")
Back to Journals » OncoTargets and Therapy » Volume 14
Propofol Inhibits the Progression of Cervical Cancer by Regulating HOTAIR/miR-129-5p/RPL14 Axis
Authors Sun N, Zhang W, Liu J, Yang X, Chu Q
Received 1 September 2020
Accepted for publication 10 November 2020
Published 19 January 2021 Volume 2021:14 Pages 551—564
DOI https://doi.org/10.2147/OTT.S279942
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr William C. Cho
Nai Sun,* Wei Zhang,* Jiaying Liu, Xiaochen Yang, Qinjun Chu
Department of Anesthesiology, Zhengzhou Central Hospital Affiliated to Zhengzhou University, Zhengzhou City, Henan Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qinjun Chu
Department of Anesthesiology, Zhengzhou Central Hospital Affiliated to Zhengzhou University, No. 16 Tongbai North Road, Zhongyuan District, Zhengzhou 450007, Henan Province, People’s Republic of China
Tel/Fax +86 371-67690273
Email [email protected]
Background: Propofol has been proposed to function as a tumor suppressor in various human cancers. In this study, we aimed to investigate the anti-tumor effect of propofol on cervical cancer (CC).
Methods: Cell Counting Kit-8 (CCK-8) assay, colony formation assay, flow cytometry analysis, transwell assay and wound healing assay were conducted for cell viability, colony formation, apoptosis, invasion and migration, respectively. Western blot assay was used for protein levels. Quantitative real-time polymerase chain reaction (qRT-PCR) was used for HOX antisense intergenic RNA (HOTAIR), miR-129-5p and RPL14 levels. Dual-luciferase reporter assay and RNA immunoprecipitation (RIP) assay were executed to verify the interaction between miR-129-5p and HOTAIR or RPL14. Murine xenograft model assay was used for the role of propofol in tumor progression in vivo.
Results: Propofol treatment suppressed CC cell viability, colony formation, invasion and migration and facilitated apoptosis. Propofol treatment led to a marked reduction in HOTAIR level in CC cells. HOTAIR overexpression promoted cell colony formation, invasion and migration and repressed apoptosis in CC cells and propofol-treated CC cells. For mechanism analysis, HOTAIR positively regulated RPL14 expression via acting as the sponge of miR-129-5p. MiR-129-5p overexpression reversed the impacts of HOTAIR on the malignant behaviors of propofol-treated CC cells. Furthermore, miR-129-5p inhibition accelerated the progression of CC cells, while RPL14 interference rescued the effect. In addition, propofol treatment restrained tumor growth of CC in vivo.
Conclusion: Propofol inhibited CC development by modulation of HOTAIR/miR-129-5p/RPL14 axis.
Keywords: propofol, cervical cancer, HOTAIR, miR-129-5p, RPL14
Introduction
Cervical cancer (CC) is a common gynecological malignancy in the world, ranking fourth in the leading cause of cancer-associated deaths in female.1,2 Although the therapeutic methods, including radiotherapy, chemotherapy and surgery, have made great advancements, the overall survival rate of CC patients remains unfavorable because of the metastasis and recurrence of tumors.3–5 Therefore, it is urgently needed to clarify the underlying mechanism of CC and discover novel targets for CC therapy.
Propofol is an intravenous anesthetic used for the induction and maintenance of general anesthesia during surgery.6 Apart from its anesthetic effect, propofol also has been demonstrated to play an anti-tumor effect on diverse human cancers, such as osteosarcoma,7 hepatocellular carcinoma8 and pancreatic cancer.9 Moreover, it has been documented that propofol can inhibit tumor progression through regulating long non-coding RNAs (lncRNAs), which are identified as a sort of non-coding RNAs (ncRNAs) with >200 nucleotides (nts).10 For example, Chen et al claimed that propofol suppressed the malignancy of papillary thyroid cancer cells by reducing ANRIL.11 Ren and Zhang suggested that propofol contributed to the apoptosis and restrained the proliferation of colorectal cancer cells through decreasing HOXA11-AS and increasing miR-let-7i.12 In CC, the oncogenic lncRNA HOX antisense intergenic RNA (HOTAIR) level has been confirmed to be abnormally expressed and aggravated CC carcinogenesis.13 Moreover, Zhang et al disclosed that propofol hampered CC development by repressing mTOR/p70S6K pathway-mediated by HOTAIR.14 Even so, the relationship between propofol and HOTAIR in CC progression and related mechanisms still need to be further investigated.
MicroRNAs (miRNAs) are a class of highly conserved ncRNAs with ~22 nts which can bind to the 3ʹ untranslated regions (3ʹUTR) of target genes to modulate gene expression.15,16 Emerging evidence has shown that miRNAs serve as essential regulators in CC. For instance, miR-342-3p blocked CC cell growth and metastasis through recognizing FOXM1.17 MiR-486-3p directly targeted ECM1 to repress CC cell proliferation and motility.18 As for miR-129-5p, Wang et al demonstrated its anti-tumor activity in CC.19 Ribosomal protein L14 (RPL14) has been proved to participate in regulating the malignant phenotypes of CC cells via acting as the target gene of miR-1285-3p.20 Nevertheless, there is no report on RPL14 as a target of miR-129-5p.
The study aimed to reveal the effects of propofol on CC carcinogenesis. Moreover, whether HOTAIR took part in the regulation of CC progression and its related molecular mechanisms were investigated.
Materials and Methods
Cell Culture
Two CC cell lines (HeLa and SiHa) and 293T cell line were purchased from Procell (Wuhan, China) and cultivated in Dulbecco’s modified Eagle’s medium (DMEM; Procell) added with 10% fetal bovine serum (FBS; Procell) and 1% penicillin/streptomycin (Procell) at 37°C in a humid atmosphere containing 5% CO2.
Cell Transfection
The overexpression vector of HOTAIR (oe-HOTAIR) and its control (vector), miR-129-5p mimic and mimic NC, miR-129-5p inhibitor and inhibitor NC, small interfering RNA targeting RPL14 (si-RPL14) and scramble siRNA (si-NC) were all synthesized by GenePharma (Shanghai, China). Then, HeLa and SiHa cells were plated into 12-well plates and transfected with indicated vectors or oligonucleotides utilizing Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) based on the manufacturers’ protocols.
Propofol Treatment
Propofol (Sigma-Aldrich, St. Louis, MO, USA) was dissolved in DMSO (Sigma-Aldrich) and made up with the culture medium until the final dose of Dimethyl sulfoxide (DMSO; Sigma-Aldrich) <0.1%. Then, HeLa and SiHa cells were treated with 1 µg/mL, 5µg/mL, 10 µg/mL and 20 µg/mL propofol (Sigma-Aldrich) for 48 h. The control groups (0 µg/mL propofol) were treated with the same volume of DMSO (Sigma-Aldrich) for 48 h.
Cell Counting Kit-8 (CCK-8) Assay
The viability of HeLa and SiHa cells were tested by CCK-8 assay. Briefly, CC cells (1×104 cells/well) were seeded into 96-well plates and treated with various doses of propofol (Sigma-Aldrich). After incubation for 48 h, 20 μL CCK-8 (Sigma-Aldrich) was added into each well with a further incubation for 2 h at 37°C. The absorption at 450 nm was measured by a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
Colony Formation Assay
Following relevant transfection and treatment, the colony formation capacity of HeLa and SiHa cells was tested through colony formation assay. Briefly, the cells (250 cells/well) were sowed into the 6-well plates and maintained in the culture medium for about 14 days. After that, the colonies were fixed with 4% paraformaldehyde (Sangon, Shanghai, China) and then mixed with 0.1% crystal violet (Solarbio, Beijing, China). After 15 min, the colonies contained more than 50 cells were counted with a microscope (Olympus Corporation, Tokyo, Japan).
