")
Back to Journals » Infection and Drug Resistance » Volume 13
Prevalence of Virulence Genes and Their Association with Antimicrobial Resistance Among Pathogenic E. coli Isolated from Egyptian Patients with Different Clinical Infections
Authors Abd El-Baky RM , Ibrahim RA , Mohamed DS, Ahmed EF, Hashem ZS
Received 4 December 2019
Accepted for publication 4 April 2020
Published 28 April 2020 Volume 2020:13 Pages 1221—1236
DOI https://doi.org/10.2147/IDR.S241073
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sahil Khanna
Rehab Mahmoud Abd El-Baky,1,2 Reham Ali Ibrahim,1 Doaa Safwat Mohamed,2 Eman Farouk Ahmed,2 Zeinab Shawky Hashem1
1Department of Microbiology and Immunology, Faculty of Pharmacy, Minia University, Minia 61519, Egypt; 2Department of Microbiology and Immunology, Faculty of Pharmacy, Deraya University, Minia 11566, Egypt
Correspondence: Rehab Mahmoud Abd El-Baky
Department of Microbiology and Immunology, Faculty of Pharmacy, Minia University Tel +20 1092487412
Email [email protected]
Introduction: Escherichia (E.) coli can cause intestinal and extra-intestinal infections which ranged from mild to life-threatening infections. The severity of infection is a product of many factors including virulence properties and antimicrobial resistance.
Objectives: To determine the antibiotic resistance pattern, the distribution of virulence factors and their association with one another and with some selected resistance genes.
Methods: Virulence properties were analyzed phenotypically while antimicrobial susceptibility was tested by Kirby-Bauer agar disc diffusion method. In addition, 64 E. coli isolates were tested for 6 colicin genes, fimH, hlyA, traT, csgA, crl virulence genes and bla−CTX-M-15, bla−oxa-2, and bla−oxa-10 resistance genes by polymerase chain reaction (PCR).
Results: Extra-intestinal pathogenic E. coli isolated from urine and blood samples represented a battery of virulence factors and resistance genes with a great ability to produce biofilm. Also, a significant association (P< 0.05) among most of the tested colicin, virulence and resistance genes was observed. The observed associations indicate the importance and contribution of the tested factors in the establishment and the progress of infection especially with Extra-intestinal E. coli (ExPEC) which is considered a great challenging health problem.
Conclusion: There is a need for studying how to control these factors to decrease the rate and the severity of infections. The relationship between virulence factors and resistance genes is complex and needs more studies that should be specific for each area.
Keywords: E. coli, virulence, resistance, colicin genes, ESBL, bla−CTX-M-15, bla−oxa-2, bla−oxa-10
Introduction
The acquisition of virulence and resistance genes is believed to increase the pathogenicity of microorganisms and the severity of infection with the great possibility of therapy failure. Escherichia coli is an opportunistic pathogen, commensal bacteria that can be found as normal flora in humans and animals. It can be classified according to the site of its existence into commensal, intestinal and extra-intestinal E. coli. Commensal E. coli may acquire many virulence genes that may, in combination, result in intestinal and extra-intestinal E. coli infections. Also, diarrheagenic E. coli strains can be classified according to their virulence into different pathotypes which are enterotoxigenic (ETEC), enteropathogenic (EPEC), enterohemorrhagic (EHEC), enteroinvasive (EIEC), and enteroaggregative (EaggEC). Each one of these pathotypes has some virulence factors that in combination are responsible for their pathogenicity of each type and have an important role in infection. For instance, toxin production, hemolysins, proteases, cell surface hydrophobicity, colicins, etc.1,2 E. coli produces bacteriocins such as colicins and microcins which are bactericidal peptides and considered as virulence factors in different E. coli strains or pathotypes.3 Colicins produced by E. coli under stress; prohibiting the colonization of other E. coli strains without affecting their producing strains. Also, colicins increase the host colonization by their producing strain. So, the resistance of other strains to colicins is considered as a cause of their pathogenicity.4
Over 25 different colicins have been specified and categorized on the base of their pathways. Colicins E1 inhibit the synthesis of all macromolecular without respiration arrest.5 Genes encoding colicins E1 were found in ExPEC4 especially uropathogenic E. coli (UPEC). Colicins E2, E7, E8 and E9 cause cleavage of DNA,6 colicins E3, E4 and E6 lead to rRNA hydrolysis,7 while colicins E5 cleave tRNA.8 Microcins V ar urovirulence,9 while colicins B are highly identified in bowel diseases caused by E. coli.10
Furthermore, virulence factors in UPEC are mainly genes encoding adhesion like fimbriae which facilitate invasion and colonization of epithelial cells11 and thin flexible aggregative protein filaments which are known as curli and their encoding genes are crl and csgA.12 Adhesion increases the expression of bacterial toxins, iron acquisition and eschewal the host defense mechanisms.13 The adhesion genes which are mostly prevalent are iha and fimH;14 while toxin gene is hlyA.15 ExPEC in bloodstream is able to overcome the serum bactericidal effect encoded by traT gene.16
In addition to the role of acquisition of virulence genes and its effect on the pathogenicity, the acquisition of resistance genes plays an important role in therapeutic failure and the increase of mortality rate. The inability to control the emergence of multi-drug (MDR), extensive drug (XDR) and pandrug resistance will increase the mortality rates to 10 million people by 2050. Misuse and overuse of antibiotics in therapeutic purposes for human and therapeutic purposes and as growth promotors in livestock lead to the emergence of resistance to many antimicrobial classes as penicillins, cephalosporines, tetracyclines, sulfonamides, macrolides and polymyxins.17
One of the most prevalent resistance factors is extended spectrum ß-lactamase (ESBL) produced by Enterobacteriaceae which first identified in the 1980s.18 The most prevalent ESBL is CTX-M type (cefotaxinase)19 and E. coli having CTX-M ESBL was multidrug-resistant to most drugs.20 OXA ß-lactamase (oxacillinase) is another type which was identified as chromosomally mediated enzyme and revealed a high resistance to penicillin and showed more carbapenems resistance activity.21 Variation of OXA ß-lactamases takes place by alteration of their amino acids and some E. coli isolates may have many variants of OXA ß-lactamases. So, the single strain may have variable types of ESBLs.22
The aim of this study was to determine the antibiotic resistance pattern, the distribution of different virulence factors of E. coli isolated from different sources of infections and their association with one another and with some selected resistance genes.
Methods
Patients, Samples and Identification of Isolates
Our study comprised 200 patients, who were admitted to Minia university hospital during the period from January to July 2019. In brief, 125 males and 75 females were included with a median age of 46 (range 23–56). Samples were collected from patients with urinary tract infections (80 urine samples), gastroenteritis (50 stool samples), septicemia (30 blood samples) and wound infections (40 wound swab samples) as part of the routine hospital laboratory procedures. Samples were obtained after 2 days of their hospitalization. The following antibiotics were used until the results of cultures were available: two doses of ciprofloxacin (urinary tract infections), one dose of cefotaxime (wound infections), two doses of metronidazole and ciprofloxacin (diarrhea) and one dose of imipenem and vancomycin (septicemia).
All samples were cultured and identified using the conventional microbiological procedures. Samples positive for E. coli showed pink colonies on MacConkey agar (Oxoid, UK). E. coli positive cultures were confirmed by different biochemical tests (catalase, sugar fermentation, indole and nitrate reduction tests) and the formation of metallic sheen on Eosin methylene blue agar (EMB) (Himedia, India).
