertion at position 203 were all shown to be susceptible to PZA. In addition, the DST result of “56 T→C” strain were susceptible, which was in conflict with that reported in the WHO TB GUIDE too. Conversely, one strain with a “195 C→T” mutation, which induced a synonymous change, recorded a resistant phenotype to PZA.49 These conflicting results were further verified by evaluating the PZAse activity of these strains, which revealed that the PZAse activity of the “246 del T”, “203 in GT”, “56 T→C” and “435 bp del” was lower than 10%, while the relative PZAse activity of the “195 C→T” strain remained higher than 90%, which is consistent with synonymous mutation. All of the PZAse activity results were in conflict with the drug sensitivity assays but were consistent with the sequencing results. In addition, there were 23 strains with mutations in pncA that all demonstrated a PZA-susceptible phenotype despite an overall reduction in their PZAse activity to less than 40% of the wild-type (WT). In fact, 13 strains reported a relative activity of less than 10% of the WT strain. These results indicate that the phenotypic assays are not necessarily reliable as they suffer from some degree of user inconsistencies.
The inconsistency has been noted in other studies, and some authors have suggested that these discordances might be overcome by using phenotypic evaluations of several critical concentrations of PZA (MICs of ≤64 mg/L, = 128 mg/L, and >128 mg/L) within the BACTEC systems. Werngren et al also suggested that isolates could be classified as susceptible, intermediary, and resistant, respectively.43 This strategy allows for a potential decrease in both the false resistance and false susceptibility rates by adding evaluations at both lower and higher breakpoints for defining susceptibility and resistance, respectively. Other studies have suggested evaluations at a higher concentration (300 mg/L or 200 mg/L) of PZA as the breakpoint for PZA drug susceptibility.26,31,48 Thus, we continued to evaluate this higher (300 mg/L) breakpoint and reevaluated our data at this concentration. These evaluations resulted in only a slight increase in the sensitivity of the pncA mutation assays (84.6%), but the specificity of these assays was reduced to as low as 55.1%, suggesting that adjusting the critical concentration of PZA is not enough to optimize PZA DST. More detailed experiments are needed to develop a more robust protocol for PZA susceptibility evaluations in MDR-TB strains.
Previous studies have shown that the concordance between the phenotypic and genotypic evaluations of PZA susceptibility in MDR-TB was closely associated with the specific genetic lineage of these strains, such as Beijing/non-Beijing genotypes.40,46 Dormandy et al reported that all 29 W-Beijing family MDR M. tuberculosis strains carrying the same type of nonsynonymous pncA mutation showed susceptibility to PZA.40 One study conducted in Taiwan using 66 MDR M. tuberculosis strains produced a higher concordance for pncA mutation detection and PZA resistance testing using the MGIT 960 method in non-Beijing strains compared to Beijing strains.42 In their study, the sensitivity and specificity of their pncA mutation detection in Beijing strains were 76.2% and 93.8%, respectively, with both values increasing in non-Beijing strains (86.7% and 100.0%, respectively).40 These studies suggest that the concordance between pncA mutation detection and automatic broth-based PZA resistance detection systems may not be as good in Beijing family MDR M. tuberculosis strains as that in non-Beijing family strains. Since 96% of the MDR strains in our study were classified as the Beijing genotype, their genetic lineage might explain the differences in the sensitivity and specificity of our assays when compared to other studies conducted in different places.
Although this study expands our understanding of pncA mutations in MDR-TB, there were several limitations. First, all of the MDR strains included in this study were sampled from Henan, China preventing any geographical evaluation. Second, we did not include any upstream pncA mutations, which may decrease the sensitivity of the evaluations. Nevertheless, our research may be used to determine the MIC of PZA using a reduced inoculum, via the BACTEC MGIT 960 method, to aid the interpretation of genotypic DST data while minimizing false resistance. Further, we intend to collect more samples from different regions for analysis to overcome the geographic limitations.
Conclusion
Our study illustrates the complexities and challenges associated with PZA susceptibility of M. tuberculosis using DST, molecular analysis, and enzyme activity determination. We found that the association between pncA mutation and PZA resistance was significant, and the molecular method for detection of PZA resistance is more reliable than the phenotype method. The introduction of the PZAse assay also has a higher consistency with the molecular method. In addition, the BACTEC MGIT 960 automatic method, needs to be established to facilitate both the epidemiological analysis and clinical evaluation of MDR-TB treatment and further our understanding of the relationship between specific mutations and phenotypes.
Acknowledgments
We thank Yuling Zhao and Jie Shi from Henan CDC and Wei Zhen from IPB-CAMS for their technical assistance in the procedures for bacterium isolation, culture, drug susceptibility testing, and PCR amplification.
