")
Back to Journals » OncoTargets and Therapy » Volume 8
Protein tyrosine phosphatase SHP-1 sensitizes EGFR/HER-2 positive breast cancer cells to trastuzumab through modulating phosphorylation of EGFR and HER-2
Authors Wu Y, Li R, Zhang J, Wang G, Liu B, Huang X, Zhang T, Luo R
Received 4 February 2015
Accepted for publication 2 April 2015
Published 14 September 2015 Volume 2015:8 Pages 2577—2587
DOI https://doi.org/10.2147/OTT.S82225
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jianmin Xu
Yifen Wu,1,2,* Rong Li,3,* Junyi Zhang,4 Gang Wang,5 Bin Liu,6 Xiaofang Huang,7 Tao Zhang,7 Rongcheng Luo8
1Traditional Chinese Medicine-Integrated Hospital, Southern Medical University, Guangzhou, 2Department of Oncology, Dongguan People’s Hospital, Dongguan, 3Department of Oncology, Nanfang Hospital, 4Department of Oncology, Traditional Chinese Medicine-Integrated Hospital, Southern Medical University, Guangzhou, 5Department of Radiology, Dongguan People’s Hospital, Dongguan, 6Second Affiliated Hospital of Guangzhou Medical College, 7College of Traditional Chinese medicine, Southern Medical University, 8Cancer Center, Traditional Chinese Medicine-Integrated Hospital, Southern Medical University, Guangzhou, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Background: Trastuzumab resistance in HER-2 positive breast cancer cells is closely related to overexpression of both epidermal growth factor receptor (EGFR) and human epidermal receptor (HER-2). SHP-1 has been demonstrated to downregulate tyrosine kinase activity including EGFR via its phosphatase function, but its effect on HER-2 activity is still unknown. Here, we examined the hypothesis that SHP-1 enhances the anticancer efficacy of trastuzumab in EGFR/HER-2 positive breast cancer cells through combining dual inhibition of EGFR and HER-2.
Methods: Trastuzumab-resistant breast cancer SKBr-3 cells were generated by long-term in vitro culture of SKBr-3cells in the presence of trastuzumab. The SHP-1 was ectopically expressed by stable transfection. The activity and expression of EGFR, HER-2, and downstream signaling pathways were tested by Western blot. Cell viability was examined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, and apoptosis was examined by flow cytometry. The binding between SHP-1 and EGFR/HER-2 was evaluated by immunoprecipitation assay and bimolecular fluorescence complementation. The effects of SHP-1 on tumorigenicity and trastuzumab sensitivity were confirmed via in vivo xenograft model.
Results: Trastuzumab-resistant SKBr-3 cells showed aberrant co-expression of EGFR and HER-2. Introduction of wild-type SHP-1 inhibited cell proliferation, clone formation, and promoted the apoptosis induced by trastuzumab. Meanwhile, SHP-1 overexpression reduced phosphorylation levels of EGFR and HER-2 both in parental and trastuzumab-resistant SKBr-3 cells. In vivo study showed an increased antitumor effect of trastuzumab in SHP-1 overexpressed xenografts. At last, we discovered that SHP-1 can make complexes with both EGFR and HER-2, and both phospho-EGFR and phosphor-HER-2 levels in wild-type SHP-1 immunoprecipitates were less than those in phosphatase-inactive SHP-1 (C453S) immunoprecipitates, indicating that EGFR and HER-2 are potential substrates of SHP-1.
Conclusion: Taken together, we have demonstrated that the SHP-1 is a negative regulatory factor of the tyrosine kinase activity of HER-2 and EGFR through inhibiting phosphorylation. Dual targeting of EGFR and HER-2, by combining trastuzumab with SHP-1 overexpression, may improve response in HER-2 overexpressing breast cancer cells that also express high levels of EGFR.
Keywords: breast cancer, trastuzumab, EGFR, HER-2, SHP-1, drug resistance
Background
Breast cancer is a leading cause of cancer-related death in women; it is estimated that there will be 39,620 breast cancer related deaths among women in the United States1 and 88,886 in Europe2 in 2013. HER-2 is a transmembrane tyrosine kinase receptor that belongs to the human epidermal growth factor receptor (EGFR/HER) family, which activates various intracellular downstream oncogenic signaling pathways.3 HER-2 amplification occurs in approximately 20%–30% of patients with breast cancer,4,5 and it correlates with more aggressive biological features of tumor such as poor clinical outcome, high rates of metastasis, and resistance to conventional treatment modalities.6 Trastuzumab (Herceptin®), a humanized recombinant monoclonal antibody that binds to the extracellular domain of HER-2, has been recommended as first-line treatment for HER-2 positive patients and shown to improve outcome. However, not all patients with HER-2 positive respond to trastuzumab treatment; nearly 15% of patients will relapse with distant disease within 12 months of treatment with trastuzumab,7 and only about 26% of HER-2-overexpressing metastatic breast cancer patients showed response to trastuzumab monotherapy as first-line treatment.8 The innate and acquired trastuzumab resistance has increasingly occurred and remains the biggest clinical challenge.9
Epidermal growth factor receptor (EGFR) is overexpressed in 35%–65% of HER-2-positive breast cancers.10–12 There is evidence that the EGFR overexpression influences response to HER-2 inhibition, impairs response to trastuzumab in HER-2 positive breast cancers, and correlates with poor survival.13,14 Therefore, combining trastuzumab with EGFR-targeted strategy may improve response to trastuzumab treatment.15 It is well known that phosphorylation of proteins at specific tyrosine residues has an important role in regulating the activity of EGFR/HER family proteins and downstream pathways. Biologically, the de-phosphorylation of tyrosine residues in proteins is precisely regulated by protein tyrosine phosphatases (PTPs), and defects in PTPs can lead to the intracellular accumulation of tyrosine-phosphorylated proteins, which can cause abnormal cell proliferation and differentiation.16,17 HERs are major substrates of PTPs in breast cancer18 cells, and ectopic expression of some PTPs in human breast cancer cells decreased cell proliferation and tumorigenicity,19,20 which suggests their role as a tumor suppressor. In consideration of the dual inhibitory effect of PTPs on both EGFR and HERs, it is reasonable to speculate that some members of PTPs might be potential therapy targets and markers.