Phenotypic Identification of Virulence Properties of E. coli Isolates
Virulence properties were determined phenotypically as follows: Hemolysis was tested by inoculating isolates into 5% sheep blood agar plates. Clear zones around colonies indicate hemolysin production. Biofilm formation was tested using the tissue culture plate method (TCP).23 Cell surface hydrophobicity was determined by the salt aggregation test (SAT). Protease test was detected by culturing the tested isolates on skim milk agar and the formation of clear zones around colonies is considered positive. Mannose resistant and mannose sensitive haemagglutination test was done by mixing one drop of blood group “O” with a drop of bacterial cultures on a slide, followed by rotating the slide for 5 min at 37̊C. Clumps formed were indicated as haemagglutination. Mannose sensitive haemagglutination was indicated by the addition of 2% w/v of d-mannose. The absence of haemagglutination indicates Mannose sensitivity while the presence of Haemagglutination indicates Mannose resistance. Curli fimbriae expression was determined by growing the tested isolates on agar plates containing 0.1% tryptone, 0.05% yeast extract, 0.002% Coomassie brilliant blue, 0.004% Congo red and 1.5% agar. Curli production can be detected by the presence of red colonies while white colonies were negative for curli. Colicin production: fresh E. coli cultures were cultured on the surface of nutrient agar for 24 hrs at 37°C. Plates were exposed for chloroform vapors for 2 hrs, and then left for 30 mins to evaporate chloroform. Isolates were inoculated perpendicular to the original cultures. The tested isolates were examined for inhibition of growth.24–29
Antimicrobial Susceptibility Testing and Phenotypic Detection of ESBL
Antimicrobial susceptibility testing of E. coli isolates was performed on Mueller Hinton agar plates using the Kirby–Bauer agar disc diffusion method.30 Antibiotic discs used in this study were obtained from Bioanalyse (Turkey). Isolates were defined as sensitive or resistant depending on the measurement of inhibition zone diameters following the criteria of Clinical Laboratory standards Institute (CLSI). The incidence of resistance to each antibiotic was calculated by the number of resistant isolates to specific antibiotics/total number of isolates Multiplied by 100. Detection of ESBL production was performed by a double-disk synergy test using E. coli ATCC 25922 as a control.31,32
DNA Extraction
The DNA template was prepared by the boiling of the suspensions of bacterial pellets for 10 min and using the supernatant directly in the PCR assay.33
PCR Primers and Testing Conditions
PCR was used to amplify the targeted virulence and resistance genes. PCR reaction mixture was done in 25 μL reaction volumes containing 12.5 μL master mix (Bioline, USA), 1 µL of each 10 μmol−1 forward and reverse primers (Laboratories Midland Certified Reagent Company Inc.), 2 µL DNA template and 8.5 µL pyrogen-free water. PCR cycling was performed using the conditions summarized in Table 1 according to Yamamoto et al,34 Johnson et al,35 Schamberger et al,36 Schamberger et al,37 Bhattacharjee et al,38 Pal and Singh,12 Lin et al,39 and Tahamtan et al40 PCR products were analyzed using 1.5% agarose gel electrophoresis containing ethidium bromide at 8 V/cm. Then, the reaction product was visualized under Gel doc/UV transilluminator.41
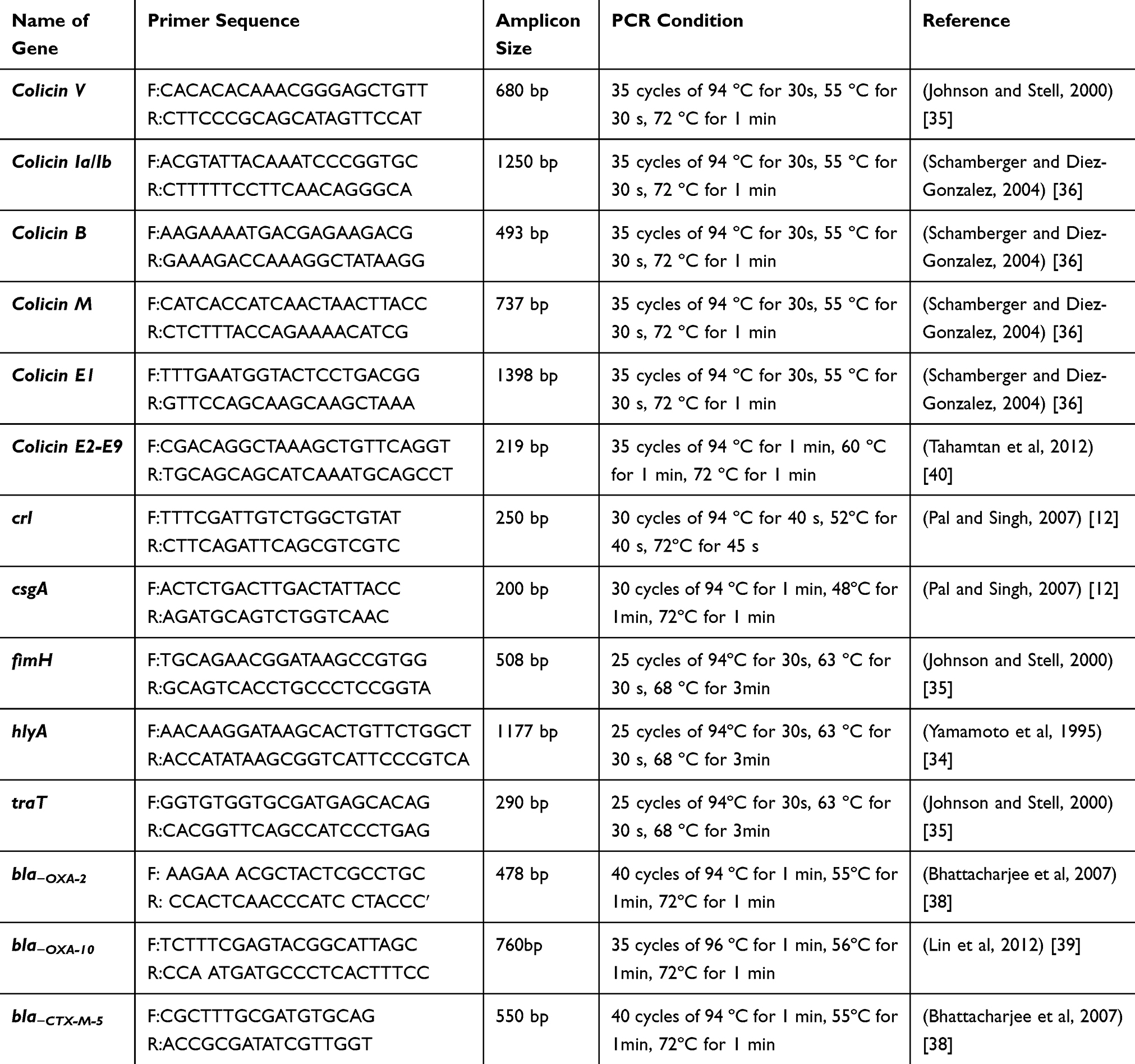 |
Table 1 List of Primers Used in This Study |
Statistical Analysis
Statistical analysis was performed using SPSS, 17 statistical software (SPSS Inc., Chicago, IL). In order to compare the frequencies obtained for phenotypic properties, virulence genes and antibiotic resistance, chi-square (X2) and Fisher’s exact test were used. Correlations were established using Pearson’s correlation coefficient (r2) in bivariate linear correlations (P< 0.05). P-value is significant if it is ≤0.05.
Results
Out of 200 samples, 96 (48%) and 88 (44%) samples were positive for Gram-negative and Gram-positive bacteria, respectively, while 16 samples (8%) were negative for growth.
Out of 96 Gram-negative bacteria, 64 isolates (66.66%) were confirmed as E. coli (10 (15.6%) from wound samples, 22 (34.4%) from urine samples, 23 (35.9%) from stool samples and 9 (14%) from blood samples).
Phenotypic Characteristics
E. coli isolated from urine samples showed greater hemolytic activity (19/22, 86.3%), MRHA (14/22, 63.6%) (Adherence mediated by non-type I pili) and curli fimbriae production (20/22, 90.9%) compared to E. coli isolated from other sources. On the other hand, about 88.8% (8/9 isolates) and 66.6% (6/9 isolates) of E. coli isolated from blood samples showed high cell surface hydrophobicity and mannose sensitivity (adherence mediated by type I pili). A high incidence of strong biofilm production was observed among E. coli isolated from urine (5/22, 22.7%) followed by those isolated from blood samples (2/9, 22.2%) (Table 2).
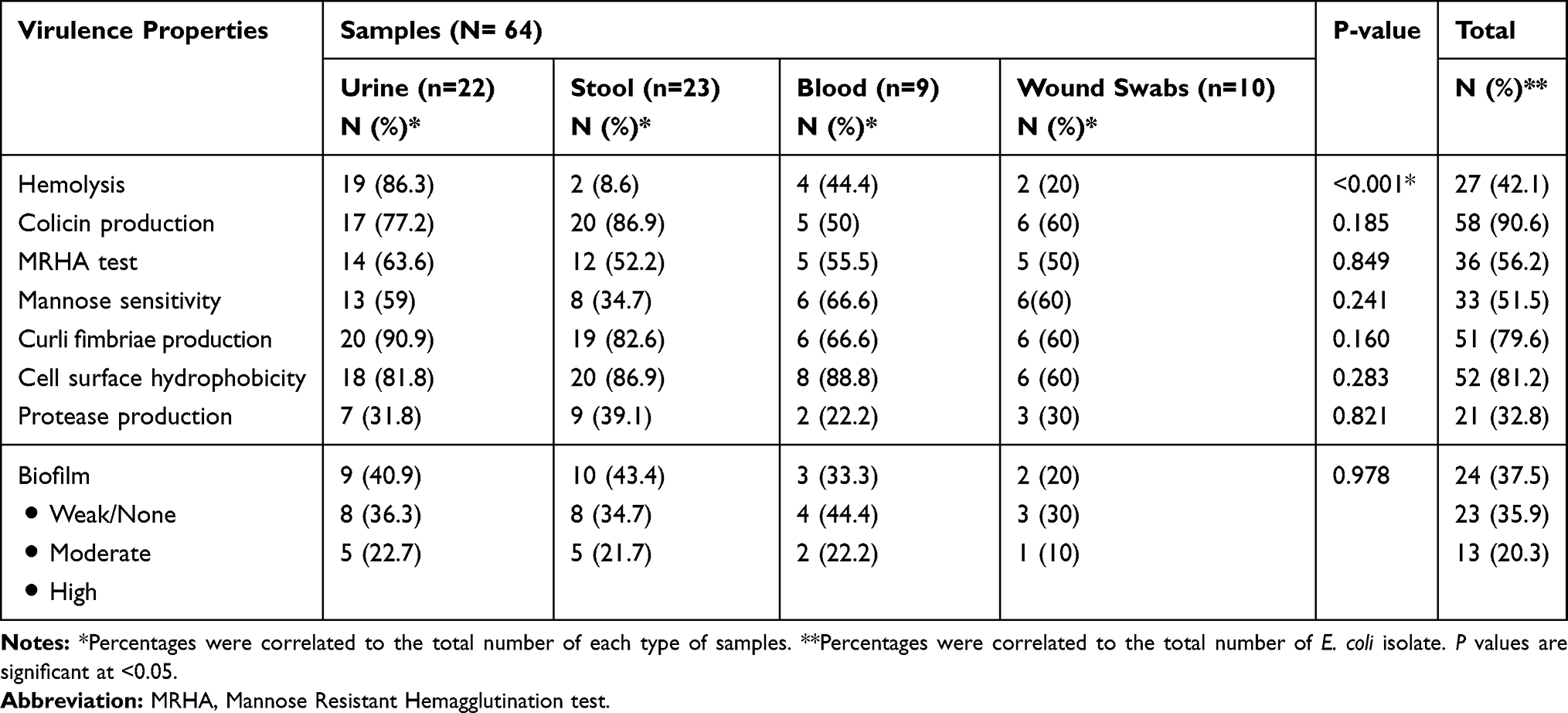 |
Table 2 Distribution of Virulence Properties Among E. coli Clinical Isolates Collected from Different Infections |
Prevalence of Virulence Genes Among the Tested Isolates
Irrespective of the source of E. coli isolates, the most common virulence genes among all isolates were csgA and crl (50/64 (78.1%) and 49/64 (76.5%), respectively) and fimH (48/64, 75%). The distribution of virulence factors among E. coli isolates isolated from different sources is represented in Table 3 which showed that all eight virulence genes tested appeared in the tested urine and stool samples. On the other hand, col la-lb was determined among E. coli isolated from different sources with high prevalence among E. coli isolated from stool (19/23, 82.6%) while colV was more common among fecal and blood isolates (14/23, 60.8% and 5/9, 55.5%, respectively) (Table 3).