Flow Cytometry Analysis
The Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis Detection Kit (Beyotime, Shanghai, China) was adopted to analyze the apoptosis of HeLa and SiHa cells. The cells were harvested and washed twice with PBS (Solarbio) after indicated transfection and treatment. Next, the cells were resuspended and subjected to 5 μL AnnexinV-FITC and 5 μL PI for 15 min in the dark. At last, the apoptotic cells were examined utilizing a flow cytometry (BD Biosciences, San Jose, CA, USA) and analyzed with FlowJo 7.6.1.
Transwell Assay
After indicated transfection and treatment, the invasion of HeLa and SiHa cells was assessed utilizing the 24-well transwell inserts (BD Biosciences) which were coated with Matrigel (BD Biosciences). 1×104 cells were inoculated to the upper chamber and the bottom chamber was added with 600 μL complete medium as chemoattractive. After 24 h of incubation at 37°C, the invaded cells were fixed with 4% paraformaldehyde (Sangon) followed by 0.1% crystal violet dying (Solarbio). The invaded cells were counted with an inverted microscope (Olympus) at the magnification of 100×.
Wound Healing Assay
HeLa and SiHa cells (1×105 cells/well) were inoculated onto 6-well plates. When the cells were cultured 100% confluence, a pipette tip was utilized to make a scratch and then the scratch was washed three times with PBS (Solarbio). Afterward, the widths of the scratch wound were examined at 0 h and 24 h at the magnification of 100× to assess cell migration ability.
Western Blot Assay
Total protein in CC cells was extracted by RIPA (Beyotime) and determined with a BCA protein assay kit (Tiangen, Beijing, China). Equal amounts of proteins (20 μg) were resolved with sodium dodecyl sulfonate-polyacrylamide gel (Solarbio) electrophoresis and then blotted onto polyvinylidene difluoride membranes (Millipore, Billerica, MA, USA). Next, the membranes were blocked with 5% defatted milk in Tris Buffered Saline with Tween-20 (TBST; Solarbio) for 1 h at indoor temperature. Thereafter, the membranes were kept overnight with primary antibodies: matrix metalloprotein 9 (MMP9; ab38898; Abcam, Cambridge, MA, USA), proliferating cell nuclear antigen (PCNA; ab18197; Abcam), BCL2-Associated X (Bax; ab182734; Abcam), RPL14 (ab181200; Abcam) or β-actin (ab8227; Abcam) and then probed with HRP-conjugated secondary antibody (ab6789; Abcam) for 2 h at indoor temperature. At last, the protein bands were visualized with an ECL kit (Vazyme, Nanjing, China) and analyzed using ImageJ v1.8.0 (National Institutes of Health). The gray scale of the target protein = the IOD value of the target protein/the IOD value of the corresponding internal reference. Next, the target proteins were normalized.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Total RNA extraction was done through the usage of TRIzol (Invitrogen). The extracted RNAs were treated with RNase-free DNase (Promega, Madison, WI, USA). The synthesis of cDNAs was finished utilizing M-MLV Reverse Transcriptase Kit (Promega) or miRNA 1st Strand cDNA Synthesis Kit (Vazyme). After that, qRT-PCR assay was conducted on an ABI 7500 PCR system (Applied Biosystems, Foster City, CA, USA) using AceQ Universal SYBR qPCR Master Mix (Vazyme) and specific primers (GenePharma). The primers were: HOTAIR: (F: 5ʹ-GGAAAGATCCAAATGGGACCA-3ʹ and R: 5ʹ-CTAGGAATCAGCACGAAGCAAA-3ʹ); miR-129-5p: (F: 5ʹ-AGTGCGTGTCGTGGAGTC-3ʹ and R: 5ʹ-AGAGGCAGGGATGATGTTCTG-3ʹ); RPL14: (F: 5ʹ-TTGAAGCCAGAGAAAGGGTAA-3ʹ and R: 5ʹ-CCTGAAATGGGATTTGGCTA-3ʹ); β-actin: (F: 5ʹ-AGCCATGTACGTAGCCATC-3ʹ and R: 5ʹ-TCCCTCTCAGCTGTGGTGGTGAA-3ʹ); U6: (F: 5ʹ-CTCGCTTCGGCAGCACA-3ʹ and R: 5ʹ-AACGCTTCACGAATTTGCGT-3ʹ). The relative expression was computed via the 2−ΔΔCt strategy with β-actin or U6 as an internal reference.
Dual-Luciferase Reporter Assay
The sequences of HOTAIR or RPL14 3ʹUTR consisting of the predicted miR-129-5p binding sites were cloned and inserted into pmirGLO plasmid (Promega), generating HOTAIR wt and RPL14 3ʹUTR wt, respectively. The mutant sequences of HOTAIR or RPL14 3ʹUTR lacking miR-129-5p binding sites were cloned and inserted into pmirGLO plasmid (Promega) to construct HOTAIR mut and PRL14 3ʹUTR mut, respectively. Then, the constructs were administrated into 293T cells in combination with mimic NC or miR-129-5p mimic. The luciferase activity was tested with Dual-Luciferase Reporter Assay Kit (Promega) after 48 h. The Renilla luciferase activity was normalized to firefly luciferase activity.
RNA Immunoprecipitation (RIP) Assay
The Magna RIPTM RNA Binding Protein Immunoprecipitation Kit (Millipore) was employed for RIP assay. Briefly, HeLa and SiHa cells were disrupted in RIP lysis buffer and then cell lysates were maintained with magnetic beads conjugated with anti-immunoglobulin G (anti-IgG) or anti-Argonaute-2 (anti-Ago2) for 8 h at 4°C. The anti-IgG and input were used as controls. Finally, the immunoprecipitated RNAs were purified and then subjected to qRT-PCR test for the enrichment of HOTAIR, miR-129-5p and RPL14.
Murine Xenograft Model
The female nude mice were obtained from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China) and divided into 6 groups (n=6/group): Model, Propofol, Propofol+vector, Propofol+oe-HOTAIR, Propofol+miR-NC and Propofol+miR-129-5p. The Model groups and Propofol groups were transplanted with HeLa cells (1×106 cells) and intraperitoneally injected with DMSO (DMSO; Sigma-Aldrich) or propofol (35mg/kg; Sigma-Aldrich) every week for 4 continuous weeks after 10 days of HeLa cells injection. For Propofol+vector, Propofol+oe-HOTAIR, Propofol+miR-NC and Propofol+miR-129-5p groups, the mice were implanted with HeLa cells (1×106 cells) transfected with vector, oe-HOTAIR, miR-NC or miR-129-5p and intraperitoneally given propofol (35mg/kg; Sigma-Aldrich) once per week for 4 continuous weeks. Tumor volume was monitored on days 5, 10, 17, 24, 31 and 38 and computed with the formula: (Length×width2)/2. On day 38, the mice were euthanized and tumor tissues were harvested and then weighed. The harvested tissues were saved at −80°C. The animal study was performed in accordance with the Guidelines for Care and Use of Laboratory Animals of “National Institutes of Health” and approved by the Ethics Committee of Animal Research of Zhengzhou Central Hospital Affiliated to Zhengzhou University.
Statistical Analysis
The experiments were conducted in triple times. The results were analyzed by GraphPad Prism 7 and presented as mean ± standard deviation. The differences in two groups and multiple groups were estimated by Student’s t-test and one-way analysis of variance, respectively. It was deemed as a significant difference if P<0.05.
