")
Back to Journals » Infection and Drug Resistance » Volume 16
Prevalence and Characterization of Serratia marcescens Isolated from Clinical Bovine Mastitis Cases in Ningxia Hui Autonomous Region of China
Authors Liang Z , Shen J, Liu J, Sun X , Yang Y, Lv Y, Zheng J, Mou X, Li H, Ding X, Yang F
Received 14 February 2023
Accepted for publication 15 April 2023
Published 4 May 2023 Volume 2023:16 Pages 2727—2735
DOI https://doi.org/10.2147/IDR.S408632
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Zeyi Liang,* Jiahao Shen,* Jing Liu, Xu Sun, Yayuan Yang, Yanan Lv, Juanshan Zheng, Xiaoqing Mou, Hongsheng Li, Xuezhi Ding, Feng Yang
Key Laboratory of New Animal Drug Project of Gansu Province/Key Laboratory of Veterinary Pharmaceutics Discovery, Lanzhou Institute of Husbandry and Pharmaceutical Sciences of Chinese Academy of Agricultural Science, Lanzhou, Gansu, 730050, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Feng Yang; Xuezhi Ding, Lanzhou Institute of Husbandry and Pharmaceutical Sciences of Chinese Academy of Agricultural Science, No. 335 Jiangouyan, Qilihe District, Lanzhou, Gansu, 730050, People’s Republic of China, Tel +86-931-2115262, Fax +86-931-2114180, Email [email protected]; [email protected]
Purpose: This study aimed to investigate the prevalence and genetic characterization of Serratia marcescens isolates from clinical bovine mastitis in Ningxia Hui Autonomous Region of China.
Methods: S. marcescens was identified by the polymerase-chain reaction of 16S rRNA gene and sequencing. Antimicrobial susceptibility was tested by the disk diffusion method. Genes of resistance and virulence were determined by the PCR.
Results: Overall, S. marcescens were confirmed from 32 of 2897 (1.1%) mastitis milk samples. These isolates showed high resistance to cefazolin (30/32, 93.8%) and chloramphenicol (28/32, 87.5%). A 12.5% (4/32) of the isolates displayed multidrug resistance (MDR). The most prevalent resistant genes found in S. marcescens were TEM (32/32, 100%) and CTX-M (24/32, 75.0%; CTX-M-15, 14/32, 43.8%; CTX-M-14, 8/32, 25.0%; CTX-M-65, 2/32, 6.3%) for extended-spectrum beta-lactamase, cmlA (28/32, 87.5%) and floR (16/32, 50.0%) for chloramphenicol resistance, SIM-1 (2/32, 6.3%) for carbapenemases, and sdeB (28/32, 87.5%), sdeY (26/32, 81.3%), sdeR (26/32, 81.3%) and sdeD (20/32, 62.5%) for efflux pumps. Moreover, all isolates carried virulence genes flhD, entB, and kpn, and most of them contained mrkD (30/32, 93.8%), ycfM (26/32, 81.3%), bsmB (26/32, 81.3%), pigP (26/32, 81.3%), kfu (24/32, 75.0%) and shlB (24/32, 75.0%).
Conclusion: To our knowledge, this is the first report of genetic determinants for antimicrobial resistance and virulence in S. marcescens isolated from bovine mastitis cases in China. These findings are useful for developing strategies for prevention and treatment of bovine mastitis caused by S. marcescens in China.
Keywords: Serratia marcescens, bovine mastitis, antimicrobial resistance, virulence
Introduction
Bovine mastitis is one of the most prevalent and costly diseases affecting the dairy cattle industry.1 The etiopathology of this disease is usually involved three factors: exposure to microorganisms, host defense mechanisms, and environmental conditions.2 Although considerable progress has been made in controlling contagious mastitis pathogens through improved milking hygiene, little has been achieved against environmental pathogens, such as Serratia marcescens, which is considered an important opportunistic pathogen and has been found to be associated with outbreaks of mastitis in dairy cows.3
Antibiotic therapy is the main method of treating infections caused by S. marcescens.4 However, the therapeutic effectiveness has been attenuated by emerging resistant strains.5 Meanwhile, the common multidrug-resistant nature of S. marcescens complicates the treatment of its infections.6 Indeed, lines of studies revealed an alarming increase in S. marcescens resistance to the commonly used beta-lactams.7 Antimicrobial resistance of S. marcescens is mainly attributed to different resistance determinants, such as genes of extended-spectrum beta-lactamase (eg, SHV, TEM, and CTX) and carbapenemases (eg, OXA-48, KPC, and NDM) for beta-lactam resistance and bacterial effector proteins (eg, sdeB, sdeD, and sdeY) for multidrug resistance. Moreover, the pathogenicity of S. marcescens is mediated by an arsenal of virulence factors including hemolysin, lipase, protease, prodigiosin, and motility.8
At present, reports of S. marcescens are limited worldwide, especially isolates from bovine mastitis in China. The purpose of this study was to investigate the prevalence, antimicrobial resistance, and virulence genes of S. marcescens in bovine mastitis cases in Ningxia Hui Autonomous Region of China. To the best of our knowledge, this is the first report of genetic determinants for antimicrobial resistance and virulence in S. marcescens isolated from bovine mastitis cases in China.