This work was supported by the Xiamen Major Science and Technology Projects (3502Z20191007), the Xiamen City Science and Technology Plan Project (3502Z20203013), the Social Development Project Cooperation Program for University and Industry in Fujian Province (2021Y4001), and the Shenzhen Fundamental Research Program (JCYJ20180306170526435).
Disclosure
The authors report no conflicts of interest in this work.
References
1. World Health Organization. Global tuberculosis report 2021. World Health Organization; 2021.
2. Gandhi NR, Nunn P, Dheda K, et al. Multidrug-resistant and extensively drug-resistant tuberculosis: a threat to global control of tuberculosis. Lancet. 2010;375(9728):1830–1843. doi:10.1016/S0140-6736(10)60410-2
3. Sun F, Li Y, Chen Y, et al. Introducing molecular testing pyrazinamide susceptibility improves multidrug-resistant tuberculosis treatment outcomes: a prospective cohort study. Eur Respir J. 2019;53(3):1801770. doi:10.1183/13993003.01770-2018
4. Zhang Y, Mitchison D, Shi W, Zhang W. Mechanisms of pyrazinamide action and resistance. Microbiol Spectr. 2014;2:1–12. doi:10.1128/microbiolspec.MGM2-0023-2013
5. Nahid P, Dorman SE, Alipanah N, et al. Executive Summary: official American Thoracic Society/Centers for Disease Control and Prevention/Infectious Diseases Society of America Clinical Practice Guidelines: treatment of drug-susceptible tuberculosis. Clin Infect Dis. 2016;63:e147. doi:10.1093/cid/ciw376
6. Kempker RR, Heinrichs MT, Nikolaishvili K, et al. Lung tissue concentrations of pyrazinamide among patients with drug-resistant pulmonary tuberculosis. Antimicrob Agents Chemother. 2017;61. doi:10.1128/AAC.00226-17
7. Somner AR. A controlled trial of six months chemotherapy in pulmonary tuberculosis. First report: results during chemotherapy. Br J Dis Chest. 1981;75:141–153. doi:10.1016/0007-0971(81)90046-2
8. World Health Organization. Rapid communication: key changes to treatment of multidrug-and rifampicin- resistant tuberculosis (MDR/RR-TB). World Health Organization; 2018.
9. Van Deun A, Maug AKJ, Salim MAH, et al. Short, highly effective, and inexpensive standardized treatment of multidrug-resistant tuberculosis. Am J Respir Crit Care Med. 2010;182(5):684–692. doi:10.1164/rccm.201001-0077OC
10. Falzon D, Schünemann HJ, Harausz E, et al. World Health Organization treatment guidelines for drug-resistant tuberculosis, 2016 update. Eur Respir J. 2017;49:1602308. doi:10.1183/13993003.02308-2016
11. Moodley R, Godec TR. Short-course treatment for multidrug-resistant tuberculosis: the STREAM trials. Eur Respir Rev. 2016;25:29–35. doi:10.1183/16000617.0080-2015
12. Lan N, Master I, Phillips PP, et al. A trial of a shorter regimen for rifampin-resistant tuberculosis. NEJM. 2019;380:1201–1213.
13. Bouton TC, Phillips PPJ, Mitnick CD, et al. An optimized background regimen design to evaluate the contribution of levofloxacin to multidrug-resistant tuberculosis treatment regimens: study protocol for a randomized controlled trial. Trials. 2017;18:563. doi:10.1186/s13063-017-2292-x
14. Cellamare M, Ventz S, Baudin E, Mitnick CD, Trippa L. A Bayesian response-adaptive trial in tuberculosis: the end-TB trial. Clin Trials. 2016;14:17–28.
15. Weng T, Sun F, Li Y, et al. Refining MDR-TB treatment regimens for ultra short therapy (TB-TRUST): study protocol for a randomized controlled trial. BMC Infect Dis. 2021;21(1). doi:10.1186/s12879-021-05870-w
16. Hussain Z, Zhu J, Ma X. Metabolism and hepatotoxicity of pyrazinamide, an antituberculosis drug. Drug Metab Dispos. 2021;49(8):679–682. doi:10.1124/dmd.121.000389
17. Scorpio A, Lindholm-Levy P, Heifets L, et al. Characterization of pncA mutations in pyrazinamide-resistant Mycobacterium tuberculosis. Antimicrob Agents Chemother. 1997;41(3):540–543. doi:10.1128/AAC.41.3.540
18. Mestdagh M, Fonteyne PA, Realini L, et al. Relationship between pyrazinamide resistance, loss of pyrazinamidase activity, and mutations in the pncA locus in multidrug-resistant clinical isolates of Mycobacterium tuberculosis. Antimicrob Agents Chemother. 1999;43(9):2317–2319. doi:10.1128/AAC.43.9.2317
19. Stoffels K, Mathys V, Fauville-Dufaux M, Wintjens R, Bifani P. Systematic analysis of pyrazinamide-resistant spontaneous mutants and clinical isolates of Mycobacterium tuberculosis. Antimicrob Agents Chemother. 2012;56(10):5186–5193.