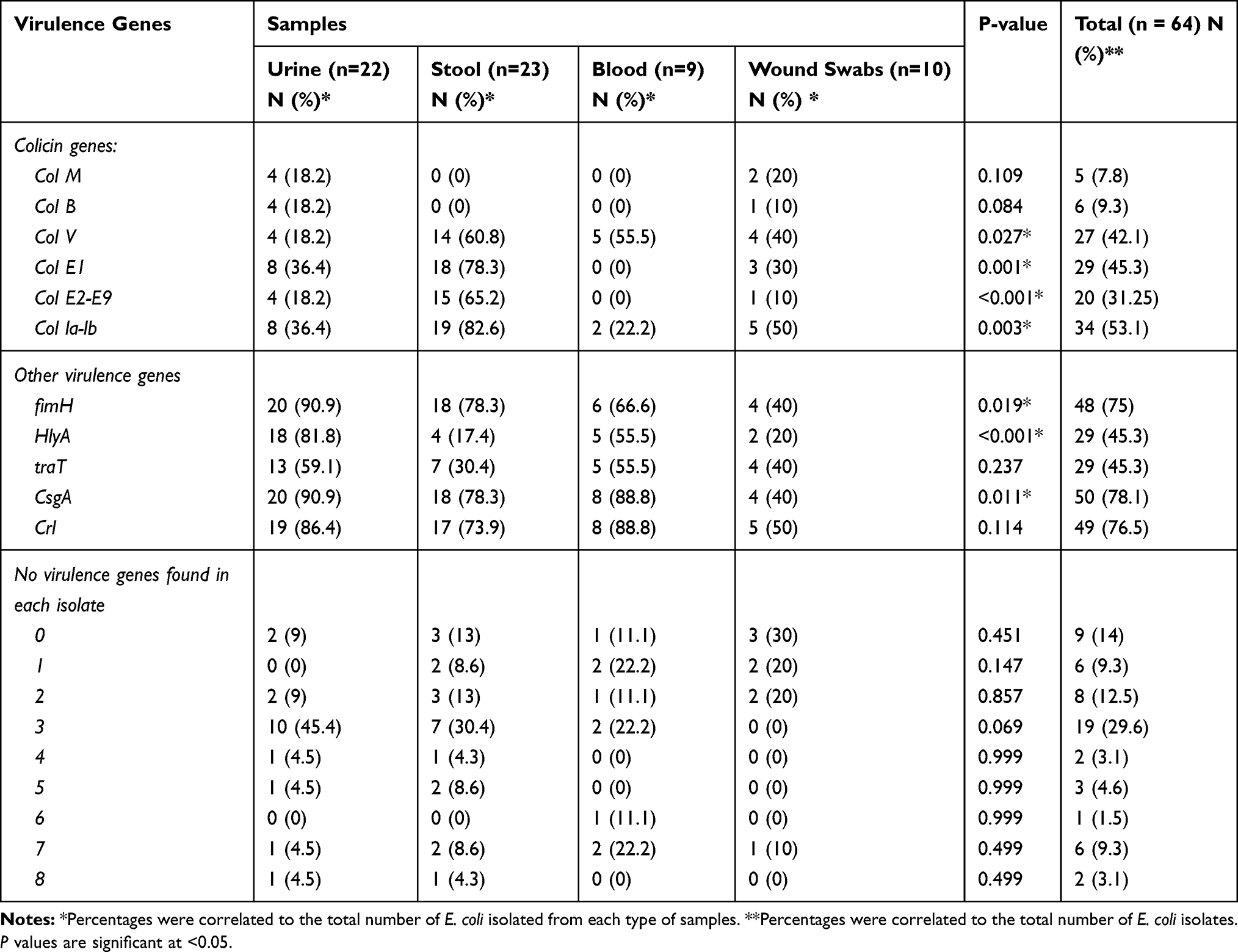 |
Table 3 Distribution of Virulence Genes Among E. coli Clinical Isolates Collected from Different Infections |
High prevalence of colE1 genes was observed among E. coli isolated from stool and urine samples (18/23, 78.3% and 8/22, 36.4%, respectively). colM and colB genes were not identified in any isolates obtained from blood and stool samples but low prevalence for both genes was observed among E. coli isolated from wound swab samples (20% for colM and 10% for colB, respectively) and uropathogenic E. coli (4/22, 18.2% each) was observed. Also, all E. coli isolates obtained from blood samples were negative for col E1, col E2-E9. On the other hand, fimH (90.9%), csgA (90.9%), HlyA (81.8%) and crl (86.4%) genes were the most common virulence genes in UPEC while crl and csgA (88.8% each) genes were the most common virulence genes in E. coli isolated from blood samples (Table 3). The difference in the distribution and frequency of virulence genes among E. coli isolates with respect to the source of samples is illustrated in Tornado Figure 1.
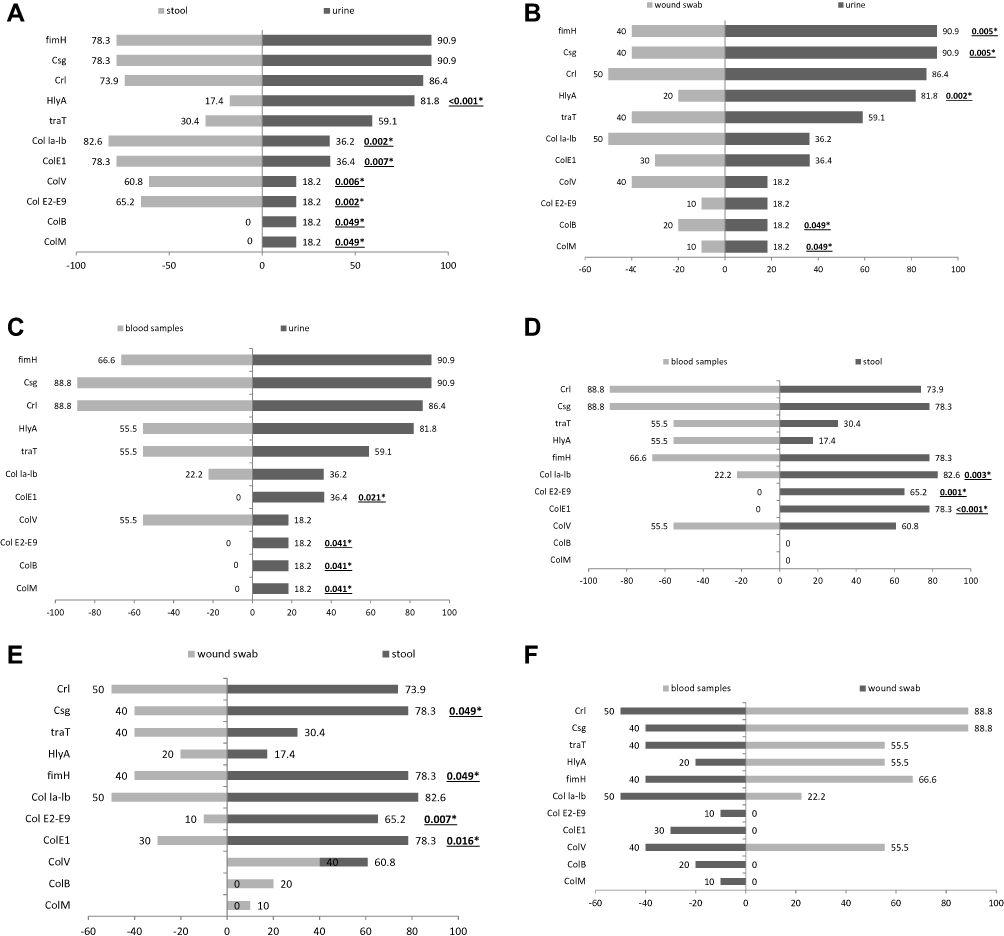 |
Figure 1 Virulence genotypes of the tested E. coli isolates based on the type of samples. (A): Stool and urine samples. (B): Wound and urine samples. (C): Blood and urine samples. (D): Blood and stool samples. (E): Wound and stool samples. (F): Blood and wound samples. |
Antimicrobial Resistance of the Tested Isolates
No significant differences (P-value >0.05) among E. coli isolates of different sources in the antibiotic resistance pattern were observed. Figure 2 shows that Meropenem and imipenem were the most effective antibiotics. E. coli isolated from stool samples showed the lowest resistance to meropenem (13%) while those isolated from wound samples showed the lowest resistance to imipenem (20%).