Results
Propofol Treatment Inhibited CC Cell Viability, Colony Formation, Invasion and Migration and Promoted Apoptosis
To elucidate the roles of propofol in CC, HeLa and SiHa cells were exposed to various doses of propofol (0 µg/mL, 1 µg/mL, 5 µg/mL, 10 µg/mL or 20 µg/mL) for 48 h. Then, we determined the viability of HeLa and SiHa cells using CCK-8 assay. The results showed that the viability of propofol-treated cells was apparently suppressed in a concentration-dependent way (Figure 1A). Then, we selected 10 µg/mL propofol for further experiments for the significant suppressive effect on cell viability. The results of colony formation assay indicated that the colony formation ability of HeLa and SiHa cells was markedly inhibited by propofol treatment (Figure 1B). As demonstrated by flow cytometry analysis, propofol treatment led to a remarkable promotion in the apoptosis of HeLa and SiHa cells (Figure 1C). Moreover, we demonstrated that the invasion and migration of HeLa and SiHa cells were evidently restrained following the exposure of propofol (Figure 1D and E). Additionally, we determined the protein levels of MMP9, PCNA and Bax in HeLa and SiHa cells after propofol treatment through Western blot assay. The results implied that MMP-9 and PCNA levels were reduced and Bax level was elevated in propofol-treated HeLa and SiHa cells (Figure 1F). All these data indicated that propofol treatment relieved the malignancy of CC cells.
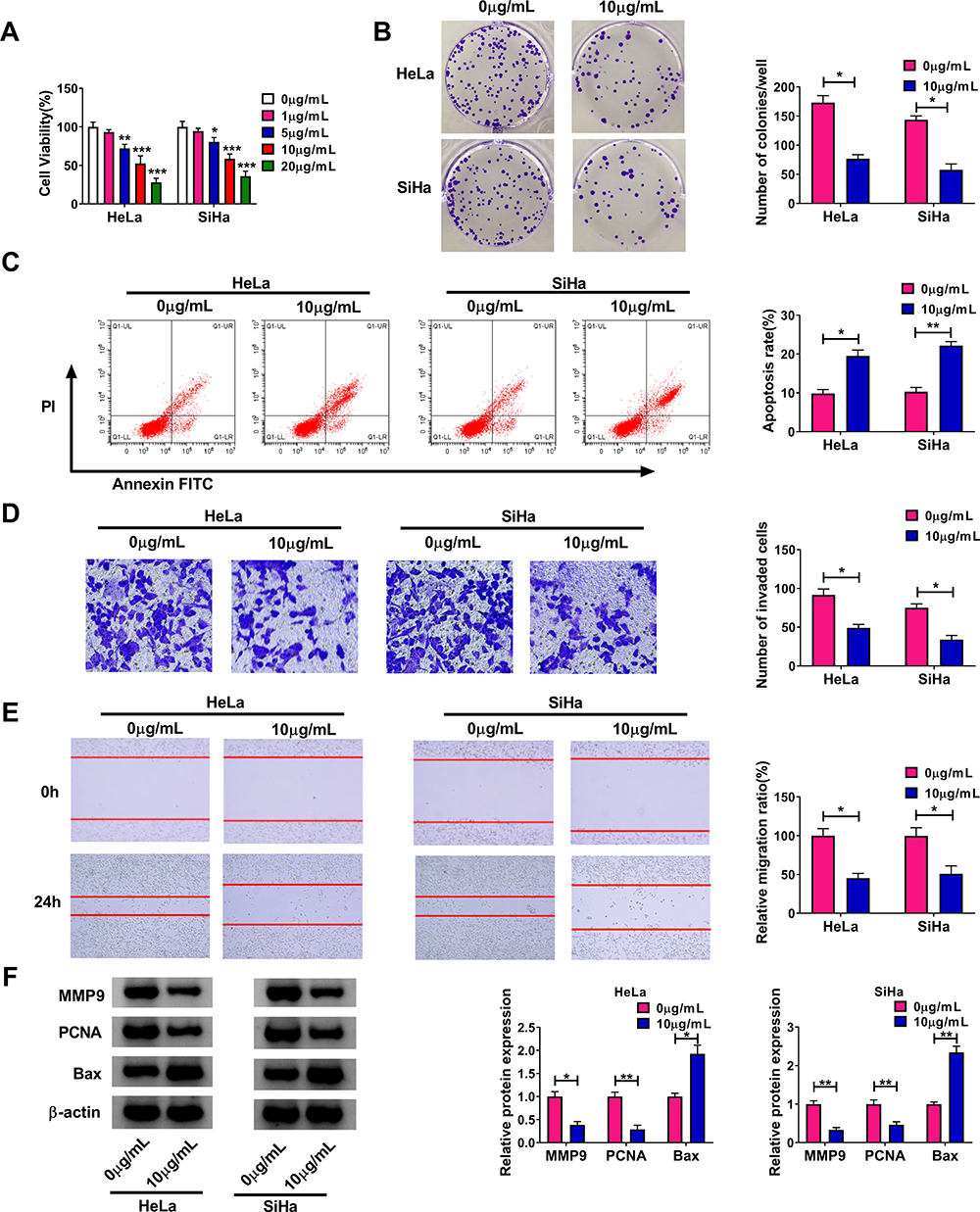 |
Figure 1 Effects of propofol treatment on CC cell viability, colony formation, apoptosis, invasion and migration. (A) After different concentrations of propofol treatment, the viability of HeLa and SiHa cells was assessed by CCK-8 assay. (B–E) The colony formation, apoptosis, invasion and migration of HeLa and SiHa cells treated with 10 µg/mL propofol were evaluated by colony formation assay, flow cytometry analysis, transwell assay and wound healing assay, respectively. (F) The protein levels of MMP9, PCNA and Bax in propofol-treated HeLa and SiHa cells were measured by Western blot assay. *P<0.05, **P<0.01, ***P<0.001. |
Overexpression of HOTAIR Promoted the Colony Formation, Invasion and Migration and Suppressed the Apoptosis of CC Cells
To explore the underlying mechanism of the inhibitory roles of propofol in CC cell progression, HeLa and SiHa cells were treated with different concentrations of propofol (0 µg/mL, 1 µg/mL, 5 µg/mL, 10 µg/mL or 20 µg/mL) for 48 h. Then, qRT-PCR assay was utilized to determine the expression level of HOTAIR in HeLa and SiHa cells. The results showed that HOTAIR level was conspicuously reduced in HeLa and SiHa cells in a dose-dependent manner (Figure 2A). Next, oe-HOTAIR or vector was transfected into HeLa and SiHa cells to investigate the functional roles of HOTAIR in CC cells. The results of qRT-PCR assay showed that oe-HOTAIR transfection distinctly enhanced the expression level of HOTAIR in HeLa and SiHa cells compared to control groups (Figure 2B). The results of colony formation assay exhibited that HOTAIR overexpression markedly enhanced the colony formation ability of HeLa and SiHa cells in both control and propofol groups (Figure 2C). Flow cytometry analysis showed that overexpression of HOTAIR drastically repressed the apoptosis of HeLa and SiHa cells and also reversed the promotion of cell apoptosis in HeLa and SiHa cells caused by propofol (Figure 2D). Overexpression of HOTAIR notably promoted cell invasion and migration in HeLa and SiHa cells treated with or without propofol, as illustrated by transwell assay and wound healing assay (Figure 2E and F). Furthermore, HOTAIR elevation prominently increased the protein levels of MMP9 and PCNA and dramatically decreased the protein level of Bax in HeLa and SiHa cells in both control and propofol groups (Figure 2G). These results indicated that HOTAIR was a regulator in propofol-mediated CC cell progression.