SHP-1, a non-receptor PTP containing an SH2 domain, is a key regulator that controls the intracellular phosphotyrosine level. Extensive studies on SHP-1 revealed that the expression of SHP-1 protein was decreased or absent in many kinds of cancer cell lines and tissues. Abolished expression in leukemia and lymphoma has been related to both malignant transformation and malignant tumor behavior.21,22 Ectopic expression of SHP-1 in the breast cancer cell line MDAMB-231 impaired cell proliferation two- to threefold. In addition, SHP-1 has also been shown to be involved in regulating several signals including EGFR and other tyrosine kinase receptors.23 The findings mentioned earlier suggests a potential key role of SHP-1 in resistance to trastuzumab, but the role of SHP-1 in trastuzumab resistance remains unclear. The aim of this study is to elucidate the role of SHP-1 protein expression in relation to trastuzumab treatment and resistance in HER-2 positive breast cancer.
Materials and methods
Cell culture
The human breast cancer SKBr-3 and human embryonic kidney 293 (HEK293) cell lines were obtained from American Type Culture Collection (Manassas, VA, USA). SKBr-3 cells were cultured in RPMI 1640 media (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum and HEK293 cells were cultured in DMEM high glucose medium (Thermo Fisher Scientific) containing 10% fetal bovine serum in a humidified 5% CO2 atmosphere at 37°C.
Establishment of trastuzumab-resistant cells
Trastuzumab/Herceptin (Hoffman-La Roche Ltd., Basel, Switzerland) was dissolved in sterile water. Trastuzumab-resistant cells were generated through continuous culture of SKBr-3 cells in the presence of 5 μg/mL trastuzumab for 6 months, as reported previously.24 Subsequently, the established trastuzumab-resistant SKBr-3 cells were cultured in medium containing 5 μg/mL trastuzumab.
Plasmids and virus infection
SHP-1 cDNA was amplified by polymerase chain reaction (PCR) from total RNA extracted from SHP-1 overexpressed MCF-7 cells and ligated into pCDNA3.1. The catalytically inactive (C453S) SHP-1 was generated by introducing a point mutation using standard PCR techniques. Lentiviral particles were produced by transfecting 239T cells, the viral supernatants were collected after 72 hours following transfection, the particles were concentrated by using Lenti-X™ Concentrator overnight at 4°C (Clontech, Mountain View, CA, USA), and the aliquots were stored at −80°C. The titers of the concentrated particles were measured before using. For bimolecular fluorescence complementation (BiFC), the pBiFC-VC155 plasmid (#22011, deposited by Chang-Deng Hu) and pBiFC-VN173 plasmid (#22010, deposited by Chang-Deng Hu) were obtained from Addgene (Cambridge, MA, USA). The SHP-1 gene was amplified from MCF-7 cells, and the PCR product was digested with KpnI and EcoRI, and then ligated into pBiFC-VN173. EGFR, HER-2, HER-3, and HER-4 genes were amplified from total RNA extracted from SKBr-3 cells and ligated into pBiFC-VC155.
Cell proliferation analysis
MTT substrate (Sigma-Aldrich, St Louis, MO, USA) was used to assay cell proliferation according to the manufacturer’s instructions. Briefly, a total of 5×103 cells were seeded into 96-well dishes and allowed to adhere overnight; trastuzumab of indicated concentration was then added into the medium. The medium was replaced by 100 μL fresh serum-free medium containing 0.5 g/L MTT 24 hours later, followed by 4-hour incubation. Then the MTT medium was replaced by 50 μL of DMSO for further 10 minutes incubation at 37°C. The A490 value of each sample was measured.
Colony formation assay
For colony formation assay, transfected cells were placed in a fresh six-well plate with a density of 500 cells per well and maintained in DMEM medium containing 10% fetal bovine serum and trastuzumab of indicated concentration at 37°C and 5% CO2 for 10 days. Colonies were fixed with methanol and stained with 0.1% crystal violet and counted.
Quantitative reverse transcription PCR analysis
Total RNA was extracted from each cell line using TRIzol reagent (Thermo Fisher Scientific) according to the manufacturer’s protocol. Reverse transcription was performed using M-MLV reverse transcriptase (Thermo Fisher Scientific) and quantitative PCR was performed with SYBR Green Master Mix (Hoffman-La Roche Ltd.). Relative expression values were calculated (ΔΔCT method) using GAPDH as a normalizer. The PCR primers sequences were: SHP-1, forward, 5′-GGAGAAGTTTGCGACTCTGAC-3′, reverse, 5′-GCGGGTACTTGAGGTGGATG-3′; EGFR, forward, 5′-CCCACTCATGCTCTACAACCC-3′, reverse, 5′-TCGCACTTCTTACACTTGCGG-3′; HER-2, forward, 5′-TGCAGGGAAACCTGGAACTC-3′, reverse, 5′-ACAGGGGTGGTATTGTTCAGC-3′.
Flow cytometry
Cells were plated into six-well plates at a density of 3×105 cells per well, and 24 hours later cells were treated with trastuzumab of indicated concentrations. After 24 hours exposure to trastuzumab, cells were stained with Annexin V-FITC and propidium iodide, and then flow cytometry was performed to detect apoptosis of the transfected cells.
Immunoblotting
Cells were treated as indicated earlier and were placed on ice and washed with PBS. Washed cells were lysed in RIPA lysis buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% sodium deoxycholate, 1% Triton X-100, and 0.1% SDS) plus protease inhibitor cocktail (Hoffman-La Roche Ltd.). Equal amounts of protein sample were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS–PAGE), transferred to polyvinylidene difluoride (PVDF) membrane (EMD Millipore, Billerica, MA, USA). Membranes were blocked with Tris-buffered saline containing Tween-20 and 5% (w/v) fat-free milk powder, and then incubated with appropriate primary antibodies at 4°C overnight followed by horseradish peroxidase-conjugated secondary antibodies. All antibodies were purchased from Cell Signaling Technologies: phospho-HER2 (Tyr1248) rabbit polyclonal antibody (#2247), HER2 rabbit polyclonal antibody (#2242), phospho-EGFR (Tyr1173) rabbit monoclonal antibody (#4407), EGFR rabbit monoclonal antibody (#4405), phospho-Erk1/2 (Thr202/Tyr204) rabbit monoclonal antibody (#4370), Erk1/2 rabbit monoclonal antibody (#4695), phospho-Stat3 (Tyr705) rabbit monoclonal antibody (#9145), Stat3 rabbit monoclonal antibody (#4904), phospho-Akt (Ser473) rabbit monoclonal antibody (#4060), and Akt (pan) rabbit monoclonal antibody (#4685).
Quantification and comparison of the intensity of bands of Western blot was done by using ImageJ software. Briefly, the images of Western blot results were converted to grayscale, the background were subtracted, the blot areas were selected, and the corresponding area and mean gray value (GV) were recorded. The GV intensity = area × mean GV. The GV of each protein was normalized to that of GAPDH, and the GV of each phosphorylated protein was normalized to the GV of its own corresponding total protein.