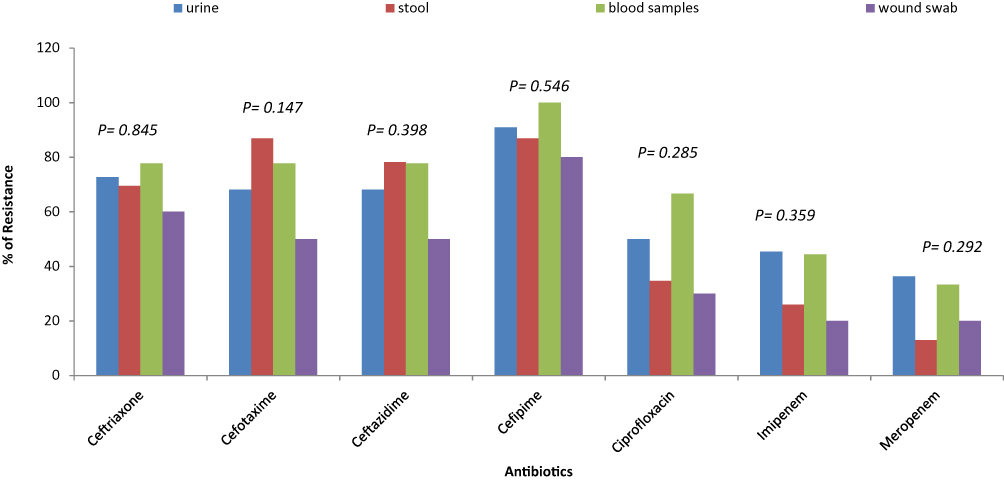 |
Figure 2 Distribution of antibiotic resistance among E. coli clinical isolates of different sources. |
Prevalence of ESBL Production and the Tested Resistance Genes Among Isolates
No significant differences (P-value >0.05) among the distribution of the resistance genes and the source of samples were reported in this study. Table 4 shows that ESBL production was common among E. coli isolates isolated from blood and urine samples (5/9, 55.5 and 12/22, 54.5%, respectively). Also, bla−CTX-M-15 was more common among blood isolates followed by those isolated from urine (4/9, 44.4 and 8/22, 36.3%, respectively). The co-existence of bla−CTX-M-15 with bla−OXA-10 (1/9, 11.1%) and bla−OXA-2 (2/9, 22.2%) were more common among E. coli isolated from blood in comparison to those obtained from other sources. In addition, no association between bla−OXA-2 and bla−OXA-10 was observed among the tested isolates (Table 4).
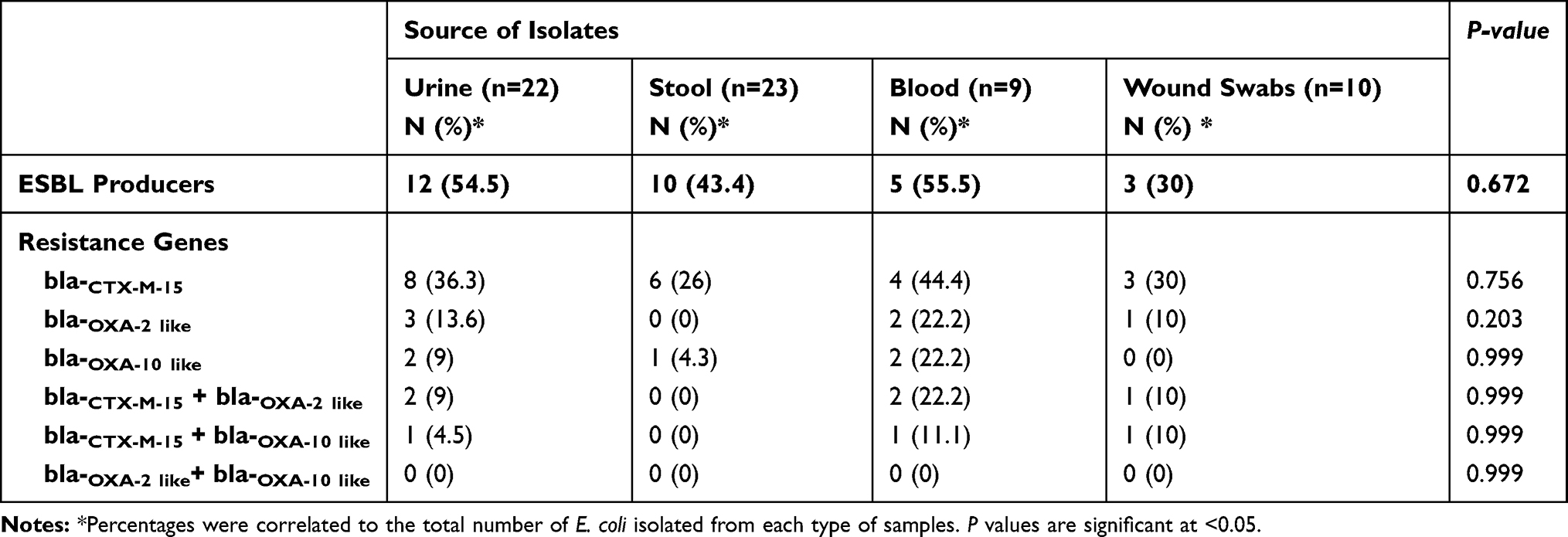 |
Table 4 Distribution of ESBL Production and Resistance Genes Among the Isolated E. coli from Different Sources |
Associations Among Virulence Factors
There were distinctive, complex associations and relationships among the tested virulence factors and with one another (Tables 5–8). Colicin genes were found to be common among E. coli isolated from urine and stool samples. In UPEC, a significant strong positive association between colM with col E1, colB, traT and csgA was reported. colB was positively associated with colV, colE1, col Ia-Ib and crl genes, while colV showed a positive association with colE1, col Ia-Ib, fimH, traT and crl genes. col E2-E9 showed a positive association with col Ia-Ib and csgA genes, while col Ia-Ib showed a positive association with hlyA, traT, csgA and crl genes (Table 5). A moderate positive association was reported among the tested genes in E. coli isolated from wound samples (Table 6).