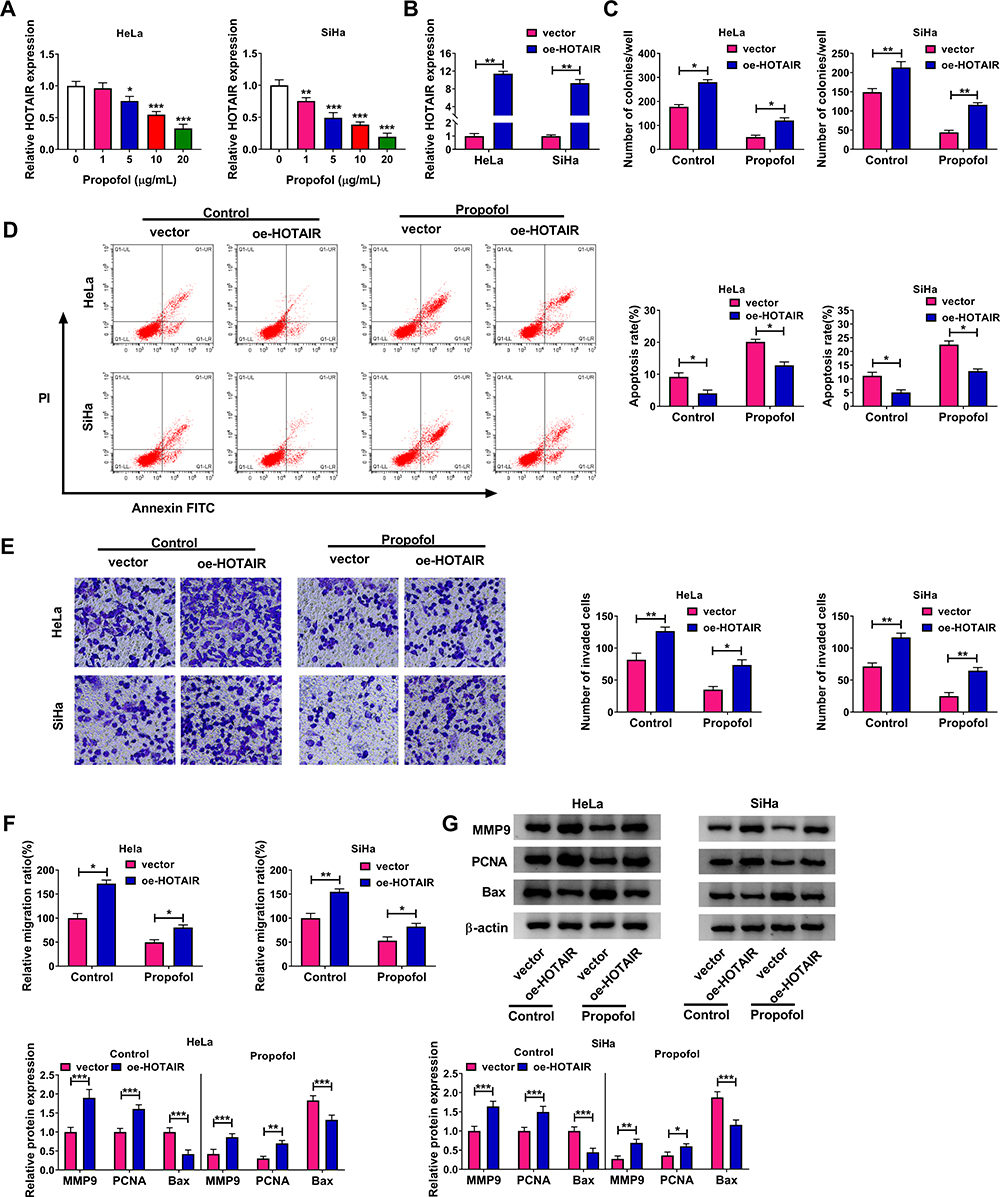 |
Figure 2 Propofol repressed CC cell progression through elevating HOTAIR expression. (A) The expression level of HOTAIR in different doses of propofol-treated HeLa and SiHa cells was examined by qRT-PCR assay. (B) The expression level of HOTAIR in HeLa and SiHa cells transfected with oe-HOTAIR or vector was detected with qRT-PCR assay. (C–G) HeLa and SiHa cells were transfected with vector or oe-HOTAIR before treatment with (Propofol group) or without propofol (Control group). (C–F) The colony formation, apoptosis, invasion and migration of HeLa and SiHa cells were assessed by colony formation assay, flow cytometry analysis, transwell assay and wound healing assay, respectively. (G) The protein levels of MMP9, PCNA and Bax in HeLa and SiHa cells were tested by Western blot assay. *P<0.05, **P<0.01, ***P<0.001. |
HOTAIR Overexpression Aggravated the Malignant Behaviors of CC Cells Upon Propofol Treatment by Sponging miR-129-5p
As shown in Figure 3A, propofol exposure obviously increased the expression level of miR-129-5p in HeLa and SiHa cells in a dose-independent way. Through analyzing online tool miRcode (http://mircode.org/?gene=HOTAIR&mirfam=&class=&cons=&trregion=), miR-129-5p was found to contain the potential binding sites of HOTAIR (Figure 3B). Then, dual-luciferase reporter assay and RIP assay were conducted to verify the interaction between HOTAIR and miR-129-5p. Dual-luciferase reporter assay presented that miR-129-5p mimic transfection strikingly repressed the luciferase activity of HOTAIR wt in 293T cells, but did not influence the luciferase activity of HOTAIR mut (Figure 3C). Moreover, RIP assay indicated that HOTAIR and miR-129-5p were all enriched by anti-Ago2 immunoprecipitate in HeLa and SiHa cells compared to anti-IgG immunoprecipitate (Figure 3D). Next, HeLa and SiHa cells were transfected with vector, oe-HOTAIR, oe-HOTAIR+mimic NC or oe-HOTAIR+miR-129-5p mimic and then treated with 10 µg/mL propofol to explore the relationship between HOTAIR and miR-129-5p in regulating CC cell behaviors. QRT-PCR assay showed that oe-HOTAIR transfection markedly reduced miR-129-5p level in propofol-treated HeLa and SiHa cells, while miR-129-5p mimic transfection partially ameliorated the effect (Figure 3E). As demonstrated by colony formation assay, flow cytometry analysis, transwell assay and wound healing assay, the promotional effects on cell colony formation, invasion and migration and the suppressive effect on cell apoptosis in propofol-exposed HeLa and SiHa cells caused by HOTAIR overexpression were all effectively rescued by increasing miR-129-5p (Figure 3F–I). In addition, our results presented that HOTAIR overexpression evidently increased MMP9 and PCNA levels and decreased Bax level in propofol-stimulated HeLa and SiHa cells, while miR-129-5p upregulation overturned the impacts (Figure 3J). Collectively, HOTAIR overexpression promoted cell progression in propofol-treated CC cells by targeting miR-129-5p.