Co-immunoprecipitation
The co-immunopreciptation experiment was carried out with the Pierce® Co-Immunoprecipitation Kit according to the manufacturer’s protocol using the following antibodies: EGF Receptor (D38B1) XP® Rabbit mAb (#4267, Cell Signaling Technology, 1:500), HER2 (D8F12) XP® Rabbit mAb (#4290, Cell Signaling Technology, 1:500), HER3 (1B2E) Rabbit mAb (#4754, Cell Signaling Technology, 1:500), HER4 (111B2) Rabbit mAb (#4795, Cell Signaling Technology, 1:500), Anti-rabbit IgG, HRP-linked Antibody (#7074S, Cell Signaling Technology, 1:1,500).
In vivo tumor growth and mouse survival assay
BALC/nu/nu mice were randomly grouped to monitor tumor growth (n=10) and mouse survival (n=20). Trastuzumab-resistant SKBr-3 cells stably transfected vectors expressing SHP-1 or blank vectors and were subcutaneously injected (2×106) into the right mammary fat pad of each nude mice. After 1 week, mice were intravenously injected with 10 mg/kg trastuzumab twice a week. Tumor size was measured with the following formula: volume = (L×W2)/2, where L and W are the longest and shortest diameters, respectively. Mice were sacrificed 45 days post-first trastuzumab injection, and tumors were separated for weighing. For survival assay, the survival of mice in each group was recorded, and the ratios of surviving mice were plotted. All experimental procedures involving animals were in accordance with the Guide for the Care and Use of Laboratory Animals and were performed according to the institutional ethical guidelines for animal experiment. The study protocol was also approved by the Committee on the Use of Live Animals in Teaching and Research, Southern Medical University, Guangzhou, Guangdong, People’s Republic of China.
Results
EGFR and HER-2 are aberrantly expressed and phosphorylated in trastuzumab-resistant cells.
The HER-2 overexpressed human breast cancer SKBr-3 cells were cultured in medium containing 5 μg/mL trastuzumab continuously for 6 months, which resulting in the acquisition of trastuzumab resistance. Compared with the parental SKBr-3 cells, MTT assay showed that the trastuzumab-resistant cells displayed a significantly higher viability or proliferative capacity in the presence of every different concentration of trastuzumab (Figure 1A). Besides, the resistant SKBr-3 cells displayed dramatically increased colony formation ability under the treatment of trastuzumab (Figure 1B). We next assessed EGFR and HER-2 expression in the parental cells and trastuzumab-resistant cells, and found that both the mRNA and protein levels of EGFR and HER-2 were increased in the trastuzumab-resistant cells (Figure 1C and D). Results also showed that the phosphorylation levels of EGFR and HER-2 as well as the key kinases of their downstream pathways which including STAT3, Akt, and ERK were significantly enhanced (Figure 1D and E).
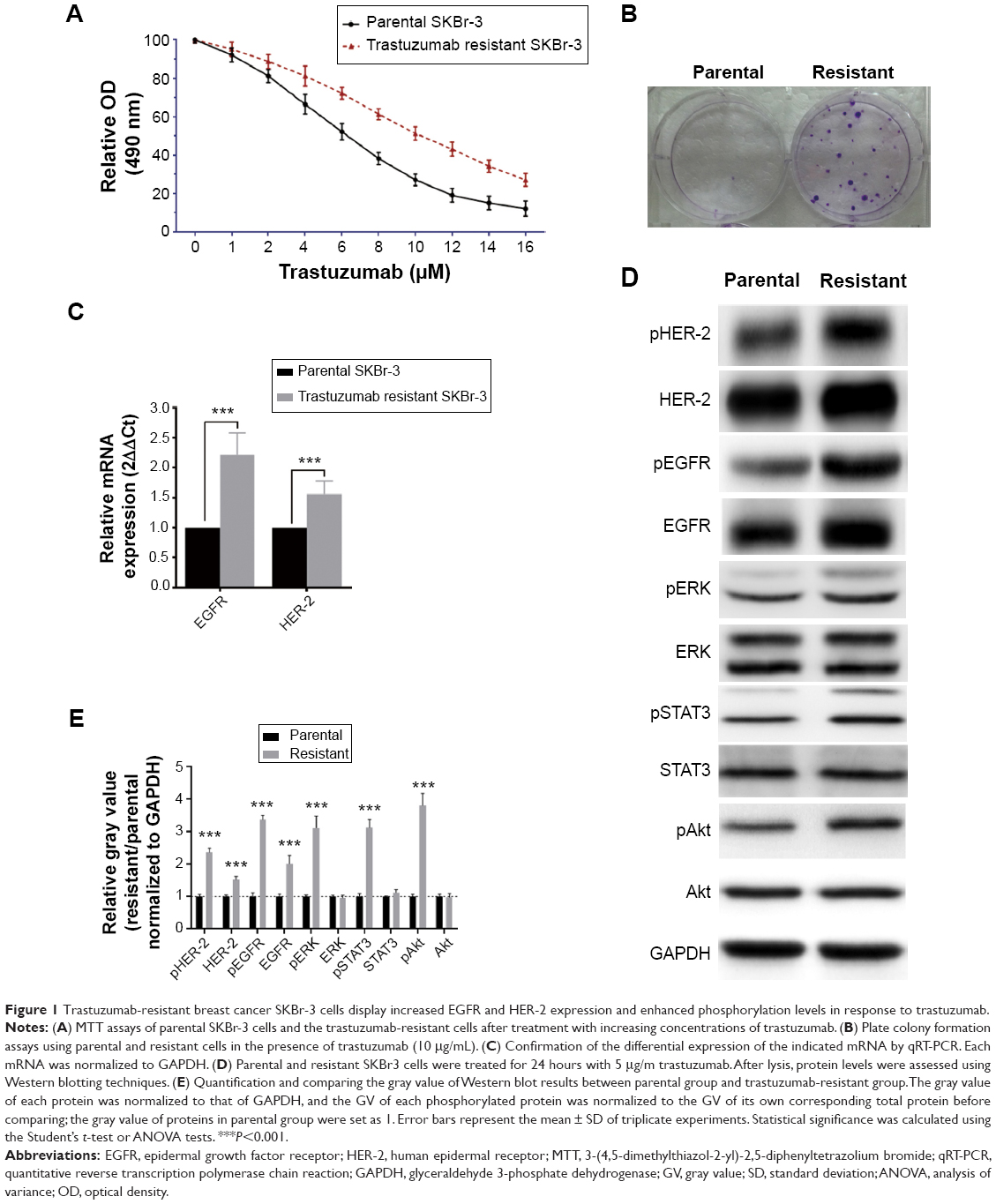 | Figure 1 Trastuzumab-resistant breast cancer SKBr-3 cells display increased EGFR and HER-2 expression and enhanced phosphorylation levels in response to trastuzumab. |
SHP-1 overexpression promotes trastuzumab sensitivity in breast cancer cells