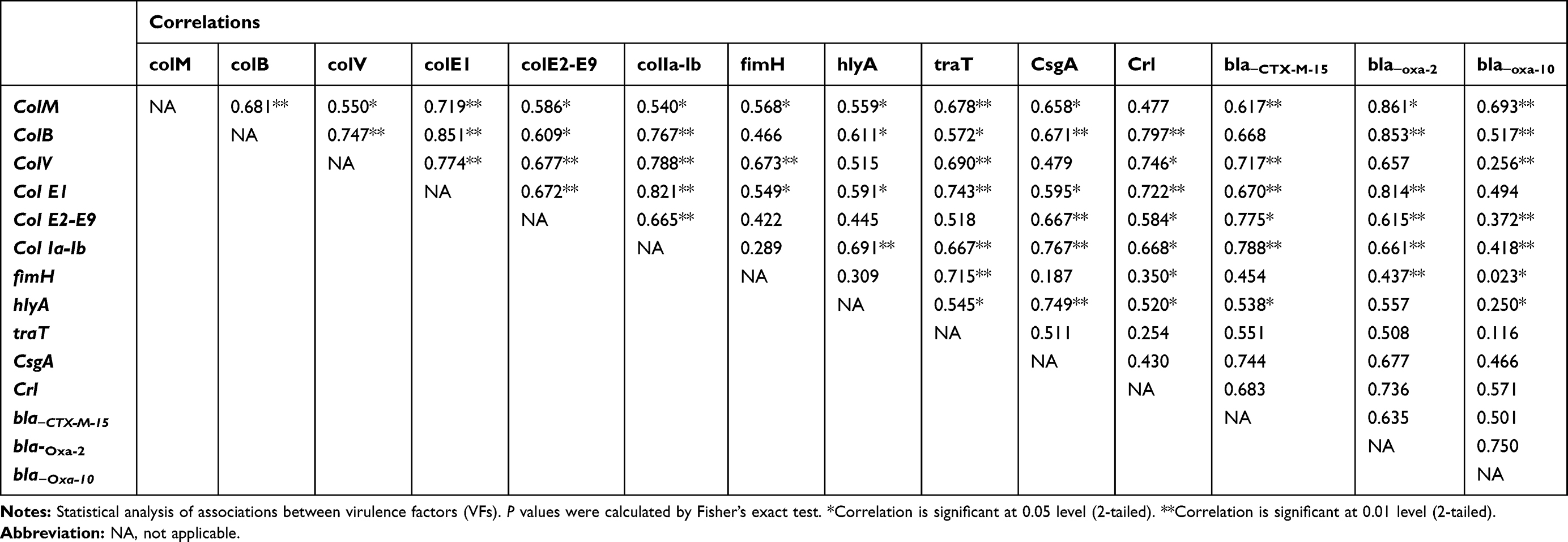 |
Table 5 Relationships Between Virulence Factors Genes and Resistance Genes in Uropathogenic E. coli Isolates |
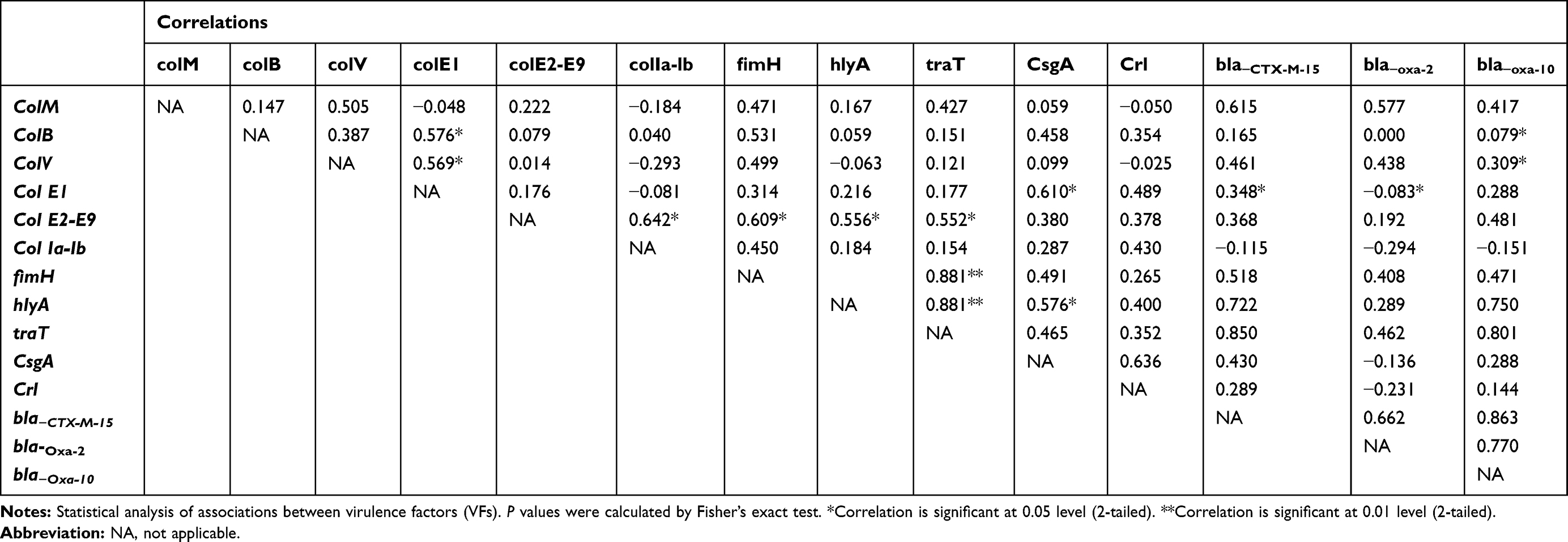 |
Table 6 Relationships Between Virulence Factors Genes and Resistance Genes in E. coli Isolated from Wound Samples |
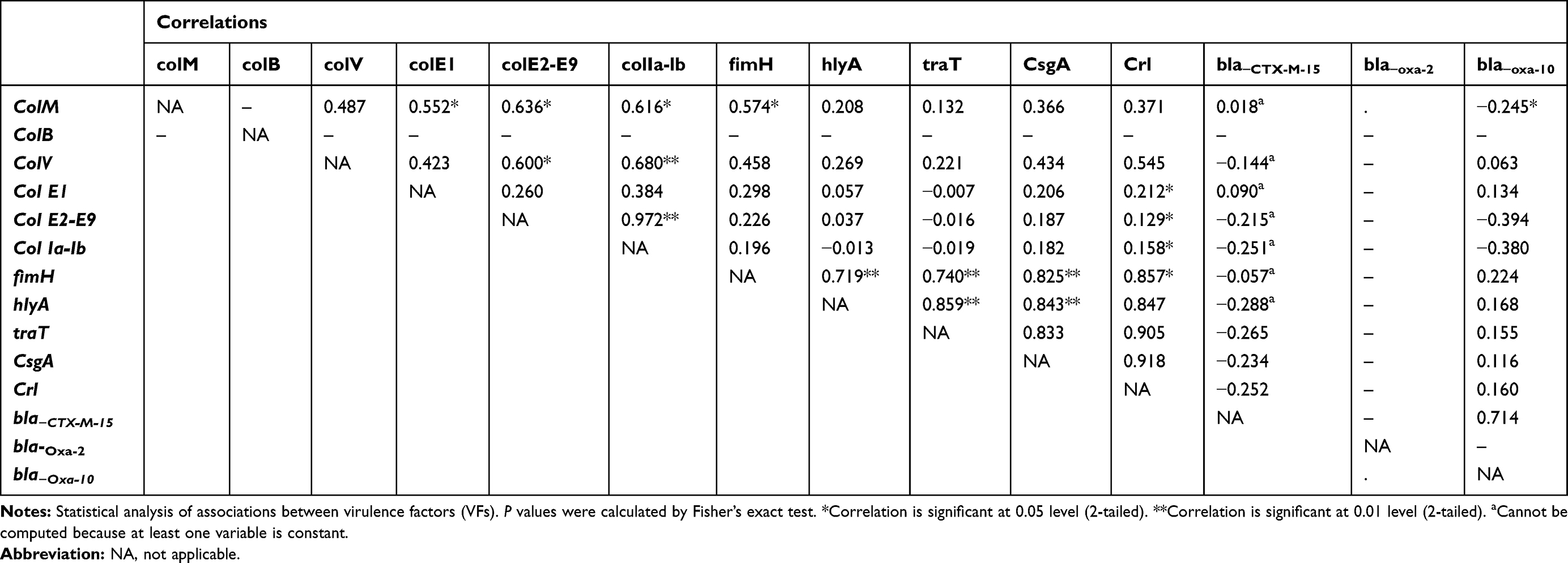 |
Table 7 Relationships Between Virulence Factors Genes and Resistance Genes in Fecal E. coli Isolates |
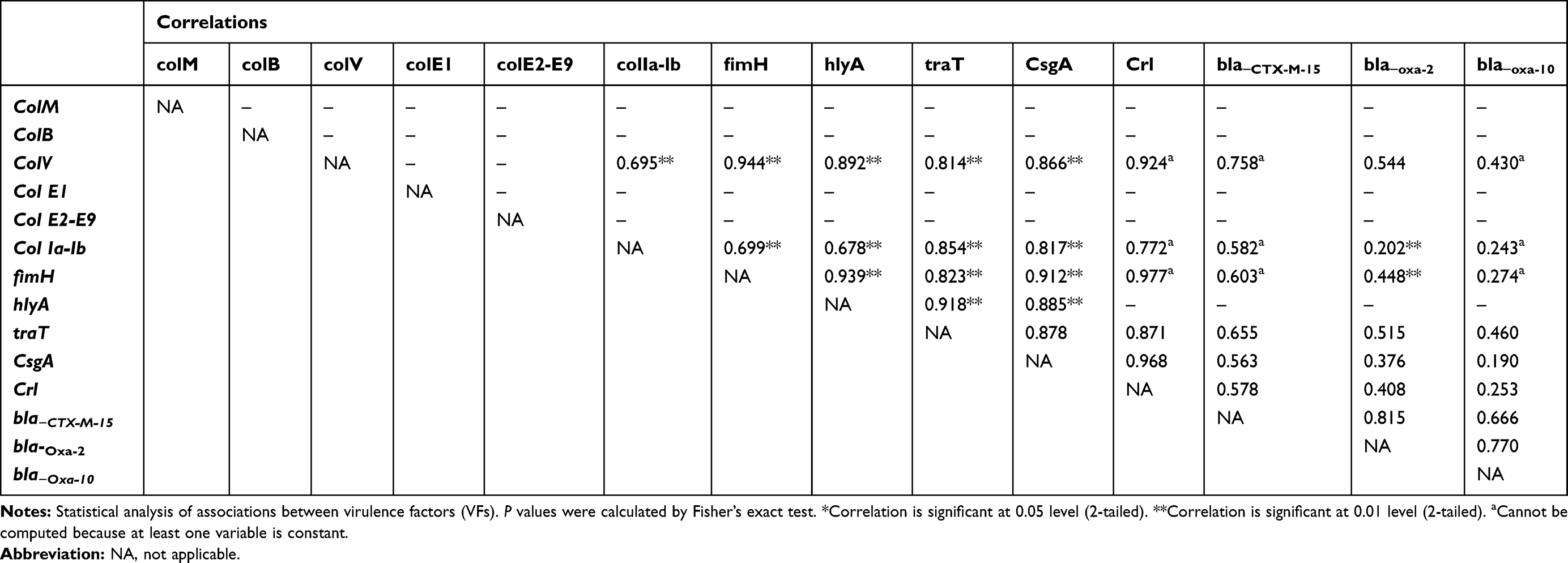 |
Table 8 Relationships Between Virulence Factors Genes and Resistance Genes in E. coli Isolated from Blood Samples |
For E. coli isolated from stool samples, colM showed positive association with colE2-E9, col Ia-Ib and fimH. colV showed a moderate association with col E2-E9, col Ia-Ib (Table 7). A forceful positive correlation between colV and fimH, hlyA, traT, csgA and crl genes was observed in E. coli isolated from blood samples (Table 8).
fimH gene showed significant strong association only with traT gene in uropathogenic E. coli (UPEC) isolates and those isolated from wound infections but showed a significant strong positive association with hlyA, traT, csgA and crl genes in E. coli isolated from stool and blood samples. On the other hand, hlyA showed a strong positive association with csgA gene in UPEC isolates. Furthermore, hlyA showed a strong correlation with fimH, traT, csgA in blood and stool isolates. In fecal isolates, a strong association between hlyA, traT, csgA and crl genes was observed (Tables 5–8).