Materials and Methods
Experimental Design
From July 2018 to May 2022, this study collected milk samples of clinical bovine mastitis from 19 commercial dairy herds in the Ningxia Hui Autonomous Region of China. Quickly place the collected sample in liquid nitrogen and transport it back to the key laboratory of new animal drug project of Gansu province. Store it in a refrigerator at −80°C until the strains in the sample are isolated and purified. All purified isolates were confirmed with PCR. Serratia marcescens was selected and tested for drug resistance to detect the type and quantity of drug resistance and virulence genes carried.
Sample Collection and S. marcescens Isolation
Mastitis milk samples of 2897 cows were collected from 19 commercial dairy herds in Ningxia Hui Autonomous Region of China during July 2018 to May 2022. In this study, clinical examinations were conducted on the breasts and nipples of lactating cows according to Jonathan et al.9 In short, check the symmetry of one quarter of each cow’s breasts. Then, possible fibrosis, inflammation and swelling, visible damage, tick infestation, tissue atrophy, and swelling of lymph nodes on the breast are detected through palpation. Test the viscosity and appearance of milk secretion from each breast segment for the presence of clots, thin sections, blood, and water secretions, in order to determine clinical mastitis.10 After delivery to the laboratory, a volume of 20 μL of each mastitis sample was cultivated on agar plates supplemented with 5% defibrinated sheep blood (Huan kai, Guangdong, China) at 37 °C aerobically for 18 to 24 h. Presumptive S. marcescens colonies based on morphology and gram strain were further identified by PCR and sequencing as described in our previous study.1 Briefly, the genomic DNA was extracted using the Bacterial DNA Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. The 16S rRNA gene was amplified by the 16S rDNA Bacterial Identification PCR Kit (Takara, Shiga, Japan) according to the manufacturer’s recommendations (https://www.takarabiomed.com.cn/DownLoad/RR176.pdf). The PCR products were purified and sequenced by Sanger sequencing by Sangon Biotech (Shanghai) Co., Ltd. in China. The nucleotide sequences obtained were analyzed with the NCBI-BLAST program (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility of S. marcescens to 15 antimicrobial agents was determined by disk diffusion method on Mueller–Hinton agar (MHA; Oxoid, United Kingdom) according to the Clinical and Laboratory Standards Institute (CLSI, 2018). Antimicrobial agents tested in this study include ampicillin (10 μg), cefazolin (30 μg), cefepime (30 μg), ceftazidime (30 μg), cefotaxime (30 μg), ertapenem (10 μg), meropenem (10 μg), imipenem (10 μg), gentamicin (10 μg), tetracycline (30 μg), levofloxacin (5 μg), chloramphenicol (30 μg), trimethoprim/sulfamethoxazole (1.25/23.75 μg). Escherichia coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853 were used as quality control strains. Multidrug resistance was defined as isolates that were resistant to at least 3 classes of the tested antimicrobial agents.11
Genetic Determinants for Antimicrobial Resistance and Virulence
Genes encoding extended-spectrum beta-lactamase (SHV, TEM, CTX-M),12 carbapenemases (IMP, SPM, AIM, VIM, OXA-48, GIM, BIC, SIM-1, NDM, DIM, KPC, GES, and IMI),13,14 chloramphenicol resistance (cmlA and floR),15 and efflux pumps (sdeB, sdeD, sdeR, sdeS, sdeY, and hasF) were detected by PCR as previously described.16
Meanwhile, virulence genes for movement (fimA and fimC: fimbriae; flhD: flagella), adhesins (fimH: viscous protein expressed in type 1 fimbriae; kpn: fimbriae adhesins; mrkD: type 3 fimbria adhesin subunit; rmpA: regulator of the mucoid phenotype; magA: mucoviscosity-associated gene; bsmB: genes for protease and lipase production; ycfM: outer membrane lipoprotein), allantoin metabolism-associated gene (allS), siderophores (iutA: iron uptake transport; entB: enterobactin; fyuA, ybtS, irp-1, and irp-2: yersiniabactin; kfu and iroN), protectines or invasins (K2: type 2 capsular polysaccharide; traT: surface exclusion protein), and toxins (pigP, pigA and pigC: prodigiosin; shlA, shlB, and hlyA: hemolysins; phlA: phospholipase) were also determined though PCR.17–22 The PCR products were analyzed by electrophoresis on 1.2% agarose gel and stained with gel-red (Tsingke, Xi’an, China). The results were visualized and photographed by a UV transilluminator. Additionally, DNA sequencing was carried out on genes of extended-spectrum beta-lactamase and carbapenemases. The sequence analysis of these genes was similar to that of the 16S rRNA gene.
Results
Bacterial Isolates
Overall, 32 (32/2897, 1.1%) S. marcescens isolates were obtained from 2897 mastitis milk samples from Ningxia Hui Autonomous Region of China. S. marcescens isolates showed resistance to cefazolin (30/32, 93.8%), chloramphenicol (28/32, 87.5%), ampicillin (12/32, 37.5%), gentamicin (4/32, 12.5%), trimethoprim/sulphamethoxazole (4/32, 12.5%), tetracycline (4/32, 12.5%), cefotaxime (2/32, 6.3%), and meropenem (2/32, 6.3%). However, all tested isolates were susceptible to ceftazidime, ertapenem, levofloxacin, cefepime, and imipenem. Besides, 4 (4/32, 12.5%) of S. marcescens isolates displayed multidrug resistance.