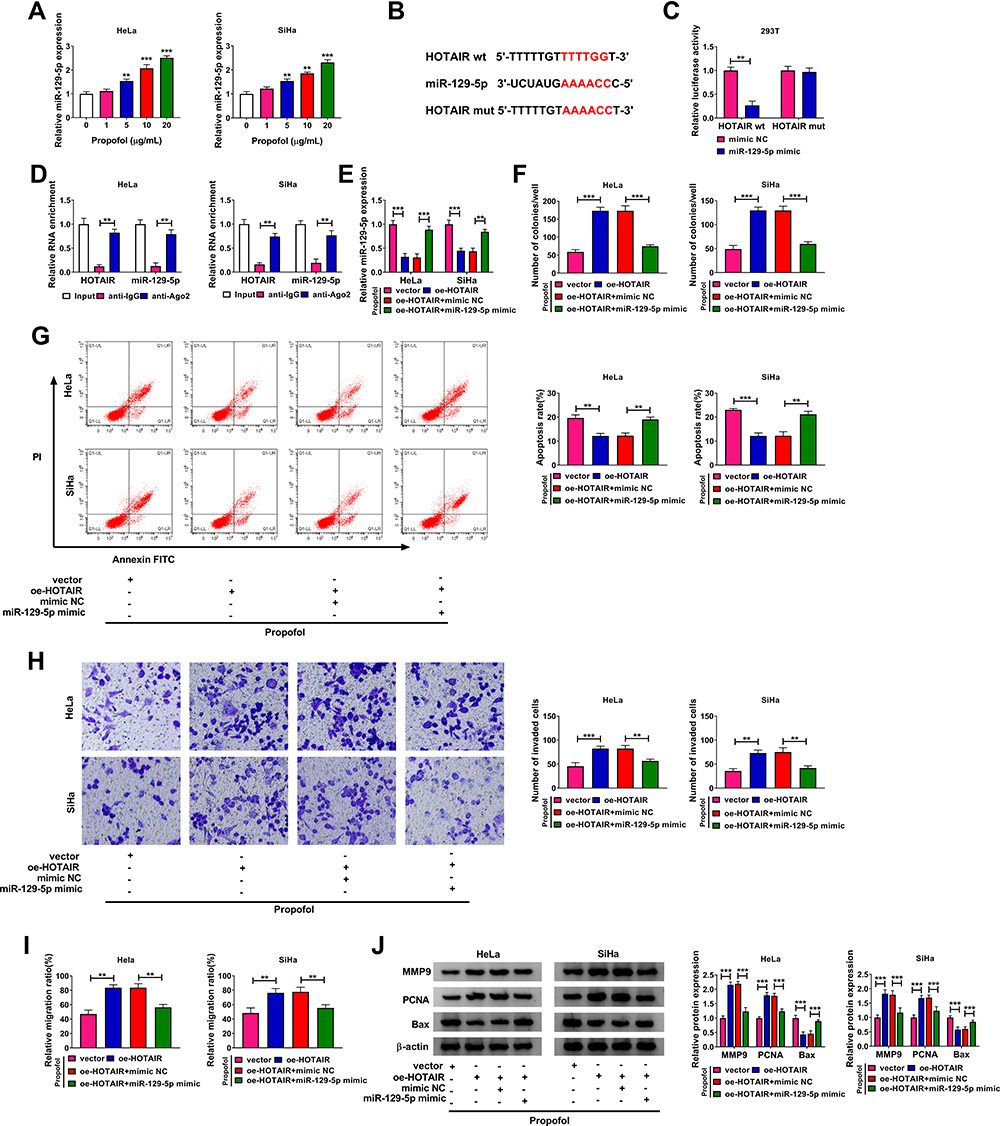 |
Figure 3 HOTAIR overexpression facilitated cell colony formation, invasion and migration and repressed apoptosis in propofol-treated CC cells by sponging miR-129-5p. (A) The level of miR-129-5p in HeLa and SiHa cells treated with different concentrations of propofol was determined by qRT-PCR assay. (B) The potential binding sites between HOTAIR and miR-129-5p were predicted by miRcode. (C) The luciferase activity in HOTAIR wt/HOTAIR mut and mimic NC/miR-129-5p mimic co-transfected 293T cells was measured by dual-luciferase reporter assay. (D) The enrichment of HOTAIR and miR-129-5p in anti-IgG or anti-Ago2 immunoprecipitate was determined by qRT-PCR assay after RIP assay. (E–J) HeLa and SiHa cells were administrated with vector, oe-HOTAIR, oe-HOTAIR+mimic NC or oe-HOTAIR+miR-129-5p mimic and then exposed to 10 µg/mL propofol. (E) The level of miR-129-5p in HeLa and SiHa cells was detected by qRT-PCR assay. (F–I) The colony formation, apoptosis, invasion and migration capacities of HeLa and SiHa cells were assessed by colony formation assay, flow cytometry analysis, transwell assay and wound healing assay, respectively. (J) The protein levels of MMP9, PCNA and Bax in HeLa and SiHa cells were measured by Western blot assay. **P<0.01, ***P<0.001. |
RPL14 Was a Direct Target Gene of miR-129-5p
As presented in Figure 4A and B, propofol treatment apparently decreased the mRNA and protein levels of RPL14 in HeLa and SiHa cells in a concentration-dependent manner. By analyzing the software TargetScan (http://www.targetscan.org/cgi-bin/targetscan/vert_71/view_gene.cgi?rs=ENST00000416518.1&taxid=9606&members=miR-129-5p&showcnc=0&shownc=0&subset=1), RPL14 was found to be a direct target gene of miR-129-5p and their potential binding sites were shown in Figure 4C. Dual-luciferase reporter assay indicated that miR-129-5p mimic transfection remarkably restrained the luciferase activity of RPL14 3ʹUTR wt in 293T cells, but had no effect on the luciferase activity of RPL14 3ʹUTR mut (Figure 4D). RIP assay further confirmed the interaction between miR-129-5p and RPL14 because the enrichment of RPL14 and miR-129-5p was increased by anti-Ago2 compared to anti-IgG groups (Figure 4E). All these outcomes suggested that miR-129-5p directly targeted RPL14.
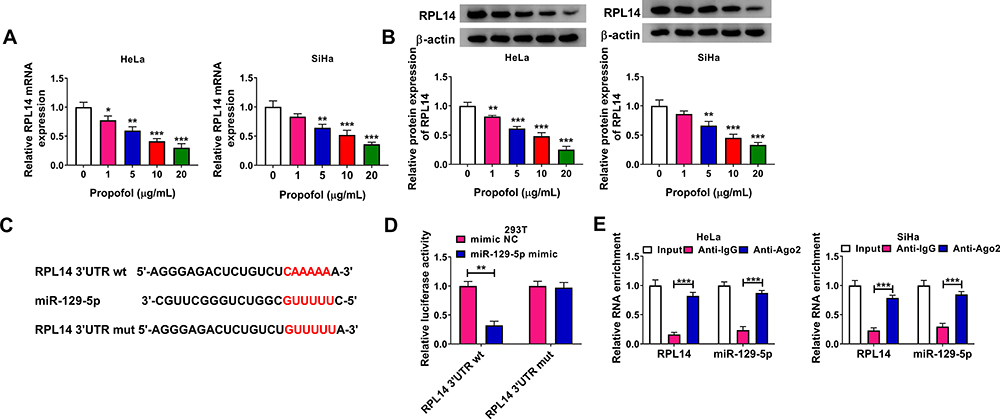 |
Figure 4 RPL14 acted as the target gene of miR-129-5p. (A and B) The mRNA and protein levels of RPL14 in different doses of propofol-treated HeLa and SiHa cells were detected by qRT-PCR assay and Western blot assay, respectively. (C) The predicted complementary sequences between miR-129-5p and RPL14. (D) The interaction between miR-129-5p and RPL14 was investigated by dual-luciferase reporter assay. (E) The interaction between miR-129-5p and RPL14 was verified by RIP assay. *P<0.05, **P<0.01, ***P<0.001. |
Inhibition of miR-129-5p Facilitated the Malignant Behaviors of Propofol-Treated CC Cells by Binding to RPL14
To further investigate the association between miR-129-5p and RPL14, HeLa and SiHa cells were transfected with inhibitor NC, miR-129-5p inhibitor, miR-129-5p inhibitor+si-NC or miR-129-5p inhibitor+si-RPL14 before 10 µg/mL propofol treatment. As exhibited in Figure 5A and B, miR-129-5p inhibition significantly elevated the mRNA and protein levels of RPL14 in propofol-treated HeLa and SiHa cells, whereas si-RPL14 administration effectively abrogated the impacts. Then, the results of colony formation assay, flow cytometry analysis, transwell assay and wound healing assay indicated that miR-129-5p inhibition notably enhanced cell colony formation, invasion and migration and conspicuously repressed apoptosis in propofol-stimulated HeLa and SiHa cells, whereas these effects were all weakened by reducing RPL14 (Figure 5C–F). Furthermore, we observed that miR-129-5p downregulation increased the protein levels of MMP9 and PCNA and decreased the protein level of Bax in propofol-treated HeLa and SiHa cells, while RPL14 suppression abated the impacts (Figure 5G). All these results suggested that miR-129-5p inhibition accelerated cell progression in propofol-treated CC cells by interacting with RPL14.