Association of Virulence Factors and Resistance Genes
It was found that bla−CTX-M-15 gene showed significant (P<0.01) positive association with colV, colE2-E9, col Ia-lb, hlyA and csgA in UPEC isolates but showed a negative association with the tested virulence genes in E. coli isolated from stool. Also, bla−CTX-M-15 showed no significant correlation with colV, fimH, traT genes in E. coli isolated from blood samples and colV, fimH, hlyA and traT in E. coli isolated from Wound samples. bla−OXA-2 showed strong correlations with colM, colB, colE and crl genes in UPEC but showed moderate correlations with colM in wound isolates. bla-OXA-10 was mostly associated with bla-CTX-M-15 in case of E. coli isolated from stool and blood samples, with traT, hlyA and bla-CTX-M-15 in case of E. coli isolated from wound samples and with colM, colB, crl and bla-CTX-M-15 genes in case of uropathogenic E. coli (Tables 5–8).
Discussion
Having many bacterial virulence factors was reported to affect the severity and the extent of infection of any pathogenic microorganisms. In addition, the ability of a microorganism to cause diseases depends not only on their virulence factors but also due to the patient underlying diseases and the other host determinants.42–44
Fimbriae and pili have a role in the hydrophobic characters of bacterial cells and in the adhesion. Hemolytic activity plays a role in tissue damage and the interference with the local immune response. Also, MRHA are adhesive factors which are essential in the well establishment of E. coli to various tissues. Cell surface hydrophobicity of the bacterial cell surface promotes the adherence of the bacteria to various surfaces like the mucosal epithelial cells. Our results showed that the hemolytic activity, mannose resistant haemagglutination and curli fimbriae production were more common in the urinary tract infection isolates in comparison to other extra-intestinal and fecal isolates which were in agreement with that reported by Fakruddin et al45 and Najar et al25 Furthermore, we found that hydrophobic E. coli were common among blood isolates that were different from that reported by Fakruddin et al45 who found that urinary isolates were more hydrophobic than isolates from other sources but blood isolates showed low hydrophobicity with high values of salt aggregation test (SAT) suggesting that surface hydrophobicity has the minor role in the pathogenesis of septicemia. Shruthi et al46 reported that MRHA was more in fecal isolates than in urine isolates.
Biofilm formation is a crucial step that facilitates the initial adhesion, exopolysaccharide production and subsequent dispersion and spread. Our results showed no significant difference among the tested E. coli isolates isolated from different sources but it was observed that biofilm production was most common among isolates obtained from blood and urine samples. Reisner et al47 reported that biofilm production is not dependent on the E. coli origin but it is affected by the composition of growth media, environmental conditions and the expression of some biofilm promoting factors such as curli and nonconjugative pili which can increase biofilm production. In addition, most E. coli isolates were moderate biofilm producers (urine (45.4%), stool (34.7), blood (44.4%) and wound swabs (30%)). Prevalence of moderate production of biofilm by urine isolates was also reported by Samie and Nkgau48 Protease production was common in fecal and urine isolates that may indicate the ability of these isolates to cause damage to urinary and the intestinal cells. Fujishige et al49 and Vermelho et al50 reported the importance of detecting the proteolytic activity of the microorganisms that help in the understanding of their role in the pathogenesis and tissue damage.
Colicin production is an important character that is observed in both pathogenic and commensal E. coli. Colicins have mainly three cytotoxic functions which are: nuclease activity causing degradation of DNA or RNA of the target cells (colicins E2-E9), pore-forming colicins which can form channels and depolarize cytoplasmic membranes (colB, col Ia-Ib, K) and the inhibition of peptidoglycan synthesis that only represented by colicin M.51
Irrespective to E. coli isolates from stool samples, Uropathogenic E. coli showed a high prevalence of colM and colE1 in comparison to other ExPEC that was in agreement with Azpiroz et al52 and Rijavec et al53 ColV (microcin V) was common among ExPEC isolated from blood which agreed with that reported by Davies et al,54 Fakruddin et al45 and Micenková et al55 E. coli isolated from wound samples showed a high prevalence of colM in comparison to the other ExPEC while 50% of isolates were positive for Col Ia-Ib which is in agreement with that reported by Micenková et al.55
There was a positive correlation among colicin production and the expression of other virulence factors. Many researchers showed the high frequency and the positive correlation of bacteriocin production and the expression of many virulence factors indicating the possibility of their contribution to E. coli pathogenicity.9,55-57 Ozanne et al58 and Smith59 showed that mice injected by colV showed 100% death for macrophages in the peritoneal fluids. Also, mice showed symptoms resemble that observed in endotoxin shock. These findings suggested the contribution of colV in E. coli pathogenesis.
In this study we tried to determine the most frequently occurring virulence factors among E. coli isolates isolated from different origins. fimH gene is the gene encoding type 1 fimbriae and is important in the establishment of infections. It was found that fimH, hlyA, traT, csgA, and crl genes were more common among Uropathogenic E. coli compared to isolates of other ExPEC and intestinal isolates indicating that uropathogenic isolates were more virulent than other tested isolates. Cergole-Novella et al60 showed that there was an association among fimH, crl, csg, traT and colV with bla−CTX-M-15 in E. coli isolated from gastroenteritis which was in agreement with our results.
The high prevalence and association of fimH (90.9%) with uropathogenic isolates were reported by many studies.61–63 On the other hand, some studies showed a lower incidence of fimH among uropathogenic isolates such as Tabasi et al64 and Paniagua-Contreras et al14 HlyA gene was more frequently common among uropathogenic E. coli and those isolated from blood (81.8% and 55.5%). Prevalence of hemolysin protein contributes to virulence of both E. coli of urine or blood origin as it is a pore-forming protein. Also, it was found associated with isolates of urinary tract infections that may give rise to bacteremia.65,66 This study showed that traT was more common among E. coli isolated from urine and blood samples (59.1% and 55.5%, respectively). E. coli with serum resistance were highly virulent, as they can escape the complement system and promote serum survival and increase the risk of developing septic shock and the increase in mortality.3,16 In our study, Multi-drug resistance (MDR) (resistance to ≥3 antimicrobials of different classes) to most of the tested antibiotics was more common among urine and blood E. coli isolates especially to cefepime (100% resistance) and the other tested cephalosporins. Ciprofloxacin resistance was observed mostly among urine and blood isolates (50% and 66.6%, respectively) which in agreement with Raeispour and Ranjbar,67 Abdi et al68 and Hashemizadeh et al69 Cergole-Novella et al60 showed that fimH, crl, csg, traT and colV were common among all tested E. coli obtained from different sources (urinary tract infection, septicemia, respiratory infection and gastroenteritis) with 100% prevalence of fimH, crl, csg genes among all isolates followed by traT (83.3%). Also, they showed that biofilm production was observed among isolates from gastroenteritis, sepsis and UTI.
ESBL production was found to be more common among E. coli isolated from blood and urine samples. Also, all tested resistance genes (bla−CTX-M-15, bla−oxa-2, and bla−oxa-10) were found to be more prevalent among E. coli isolates obtained from urine and blood.70 OXA-type β- lactamases have high hydrolytic activity against oxacillin and cloxacillin but they are poorly inhibited by clavulanic acid. OXA-2 and OXA-10 have recently reported to have extended hydrolytic spectrum to include oxyimino cephalosporins. OXA-2 was first reported in pseudomonas spp. then in E. coli from Israel in 2005. Many studies reported that the increase in the expression of different virulence factors results in the increase of the microbial pathogenicity. Also, antibiotic resistance genes expression was found to increase the microbial pathogenicity. So, antibiotic resistance genes were considered as a subtype of virulence factors. By using biofilm as an example, the presence of pili, fimbriae, flagella promotes the adhesion of microbes to biotic or abiotic surfaces.71,72 In addition, these factors promote the formation of biofilm and the expression of quorum sensing signals (regulates cellular functions depending on the cellular density) resulting in the increase of pathogenicity and resistance to antibiotics.73 Furthermore, it was found that there are large plasmids that carry many virulence genes in association with antibiotic resistance genes (hybrid plasmids) which means that selection of these plasmids by antibiotics may select for some virulence characteristics (horizontal gene transfer) as an adverse effect to the antibiotic therapy.72 In another study done by Escudeiro et al,74 the authors reported that there is a strong correlation among virulence factors and antibiotic resistance and the acquisition of new virulence genes is followed by the acquisition of new resistance genes. From the previous findings, there are a widespread of virulence genes in association with resistance genes which increase the need for enhanced surveillance and the emergence of new antimicrobials with anti-virulence ability.
Conclusion
The relationship between virulence factors and resistance genes is complex and needs more studies that should be specific for each area. There is a significant association among colicin, virulence and resistance genes indicating their contribution in the establishment and the progress of infection, especially with ExPEC. Extra-intestinal E. coli isolated from urine and blood samples represent a battery of virulence factors and resistance genes with a great ability to produce biofilm which is considered a great challenging health problem. So, there is a need for studying how to control these factors to decrease the rate and the severity of infections by the emergence of new antimicrobials with anti-virulence ability.