To further investigate the role of SHP-1 in trastuzumab resistance of breast cancers, stable SHP-1 overexpressing SKbr-3 cells were established through retrovirus infection. In order to assess the role of the phosphatase activity of SHP-1 in the regulation of trastuzumab resistance, vectors expressing a catalytic domain mutant (C453S) of SHP-1 (SHP-1 mutant-type, SHP-1 MT) were established subsequently. We first treated cells with different concentrations of trastuzumab, and evaluated cell viability by using MTT assay. Results showed that, compared with cells expressing control scramble shRNA, both parent cells and resistant cells with wild-type SHP-1 overexpression displayed enhanced sensitivity to trastuzumab (Figure 2A); as expected, cells transfected with mutant-type SHP-1 showed similar trastuzumab sensitivity with controls. We also examined the apoptosis of cells after treatment with 5 μg/mL trastuzumab for 24 hours. Wild-type SHP-1 overexpression caused a significant increase in apoptosis of both parental cells and trastuzumab-resistant cells (Figure 2B). Notably, trastuzumab-resistant cells with overexpression of SHP-1 displayed a parallel trastuzumab sensitivity compared with SHP-1 overexpression parental cells. In addition, overexpression of wild-type SHP-1 also significantly suppressed in vitro colony formation by both the parental and trastuzumab-resistant cells (Figure 2C). At last, we investigated the effect of SHP-1 overexpression on the expression levels and activity of EGFR and HER-2 proteins through Western blot analysis. We found that the protein levels of EGFR and HER-2, as well as their phosphorylation levels, were significantly decreased in cells stably expressing wild-type SHP-1 compared with controls (Figure 2D and E).
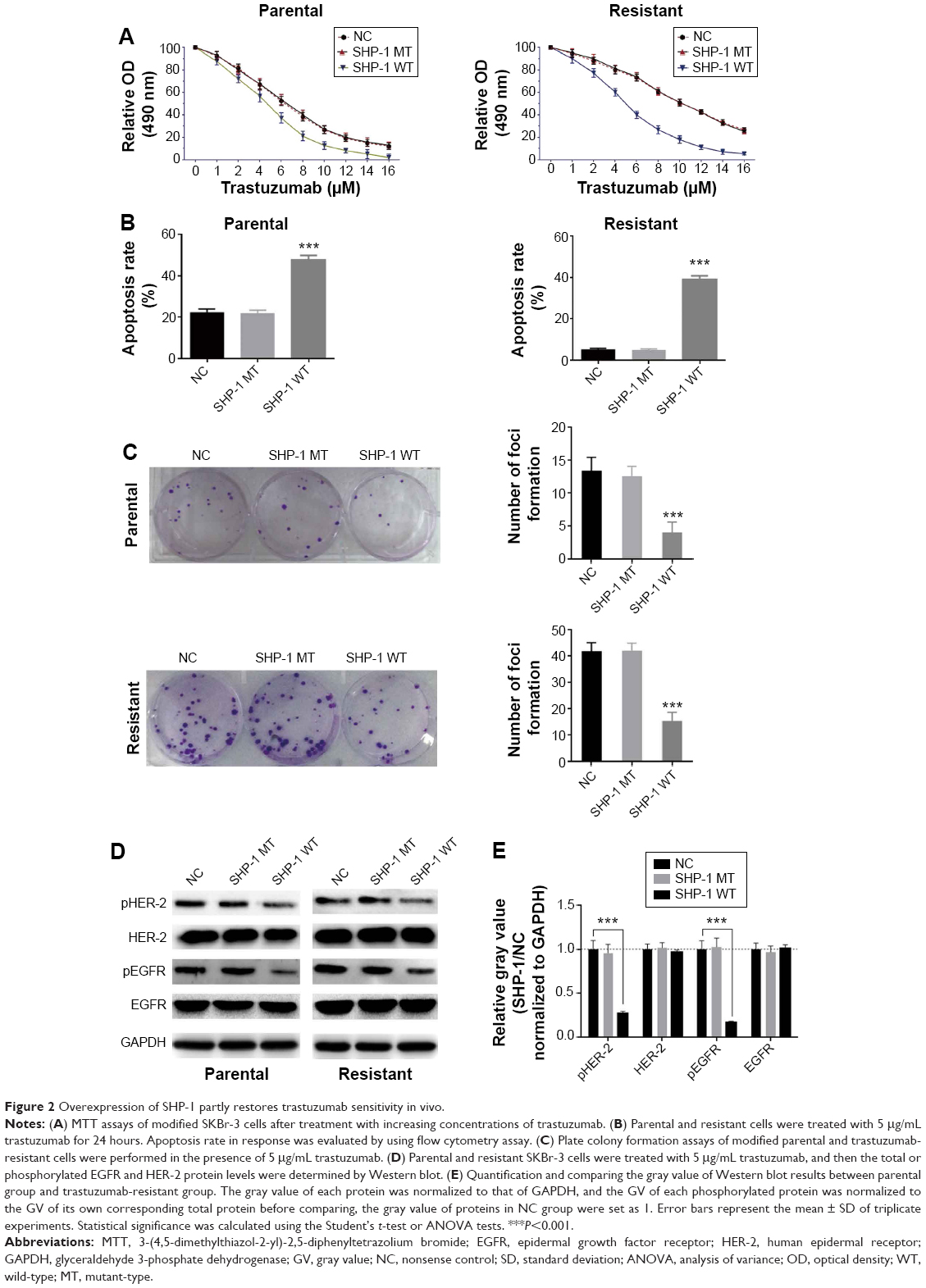 | Figure 2 Overexpression of SHP-1 partly restores trastuzumab sensitivity in vivo. |
SHP-1 binds to EGFR and HER-2 proteins
To verify the interactions between SHP-1 and EGFR, as well as HER-2, we conducted immunoprecipitation studies in order to ascertain whether SHP-1 and EGFR could be coprecipitated – an indicator of their co-association or heterodimerization. Anti-EGFR and anti-HER-2 antibodies were used to detect EGFR and HER-2 in anti-SHP-1 immunoprecipitates, respectively. Pull-down of SHP-1 with an anti-SHP-1 antibody showed co-association with both EGFR and HER-2 (Figure 3A and B). Meanwhile, we also detected the interaction between SHP-1 and phosphorylated EGFR (pEGFR) or pHER-2 in anti-SHP-1 immunoprecipitates. As expected, the corresponding bands were observed (Figure 3C and D). Notably, both pEGFR and pHER-2 protein levels in the wild type SHP-1 immunoprecipitates were less than those of C453S SHP-1. Besides, we also examined whether SHP-1 could interact with HER-3 and HER-4. We did not detect any specific band by using anti-HER-3 (Figure 3E) and anti-HER-4 antibodies (Figure 3F). The interaction between SHP-1 and both EGFR and HER-2 was further investigated by using BiFC. Cells were cotransfected with pBiFC-VC155-EGFR vectors and pBiFC-VN173-SHP1 vectors, and green fluorescence was detected (Figure 3G), which indicated protein interactions between EGFR and SHP-1. Fluorescent was also detected in cells cotransfected with pBiFC-VC155-HER2 vectors and pBiFC-VN173-SHP1 vectors (Figure 3E). However, we did not detect fluorescence in cells cotransfected with pBiFC-VN173-SHP1 and either pBiFC-VC155-HER3 or pBiFC-VC155-HER4 (Figure 3G).
