")
Back to Journals » Cancer Management and Research » Volume 12
Pickled Vegetables Intake Impacts the Metabolites for Gastric Cancer
Authors Yang S , Lv Y , Wu C, Liu B, Shu Z, Lin Y
Received 8 July 2020
Accepted for publication 20 August 2020
Published 9 September 2020 Volume 2020:12 Pages 8263—8273
DOI https://doi.org/10.2147/CMAR.S271277
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eileen O'Reilly
Shuangfeng Yang,1,2 Yanping Lv,1,2 Chuancheng Wu,1,2,* Baoying Liu,1,2,* Zhixiong Shu,1,2 Yulan Lin1
1School of Public Health, Fujian Medical University, Fuzhou, People’s Republic of China; 2Fujian Provincial Key Laboratory of Environment Factors and Cancer, Fuzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Chuancheng Wu; Baoying Liu
School of Public Health, Fujian Medical University, No. 1 Xueyuan Road, Minhou, Fuzhou 350122, People’s Republic of China
Tel +86-18046043729
; Tel +81-0591-22862582
Email [email protected]; [email protected]
Purpose: An increased risk of gastric cancer (GC) for pickled vegetables intake has been suggested, but a complete understanding of its pathogenic origin is still lacking, especially from a metabolic viewpoint. We investigated the plasma metabolites and metabolic pathway alteration of GC related to pickled vegetables intake.
Methods: We analyzed plasma samples collected from 365 gastric cancer patients and 347 healthy individuals, and divided them into three subgroups according to the intake of pickled vegetables. Plasma samples were detected by untargeted metabolomics.
Results: Nine metabolites were significantly altered in GC patients among pickled vegetables intake groups (FDR P-value< 0.05). All of them were associated with the risk of gastric cancer adjusted for gender, age, smoking status, Helicobacter pylori infection. Pathway analysis showed significant alteration in the folate biosynthesis pathway.
Conclusion: In short, we provide new insights from a metabolic perspective on the relationship between pickled vegetables intake and the occurrence of gastric cancer.
Keywords: pickled vegetables intake, metabolites, metabolic pathway, gastric cancer
Introduction
Gastric cancer (GC) is the fifth most common cancer and the third leading cause of cancer-related death worldwide.1 However, the occurrence of GC varies from place to place, with “Eastern Asia Countries” like the People's Republic of China, Korea, and Japan experiencing the highest incidences while North America and Africa experience the lowest. Recent evidence shows that the GC pattern of most Chinese American immigrants is more consistent with their new country of residence than with their country of origin.2 Given this geographical variation, environmental and dietary factors may play more important roles than genetic factors in gastric oncogenesis. Therefore, there are growing interests in studying the impact of dietary changes on GC incidence.
Vegetable consumption is known to contribute to the reduction in GC risk.3 However, the incidence rates of GC remain high in Chinese, Japanese, and Korean populations, even though they have a high consumption of total vegetables.4,5 This may be due to the fact that they mainly consume pickled vegetables, which were the traditional and important part of their dietary culture. Several studies have shown that pickled vegetable intake could increase the risk of GC.4,6,7 It has been shown that the risk of GC for pickled vegetables intake is even higher than Helicobacter pylori (H. pylori) infection among Chinese populations (OR:1.49 [95% CI:1.10–2.01] vs 1.26 [95% CI: 1.12–1.62]).8 Although several mechanisms have been postulated in epidemiological studies,9–11 the molecular mechanism of pickled vegetable intake in gastric carcinogenesis remains unclear.
Metabolomics, as the end point of the signaling cascade, represents the final response of living systems to environmental, genetic, and disease factors, and is routinely applied as a tool for biomarker discovery.12–14 For example, Lario et al15 found that histidine, tryptophan and phenylacetylglutamine could possibly be used as potential GC biomarkers between non-GC and GC groups.15 The correlation between metabolomics and phenotypes enables metabolites to easily model pathophysiological changes in the molecular level in cancer. Due to innovative developments in bioinformatics and analytical technologies, metabolomics analysis is now expanded to provide insight into the mechanisms that underlie various physiological conditions and aberrant processes.16 For example, metabolomics was used to further study the potential molecular mechanism of gastric cancer caused by H. pylori.17 Therefore, metabolomics is a great tool to study the underlying molecular mechanism of gastric cancer caused by pickled vegetables intake.
In this study, we set out to examine the potential effects of pickled vegetables intake by measuring a broad range of blood metabolites and examined their correlation with gastric cancer. We applied untargeted metabolomics to plasma samples obtained from 365 GC patients and 347 healthy individuals.