Informed Consent
Informed consent is not required as samples were obtained from the laboratory of hospitals as part of the routine hospital laboratory procedure.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors. The authors received no financial support for the research or publication of this article.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Chapman TA, Wu X-Y, Barchia I, et al. Comparison of virulence gene profiles of escherichia coli strains isolated from healthy and diarrheic swine. Appl Environ Microbiol. 2006;72(7):4782. doi:10.1128/AEM.02885-05
2. Poey ME, Albini M, Saona G, Laviña M. Virulence profiles in uropathogenic Escherichia coli isolated from pregnant women and children with urinary tract abnormalities. Microb Pathog. 2012;52(5):292–301. doi:10.1016/j.micpath.2012.02.006
3. Micenková L, Beňová A, Frankovičová L, et al. Human Escherichia coli isolates from hemocultures: septicemia linked to urogenital tract infections is caused by isolates harboring more virulence genes than bacteraemia linked to other conditions. Int J Med Microbiol. 2017;307(3):182–189. doi:10.1016/j.ijmm.2017.02.003
4. Petkovšek Ž, Žgur-Bertok D, Erjavec MS. Colicin insensitivity correlates with a higher prevalence of extraintestinal virulence factors among Escherichia coli isolates from skin and soft-tissue infections. J Med Microbiol. 2012;61(6):762–765. doi:10.1099/jmm.0.037234-0
5. Konisky J. Colicins and other bacteriocins with established modes of action. Annu Rev Microbiol. 1982;36(1):125–144. doi:10.1146/annurev.mi.36.100182.001013
6. Zakharov SD, Zhalnina MV, Sharma O, Cramer WA. The colicin E3 outer membrane translocon: immunity protein release allows interaction of the cytotoxic domain with OmpF porin. Biochemistry. 2006;45(34):10199–10207.
7. Stahl CH, Callaway TR, Lincoln LM, Lonergan SM, Genovese KJ. Inhibitory activities of colicins against Escherichia coli strains responsible for postweaning diarrhea and edema disease in swine. Antimicrob Agents Chemother. 2004;48(8):3119–3121. doi:10.1128/AAC.48.8.3119-3121.2004
8. Tomita K, Ogawa T, Uozumi T, Watanabe K, Masaki H. A cytotoxic ribonuclease which specifically cleaves four isoaccepting arginine tRNAs at their anticodon loops. Proc Natl Acad Sci. 2000;97(15):8278–8283. doi:10.1073/pnas.140213797
9. Azpiroz MF, Laviña M. Modular structure of microcin H47 and colicin V. Antimicrob Agents Chemother. 2007;51(7):2412–2419. doi:10.1128/AAC.01606-06
10. Micenková L, Frankovičová L, Jaborníková I, et al. Escherichia coli isolates from patients with inflammatory bowel disease: ExPEC virulence-and colicin-determinants are more frequent compared to healthy controls. Int J Med Microbiol. 2018;308(5):498–504. doi:10.1016/j.ijmm.2018.04.008
11. Mulvey MA. Adhesion and entry of uropathogenic Escherichia coli. Cell Microbiol. 2002;4(5):257–271. doi:10.1046/j.1462-5822.2002.00193.x
12. Pal M, Singh S. PCR based detection of adhesive curli gene “crl” and ‘csgA’in avian pathogenic Escherichia coli. Indian J Anim Res. 2007;41(3):226–229.
13. Blum G, Ott M, Lischewski A, et al. Excision of large DNA regions termed pathogenicity islands from tRNA-specific loci in the chromosome of an Escherichia coli wild-type pathogen. Infect Immun. 1994;62(2):606–614. doi:10.1128/IAI.62.2.606-614.1994
14. Paniagua-Contreras GL, Monroy-Perez E, Rodriguez-Moctezuma JR, Dominguez-Trejo P, Vaca-Paniagua F, Vaca S. Virulence factors, antibiotic resistance phenotypes and O-serogroups of Escherichia coli strains isolated from community-acquired urinary tract infection patients in Mexico. J Microbiol Immunol Infect. 2017;50(4):478–485. doi:10.1016/j.jmii.2015.08.005
15. Chiou -Y-Y, Chen M-J, Chiu N-T, Lin C-Y, Tseng -C-C. Bacterial virulence factors are associated with occurrence of acute pyelonephritis but not renal scarring. J Urol. 2010;184(5):2098–2102. doi:10.1016/j.juro.2010.06.135
16. Miajlovic H, Smith SG. Bacterial self-defence: how Escherichia coli evades serum killing. FEMS Microbiol Lett. 2014;354(1):1–9. doi:10.1111/1574-6968.12419
17. Kallau NHG, Wibawan IWT, Lukman DW, Sudarwanto MB. Detection of multi-drug resistant (MDR) Escherichia coli and tet gene prevalence at a pig farm in Kupang, Indonesia. J Adv Vet Anim Res. 2018;5(4):388–396. doi:10.5455/javar.2018.e289
18. Matsumoto Y, Ikeda F, Kamimura T, Yokota Y, Mine Y. Novel plasmid-mediated beta-lactamase from Escherichia coli that inactivates oxyimino-cephalosporins. Antimicrob Agents Chemother. 1988;32(8):1243–1246. doi:10.1128/AAC.32.8.1243
19. Cantón R, Coque TM. The CTX-M β-lactamase pandemic. Curr Opin Microbiol. 2006;9(5):466–475. doi:10.1016/j.mib.2006.08.011
20. Touati A, Benallaoua S, Forte D, Madoux J, Brasme L, De Champs C. First report of CTX-M-15 and CTX-M-3 β-lactamases among clinical isolates of Enterobacteriaceae in Béjaia, Algeria. Int J Antimicrob Agents. 2006;27(5):397–402. doi:10.1016/j.ijantimicag.2005.12.007
21. Poirel L, Girlich D, Naas T, Nordmann P. OXA-28, an extended-spectrum variant of OXA-10 β-lactamase from Pseudomonas aeruginosa and its plasmid-and integron-located gene. Antimicrob Agents Chemother. 2001;45(2):447–453. doi:10.1128/AAC.45.2.447-453.2001
22. Ullah W, Qasim M, Rahman H, et al. CTX-M-15 and OXA-10 beta lactamases in multi drug resistant Pseudomonas aeruginosa: first report from Pakistan. Microb Pathog. 2017;105:240–244. doi:10.1016/j.micpath.2017.02.039
23. Christensen GD, Simpson WA, Younger JJ, et al. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: a quantitative model for the adherence of staphylococci to medical devices. J Clin Microbiol. 1985;22(6):996–1006. doi:10.1128/JCM.22.6.996-1006.1985
24. Kim SH, Kim YH. Escherichia coli O157: h7adherence to HEp-2 cells is implicated with curli expression and outer membrane integrity. J Vet Sci. 2004;5(2):119–124. doi:10.4142/jvs.2004.5.2.119
25. Najar AG, Nejad MM, Mansouri S. The comparison between virulence factors of Escherichia coli isolated from urinary tract infections and feacal flora. Res Pharm Sci. 2007;1(2):99–103.
26. Mansouri S, Norouzi F, Moradi M, Nakhaee N. Comparison of virulence factors among clinical isolates of Pseudomonas aeruginosa producing and non-producing extended spectrum beta-lactamases. Curr Res Bacteriol. 2011;4(3):85–93. doi:10.3923/crb.2011.85.93
27. Mattos-Guaraldi AL, Formiga LCD, Andrade AFB. Cell surface hydrophobicity of sucrose fermenting and nonfermenting Corynebacterium diphtheriae strains evaluated by different methods. Curr Microbiol. 1999;38(1):37–42. doi:10.1007/PL00006769
28. Maheswari UB, Palvai S, Anuradha PR, Kammili N. Hemagglutination and biofilm formation as virulence markers of uropathogenic Escherichia coli in acute urinary tract infections and urolithiasis. Indian J Urol. 2013;29(4):277–281. doi:10.4103/0970-1591.120093
29. Reichhardt C, Jacobson AN, Maher MC, et al. Congo red interactions with curli-producing E. coli and native curli amyloid fibers. PLoS One. 2015;10(10):e0140388. doi:10.1371/journal.pone.0140388
30. Bauer A, Kirby W, Sherris JC, Turck M. Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol. 1966;45(4_ts):493–496. doi:10.1093/ajcp/45.4_ts.493
31. Tzelepi E, Giakkoupi P, Sofianou D, Loukova V, Kemeroglou A, Tsakris A. Detection of extended-spectrum beta-lactamases in clinical isolates of Enterobacter cloacae and Enterobacter aerogenes. J Clin Microbiol. 2000;38(2):542–546. doi:10.1128/JCM.38.2.542-546.2000
32. CLSI. Performance Standards for Antimicrobial Susceptibility Tests.
33. Wilson K. Preparation of genomic DNA from bacteria. Curr Protoc Mol Biol. 2001;Chapter 2:
34. Yamamoto S, Terai A, Yuri K, Kurazono H, Takeda Y, Yoshida O. Detection of urovirulence factors in Escherichia coli by multiplex polymerase chain reaction. FEMS Immunol Med Microbiol. 1995;12(2):85–90. doi:10.1111/j.1574-695X.1995.tb00179.x
35. Johnson JR, Stell AL. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J Infect Dis. 2000;181(1):261–272. doi:10.1086/315217
36. Schamberger GP, Diez-Gonzalez F. Characterization of colicinogenic Escherichia coli strains inhibitory to enterohemorrhagic Escherichia coli. J Food Prot. 2004a;67(3):486–492. doi:10.4315/0362-028X-67.3.486
37. Schamberger GP, Phillips RL, Jacobs JL, Diez-Gonzalez F. Reduction of Escherichia coli O157: H7 populations in cattle by addition of colicin E7-producing E. coli to feed. Appl Environ Microbiol. 2004b;70(10):6053–6060. doi:10.1128/AEM.70.10.6053-6060.2004
38. Bhattacharjee A, Sen MR, Anupurba S, Prakash P, Nath G. Detection of OXA-2 group extended-spectrum-β-lactamase-producing clinical isolates of Escherichia coli from India. J Antimicrob Chemother. 2007;60(3):703–704. doi:10.1093/jac/dkm267
39. Lin SP, Liu MF, Lin CF, Shi ZY. Phenotypic detection and polymerase chain reaction screening of extended-spectrum beta-lactamases produced by Pseudomonas aeruginosa isolates. J Microbiol Immunol Infect. 2012;45(3):200–207. doi:10.1016/j.jmii.2011.11.015
40. Tahamtan Y, Shirazi Z, Pourbakhsh A, et al. Detection of colicin genes by PCR in Escherichia coli isolated from cattle in Shiraz-Iran. Arch Razi Inst. 2012;67(1):63–67.