Genotypic Resistance Profiles of S. marcescens
The results of resistant genes showed that all S. marcescens isolates carried TEM (32/32, 100%). And most of them contained cmlA (28/32, 87.5%), sdeB (28/32, 87.5%), sdeY (26/32, 81.3%), sdeR (26/32, 81.3%), sdeD (20/32, 62.5%), CTX-M (24/32, 75.0%; CTX-M-15, 14/32, 43.8%; CTX-M-14,8/32, 25.0%; CTX-M-65, 2/32, 6.3%) and floR (16/32, 50.0%). A few isolates harbored hasF (10/32, 31.3%), and SIM-1 (2/32, 6.3%). While genes encoding for extended-spectrum beta-lactamase (SHV), carbapenemases (IMP, SPM, AIM, VIM, OXA-48, GIM, BIC, NDM, DIM, KPC, GES, and IMI) and efflux pumps (sdeS) were not detected in any of the isolates (Table 1 and Supplementary Figure 1–24).
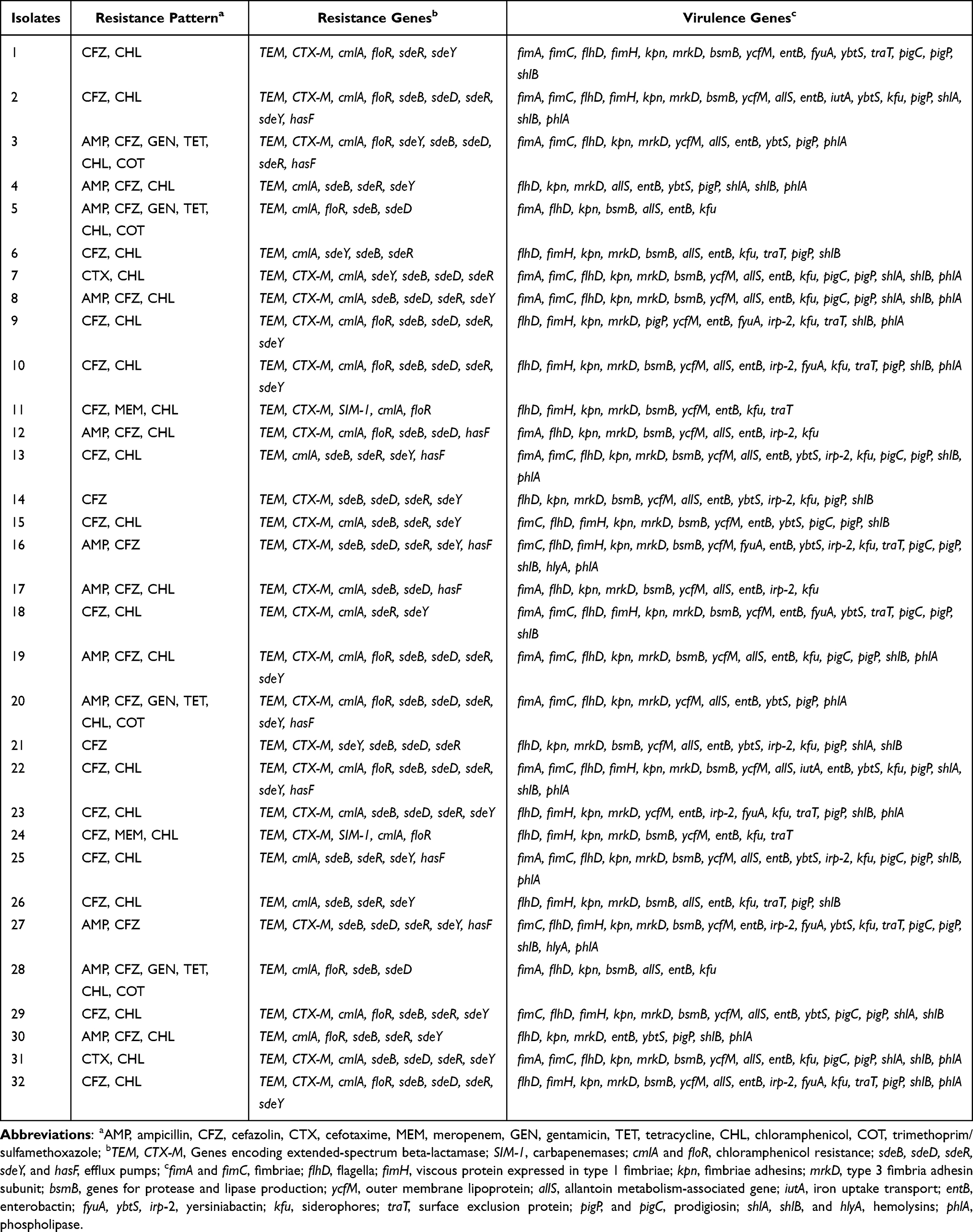 |
Table 1 Resistance Determinants and Virulence Genes of Serratia marcescens Isolates from Clinical Bovine Mastitis |
Genotypic Virulence Profiles of S. marcescens
We also detected the virulence-encoding genes of the S. marcescens isolates (Table 1). All S. marcescens isolates carried flhD, kpn, and entB, followed by mrkD (30/32, 93.8%), bsmB (26/32, 81.3%), ycfM (26/32, 81.3%), pigP (26/32, 81.3%), kfu (24/32, 75.0%), shlB (24/32, 75.0%), allS (22/32, 68.8%), phlA (18/32, 56.3%) fimA (16/32, 50.0%), fimC (16/32, 50.0%), fimH (16/32, 50.0%), irp-2 (12/32, 37.5%), traT (12/32, 37.5%), pigC (12/32, 37.5%), ybtS (16/32, 50.0%), fyuA (8/32, 25.0%), shlA (8/32, 25.0%), hlyA (2/32, 6.3%), and iutA (2/32, 6.3%). Notably, each of the tested isolates carried at least 8 virulent genes. In addition, all isolates were negative for rmpA, magA, irp-1, iroN, K2, and pigA (Table 1 and Supplementary Figure 25–52).