Methods
Study Population
This study was based on a population-based case–control study, in which GC patients and healthy individuals were consecutively enrolled in Xianyou County between March 2013 and December 2017. All newly diagnosed cases of GC which were confirmed histologically based on tissue specimens and who had lived in Xianyou for at least 10 years were included. GC patients with other cancers, secondary or recurrent gastric cancer, gastritis, previously received neo-adjuvant chemotherapy or chemoradiotherapy or radiotherapy, pregnant, metabolic diseases such as diabetes, gout, hyperlipidemia, systemic administration of corticosteroids, neurological and psychiatric diseases, severe hepatic and renal dysfunction; and severe respiratory disease requiring continuous oxygen treatment, etc, were excluded. For the healthy individuals, there was no abnormality of blood tests and endoscopic examinations. A total of 365 GC cases and 347 healthy controls were recruited, and based on the median intake of pickled vegetables of subjects 10 years ago, all of them were divided into three subgroups: the group of no pickled vegetable intake 10 years ago (NPVI = 0g/week) (62 GC cases and 76 healthy controls), the group of low level pickled vegetable intake 10 years ago (LPVI ≤350 g/week) (133 GC cases and 110 healthy control) and the group of high level pickled vegetable intake 10 years ago (HPVI >350 g/week) (170 GC cases and 161 healthy controls). This study was approved by the Biomedical Research Ethics Committee of Fujian Medical University, China (no.97,2014), in line with the relevant regulations and requirements of medical ethics. Written consents were obtained from all participants at study enrollment.
Questionnaire Data Collection
Face-to-face interviews were conducted by trained investigators. A structured questionnaire was applied to collect information on demographic features, lifestyles, family cancer history, cigarette smoking and alcohol consumption, which has been described previously.8 It is noteworthy that a food frequency questionnaire (FFQ) was also used to obtain participants’ dietary habits and food intake, including the frequency and amount 10 years before diagnosis in the GC cases and 10 years before interview in the controls. For each kind of food, participants were asked to describe how often (per day, per week and per month) and the serving size they had. We made an estimation of average pickled vegetable intake per week for each subject, expressed as the product of frequency per week and usual serving size in grams. A commonly used serving bowl and some food pictures were presented to help subjects recall their past consumption of specific food when investigators collected dietary information. The study was introduced to the participants as a diet and health study in order to reduce recall bias.
Sample Collection and Preparation
All the study participants were in an overnight fasting state and 5 mL of peripheral venous blood was taken in the morning. The blood was then allowed to clot for 30 minutes at 37°C water bath and followed by centrifugation at 3000 rpm for 10 minutes, then was divided into plasma, white blood cells, and red blood cells and stored at −80°C until further analysis.
Plasma samples were thawed on ice, 300 μL of cold acetonitrile were added to 150 μL of plasma, vortexed and kept at 4°C for 30 min, 150 μL of cold acetonitrile were added again and vortexed and kept at 4°C for 3 hours, for protein precipitation. After centrifugation at 16,000 g (30 min, 4°C) and 500 μL supernatant filtration with 0.22 μm filter, 100–150 μL resulting supernatant were transferred to liquid chromatography-mass spectrometry (LC-MS) vials and stored at −20°C until subsequent LC-MS analyses. Quality control (QC) was prepared with standard plasma sample.
Metabolic Profiling
Each sample was processed by Agilent 1200 high performance liquid chromatography combined with a 6520 accurate electrospray ionization/quadrupole-time-of-flight mass system (Agilent Technologies, CA, USA). Plasma samples were separated on an Eclipse Plus C18 column (2.1 x 50 mm, 1.8 μm, Agilent Technologies), with the condition of 150 μL injection volume, 0.25 mL/min flow rate and 37°C column temperature, by using a gradient program of the mobile phase A 0.1% formic acid solution (ESI+)/water (ESI-) and mobile phase B was acetonitrile with 0.1% formic acid solution (ESI+)/acetonitrile (Merck, Darmstadt, Germany) (ESI-). The gradient program started from 2% B for 0–9 minutes, linearly increased from 2 to 60% B for 9–18 minutes, linearly increased from 60 to 100% B for 18–20 minutes, and kept at 100% B for 20–30 minutes. To avoid cross contamination from plasma samples, plusing an ultra-pure water elution after each sample. All the data were collected in ionization quadrupole time-of-flight mass spectrometry with both positive (ESI+) and negative (ESI-) full scan mode. The parameters of mass spectrometry were as following: GAS1, 80; GAS2, 70; CUR, 40; TEM, 375°C; and ISVF, 5500 V; a mass range of 50 to 1500; scan time of 0.3 seconds.
Data Processing and Statistical Analysis
mzData files were converted to the mzXML format using Agilent MassHunter qualitative analysis software and processed using XCMS online (https://xcmsonline.scripps.edu), which performed feature detection, retention time correction, alignment, annotation, etc. Metabolic peaks detected less than 80% in all QC samples were discarded.18 Identification of metabolites was carried out by matching m/z (Da) values against the Human Metabolome Database (HMDB, http://www.hmdb.ca).