20. Whitfield MG, Soeters HM, Warren RM, et al. A global perspective on pyrazinamide resistance: systematic review and meta-analysis. PLoS One. 2015;10(7):e0133869. doi:10.1371/journal.pone.0133869
21. Shi D, Li L, Zhao Y, et al. Characteristics of embB mutations in multidrug-resistant Mycobacterium tuberculosis isolates in Henan, China. J Antimicrob Chemother. 2011;66:2240–2247. doi:10.1093/jac/dkr284
22. Sheen P, Ferrer P, Gilman RH, et al. Effect of pyrazinamidase activity on pyrazinamide resistance in Mycobacterium tuberculosis. Tuberculosis. 2009;89:109–113. doi:10.1016/j.tube.2009.01.004
23. Zhang L, Chen J, Shen X, et al. Highly polymorphic variable-number tandem repeats loci for differentiating Beijing genotype strains of Mycobacterium tuberculosis in Shanghai, China. FEMS Microbiol Lett. 2008;282:22–31. doi:10.1111/j.1574-6968.2008.01081.x
24. Chen J, Tsolaki AG, Shen X, Jiang X, Mei J, Gao Q. Deletion-targeted multiplex PCR (DTM-PCR) for identification of Beijing/W genotypes of Mycobacterium tuberculosis. Tuberculosis. 2007;87:446–449. doi:10.1016/j.tube.2007.05.014
25. O’Mahony J, Hill C. Rapid real-time PCR assay for detection and quantitation of Mycobacterium avium subsp. paratuberculosis DNA in artificially contaminated milk. Appl Environ Microbiol. 2004;70(8):4561–4568. doi:10.1128/AEM.70.8.4561-4568.2004
26. Sheen P. Molecular Diagnosis of Pyrazinamide Resistance and Molecular Understanding of the Pyrazinamidase Functionality in Mycobacterium Tuberculosis. Baltimore, MD: Johns Hopkins University; 2008.
27. Gopal P, Grüber G, Dartois V, Dick T. Pharmacological and molecular mechanisms behind the sterilizing activity of pyrazinamide. Trends Pharmacol Sci. 2019;40(12):930–940. doi:10.1016/j.tips.2019.10.005
28. Aono A, Chikamatsu K, Yamada H, Kato T, Mitarai S. Association between pncA gene mutations, pyrazinamidase activity, and pyrazinamide susceptibility testing in Mycobacterium tuberculosis. Antimicrob Agents Chemother. 2014;58:4928–4930. doi:10.1128/AAC.02394-14
29. Singh P, Mishra AK, Malonia SK, et al. The paradox of pyrazinamide: an update on the molecular mechanisms of pyrazinamide resistance in Mycobacteria. J Commun Dis. 2006;38:288–298.
30. Chinese Ministry of Health. National Baseline Survey of Drug-Resistant Tuberculosis (2007–2008). People’s Medical Publishing House; 2010.
31. World Health Organization. Multidrug and extensively drug-resistant TB (M/XDR-TB): 2010 global report on surveillance and response. WHO/HTM/TB/2010.3. Geneva, Switzerland: World Health Organization; 2010.
32. World Health Organization. Guidelines for the programmatic management of drug-resistant tuberculosis −2011 update. WHO/HTM/TB/2011.6. Geneva, Switzerland: World Health Organization; 2011.