41. Sambrook J, Russell D. Molecular cloning: a laboratory manual. Mol Cloning a Lab Man. 2001.
42. Dale AP, Woodford N. Extra-intestinal pathogenic Escherichia coli (ExPEC): disease, carriage and clones. J Infect. 2015;71(6):615–626. doi:10.1016/j.jinf.2015.09.009
43. Lefort A, Panhard X, Clermont O, et al. Host factors and portal of entry outweigh bacterial determinants to predict the severity of Escherichia coli bacteremia. J Clin Microbiol. 2011;49(3):777–783. doi:10.1128/JCM.01902-10
44. Jauréguy F, Carbonnelle E, Bonacorsi S, et al. Host and bacterial determinants of initial severity and outcome of Escherichia coli sepsis. Clin Microbiol Infect. 2007;13(9):854–862. doi:10.1111/j.1469-0691.2007.01775.x
45. Fakruddin M, Mazumdar RM, Chowdhury A, Mannan KSB. A preliminary study on virulence factors & antimicrobial resistance in extra-intestinal pathogenic Escherichia coli (ExPEC) in Bangladesh. Indian J Med Res. 2013;137(5):988–990.
46. Shruthi N, Kumar R. Phenotypic study of virulence factors in Escherichia coli isolated from antenatal cases, catheterized patients, and faecal flora. J Clin Diagn Res. 2012;6(10):1699–1703. doi:10.7860/JCDR/2012/4669.2634
47. Reisner A, Krogfelt KA, Klein BM, Zechner EL, Molin S. In vitro biofilm formation of commensal and pathogenic escherichia coli strains: impact of environmental and genetic factors. J Bacteriol. 2006;188(10):3572–3581. doi:10.1128/JB.188.10.3572-3581.2006
48. Samie A, Nkgau T. Biofilm production and antibiotic susceptibility profile of Escherichia coli isolates from HIV and AIDS patients in the Limpopo Province. Afr J Biotechnol. 2012;11(34):8560–8570. doi:10.5897/AJB11.2865
49. Fujishige A, Smith KR, Silen JL, Agard DA. Correct folding of alpha-lytic protease is required for its extracellular secretion from Escherichia coli. J Cell Biol. 1992;118(1):33–42. doi:10.1083/jcb.118.1.33
50. Vermelho AB, Meirelles MNL, Lopes A, Petinate SDG, Chaia AA, Branquinha MH. Detection of extracellular proteases from microorganisms on agar plates. Mem Inst Oswaldo Cruz. 1996;91(6):755–760. doi:10.1590/S0074-02761996000600020
51. Hahn-Löbmann S, Stephan A, Schulz S, et al. Colicins and Salmocins – new classes of plant-made non-antibiotic food antibacterials. Front Plant Sci. 2019;10:437. doi:10.3389/fpls.2019.00437
52. Azpiroz MF, Poey ME, Laviña M. Microcins and urovirulence in Escherichia coli. Microb Pathog. 2009;47(5):274–280. doi:10.1016/j.micpath.2009.09.003
53. Rijavec M, Budič M, Mrak P, Müller-Premru M, Podlesek Z, Žgur-Bertok D. Prevalence of ColE1-like plasmids and colicin K production among uropathogenic Escherichia coli strains and quantification of inhibitory activity of colicin K. Appl Environ Microbiol. 2007;73(3):1029–1032. doi:10.1128/AEM.01780-06
54. Davies D, Falkiner F, Hardy K. Colicin V production by clinical isolates of Escherichia coli. Infect Immun. 1981;31(2):574–579. doi:10.1128/IAI.31.2.574-579.1981
55. Micenková L, Bosák J, Vrba M, Ševčíková A, Šmajs D. Human extraintestinal pathogenic Escherichia coli strains differ in prevalence of virulence factors, phylogroups, and bacteriocin determinants. BMC Microbiol. 2016;16(1):218. doi:10.1186/s12866-016-0835-z
56. Budič M, Rijavec M, Petkovšek Ž, Žgur-Bertok D. Escherichia coli bacteriocins: antimicrobial efficacy and prevalence among isolates from patients with bacteraemia. PLoS One. 2011;6(12):e28769. doi:10.1371/journal.pone.0028769
57. Šmajs D, Micenková L, Šmarda J, et al. Bacteriocin synthesis in uropathogenic and commensal Escherichia coli: colicin E1 is a potential virulence factor. BMC Microbiol. 2010;10(1):288. doi:10.1186/1471-2180-10-288
58. Ozanne G, Mathieu L, Baril J. Production of colicin V in vitro and in vivo and observations on its effects in experimental animals. Infect Immun. 1977;17(3):497–503. doi:10.1128/IAI.17.3.497-503.1977
59. Smith HW. A search for transmissible pathogenic characters in invasive strains of Escherichia coli: the discovery of a plasmid-controlled toxin and a plasmid-controlled lethal character closely associated, or identical, with colicine V. Microbiology. 1974;83(1):95–111.
60. Cergole-Novella MC, Pignatari AC, Guth BE. Adhesion, biofilm and genotypic characteristics of antimicrobial resistant Escherichia coli isolates. Braz J Microbiol. 2015;46(1):167–171. doi:10.1590/S1517-838246120140077
61. Malekzadegan Y, Khashei R, Sedigh Ebrahim-Saraie H, Jahanabadi Z. Distribution of virulence genes and their association with antimicrobial resistance among uropathogenic Escherichia coli isolates from Iranian patients. BMC Infect Dis. 2018;18(1):572. doi:10.1186/s12879-018-3467-0
62. Lee J, Subhadra B, Son YJ, et al. Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea. Lett Appl Microbiol. 2016;62(1):84–90. doi:10.1111/lam.12517
63. Gao Q, Zhang D, Ye Z, et al. Virulence traits and pathogenicity of uropathogenic Escherichia coli isolates with common and uncommon O serotypes. Microb Pathog. 2017;104:217–224. doi:10.1016/j.micpath.2017.01.027
64. Tabasi M, Karam MRA, Habibi M, Yekaninejad MS, Bouzari S. Phenotypic assays to determine virulence factors of uropathogenic Escherichia coli (UPEC) isolates and their correlation with antibiotic resistance pattern. Osong Public Health Res Perspect. 2015;6(4):261–268. doi:10.1016/j.phrp.2015.08.002
65. Daga AP, Koga VL, Soncini JGM, et al. Escherichia coli Bloodstream Infections in Patients at a University Hospital: virulence Factors and Clinical Characteristics. Front Cell Infect Microbiol. 2019;9:191. doi:10.3389/fcimb.2019.00191
66. Sonnen AF-P, Henneke P. Role of pore-forming toxins in neonatal sepsis. Clin Dev Immunol. 2013;2013.
67. Raeispour M, Ranjbar R. Antibiotic resistance, virulence factors and genotyping of Uropathogenic Escherichia coli strains. Antimicrob Resist Infect Control. 2018;7(1):118. doi:10.1186/s13756-018-0411-4
68. Abdi S, Ranjbar R, Vala MH, Jonaidi N, Bejestany OB, Bejestany FB. Frequency of bla TEM, bla SHV, bla CTX-M, and qnrA among Escherichia coli isolated from urinary tract infection. Arch Clin Infect Dis. 2014;9(1):e18690. doi:10.5812/archcid.18690
69. Hashemizadeh Z, Kalantar-Neyestanaki D, Mansouri S. Correlation between hlyA and cnf1 virulent genes with antibiotic resistance and non-ESBLs escherichia coli isolates collected from patient with urinary tract infections in Kerman, Iran. Arch Pediatr Infect Dis. 2017;5(4):e61653. doi:10.5812/pedinfect.61653
70. Alqasim A, Abu Jaffal A, Alyousef AA. Prevalence and molecular characteristics of sequence type 131 clone among clinical uropathogenic Escherichia coli isolates in Riyadh, Saudi Arabia. Saudi J Biol Sci. 2020;27(1):296–302. doi:10.1016/j.sjbs.2019.09.020
71. Ramirez MS, Traglia GM, Lin DL, Tran T, Tolmasky ME. Plasmid-mediated antibiotic resistance and virulence in gram-negatives: the klebsiella pneumoniae paradigm. Microbiol Spectr. 2014;2(5):1–15. doi:10.1128/microbiolspec.PLAS-0016-2013
72. Turton J, Davies F, Turton J, Perry C, Payne Z, Pike R. Hybrid resistance and virulence plasmids in “high-risk” clones of klebsiella pneumoniae, including those carrying blaNDM-5. Microorganisms. 2019;7(9):326. doi:10.3390/microorganisms7090326
73. Rubini D, Varthan PV, Jayasankari S, Vedahari BN, Nithyanand P. Suppressing the phenotypic virulence factors of Uropathogenic Escherichia coli using marine polysaccharide. Microb Pathog. 2020;141:103973. doi:10.1016/j.micpath.2020.103973
74. Escudeiro P, Pothier J, Dionisio F, Nogueira T. Antibiotic resistance gene diversity and virulence gene diversity are correlated in human gut and environmental microbiomes. mSphere. 2019;4(3):e00135–e00119. doi:10.1128/mSphere.00135-19
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.