Prior to statistical analysis, each metabolic peak in all subject samples was normalized based on QC samples for removing the unwanted analytical variations occurring intra- and inter-batches. And the plasma abundant values of metabolites investigated were set to a log scale and auto-scaled (mean-centered and divided by the standard deviation of each variable) using MetaboAnalyst 4.0. An orthogonal partial least squares discriminant analysis (OPLS-DA) and univariate two-sided t-test were used for metabolic profile description and metabolic signature discovery between GC cases and healthy controls among the NPVI, LPVI and HPVI groups, respectively. The false discovery rate (FDR) method was used to correct for multiple hypothesis testing and to reduce false positives. Those metabolic features with variable importance in the projection (VIP) values >1.0 in the OPLS-DA model and FDR adjusted p-values < 0.05 in the t-test were considered to be significantly different between GC cases and healthy controls. Then, using a venn diagram we selected the discriminant metabolites associated with pickled vegetables intake. Metabolites in groups LPVI and HPVI but outside the overlap part with group NPVI were considered to be related to pickled vegetables intake. The boxplots were used for depiction of highly discriminant metabolites related to intake of pickled vegetables between GC cases and healthy controls in the LPVI and HPVI groups. Subsequently, odds ratio (OR) and 95% confidence intervals (95% CI) for the association between metabolites related to pickled vegetables intake and GC risk were determined by logistic regression. Finally, pathway analysis was run with MetaboAnalyst 4.0.
Results
Participant Characteristics
A total of 365 GC cases and 347 healthy controls were included, with a mean age of (65.15±9.03) years and (64.95±10.07) years, respectively. As shown in Table 1, there was no statistically significant difference in gender, age, smoking status, H. pylori infection, among the three subgroups (p>0.05), except a significant difference in gender among the LPVI group (p<0.05).
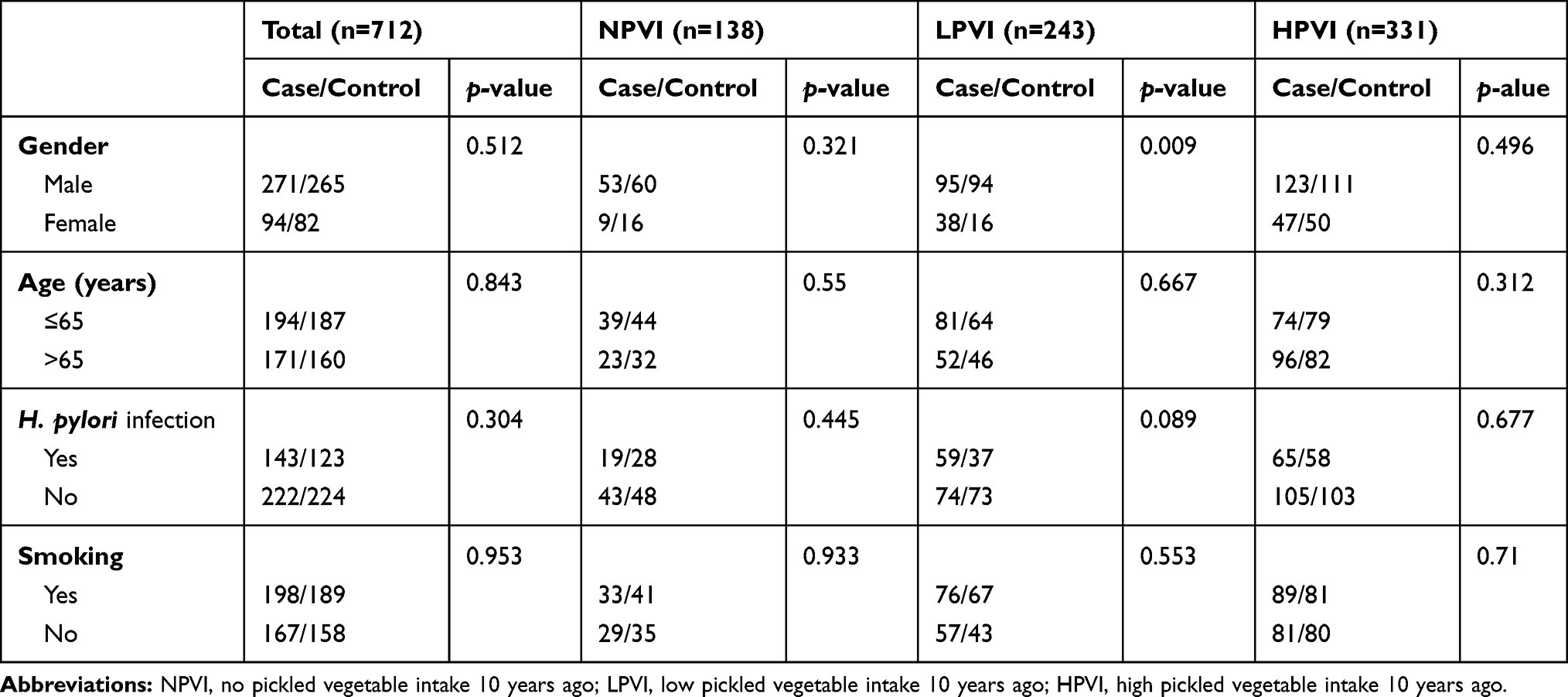 |
Table 1 Demographic Characteristics of Study Participants |
Metabolic Profiles of Plasma Samples
A total of 225 compounds were extracted from the LC-MS data. After being excluded based on Extracted Ion Chromatogram (EIC), 131 compounds were selected for subsequent analyses. After structurally identified by comparing with data shown in HMDB (www.hmdb.ca) databases, 51 compounds were identified from seven chemical classes, of which 14 were lipids, 17 were nucleotides, 4 were amino acids, 4 were peptides, 5 were carbohydrates/other, 5 were cofactors/vitamins, and 2 were energy pathway metabolites (Figure 1A). OPLS-DA score plots showed the differences between GC patients and healthy individuals in the three groups, suggesting variations in their metabolomes (Figure 1B–D).