33. Li S-Y, Tasneen R, Tyagi S, et al. Bactericidal and sterilizing activity of a novel regimen with Bedaquiline, Pretomanid, Moxifloxacin, and pyrazinamide in a murine model of tuberculosis. Antimicrob Agents Chemother. 2017;61(9):
34. Njire M, Tan Y, Mugweru J, et al. Pyrazinamide resistance in mycobacterium tuberculosis: review and update. Adv Med Sci. 2016;61(1):63–71. doi:10.1016/j.advms.2015.09.007
35. Salfinger M, Heifets LB. Determination of pyrazinamide MICs for Mycobacterium tuberculosis at different pHs by the radiometric method. Antimicrob Agents Chemother. 1988;32:1002–1004. doi:10.1128/AAC.32.7.1002
36. Heifets L. Susceptibility testing of Mycobacterium tuberculosis to pyrazinamide. J Med Microbiol. 2002;51:11–12.
37. Pfyffer GE, Palicova F, Rusch-Gerdes S. Testing of susceptibility of Mycobacterium tuberculosis to pyrazinamide with the nonradiometric BACTEC MGIT 960 system. J Clin Microbiol. 2002;40:1670–1674. doi:10.1128/JCM.40.5.1670-1674.2002
38. Zhang Y, Mitchison D. The curious characteristics of pyrazinamide: a review. Int J Tuberc Lung Dis. 2003;7:6–21.
39. Lemaitre N, Sougakoff W, Truffot-Pernot C, Jarlier V. Characterization of new mutations in pyrazinamide-resistant strains of Mycobacterium tuberculosis and identification of conserved regions important for the catalytic activity of the pyrazinamidase PncA. Antimicrob Agents Chemother. 1999;43:1761–1763. doi:10.1128/AAC.43.7.1761
40. Dormandy J, Somoskovi A, Kreiswirth BN, Driscoll JR, Ashkin D, Salfinger M. Discrepant results between pyrazinamide susceptibility testing by the reference BACTEC 460TB method and pncA DNA sequencing in patients infected with multidrug-resistant W-Beijing Mycobacterium tuberculosis strains. Chest. 2007;131:497–501. doi:10.1378/chest.06-1899
41. Scarparo C, Ricordi P, Ruggiero G, Piccoli P. Evaluation of the fully automated BACTEC MGIT 960 system for testing susceptibility of Mycobacterium tuberculosis to pyrazinamide, streptomycin, isoniazid, rifampin, and ethambutol and comparison with the radiometric BACTEC 460TB method. J Clin Microbiol. 2004;42:1109–1114. doi:10.1128/JCM.42.3.1109-1114.2004
42. Chedore P, Bertucci L, Wolfe J, Sharma M, Jamieson F. Potential for erroneous results indicating resistance when using the Bactec MGIT 960 system for testing susceptibility of Mycobacterium tuberculosis to pyrazinamide. J Clin Microbiol. 2010;48:300–301. doi:10.1128/JCM.01775-09
43. Werngren J, Sturegard E, Jureen P, Angebyc K, Hoffner S, Schon T. Reevaluation of the critical concentration for drug susceptibility testing of Mycobacterium tuberculosis against pyrazinamide using wild-type MIC distributions and pncA gene sequencing. Antimicrob Agents Chemother. 2012;56:1253–1257. doi:10.1128/AAC.05894-11
44. Simons SO, van Ingen J, van der Laan T, et al. Validation of pncA gene sequencing in combination with the mycobacterial growth indicator tube method to test susceptibility of Mycobacterium tuberculosis to pyrazinamide. J Clin Microbiol. 2012;50:428–434. doi:10.1128/JCM.05435-11
45. Jonmalung J, Prammananan T, Leechawengwongs M, Chaiprasert A. Surveillance of pyrazinamide susceptibility among multidrug-resistant Mycobacterium tuberculosis isolates from Siriraj Hospital, Thailand. BMC Microbiol. 2010;10:223. doi:10.1186/1471-2180-10-223
46. Chiu YC, Huang SF, Yu KW, Lee YC, Feng JY, Su WJ. Characteristics of pncA mutations in multidrug-resistant tuberculosis in Taiwan. BMC Infect Dis. 2011;11:240. doi:10.1186/1471-2334-11-240
47. Mphahlele M, Syre H, Valvatne H, et al. Pyrazinamide resistance among South African multidrug-resistant Mycobacterium tuberculosis isolates. J Clin Microbiol. 2008;46:3459–3464. doi:10.1128/JCM.00973-08
48. Shi W, Zhang X, Jiang X, Yuan H, Lee JS, Barry CE 3rd. Pyrazinamide inhibits trans-translation in Mycobacterium tuberculosis. Science. 2011;333(6049):1630–1632. doi:10.1126/science.1208813
49. Senqstake S, Berqval IL, Schuitema AR, et al. Pyrazinamide resistance-conferring mutations in pncA and the transmission of multidrug-resistant TB in Georgia. BMC Infect Dis. 2017;17(1):491. doi:10.1186/s12879-017-2594-3
50. World Health Organization. The use of next-generation sequencing technologies for the detection of mutations associated with drug resistance in Mycobacterium tuberculosis complex: technical guide. World Health Organization; 2018.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.