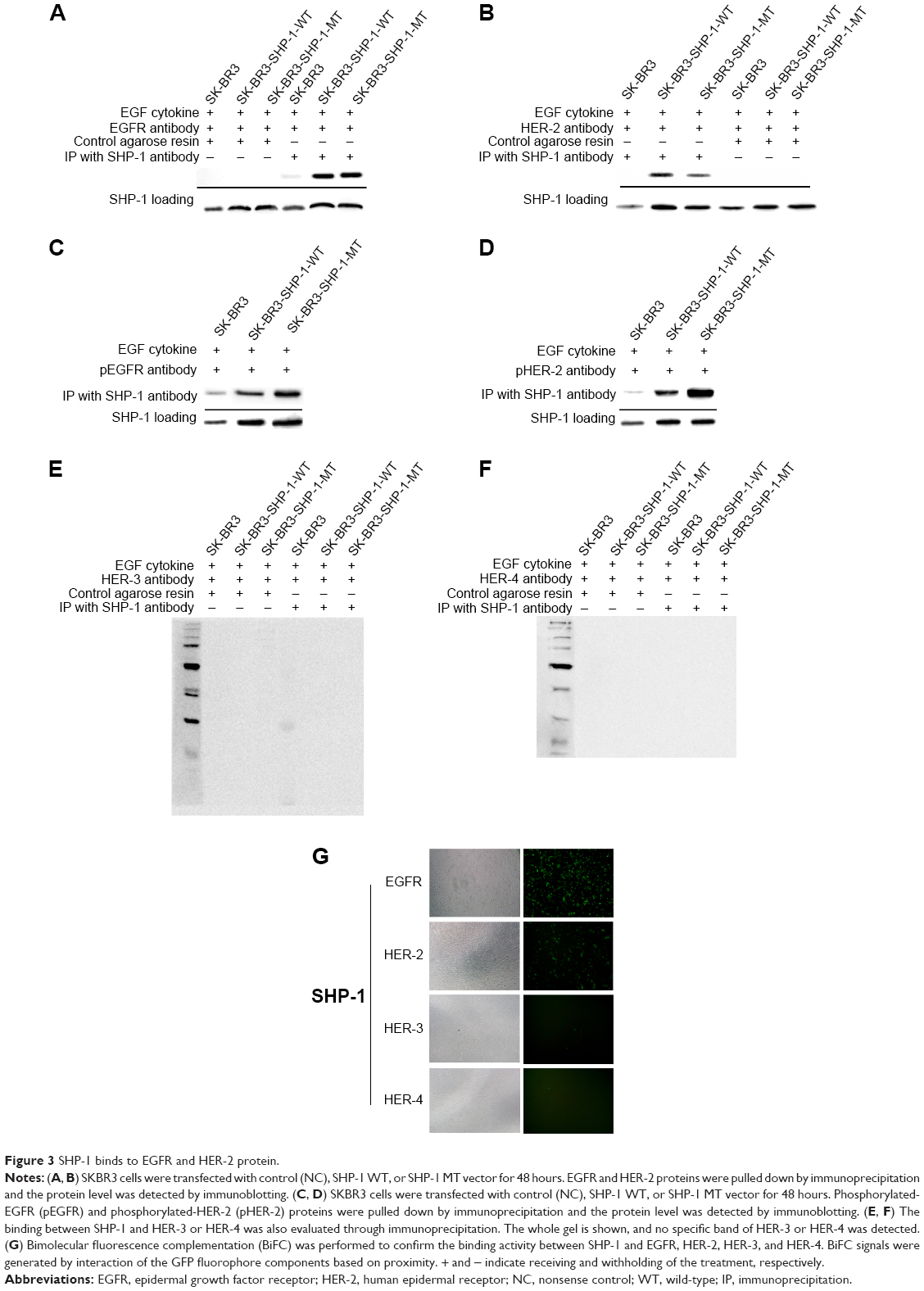 | Figure 3 SHP-1 binds to EGFR and HER-2 protein. |
SHP-1 promotes trastuzumab sensitivity in vivo
To further investigate the effect of SHP-1 on the trastuzumab resistance in vivo, xenograft models were generated using trastuzumab-resistant SKBr-3 cells modified to overexpress SHP-1 or control vectors. Compared with mice bearing tumors derived from SKBr-3 cells expressing control vectors, mice inoculated with SHP-1-modified SKBr-3 cells displayed significantly suppressed tumor development and growth (Figure 4A–C). Besides, we also evaluated the effect of SHP-1 overexpression on the survival of mice with tumor burden. A Kaplan–Meier survival analysis showed a better overall survival in mice bearing tumors developed from cells overexpressing SHP-1, compared with those injected with the control cells after treatment with trastuzumab (P=0.0036, Figure 4D). These data suggest that the overexpression of SHP-1 may sensitize trastuzumab-resistant breast cancers to trastuzumab in vivo.