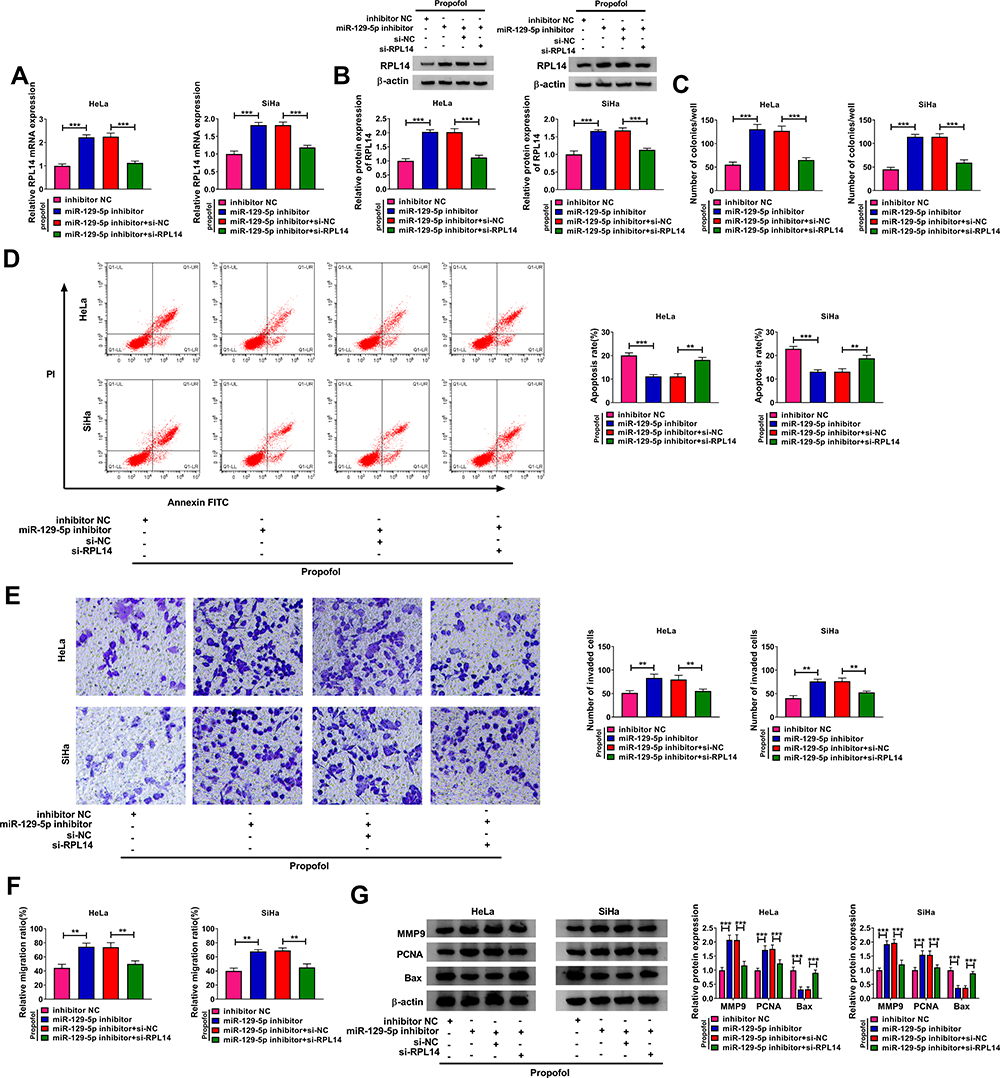 |
Figure 5 MiR-129-5p inhibition facilitated the malignant phenotypes of propofol-treated CC cells by targeting RPL14. Inhibitor NC, miR-129-5p inhibitor, miR-129-5p inhibitor+si-NC or miR-129-5p inhibitor+si-RPL14 was transfected into HeLa and SiHa cells before 10 µg/mL propofol exposure. (A and B) The mRNA and protein levels of RPL14 in HeLa and SiHa cells were detected by qRT-PCR assay and Western blot assay, respectively. (C–F) The colony formation, apoptosis, invasion and migration capacities of HeLa and SiHa cells were assessed by colony formation assay, flow cytometry analysis, transwell assay and wound healing assay, respectively. (G) The protein levels of MMP9, PCNA and Bax in HeLa and SiHa cells were measured via Western blot assay. **P<0.01, ***P<0.001. |
HOTAIR Promoted RPL14 Expression by Targeting miR-129-5p in CC Cells
In order to further explore the relationship among HOTAIR, miR-129-5p and RPL14 in CC cells, vector, oe-HOTAIR, oe-HOTAIR+mimic NC or oe-HOTAIR+miR-129-5p mimic was transfected into HeLa and SiHa cells. Next, we determined the mRNA and protein levels of RPL14 in the transfected HeLa and SiHa cells by qRT-PCR assay and Western blot assay. The results showed that RPL14 mRNA and protein levels were all increased by HOTAIR overexpression, while miR-129-5p mimic transfection abrogated the impacts (Figure 6A–D). Taken together, HOTAIR positively regulated RPL14 expression by targeting miR-129-5p in CC cells.
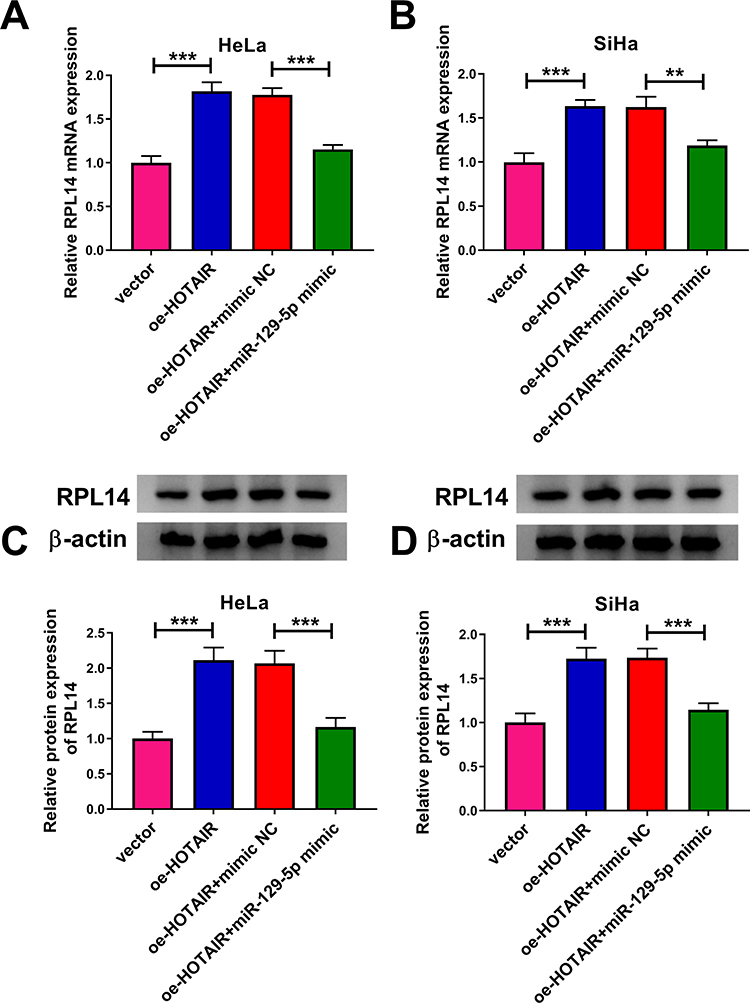 |
Figure 6 HOTAIR overexpression sponged miR-129-5p to modulate RPL14 expression in CC cells. (A–D) After HeLa and SiHa cells were transfected with vector, oe-HOTAIR, oe-HOTAIR+mimic NC or oe-HOTAIR+miR-129-5p mimic, the mRNA and protein levels of RPL14 were examined by qRT-PCR assay and Western blot assay, respectively. **P<0.01, ***P<0.001. |
Propofol Blocked Tumor Growth of CC in vivo
At last, the effect of propofol on the tumorigenesis of CC in vivo was explored by establishing a murine xenograft model. As a result, tumor volume and tumor weight were evidently reduced in Propofol groups and Propofol+vector groups when compared to corresponding control groups (Model groups and Propofol+oe-HOTAIR groups) (Figure 7A and B). In addition, we found that HOTAIR, RPL14 mRNA and RPL14 protein levels were declined and miR-129-5p level was raised in Propofol groups and Propofol+vector groups compared to Model groups and Propofol+oe-HOTAIR groups (Figure 7C and D). Moreover, Propofol groups and Propofol+vector groups exhibited a lower expression of MMP9 and PCNA and a higher expression of Bax compared to Model groups and Propofol+oe-HOTAIR groups (Figure 7D). Additionally, our results showed that miR-129-5p overexpression suppressed tumor growth in vivo (Supplemental Figure 1A and B). Overall, propofol blocked tumorigenesis of CC in vivo by HOTAIR/miR-129-5p axis.