Discussion
S. marcescens is an Enterobacteriaceae microorganism that is widespread in the environment,23 which can cause both clinical and subclinical mastitis outbreaks during the lactation and the dry period in dairy cows.24,25 In this study, S. marcescens was identified in 1.10% of the mastitis milk samples. The incidence was similar to a previous study reported in China that 1.5% of the bovine mastitis samples were positive for S. marcescens in bovine mastitis,26 but lower than the 4.5% reported in Korea and 35–39% in outbreaks of mastitis in Finland.3 These frequency variations may be due to different sample sizes, different seasons, or geographical discrepancies.27
S. marcescens is an emerging pathogen with increasing clinical importance due to its intrinsic resistance to different antimicrobials.28 In this study, S. marcescens isolates showed high resistance to cefazolin and chloramphenicol. Similar results were reported that S. marcescens and other Enterobacteriaceae isolates from bovine mastitis frequently exhibited resistance to these antimicrobials.1,29,30 We found that Serratia marcescens is resistant to cefazolin and sensitive to Cefepime, Ceftazidime, Cefotaxime, etc. This result is the same as that of previous studies.31 Cefazolin and chloramphenicol are broad-spectrum antibiotics with therapeutic effects on bovine pathogens.32 The high resistance could be due to the long-term and widespread use of these antimicrobials on dairy farms.33 In addition, an increase in exposure to antimicrobial drugs can lead to an increase in drug-resistant strains.34 Veterinarians on farms may be exposed to chloramphenicol environments, and the chloramphenicol-resistant bacteria they carry are then transferred to Serratia marcescens through plasmid mediated transfer. Carbapenems are the last resort β-lactam antibiotics for treating infections caused by multidrug-resistant Gram-negative bacteria. Unfortunately, carbapenem-resistant pathogens have emerged with the increasing clinical use of carbapenems and now pose a great threat to human health.35 In our study, although only 2 S. marcescens isolates exhibited carbapenem resistance, their potential threat should not be ignored.
The phenotypic resistance is mainly conferred by corresponding genes, which can be transferred to other bacteria and pose a serious threat to public health through the food chain.36 The most common antimicrobial resistance found in our study was to cefazolin and chloramphenicol. Thus, the resistant genes against β-lactams and chloramphenicol were detected. We found that all S. marcescens isolates carried TEM alone or combined with CTX-M and showed resistance to at least one of the tested β-lactams. Similarly, the cmlA was found in all chloramphenicol-resistant isolates alone or in combination with floR. These results indicating that genes TEM and cmlA may play an important role in β-lactams and chloramphenicol resistance in S. marcescens isolates from bovine mastitis in Ningxia Hui Autonomous Region of China, respectively. These findings were consistent with other reports that these genes were frequently observed in Enterobacteriaceae isolates from food producing animal in China and other countries.30,37,38 Carbapenems have been the most successful β-lactams in evading bacterial resistance. However, carbapenem resistance, mediated by acquired carbapenemase genes, has been increasingly reported.39 It is noteworthy that we found the two carbapenem-resistant S. marcescens isolates were positive for the carbapenemase gene SIM-1. To the best of our knowledge, this is the first report of a SIM-1 producing S. marcescens from bovine mastitis in China. Additionally, efflux pumps also play important roles in β-lactams and chloramphenicol resistance, as well as the multidrug resistance.40 Past studies reported that S. marcescens enhanced its infectivity and its inherent resistance to antibiotics through plasmids and efflux pumps, which greatly contributed to the emergence of multiple drug-resistant strains and finally complicated the treatment process,41,42 and the underlying mechanisms were complicated.43 In our study, regardless of the two S. marcescens isolates resistant to cefazolin, meropenem, and chloramphenicol simultaneously, the efflux pump-related genes sdeB, sdeD, sdeR, sdeS, sdeY, or hasF (in combination) were observed in all β-lactams and chloramphenicol-resistant isolates, respectively. Efflux pump is helpful for bacteria to excrete antibiotics, and it is the main cause of clinical intrinsic and acquired multidrug resistance in bacteria.44 Our results showed that only 12.5% of the S. marcescens isolates were multidrug resistance. This disparity could be due to the limited antimicrobial classes tested in our study.
Pathogenic microorganisms normally express several virulence determinants facilitating their invasion and evasion of the host defenses and cause disease.45 Bacteria adhere to the tissues employing fimbria or pili, thereby establishing infection.46 Fimbriae of S. marcescens encoded by fimA, fimC and fimH play vital roles in surface attachment and colonization.47 Additionally, flagella-controlled swarming motility is governed by flhD and contributes to cell surface attachment.18,48 In this study, all S. marcescens carried flhD gene, and 50.0% of the isolates were positive for fimA and fimC. Besides, we also found that all S. marcescens isolates harbored the adhesions protein-related virulence gene kpn, the majority of the isolates carried mrkD (93.8%), ycfM (81.3%) and bsmB (81.3%). The kpn gene encodes fimH-like pilus adhesin, and the ycfM gene encodes outer membrane lipoproteins. Both of them play an important role in adhesion and colony formation.49 The bsmB gene plays a key role in virulence factor production such as protease, serralysin, lipase and S-layer protein synthesis.50 Moreover, previous study confirmed that nitrogen sources in environment may affect the adhesion of S. marcescens.51 In our study, 68.8% of the S. marcescens isolates were positive for the allantoin metabolism-related gene allS, which helps pathogens compete for nitrogen sources with other bacteria.52 The high prevalence of the adhesion-related factors mentioned above in our study may provide strong abilities to attach both living tissues and non-living objects, a feature that enables S. marcescens to colonize within host tissues and participate in the first stage of infections.