 |
Figure 1 Metabolic profiles of plasma samples. (A) Total identified metabolites from plasma of GC cases and healthy controls. Orthogonal partial least-squares discrimination analysis (OPLS-DA) score plots in the groups NPVI (B), LPVI (C), and HPVI (D). |
Changes in Plasma Metabolites Associated with Pickled Vegetables Intake
To find metabolites associated with pickled vegetables intake, comparisons were made between GC cases and healthy controls among the NPVI, LPVI, and HPVI groups. First, univariate two-sided t-test and the variable importance in projection (VIP) scores from OPLS-DA models were carried out to identify different metabolites between GC patients and healthy individuals, of which 8 were different in the NPVI group, 14 in the LPVI group, and 14 in the HPVI group. Metabolites with FDR adjusted p-values < 0.05 and VIP >1 were selected as discriminants. Next, a venn diagram was used to find the important metabolites associated with pickled vegetables intake (Figure 2A). Eleven metabolites, which were in groups LPVI and HPVI but outside the overlapping part with group NPVI, were regarded as the changed metabolites related to pickled vegetables intake. They are glutathione, paraxanthine, L-histidinol, guanosine triphosphate, testosterone, N-acetylmuramate, porphobilinogen, guanosine, dihydrobiopterin, cytidine triphosphate, and 11-trans-leukotriene C4. Among them, the unique metabolites for groups LPVI and HPVI might be due to the intake of pickled vegetables. Figure 2B shows boxplots of relative abundance of changed metabolites related to pickled vegetable intake in the LPVI and HPVI groups. The change trend of metabolites related to pickled vegetable intake in the overlapping parts of LPVI and HPVI is the same, suggesting that the relative abundance of testosterone, dihydrobiopterin, and guanosine were increased in the GC group. On the contrary, guanosine triphosphate, N-acetylmuramate and porphobilinogen were decreased, compared with the control group. The difference of metabolites between GC cases and healthy controls is statistically significant (FDR p <0.05). To further certify that the metabolites change in plasma is related to the intake of pickled vegetables, we compared the relative abundance of plasma metabolites among subjects of NPVI, LPVI, and HPVI in the controls and the cases, respectively. Figure 2C shows that the changing trends of metabolites among NPVI, LPVI, and HPVI in the GC cases and the controls were similar. Only three metabolites (glutathione, paraxanthine, and 11-trans-leukotriene C4) were statistically significantly different between LPVI and HPVI in GC cases (p<0.05). However, except for porphobilinogen and L-histidinol, the relative abundance of plasma metabolites between NPVI and HPVI in cases or in controls was statistically different (p<0.05), suggesting that nine metabolites were associated with pickled vegetables intake.