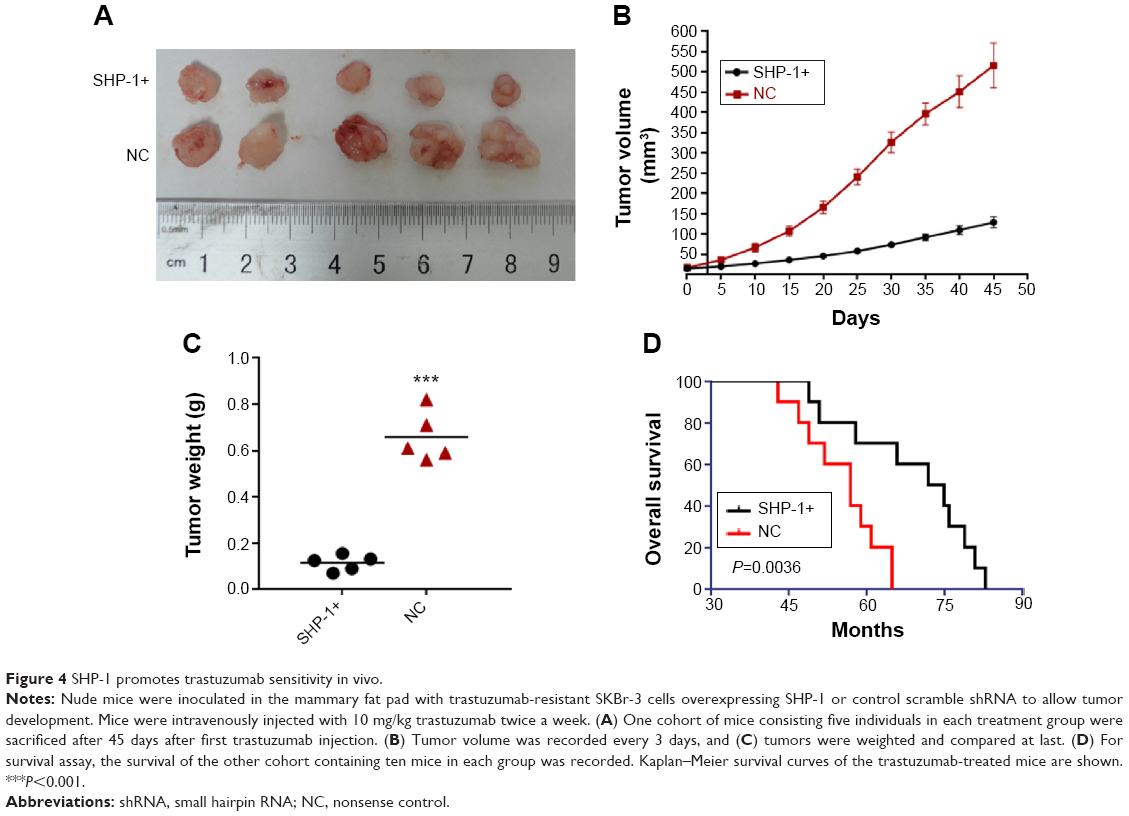 | Figure 4 SHP-1 promotes trastuzumab sensitivity in vivo. |
Discussion
Over the past decade, the humanized monoclonal erbB2/HER-2 antibody trastuzumab (Herceptin) has been successfully used for clinical treatment of patients with HER-2-positive breast cancers. Nevertheless, primary or acquired resistance to this antitumor antibody has become the major obstacle to its clinical efficacy.25 Here, we provided insight into the role of SHP-1 in the treatment of trastuzumab-resistant breast cancer. We showed that increased expression of SHP-1, a phosphatase, induces dephosphorylation of both EGFR and HER-2 and contributes to trastuzumab sensitivity of HER-2-positive, trastuzumab-resistant breast cancer cells.
Human EGFR 1 is a key member of the HER signaling pathway that may be associated with trastuzumab resistance. In earlier research, Ritter et al26 report an association between active involvement of EGFR initiated signal pathways and trastuzumab resistance; they determined that EGFR activation is causally connected to HER-2 phosphorylation, and the trastuzumab resistance cells exhibits higher levels of p-EGFR and EGFR/HER-2 heterodimers. They draw the conclusion that EGFR activation is a potential significant mechanism for sustaining the proliferation of trastuzumab-resistant breast cancer cells which continue to proliferate even in the presence of trastuzumab. Several previous cell-line model studies have also reported that overexpression and activation of EGFR induces trastuzumab resistance in HER-2-positive cell lines.26–28 Most recently, a randomized trial (NCCTG N9831) showed that high expression of EGFR appears to be associated with decreased benefit from adjuvant concurrent trastuzumab.29 These findings are supported by evidence showing that such trastuzumab-resistant breast cancer cells showed sensitivity in the presence of EGFR tyrosine kinase inhibitor.30–32 Ritter et al26 also reported that phosphorylation of HER-2 was inhibited by EGFR-specific concentrations of the EGFR tyrosine kinase inhibitors. The mentioned earlier findings emphasize the strategy of inhibiting EGFR activation in the treatment of trastuzumab-resistant cases.
The Src-homology PTP SHP-1 is a non-receptor phosphatase that contains two SH2 domains at its N-terminus, a single catalytic domain, and a C-terminal tail that contains tyrosine residues.33 SHP-1 has been suggested to contribute to the termination of mitogenic signals of growth factors by dephosphorylating critical phosphorylated molecules.34 SHP-1 has also been shown to be associated with dephosphorylation of cell membrane receptors such as platelet-derived growth factor receptor35 and the interleukin-3 receptor36 and/or other downstream substrates such as JAK/STAT3.37,38 Many observations suggest that SHP-1 is associated with proliferation of hematopoietic malignancies and epithelium-derived carcinomas.39–42 In breast cancer, Wu et al10 have reported that introduction of SHP-1 into breast cancer cells with low endogenous expression levels results in decreased cell proliferation. Several studies have demonstrated that enhanced SHP-1 expression inhibits STAT3 phosphorylation in breast cancer cells.43,44 In this study, we validated the effect of SHP-1 on the activation of EGFR and HER-2 in breast cancer cells for the first time. We observed an obvious inhibition effect of SHP-1 on EGFR and HER-2 phosphorylation, and found that introduction of SHP-1 to trastuzumab-resistant SKBr-3 cells dramatically promoted trastuzumab sensitivity. Also HER-3 and HER-4, the other members of ErbB family, may also mediate the acquirement of resistance to trastuzumab in breast cancer; we did not observe a combination between SHP-1 and neither HER-3 nor HER-4, which suggested SHP-1 mainly affected ErbB1 and ErbB2 among proteins of this family.