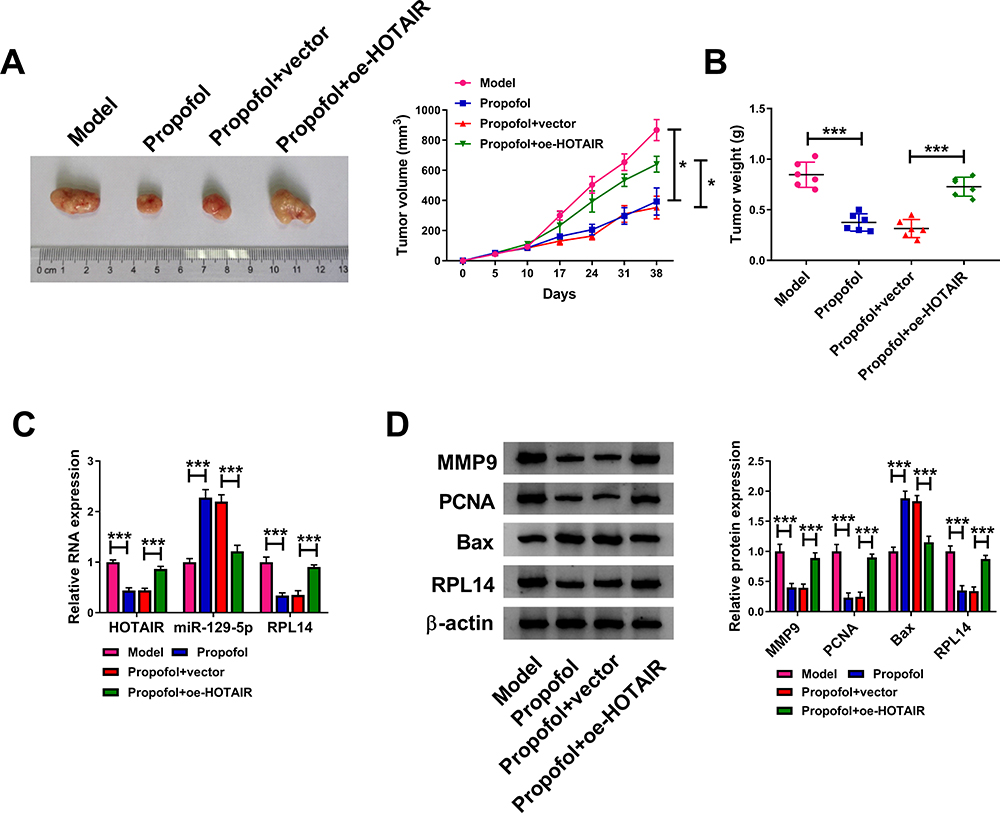 |
Figure 7 Propofol suppressed tumor growth of CC in vivo. The nude mice were divided 4 groups: Model, Propofol, Propofol+vector and Propofol+oe-HOTAIR. (A) Tumor volume was monitored at 0 d, 5 d, 10 d, 17 d, 24 d, 31 d and 38 d. (B) Tumor weight was examined on day 38. (C) The levels of HOTAIR, miR-129-5p and RPL14 in the harvested tumors were detected by qRT-PCR assay. (D) The protein levels of MMP9, PCNA, Bax and RPL14 in the harvested tumors were measured by Western blot assay. *P<0.05, ***P<0.001. |
Discussion
Currently, the anti-tumor effect of propofol has been gradually identified in numerous cancers.21 For example, Zhu et al uncovered that propofol led to a remarkable suppression in gastric cancer cell proliferation and motility and a distinct promotion in apoptosis.22 Sun et al demonstrated that propofol relieved the malignant behaviors of lung cancer cells through downregulating miR-372.23 Herein, we determined the functional roles of propofol in CC. As a result, propofol treatment obviously repressed cell viability, colony formation, invasion and migration and facilitated apoptosis in CC cells.
Subsequently, we explored the underlying mechanisms of propofol in regulating the malignancy of CC cells. Zhang et al claimed that propofol could retard colon cancer progression by regulation of STAT3/HOTAIR axis through activating WIF-1 and repressing Wnt pathway.24 Moreover, propofol was identified to inhibit tumor size and cell viability and facilitate apoptosis in CC by decreasing HOTAIR and mTOR pathway.14 Our results presented that HOTAIR level was evidently declined in CC cells following propofol treatment in a dose-dependent way. HOTAIR has been clarified to serve as a tumor promoter in multiple cancers, including CC.25–27 In this study, we also demonstrated that HOTAIR overexpression could aggravate the malignant phenotypes of CC cells. We therefore further explored the association between propofol and HOTAIR in CC carcinogenesis. Our results exhibited that the impacts on CC cell colony formation, apoptosis, invasion and migration mediated by propofol exposure were all reversed by elevating HOTAIR. Furthermore, we verified that propofol treatment blocked tumorigenesis of CC in vivo, while HOTAIR overexpression partially reversed the impact. All these findings indicated that propofol played a tumor-suppressive role in CC by modulation of HOTAIR.
The regulatory network of lncRNA/miRNA/mRNA axis in tumor progression has attracted more and more attention. For example, HOTAIR contributed to CC development through HOTAIR/miR-143-3p/BCL2 or HOTAIR/miR-206/MKL1 regulatory network.25,26 Herein, we found that HOTAIR positively regulated RPL14 expression by serving as the sponge of miR-129-5p. Zhang et al indicated that the exogenous miR-129-5p was able to restrain CC cell growth and cell cycle process and accelerate apoptosis by binding to SP1.28 Wang et al manifested that miR-129-5p hampered CC cell motility by targeting ZIC2 through inhibiting Hedgehog signaling pathway.19 In our research, we found that miR-129-5p elevation effectively ameliorated the promotional effects on cell colony formation, invasion and migration and the suppressive effect on apoptosis in propofol-treated CC cells mediated by HOTAIR overexpression, suggesting that HOTAIR facilitated propofol-mediated CC cell progression by sponging miR-129-5p. Though miR-129-5p could target diverse genes, such as RET,29 APC30 and COL1A1,31 our study was the first to explore the relationship between miR-129-5p and RPL14. Furthermore, we demonstrated that miR-129-5p suppression facilitated cell colony formation, invasion and migration and hampered apoptosis in propofol-treated CC cells, whereas RPL14 interference overturned the influences.