Iron carriers are critical virulence determinants involved in bacterial pathogenesis, invasiveness, and molecular competition at the host pathogen interface.53 Meanwhile, the carriers enable pathogens to overcome the isolation of iron ions by the immune system of eukaryotic cells and capture metals.53 In this study, all S. marcescens isolates carried the metal-chelating siderophore enterobactin encoding gene entB. The phenolate-type siderophore yersiniabactin encoding genes ybtS (50.0%), irp-2 (37.5%), and fyuA (25.0%) were frequently observed in the tested isolates alone or in combination. Meanwhile, the iron-uptake system-related genes iutA and kfu were determined in 6.3% and 75.0% of the S. marcescens isolates, respectively. The iutA gene encoding the ferric aerobactin receptor involves typical ATP binding cassette (ABC)-type transporters.54 The ferric uptake transporter encoded by kfu mainly mediates the uptake of ferric iron.19 Previous study found that the highly virulent strains were more likely to carry the ferric ion carriers.19 High frequency of the genes associated with siderophore activation and the iron-uptake system imply that the S. marcescens isolates tested in this study may have a strong ability to capture ions, which is contributive to invade the mammary epithelial cells and induce bovine mastitis.
The traT gene, encoding a surface exclusion protein, is the most common determinant of virulence factors, and generally considered to be related to serum drug resistance.55,56 In this study, 37.5% of S. marcescens isolates carried the traT gene. To our knowledge, this gene was rarely reported in bovine S. marcescens. Prodigiosin is a prominent red pigment produced by S. marcescens and is essential for invasion, survival, and pathogenicity.18,57 We tested the prodigiosin biosynthesis-related genes and found the most frequent gene was pigP (81.3%), followed by pigC (37.5%). PigP is a positive regulator of prodigiosin production that regulates swarming and hemolysis through serratamolide production.16 Other studies also reported that the pigP was more prevalent in S. marcescens isolates than pigA and pigC.16,58
Hemolysin is a kind of exotoxin, which has hemolytic and cytotoxic activities and plays an important role in the occurrence or aggravation of mastitis.59,60 The hemolysin is encoded by shlA in S. marcescens. The outer membrane protein encoding gene shlB gene is responsible for secretion and activation of shlA.61 shlA/shlB system can secrete hemolysin under low iron conditions, and may make S. marcescens to use hemoglobin released by hemolysis as iron source.62 Therefore, the significant increase in hemolytic activity of hemolysin may transform the opportunistic pathogenic S. marcescens strain into a hypervirulent type.63 It is noteworthy that 25.0% and 75.0% of the tested isolates in this study carried both shlA and shlB, which may be potentially hypervirulent strains. In addition, the phlA gene is also considered to be related to hemolysis and cytotoxicity. phlA itself does not directly induce membrane instability of target cells. Phospholipase A encoded by phlA can catalyze the production of phospholipids, which lead to hemolysis and cell death.64 In this study, S. marcescens isolates also evaluated for the presence of toxin genes phlA. This is similar to the observed results of S. marcescens isolates in previous studies.17
Conclusion
In conclusion, although the prevalence of S. marcescens is low in our study, the high frequencies of phenotypic and genotypic resistance to cefazolin and chloramphenicol as well as the multidrug resistance remind the government to pay special attention to the antimicrobial agents used in dairy industry. Importantly, the emergence of carbapenem-resistant S. marcescens pose an alarming threat to public health due to the transmission of resistant determinants through the food chain. Furthermore, the high incidence of virulence genes mrkD, ycfM, pigP, bsmB, flhD, and phlA detected in S. marcescens suggested their pathogenic potential in bovine mastitis. Further investigations to be conducted to understand the pathogenicity of the individual virulent factor.
Ethics Approval and Informed Consent
The animal owners were informed about the purpose of the study and consent of each animal owner was obtained before the physical examination of cows for clinical mastitis and the collection of milk samples. All procedures involved in animal care and their use were in strict accordance with the guidelines for the Care and Use of Laboratory Animals, Lanzhou Institute of Husbandry and Pharmaceutical Sciences, CAAS, China (SYXK-2019-0012).