Besides the direct effect of SHP-1 expression level on cell proliferation and apoptosis, SHP-1 seems to be a target of some antitumor drugs. For example, Liu et al reported that SC-2001, a novel obatoclax derivative, inhibited STAT3 phosphorylation through enhancing SHP-1 expression and induces apoptosis in human breast cancer cells.44 Regorafenib, an inhibitor of multiple protein kinases, dramatically downregulated p-STAT3 expression and resulted in significant suppression of growth in wild-type SHP-1-expressing colorectal cancer cells through enhancing the phosphatase activity of SHP-1.45 Other drugs, such as dovitinib, sorafenib derivatives, γ-tocotrienol, and Capillarisin all showed a SHP-1-mediated antitumor activity.46–49 Our findings may provide potential association between the antitumor mechanism of these drugs and SHP-1 mediated EGFR inhibition. In addition, it is reasonable to speculate that drugs displaying effective upregulation and/or activation effect on SHP-1 might have potential antitumor effect in trastuzumab-resistant breast cancer cells.
Conclusion
To our knowledge, this is the first study to reveal the role of SHP-1 in EGFR/HER-2 positive trastuzumab-resistant breast cancer cells. We discovered that the antitumor activity of trastuzumab was significantly triggered by introduction of SHP-1, accompanied with dephosphorylation of both EGFR and HER-2. SHP-1 may be a useful target for treatment of EGFR/HER-2 positive trastuzumab-resistant breast cancer.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (NSFC) (Grant number 81272546), and the Specialized Research Fund for the Doctoral Program of Higher Education (SRFDP) (Grant number 20124433120009). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Authors’ contributions
RC Luo conceived of and carried out experiments, analyzed and interpreted data, and drafted the manuscript; YF Wu and R Li conceived of experiments, analyzed, and interpreted data, and wrote the manuscript; JY Zhang, G Wang, B Liu, XF Huang, and T Zhang analyzed and interpreted data. All authors read, revised, and approved the final manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
Nahleh ZA. Hormonal therapy for male breast cancer: a different approach for a different disease. Cancer Treat Rev. 2006;32: 101–105. | ||
Malvezzi M, Bertuccio P, Levi F, La Vecchia C, Negri E. European cancer mortality predictions for the year 2013. Ann Oncol. 2013;24: 792–800. | ||
Yarden Y, Sliwkowski MX. Untangling the ErbB signalling network. Nat Rev Mol Cell Biol. 2001;2:127–137. | ||
Slamon DJ, Godolphin W, Jones LA, et al. Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science. 1989;244: 707–712. | ||
Slamon DJ, Clark GM, Wong SG, Levin WJ, Ullrich A, McGuire WL. Human breast cancer: correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science. 1987;235:177–182. | ||
Hynes NE, Lane HA. ERBB receptors and cancer: the complexity of targeted inhibitors. Nat Rev Cancer. 2005;5:341–354. | ||
Spector NL, Blackwell KL. Understanding the mechanisms behind trastuzumab therapy for human epidermal growth factor receptor 2-positive breast cancer. J Clin Oncol. 2009;27:5838–5847. | ||
Vogel CL, Cobleigh MA, Tripathy D, et al. Efficacy and safety of trastuzumab as a single agent in first-line treatment of HER2-overexpressing metastatic breast cancer. J Clin Oncol. 2002;20:719–726. | ||
Huang Y, Fu P, Fan W. Novel targeted therapies to overcome trastuzumab resistance in HER2-overexpressing metastatic breast cancer. Curr Drug Targets. 2013;14:889–898. | ||
Wu C, Guan Q, Wang Y, Zhao ZJ, Zhou GW. SHP-1 suppresses cancer cell growth by promoting degradation of JAK kinases. J Cell Biochem. 2003;90:1026–1037. | ||
Fox SB, Smith K, Hollyer J, Greenall M, Hastrich D, Harris AL. The epidermal growth factor receptor as a prognostic marker: results of 370 patients and review of 3009 patients. Breast Cancer Res Treat. 1994;29:41–49. | ||
Tsutsui S, Ohno S, Murakami S, Kataoka A, Kinoshita J, Hachitanda Y. Prognostic value of the combination of epidermal growth factor receptor and c-erbB-2 in breast cancer. Surgery. 2003;133:219–221. | ||
Diermeier S, Horvath G, Knuechel-Clarke R, Hofstaedter F, Szollosi J, Brockhoff G. Epidermal growth factor receptor coexpression modulates susceptibility to Herceptin in HER2/neu overexpressing breast cancer cells via specific erbB-receptor interaction and activation. Exp Cell Res. 2005;304:604–619. | ||
Hendriks BS, Opresko LK, Wiley HS, Lauffenburger D. Coregulation of epidermal growth factor receptor/human epidermal growth factor receptor 2 (HER2) levels and locations: quantitative analysis of HER2 overexpression effects. Cancer Res. 2003;63:1130–1137. | ||
O’Donovan N, Byrne AT, O’Connor AE, McGee S, Gallagher WM, Crown J. Synergistic interaction between trastuzumab and EGFR/HER-2 tyrosine kinase inhibitors in HER-2 positive breast cancer cells. Invest New Drug. 2011;29:752–759. | ||
Paulson RF, Vesely S, Siminovitch KA, Bernstein A. Signalling by the W/Kit receptor tyrosine kinase is negatively regulated in vivo by the protein tyrosine phosphatase Shp1. Nat Genet. 1996;13:309–315. | ||
Tautz L, Critton DA, Grotegut S. Protein tyrosine phosphatases: structure, function, and implication in human disease. Methods Mol Biol. 2013;1053:179–221. | ||
Nunes-Xavier CE, Martin-Perez J, Elson A, Pulido R. Protein tyrosine phosphatases as novel targets in breast cancer therapy. Biochim Biophys Acta. 2013;1836:211–226. | ||
Muller T, Choidas A, Reichmann E, Ullrich A. Phosphorylation and free pool of beta-catenin are regulated by tyrosine kinases and tyrosine phosphatases during epithelial cell migration. J Biol Chem. 1999;274:10173–10183. | ||
Forbes K, Skinner L, Aplin JD, Westwood M. The tyrosine phosphatase SHP-1 negatively regulates cytotrophoblast proliferation in first-trimester human placenta by modulating EGFR activation. Cell Mol Life Sci. 2012. | ||