In conclusion, we revealed that propofol decelerated the progression of CC through regulating HOTAIR/miR-129-5p/RPL14 axis. Our findings suggested that HOTAIR downregulation might facilitate the tumor-suppressive role of propofol in CC.
Ethics Approval
The hospital’s Institutional Review Board approved the current study.
Funding
This work was supported by Science and Technology Research in Henan Province (No. 162102310229).
Disclosure
The authors declare that they have no financial or non-financial conflicts of interest for this work.
References
1. Jemal A, Bray F, Center MM, et al. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi:10.3322/caac.20107
2. Mezei AK, Armstrong HL, Pedersen HN, et al. Cost-effectiveness of cervical cancer screening methods in low- and middle-income countries: a systematic review. Int J Cancer. 2017;141:437–446. doi:10.1002/ijc.30695
3. Small W
4. Gupta S, Maheshwari A, Parab P, et al. Neoadjuvant chemotherapy followed by radical surgery versus concomitant chemotherapy and radiotherapy in patients with Stage IB2, IIA, or IIB squamous cervical cancer: a randomized controlled trial. J Clin Oncol. 2018;36:1548–1555. doi:10.1200/JCO.2017.75.9985
5. Barker HE, Paget JT, Khan AA, et al. The tumour microenvironment after radiotherapy: mechanisms of resistance and recurrence. Nat Rev Cancer. 2015;15:409–425. doi:10.1038/nrc3958
6. Chidambaran V, Costandi A, D’mello A. Propofol: a review of its role in pediatric anesthesia and sedation. CNS Drugs. 2015;29:543–563. doi:10.1007/s40263-015-0259-6
7. Ye Z, Jingzhong L, Yangbo L, et al. Propofol inhibits proliferation and invasion of osteosarcoma cells by regulation of microRNA-143 expression. Oncol Res. 2013;21:201–207. doi:10.3727/096504014X13890370410203
8. Zhang J, Shan WF, Jin TT, et al. Propofol exerts anti-hepatocellular carcinoma by microvesicle-mediated transfer of miR-142-3p from macrophage to cancer cells. J Transl Med. 2014;12:279. doi:10.1186/s12967-014-0279-x
9. Liu Z, Zhang J, Hong G, et al. Propofol inhibits growth and invasion of pancreatic cancer cells through regulation of the miR-21/Slug signaling pathway. Am J Transl Res. 2016;8:4120–4133.
10. Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10:155–159. doi:10.1038/nrg2521
11. Chen F, Li M, Zhu X. Propofol suppresses proliferation and migration of papillary thyroid cancer cells by down-regulation of lncRNA ANRIL. Exp Mol Pathol. 2019;107:68–76. doi:10.1016/j.yexmp.2019.01.011
12. Ren YL, Zhang W. Propofol promotes apoptosis of colorectal cancer cells via alleviating the suppression of lncRNA HOXA11-AS on miRNA let-7i. Biochem Cell Biol. 2020;98:90–98. doi:10.1139/bcb-2018-0235
13. Zhou YH, Cui YH, Wang T, et al. Long non-coding RNA HOTAIR in cervical cancer: molecular marker, mechanistic insight, and therapeutic target. Adv Clin Chem. 2020;97:117–140. doi:10.1016/bs.acc.2019.12.004
14. Zhang D, Zhou XH, Zhang J, et al. Propofol promotes cell apoptosis via inhibiting HOTAIR mediated mTOR pathway in cervical cancer. Biochem Biophys Res Commun. 2015;468:561–567. doi:10.1016/j.bbrc.2015.10.129
15. Ambros V. The functions of animal microRNAs. Nature. 2004;431:350. doi:10.1038/nature02871
16. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi:10.1016/s0092-8674(04)00045-5
17. Li XR, Chu HJ, Lv T, et al. miR-342-3p suppresses proliferation, migration and invasion by targeting FOXM1 in human cervical cancer. FEBS Lett. 2014;588:3298–3307. doi:10.1016/j.febslet.2014.07.020
18. Ye H, Yu X, Xia J, et al. MiR-486-3p targeting ECM1 represses cell proliferation and metastasis in cervical cancer. Biomed Pharmacother. 2016;80:109–114. doi:10.1016/j.biopha.2016.02.019
19. Wang YF, Yang HY, Shi XQ, et al. Upregulation of microRNA-129-5p inhibits cell invasion, migration and tumor angiogenesis by inhibiting ZIC2 via downregulation of the Hedgehog signaling pathway in cervical cancer. Cancer Biol Ther. 2018;19:1162–1173. doi:10.1080/15384047.2018.1491497
20. Feng Y, Ma J, Fan H, et al. TNF-alpha-induced lncRNA LOC105374902 promotes the malignant behavior of cervical cancer cells by acting as a sponge of miR-1285-3p. Biochem Biophys Res Commun. 2019;513:56–63. doi:10.1016/j.bbrc.2019.03.079
21. Kim R. Effects of surgery and anesthetic choice on immunosuppression and cancer recurrence. J Transl Med. 2018;16:8. doi:10.1186/s12967-018-1389-7
22. Zhu F, Li Q, Yang Y, et al. Propofol suppresses proliferation, migration, invasion and promotes apoptosis by upregulating microRNA-140-5p in gastric cancer cells. Onco Targets Ther. 2019;12:10129–10138. doi:10.2147/OTT.S225360
23. Sun H, Gao D. Propofol suppresses growth, migration and invasion of A549 cells by down-regulation of miR-372. BMC Cancer. 2018;18:1252. doi:10.1186/s12885-018-5175-y
24. Zhang YF, Li CS, Zhou Y, et al. Effects of propofol on colon cancer metastasis through STAT3/HOTAIR axis by activating WIF-1 and suppressing Wnt pathway. Cancer Med. 2020;9:1842–1854. doi:10.1002/cam4.2840
25. Zheng P, Yin Z, Wu Y, et al. LncRNA HOTAIR promotes cell migration and invasion by regulating MKL1 via inhibition miR206 expression in HeLa cells. Cell Commun Signal. 2018;16:5. doi:10.1186/s12964-018-0216-3
26. Liu M, Jia J, Wang X, et al. Long non-coding RNA HOTAIR promotes cervical cancer progression through regulating BCL2 via targeting miR-143-3p. Cancer Biol Ther. 2018;19:391–399. doi:10.1080/15384047.2018.1423921
27. Lee M, Kim HJ, Kim SW, et al. The long non-coding RNA HOTAIR increases tumour growth and invasion in cervical cancer by targeting the Notch pathway. Oncotarget. 2016;7:44558–44571. doi:10.18632/oncotarget.10065
28. Zhang J, Li S, Yan Q, et al. Interferon-beta induced microRNA-129-5p down-regulates HPV-18 E6 and E7 viral gene expression by targeting SP1 in cervical cancer cells. PLoS One. 2013;8:e81366. doi:10.1371/journal.pone.0081366
29. Duan L, Hao X, Liu Z, et al. MiR-129-5p is down-regulated and involved in the growth, apoptosis and migration of medullary thyroid carcinoma cells through targeting RET. FEBS Lett. 2014;588:1644–1651. doi:10.1016/j.febslet.2014.03.002
30. Li M, Tian L, Wang L, et al. Down-regulation of miR-129-5p inhibits growth and induces apoptosis in laryngeal squamous cell carcinoma by targeting APC. PLoS One. 2013;8:e77829. doi:10.1371/journal.pone.0077829
31. Wang Q, Yu J. MiR-129-5p suppresses gastric cancer cell invasion and proliferation by inhibiting COL1A1. Biochem Cell Biol. 2018;96:19–25. doi:10.1139/bcb-2016-0254
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.