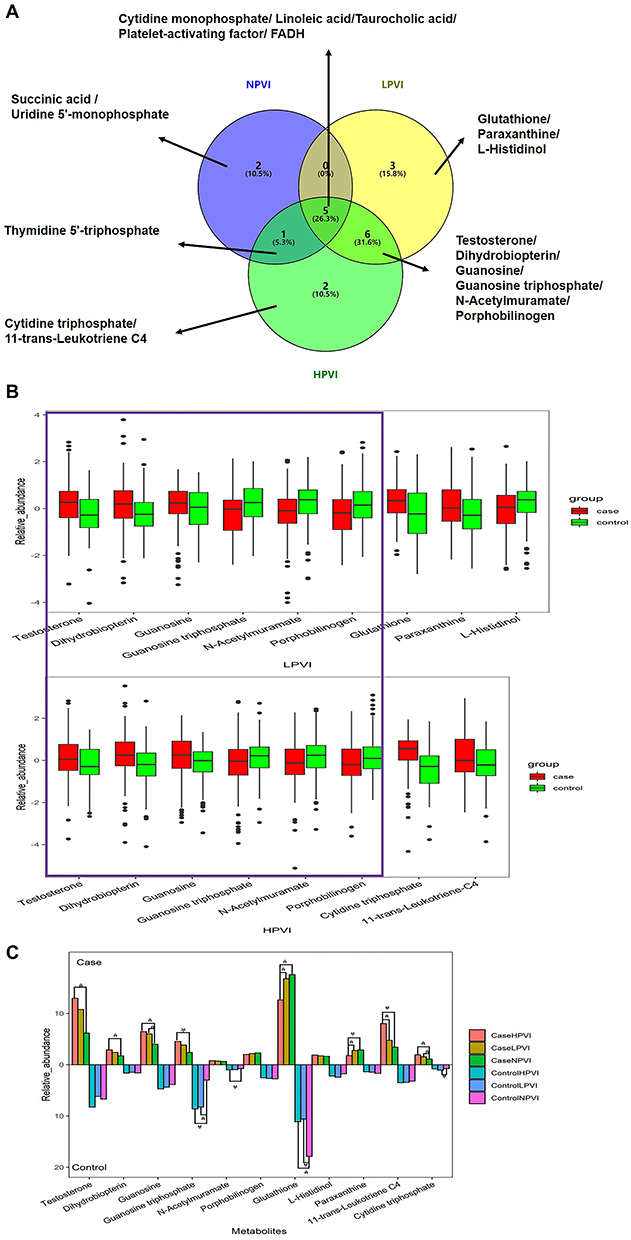 |
Figure 2 Discriminant metabolites. (A) Highly discriminant metabolites (VIP > 1, FDR adjusted t-test p-value<0.05) between GC cases and healthy controls in the groups NPVI, LPVI, and HPVI. (B) Relative abundance of highly discriminant metabolites between GC cases and healthy controls in the groups LPVI and HPVI. (C) Relative abundance of highly discriminant metabolites among NPVI, LPVI, and HPVI in the case and control. |
Association Between Metabolites Related to Pickled Vegetables Intake and Gastric Cancer
Figure 3 presents odds ratios for the association between metabolites related to pickled vegetables intake and GC. We presented the results in three groups: groups LPVI and HPVI were adjusted for gender. Total subjects including NPVI, LPVI and HPVI were adjusted for gender, age, smoking status, and H. pylori infection. Results from the three groups were similar, and the confidence intervals for the associations reported in the total subjects are notably narrower. Metabolites including testosterone, dihydrobiopterin, guanosine, paraxanthine, glutathione, cytidine triphosphate, and 11-trans-leukotriene C4, showed that their relative abundance was increased in the GC group, were associated with increased risks of GC. Conversely, metabolites including guanosine triphosphate and N-acetylmuramate, with a decreased relative abundance, were associated with a decreased risk of GC. In short, metabolites related to pickled vegetables intake are associated with the occurrence of gastric cancer.
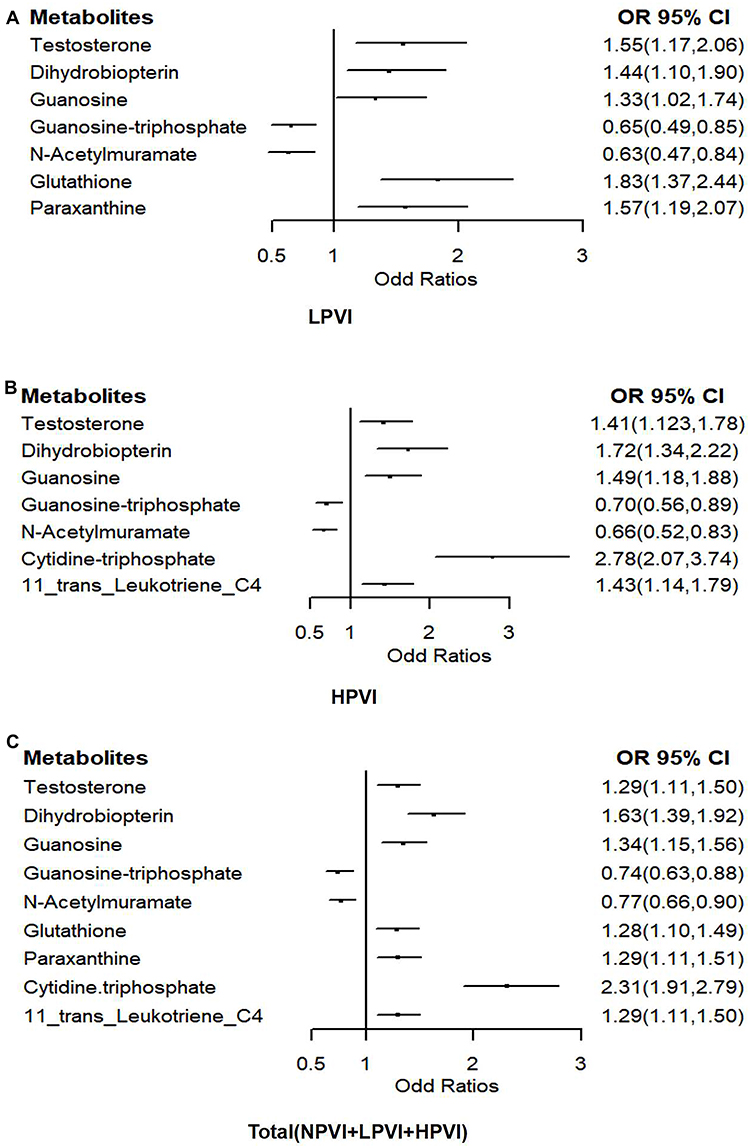 |
Figure 3 Odds ratios (OR) and 95% confidence intervals (CI) for metabolites related to pickled vegetables intake and the risk of gastric cancer (GC) in the groups LPVI (A), HPVI (B), and total (C). |
Pathway Analysis of Altered Metabolites Related to Pickled Vegetables Intake Between GC Cases and Healthy Controls