Acknowledgments
This work was financially supported by the Key Research and Development Program of Gansu Province (No. 21YF5NA141) and the National Key R&D Program of China during the 14th Five-year Plan Period (No.2022YFD1302101).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Yang F, Zhang S, Shang X, et al. Characteristics of quinolone-resistant Escherichia coli isolated from bovine mastitis in China. J Dairy Sci. 2018;101(7):6244–6252. doi:10.3168/jds.2017-14156
2. Zadoks R, Allore H, Barkema H, et al. Cow-and quarter-level risk factors for Streptococcus uberis and Staphylococcus aureus mastitis. J Dairy Sci. 2001;84(12):2649–2663. doi:10.3168/jds.S0022-0302(01)74719-4
3. Friman MJ, Eklund MH, Pitkälä AH, et al. Description of two Serratia marcescens associated mastitis outbreaks in Finnish dairy farms and a review of literature. Acta Vet Scand. 2019;61(1):1–11. doi:10.1186/s13028-019-0488-7
4. Tavares-Carreon F, De Anda-Mora K, Rojas-Barrera IC, Andrade A. Serratia marcescens antibiotic resistance mechanisms of an opportunistic pathogen: a literature review. PeerJ. 2023;11:e14399. doi:10.7717/peerj.14399
5. Iguchi A, Nagaya Y, Pradel E, et al. Genome evolution and plasticity of Serratia marcescens, an important multidrug-resistant nosocomial pathogen. Genome Biol Evol. 2014;6(8):2096–2110. doi:10.1093/gbe/evu160
6. González-Juarbe N, Mares CA, Hinojosa CA, et al. Requirement for Serratia marcescens cytolysin in a murine model of hemorrhagic pneumonia. Infect Immun. 2015;83(2):614–624. doi:10.1128/IAI.01822-14
7. Abbas HA, Hegazy WA. Repurposing anti-diabetic drug “Sitagliptin” as a novel virulence attenuating agent in Serratia marcescens. PLoS One. 2020;15(4):e0231625. doi:10.1371/journal.pone.0231625
8. Khayyat AN, Hegazy WA, Shaldam MA, et al. Xylitol inhibits growth and blocks virulence in Serratia marcescens. Microorganisms. 2021;9(5):1083. doi:10.3390/microorganisms9051083
9. Massé J, Dufour S, Archambault M. Characterization of Klebsiella isolates obtained from clinical mastitis cases in dairy cattle. J Dairy Sci. 2020;103(4):3392–3400. doi:10.3168/jds.2019-17324
10. Tezera M, Aman Ali E. Prevalence and associated risk factors of Bovine mastitis in dairy cows in and around Assosa town, Benishangul‐Gumuz Regional State, Western Ethiopia. Veter Med Sci. 2021;7(4):1280–1286. doi:10.1002/vms3.454
11. Magiorakos A-P, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. doi:10.1111/j.1469-0691.2011.03570.x
12. Yang F, Zhang SD, Shang XF, et al. Prevalence and characteristics of extended spectrum β-lactamase-producing Escherichia coli from bovine mastitis cases in China. J Integr Agric. 2018;17(006):1246–1251. doi:10.1016/S2095-3119(17)61830-6
13. Poirel L, Walsh TR, Cuvillier V, Nordmann P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn Microbiol Infect Dis. 2011;70(1):119–123. doi:10.1016/j.diagmicrobio.2010.12.002
14. Kutilova I, Valcek A, Papagiannitsis CC, et al. Carbapenemase-producing Gram-negative bacteria from American crows in the United States. Antimicrob Agents Chemother. 2020;65(1):e00586–00520. doi:10.1128/AAC.00586-20
15. Williams CT, Musicha P, Feasey NA, et al. ChloS-HRM, a novel assay to identify chloramphenicol-susceptible Escherichia coli and Klebsiella pneumoniae in Malawi. J Antimicrob Chemoth. 2019;74(5):1212–1217. doi:10.1093/jac/dky563
16. Ferreira RL, Rezende GS, Damas MSF, et al. Characterization of KPC-producing Serratia marcescens in an intensive care unit of a Brazilian tertiary hospital. Front Microbiol. 2020;11:956. doi:10.3389/fmicb.2020.00956
17. Aggarwal C, Paul S, Tripathi V, et al. Characterization of putative virulence factors of Serratia marcescens strain SEN for pathogenesis in Spodoptera litura. J Invertebr Pathol. 2017;143:115–123. doi:10.1016/j.jip.2016.12.004
18. Zhou J-W, Ruan L-Y, Chen H-J, et al. Inhibition of quorum sensing and virulence in Serratia marcescens by hordenine. J Agric Food Chem. 2019;67(3):784–795. doi:10.1021/acs.jafc.9b01005
19. Compain F, Babosan A, Brisse S, et al. Multiplex PCR for detection of seven virulence factors and K1/K2 capsular serotypes of Klebsiella pneumoniae. J Clin Microbiol. 2014;52(12):4377–4380. doi:10.1128/JCM.02316-14
20. Mamlouk K, Boutiba-Ben Boubaker I, Gautier V, et al. Emergence and outbreaks of CTX-M β-lactamase-producing Escherichia coli and Klebsiella pneumoniae strains in a Tunisian hospital. J Clin Microbiol. 2006;44(11):4049–4056. doi:10.1128/JCM.01076-06
21. Bachiri T, Bakour S, Lalaoui R, et al. Occurrence of carbapenemase-producing Enterobacteriaceae isolates in the wildlife: first report of OXA-48 in wild boars in Algeria. Microbial Drug Resist. 2018;24(3):337–345. doi:10.1089/mdr.2016.0323