Witkiewicz A, Raghunath P, Wasik A, et al. Loss of SHP-1 tyrosine phosphatase expression correlates with the advanced stages of cutaneous T-cell lymphoma. Hum Pathol. 2007;38:462–467. | ||
Oka T, Yoshino T, Hayashi K, et al. Reduction of hematopoietic cell-specific tyrosine phosphatase SHP-1 gene expression in natural killer cell lymphoma and various types of lymphomas/leukemias: combination analysis with cDNA expression array and tissue microarray. Am J Pathol. 2001;159:1495–1505. | ||
Tassidis H, Brokken LJ, Jirstrom K, et al. Immunohistochemical detection of tyrosine phosphatase SHP-1 predicts outcome after radical prostatectomy for localized prostate cancer. Int J Cancer. 2010;126:2296–2307. | ||
Gong C, Yao Y, Wang Y, et al. Up-regulation of miR-21 mediates resistance to trastuzumab therapy for breast cancer. J Biol Chem. 2011;286:19127–19137. | ||
De P, Hasmann M, Leyland-Jones B. Molecular determinants of trastuzumab efficacy: what is their clinical relevance? Cancer Treat Rev. 2013;39:925–934. | ||
Ritter CA, Perez-Torres M, Rinehart C, et al. Human breast cancer cells selected for resistance to trastuzumab in vivo overexpress epidermal growth factor receptor and ErbB ligands and remain dependent on the ErbB receptor network. Clin Cancer Res. 2007;13:4909–4919. | ||
Kumar R. ErbB-dependent signaling as a determinant of trastuzumab resistance. Clin Cancer Res. 2007;13:4657–4659. | ||
Dua R, Zhang J, Nhonthachit P, Penuel E, Petropoulos C, Parry G. EGFR over-expression and activation in high HER2, ER negative breast cancer cell line induces trastuzumab resistance. Breast Cancer Res Treat. 2010;122:685–697. | ||
Cheng H, Ballman K, Vassilakopoulou M, et al. EGFR expression is associated with decreased benefit from trastuzumab in the NCCTG N9831 (Alliance) trial. Br J Cancer. 2014;111:1065–1071. | ||
Moulder SL, Yakes FM, Muthuswamy SK, Bianco R, Simpson JF, Arteaga CL. Epidermal growth factor receptor (HER1) tyrosine kinase inhibitor ZD1839 (Iressa) inhibits HER2/neu (erbB2)-overexpressing breast cancer cells in vitro and in vivo. Cancer Res. 2001;61:8887–8895. | ||
Baselga J, Rischin D, Ranson M, et al. Phase I safety, pharmacokinetic, and pharmacodynamic trial of ZD1839, a selective oral epidermal growth factor receptor tyrosine kinase inhibitor, in patients with five selected solid tumor types. J Clin Oncol. 2002;20:4292–4302. | ||
Xia W, Gerard CM, Liu L, Baudson NM, Ory TL, Spector NL. Combining lapatinib (GW572016), a small molecule inhibitor of ErbB1 and ErbB2 tyrosine kinases, with therapeutic anti-ErbB2 antibodies enhances apoptosis of ErbB2-overexpressing breast cancer cells. Oncogene. 2005;24:6213–6221. | ||
Neel BG, Gu H, Pao L. The ‘Shp’ing news: SH2 domain-containing tyrosine phosphatases in cell signaling. Trends Biochem Sci. 2003;28:284–293. | ||
Tsui FW, Martin A, Wang J, Tsui HW. Investigations into the regulation and function of the SH2 domain-containing protein-tyrosine phosphatase, SHP-1. Immunol Res. 2006;35:127–136. | ||
Yu Z, Su L, Hoglinger O, Jaramillo ML, Banville D, Shen SH. SHP-1 associates with both platelet-derived growth factor receptor and the p85 subunit of phosphatidylinositol 3-kinase. J Biol Chem. 1998;273:3687–3694. | ||
Ayrault O, Andrique L, Fauvin D, Eymin B, Gazzeri S, Seite P. Human tumor suppressor p14ARF negatively regulates rRNA transcription and inhibits UBF1 transcription factor phosphorylation. Oncogene. 2006;25:7577–7586. | ||
Valentino L, Pierre J. JAK/STAT signal transduction: regulators and implication in hematological malignancies. Biochem Pharmacol. 2006;71:713–721. | ||
Jung JH, Kwon TR, Jeong SJ, et al. Apoptosis induced by tanshinone IIA and cryptotanshinone is mediated by distinct JAK/STAT3/5 and SHP1/2 signaling in chronic myeloid leukemia K562 cells. J Evid Based Complementary Altern Med. 2013;2013:805639. | ||
Zapata PD, Ropero RM, Valencia AM, et al. Autocrine regulation of human prostate carcinoma cell proliferation by somatostatin through the modulation of the SH2 domain containing protein tyrosine phosphatase (SHP)-1. J Clin Endocrinol Metab. 2002;87:915–926. | ||
Wu C, Sun M, Liu L, Zhou GW. The function of the protein tyrosine phosphatase SHP-1 in cancer. Gene. 2003;306:1–12. | ||
Tao XH, Shen JG, Pan WL. Significance of SHP-1 and SHP-2 expression in human papillomavirus infected Condyloma acuminatum and cervical cancer. Pathol Oncol Res. 2008;14:365–371. | ||
Vinayanuwattikun C, Chantranuwat P, Sriuranpong V, Mutirangura A. Erratum to: The role of SHP-1 promoter 2 hypermethylation detection of lymph node micrometastasis in resectable stage I non-small cell lung cancer as a prognostic marker of disease recurrence. Int J Clin Onco. 2014;19:593. | ||
Liu CY, Tseng LM, Su JC. Novel sorafenib analogues induce apoptosis through SHP-1 dependent STAT3 inactivation in human breast cancer cells. Breast Cancer Res Treat. 2013;15:R63. | ||
Liu CY, Su JC, Ni MH, et al. Obatoclax analog SC-2001 inhibits STAT3 phosphorylation through enhancing SHP-1 expression and induces apoptosis in human breast cancer cells. Breast Cancer Res Treat. 2014;146:71–84. | ||
Fan LC, Teng HW, Shiau CW, et al. SHP-1 is a target of regorafenib in colorectal cancer. Oncotarget. 2014;5:6243–6251. | ||
Lee JH, Chiang SY, Nam D, et al. Capillarisin inhibits constitutive and inducible STAT3 activation through induction of SHP-1 and SHP-2 tyrosine phosphatases. Cancer Lett. 2014;345:140–148. | ||
Huang CY, Tai WT, Hsieh CY, et al. A sorafenib derivative and novel SHP-1 agonist, SC-59, acts synergistically with radiotherapy in hepatocellular carcinoma cells through inhibition of STAT3. Cancer Lett. 2014;349:136–143. | ||
Chen KF, Tai WT, Hsu CY, et al. Blockade of STAT3 activation by sorafenib derivatives through enhancing SHP-1 phosphatase activity. Eur J Med Chem. 2012;55:220–227. | ||
Tai WT, Cheng AL, Shiau CW, et al. Dovitinib induces apoptosis and overcomes sorafenib resistance in hepatocellular carcinoma through SHP-1-mediated inhibition of STAT3. Mol Cancer Ther. 2012;11:452–463. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.