The nine altered metabolites related to pickled vegetables intake were pooled into pathway analysis, which was used to extract biological information within relevant networks of metabolic pathways integrating metabolite set enrichment and pathway topology analysis. Pathway enrichment and topology analysis were carried out using a global test and a relative-betweenness centrality measure. Results obtained are depicted in Figure 4A, where the color and the size of each circle indicate its p-value and pathway impact value, respectively. The folate biosynthesis pathway was found to be significantly altered (p<0.05). The relative abundance of two metabolites in the folate biosynthesis pathway quantified in plasma samples from GC cases and healthy controls are depicted in Figure 4B.
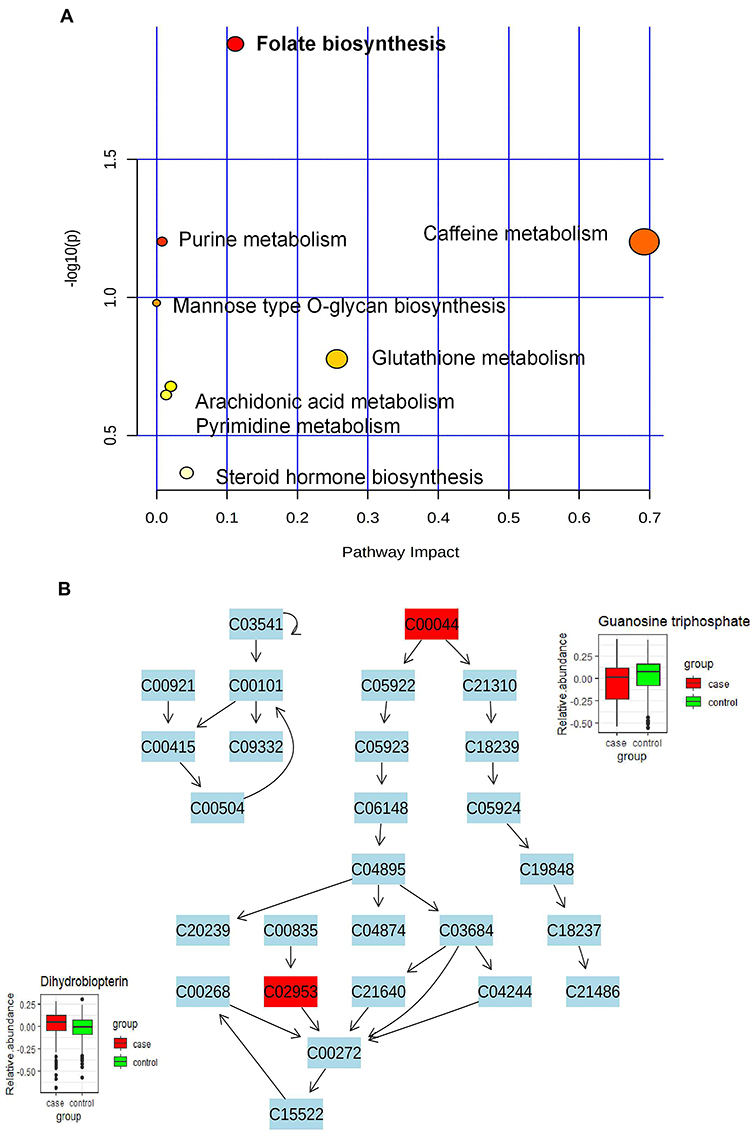 |
Figure 4 Pathway analysis. (A) Pathway analysis of the metabolites related to pickled vegetables intake. (B) Overview of differences in the relative abundance of detected metabolites between case and control in the folate biosynthesis pathway. |
Discussion
It has been reported that pickled vegetables intake increases the risk of gastric cancer,3,6,7,19 but its molecular mechanism in gastric carcinogenesis remains unclear. Metabolomics is now moving beyond biomarkers and towards mechanisms.16 In this study, we found that nine plasma metabolites were related to pickled vegetables intake, and all of them were associated with the occurrence of gastric cancer, including seven increased metabolites with increased risks of GC (testosterone, dihydrobiopterin, guanosine, paraxanthine, glutathione, cytidine triphosphate, 11-trans-leukotriene C4); and two decreased metabolites (guanosine triphosphate and N-acetylmuramate) with decreased risks of GC. Results from pathway analysis revealed folate biosynthesis pathway was significantly altered (p<0.05).