22. Ballén V, Gabasa Y, Ratia C, et al. Antibiotic resistance and virulence profiles of Klebsiella pneumoniae strains isolated from different clinical sources. Front Cell Infect Microbiol. 2021:11. doi:10.3389/fcimb.2021.738223
23. Daoudi A, Benaoui F, El Idrissi Slitine N, et al. An outbreak of serratia marcescens in a Moroccan neonatal intensive care unit. Adv Med Sci. 2018;2018:1–4. doi:10.1155/2018/4867134
24. Todhunter D, Smith K, Hogan J. Serratia species isolated from bovine intramammary infections. J Dairy Sci. 1991;74(6):1860–1865. doi:10.3168/jds.S0022-0302(91)78351-3
25. Hinthong W, Pumipuntu N, Santajit S, et al. Detection and drug resistance profile of Escherichia coli from subclinical mastitis cows and water supply in dairy farms in Saraburi Province, Thailand. PeerJ. 2017;5:e3431. doi:10.7717/peerj.3431
26. Bi Y, Wang YJ, Qin Y, et al. Prevalence of bovine mastitis pathogens in bulk tank milk in China. PLoS One. 2016;11(5):e0155621. doi:10.7717/peerj.3431
27. Silva N, Guimarães F, Manzi M, et al. Molecular characterization and clonal diversity of methicillin-susceptible Staphylococcus aureus in milk of cows with mastitis in Brazil. J Dairy Sci. 2013;96(11):6856–6862. doi:10.3168/jds.2013-6719
28. Shirshikova TV, Sierra-Bakhshi CG, Kamaletdinova LK, et al. The ABC-type efflux pump MacAB is involved in protection of Serratia marcescens against aminoglycoside antibiotics, polymyxins, and oxidative stress. Msphere. 2021;6(2):e00033–00021. doi:10.1128/mSphere.00033-21
29. Hawkey S, Choy A. Serratia marcescens: a Rare Cause of Recurrent Implantable Cardioverter Defibrillator Site Infection. Clin Res Cardiol. 2015;2015:1–3. doi:10.1155/2015/641297
30. Ahmed AM, Shimamoto T. Molecular characterization of antimicrobial resistance in Gram‐negative bacteria isolated from bovine mastitis in Egypt. Microbiol Immunol. 2011;55(5):318–327. doi:10.1111/j.1348-0421.2011.00323.x
31. Ohnishi M, Sawada T, Hirose K, et al. Antimicrobial susceptibilities and bacteriological characteristics of bovine Pseudomonas aeruginosa and Serratia marcescens isolates from mastitis. Vet Microbiol. 2011;154(1–2):202–207. doi:10.1016/j.vetmic.2011.06.023
32. Kitazaki K, Koga S, Nagatoshi K, et al. In vitro synergistic activities of cefazolin and nisin A against mastitis pathogens. J Vet Med Sci. 2017:17–0180. doi:10.1292/jvms.17-0180
33. Swinkels J, Hilkens A, Zoche-Golob V, et al. Social influences on the duration of antibiotic treatment of clinical mastitis in dairy cows. J Dairy Sci. 2015;98(4):2369–2380. doi:10.3168/jds.2014-8488
34. Dos Santos FF, Mendonça LC, de Lima Reis DR, et al. Presence of mecA-positive multidrug-resistant Staphylococcus epidermidis in bovine milk samples in Brazil. J Dairy Sci. 2016;99(2):1374–1382. doi:10.3168/jds.2015-9931
35. Chen D, Xiao L, Hong D, et al. Epidemiology of resistance of carbapenemase‐producing Klebsiella pneumoniae to ceftazidime‐avibactam in a Chinese hospital. J Appl Microbiol. 2022;132(1):237–243. doi:10.1111/jam.15166
36. Yang F, Zhang S, Shang X, et al. Antimicrobial resistance and virulence genes of Enterococcus faecalis isolated from subclinical bovine mastitis cases in China. J Dairy Sci. 2019;102(1):140–144. doi:10.3168/jds.2018-14576
37. Cao Z, Xu J, Gao M, et al. Prevalence and antimicrobial resistance of Salmonella isolates from goose farms in Northeast China. Iran J Vet Res. 2020;21(4):287. doi:10.1186/2047-2994-4-s1-p140
38. Bischoff KM, White DG, Hume ME, et al. The chloramphenicol resistance gene cmlA is disseminated on transferable plasmids that confer multiple-drug resistance in swine Escherichia coli. FEMS Microbiol Lett. 2005;243(1):285–291. doi:10.1016/j.femsle.2004.12.017
39. Lee K, Yum JH, Yong D, et al. Novel acquired metallo-β-lactamase gene, bla SIM-1, in a class 1 integron from Acinetobacter baumannii clinical isolates from Korea. Antimicrob Agents Chemother. 2005;49(11):4485–4491. doi:10.1128/AAC.49.11.4485-4491.2005
40. Ghisalberti D, Masi M, Pagès J-M, Chevalier J. Chloramphenicol and expression of multidrug efflux pump in Enterobacter aerogenes. Biochem Biophys Res Commun. 2005;328(4):1113–1118. doi:10.1016/j.bbrc.2005.01.069
41. Liou B-H, Duh R-W, Lin Y-T, et al. A multicenter surveillance of antimicrobial resistance in Serratia marcescens in Taiwan. J Microbiol Immunol Infec. 2014;47(5):387–393. doi:10.1016/j.jmii.2013.04.003
42. Yoon HJ, Choi JY, Park YS, et al. Outbreaks of Serratia marcescens bacteriuria in a neurosurgical intensive care unit of a tertiary care teaching hospital: a clinical, epidemiologic, and laboratory perspective. Am J Infect Control. 2005;33(10):595–601. doi:10.1016/j.ajic.2005.01.010