Folate metabolism is crucial for many biochemical processes, including purine and thymidine monophosphate (dTMP) biosynthesis, mitochondrial protein translation, and methionine regeneration.20 These biochemical processes in turn support critical cellular functions such as cell proliferation, mitochondrial respiration, and epigenetic regulation.20 Not surprisingly, abnormal folate metabolism has been causally linked with a myriad of diseases. For example, association between folate status and various forms of cancers has been reported.21–24 In this study, we found that the folate biosynthesis pathway was significantly altered in gastric cancer patients. The relative abundance of dihydrobiopterin (BH2) and guanosine triphosphate, the precursor of tetrahydrobiopterin (BH4) of folate biosynthesis, were increased and decreased in gastric cancer patients compared with the healthy controls among pickled vegetables intake groups, respectively. Tetrahydrobiopterin (BH4) is essential for the synthesis of nitric oxide (NO),25 and it is recognized as an essential step in initiating neoplastic transformation.26 A previous study demonstrated that the BH4:BH2 ratio is lower in tumor tissues. As a consequence, nitric oxide synthase activity generates more peroxynitrite and superoxide anion than nitric oxide, resulting in tumor growth and anti-apoptotic signaling.27 In addition, recent evidence suggests that decreasing level of BH4 will weaken the response of CD4 and CD8 T cells, thereby reducing their antitumor activity in vivo.28 Tetrahydrobiopterin (BH4) is synthesized by the combined action of three metabolic pathways, namely de novo synthesis, recycling, and salvage pathways.29 Guanosine triphosphate (GTP) is the substance of de novo synthesis, and reduced GTP will result in reduced BH4 synthesis. BH2 is the metabolic intermediates of BH4 in the salvage pathway catalyzed by dihydrofolate reductase. However, BH4 is easily oxidized to BH2. This explains why the BH2 level of GC cases is higher than that of the controls. Pickled vegetables were made with high concentrations of sodium chloride, which has been reported to induce mucosa and epithelial damage and produce chemical carcinogens–sodium nitrates and nitrites.6 We speculate that long-term intake of pickled vegetables might affect the activity of metabolic enzymes in the folate metabolism pathway, which affected BH4 levels and BH4:BH2 ratio and perturbs the folate metabolism pathway, resulting in increased production of peroxynitrite and weakened T cell responses, which ultimately leads to tumor cell proliferation and immune escape. Then tumor cells are in a state of rapid proliferation and differentiation causing nucleotide synthesis and metabolism to be upregulated significantly. For adequate energy generation, guanosine, cytidine triphosphate, and paraxanthine, were increased. Previous research results also showed that hypoxanthine and guanosine were also increased,30,31 in agreement with our result.
An increment of glutathione and 11-trans-leukotriene C4 in the GC patients among pickled vegetables intake groups are shown in Figure 2B. Glutathione is the principal intracellular antioxidant buffer against oxidative stress. It participates in many cellular metabolic activities including reactive oxygen species (ROS) removal, DNA and protein syntheses, and signal transduction.32 Cancer cells exhibit a greater ROS level than normal cells in order to meet the enhanced metabolism and proliferation. They have also developed an increased antioxidant defense system to cope with the higher oxidative state.33 Increased GSH level has been observed in different human cancer cells and is an important contributor to cancer pathology.34 11-trans-leukotriene C4 is the derivative of leukotriene, which is the major trigger of stress-induced oxidative DNA damage.35 It is well known that pickled vegetables contain exogenous sources of sodium nitrates and nitrites, which can react with amino acids in the stomach and form N-nitro compounds,6 resulting in oxidative damage.16
However, the reduction of N-acetylmuramate (a microbial cell wall component) is catalyzed by N-acetylglucosamine kinase, turning it into N-acetylmuramate 6-phosphate.36 This suggests that plasma metabolism may be affected by intestinal microbes alteration related to pickled vegetables intake. Previous research found that pickled vegetables produced a variety of flora during fermentation.37,38 Furthermore, it was reported that N-acetylmuramate was significantly decreased in the context of HLA-B27 expression and functionally modulate spondyloarthropathy in a rat model, which is associated with an altered intestinal microbiota and bowel inflammation.39
This study has some limitations. Firstly, data on pickled vegetable intake 10 years ago came from a retrospective survey, and there may be some recall bias. In addition, the detection of metabolites used non-target metabolomics analysis. Although more related metabolites can be found, the structure of some metabolites have not yet been characterized. Therefore, it is necessary to use target metabolomics to further validate specific metabolites. Finally, our metabolites were tested in plasma. Despite the fact that this can provide a starting point for finding links between diseases and metabolic pathways, it may not be the best sample for mechanistic research. Tissue analysis is perhaps more powerful for studying localized and specific responses to stimuli and pathogenesis.16 Metabolomics analysis using tissue and animal-based approaches as well as in vitro techniques will be carried out in future research.