43. Yin L, Zhang -P-P, Wang W, et al. 3-Phenylpropan-1-amine enhanced susceptibility of serratia marcescens to ofloxacin by occluding quorum sensing. Microbiol Spectr. 2022;10(5):e01829–01822. doi:10.1128/spectrum.01829-22
44. Alvarez-Ortega C, Olivares J, Martínez JL. RND multidrug efflux pumps: what are they good for? Front Microbiol. 2013;4:7. doi:10.3389/fmicb.2013.00007
45. Leitão JH. Microbial virulence factors. Int J Mol Sci. 2020;21(15):5320. doi:10.3390/ijms21155320
46. Shanks RM, Stella NA, Brothers KM, Polaski DM. Exploitation of a “hockey-puck” phenotype to identify pilus and biofilm regulators in Serratia marcescens through genetic analysis. Can J Microbiol. 2016;62(1):83–93. doi:10.1139/cjm-2015-0566
47. Srinivasan R, Devi KR, Kannappan A, et al. Piper betle and its bioactive metabolite phytol mitigates quorum sensing mediated virulence factors and biofilm of nosocomial pathogen Serratia marcescens in vitro. J Ethnopharmacol. 2016;193:592–603. doi:10.1016/j.jep.2016.10.017
48. Van Houdt R, Givskov M, Michiels CW. Quorum sensing in Serratia. FEMS Microbiol Rev. 2007;31(4):407–424. doi:10.1111/j.1574-6976.2007.00071.x
49. Marques C, Belas A, Aboim C, et al. Evidence of sharing of Klebsiella pneumoniae strains between healthy companion animals and cohabiting humans. J Clin Microbiol. 2019;57(6):e01537–01518. doi:10.1128/JCM.01537-18
50. Alexpandi R, Prasanth MI, Ravi AV, et al. Protective effect of neglected plant Diplocyclos palmatus on quorum sensing mediated infection of Serratia marcescens and UV-A induced photoaging in model Caenorhabditis elegans. J Photochem Photobiol B. 2019;201:111637. doi:10.1016/j.jphotobiol.2019.111637
51. Kalivoda EJ, Stella NA, O’Dee DM, et al. The Cyclic AMP-dependent catabolite repression system of serratia marcescens mediates biofilm formation through regulation of type 1 fimbriae. Appl Environ Microbiol. 2008;74(11):3461–3470. doi:10.1128/AEM.02733-07
52. Chou H-C, Lee C-Z, L-C M, et al. Isolation of a chromosomal region of Klebsiella pneumoniae associated with allantoin metabolism and liver infection. Infect Immun. 2004;72(7):3783–3792. doi:10.1128/IAI.72.7.3783-3792.2004
53. Klebba PE, Newton SM, Six DA, et al. Iron acquisition systems of gram-negative bacterial pathogens define TonB-dependent pathways to novel antibiotics. Chem Rev. 2021;121(9):5193–5239. doi:10.1021/acs.chemrev.0c01005
54. Weakland DR, Smith SN, Bell B, et al. The Serratia marcescens siderophore serratiochelin is necessary for full virulence during bloodstream infection. Infect Immun. 2020;88(8):e00117–00120. doi:10.1128/IAI.00117-20
55. Wenz J, Barrington G, Garry F, et al. Escherichia coli isolates’ serotypes, genotypes, and virulence genes and clinical coliform mastitis severity. J Dairy Sci. 2006;89(9):3408–3412. doi:10.3168/jds.S0022-0302(06)72377-3
56. Firoozeh F, Saffari M, Neamati F, Zibaei M. Detection of virulence genes in Escherichia coli isolated from patients with cystitis and pyelonephritis. Inter J Infec Dis. 2014;29:219–222. doi:10.1016/j.ijid.2014.03.1393
57. Liu GY, Nizet V. Color me bad: microbial pigments as virulence factors. Trends Microbiol. 2009;17(9):406–413. doi:10.1016/j.tim.2009.06.006
58. Abbas HA, Goda RM. Sotolon is a natural virulence mitigating agent in Serratia marcescens. Arch Microbiol. 2021;203:533–541. doi:10.1007/s00203-020-02039-y
59. Yang F, Zhang S, Shang X, et al. Detection and molecular characterization of methicillin-resistant Staphylococcus aureus isolated from subclinical bovine mastitis cases in China. J Dairy Sci. 2020;103(1):840–845. doi:10.3168/jds.2019-16317
60. Vallet-Gely I, Novikov A, Augusto L, et al. Association of hemolytic activity of Pseudomonas entomophila, a versatile soil bacterium, with cyclic lipopeptide production. Appl Environ Microbiol. 2010;76(3):910–921. doi:10.1128/AEM.02112-09
61. Calder A, Menkiti CJ, Çağdaş A, et al. Virulence genes and previously unexplored gene clusters in four commensal Neisseria spp. isolated from the human throat expand the neisserial gene repertoire. Microbial Genomic. 2020;6(9). doi:10.1099/mgen.0.000423
62. Schiebel E, Schwarz H, Braun V. Subcellular location and unique secretion of the hemolysin of Serratia marcescens. J Biol Chemistry. 1989;264(27):16311–16320. doi:10.1016/s0021-9258(18)71623-2
63. Lin C-S, Horng J-T, Yang C-H, et al. RssAB-FlhDC-ShlBA as a major pathogenesis pathway in Serratia marcescens. Infect Immun. 2010;78(11):4870–4881. doi:10.1128/IAI.00661-10
64. Shimuta K, Ohnishi M, Iyoda S, et al. The hemolytic and cytolytic activities of Serratia marcescens phospholipase A (PhlA) depend on lysophospholipid production by PhlA. BMC Microbiol. 2009;9:261. doi:10.1186/1471-2180-9-261
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.