Conclusion
In summary, we have reported that nine metabolites and the folate biosynthesis pathway were significantly altered in GC patients among the pickled vegetable intake groups. Our study provides new insights from a metabolic perspective on the relationship between pickled vegetable intake and the occurrence of gastric cancer.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. doi:10.3322/caac.21492
2. Liu Z, Lin C, Mu L, et al. The disparities in gastrointestinal cancer incidence among Chinese populations in Shanghai compared to Chinese immigrants and indigenous non-Hispanic white populations in Los Angeles, USA. Int J Cancer. 2020;146:329–340. doi:10.1002/ijc.32251
3. Huang L, Chen L, Gui Z-X, Liu S, Wei Z-J, Xu A-M. Preventable lifestyle and eating habits associated with gastric adenocarcinoma: a case-control study. J Cancer. 2020;11:1231–1239. doi:10.7150/jca.39023
4. Ren J-S, Kamangar F, Forman D, Islami F. Pickled food and risk of gastric cancer–a systematic review and meta-analysis of English and Chinese literature. Cancer Epidemiol Biomarkers Prev. 2012;21:905–915. doi:10.1158/1055-9965.EPI-12-0202
5. Kim HJ, Lim SY, Lee J-S, et al. Fresh and pickled vegetable consumption and gastric cancer in Japanese and Korean populations: a meta-analysis of observational studies. Cancer Sci. 2010;101:508–516. doi:10.1111/j.1349-7006.2009.01374.x
6. Yoo JY, Cho HJ, Moon S, et al. Pickled vegetable and salted fish intake and the risk of gastric cancer: two prospective cohort studies and a meta-analysis. Cancers. 2020;12:996. doi:10.3390/cancers12040996
7. Cai Q, Zhu C, Yuan Y, et al. Development and validation of a prediction rule for estimating gastric cancer risk in the Chinese high-risk population: a nationwide multicentre study. Gut. 2019;68:1576–1587. doi:10.1136/gutjnl-2018-317556
8. Lin Y, Wu C, Yan W, Guo S, Liu B. Five serum trace elements associated with risk of cardia and noncardia gastric cancer in a matched case-control study. Cancer Manag Res. 2020;12:4441–4451. doi:10.2147/CMAR.S250592
9. Iqbal A. Effect of food on causation and prevention of gastric cancer. J Cancer Prev Curr Res. 2017;8(5). doi:10.15406/jcpcr.2017.08.00289
10. Charnley G, Tannenbaum SR. Flow cytometric analysis of the effect of sodium chloride on gastric cancer risk in the rat. Cancer Res. 1985;45:5608–5616.
11. Furihata C, Ohta H, Katsuyama T. Cause and effect between concentration-dependent tissue damage and temporary cell proliferation in rat stomach mucosa by NaCl, a stomach tumor promoter. Carcinogenesis. 1996;17:401–406. doi:10.1093/carcin/17.3.401
12. Campbell SL, Wellen KE. Metabolic signaling to the nucleus in cancer. Mol Cel. 2018;71:398–408. doi:10.1016/j.molcel.2018.07.015
13. Pavlova NN, Thompson CB. The emerging hallmarks of cancer metabolism. Cell Metab. 2016;23:27–47. doi:10.1016/j.cmet.2015.12.006
14. Vander Heiden MG, DeBerardinis RJ. Understanding the intersections between metabolism and cancer biology. Cell. 2017;168:657–669. doi:10.1016/j.cell.2016.12.039
15. Lario S, Ramírez-Lázaro MJ, Sanjuan-Herráez D, et al. Plasma sample based analysis of gastric cancer progression using targeted metabolomics. Sci Rep. 2017;7:17774. doi:10.1038/s41598-017-17921-x
16. Johnson CH, Ivanisevic J, Siuzdak G. Metabolomics: beyond biomarkers and towards mechanisms. Nat Rev Mol Cell Biol. 2016;17:451–459. doi:10.1038/nrm.2016.25
17. Matsunaga S, Nishiumi S, Tagawa R, Yoshida M. Alterations in metabolic pathways in gastric epithelial cells infected with Helicobacter pylori. Microb Pathog. 2018;124:122–129. doi:10.1016/j.micpath.2018.08.033
18. Dunn WB, Broadhurst D, Begley P, et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat Protoc. 2011;6:1060–1083. doi:10.1038/nprot.2011.335
19. Zou T-H, Zheng R-H, Gao Q-Y, et al. Factors affecting occurrence of gastric varioliform lesions: a case-control study. World J Gastroenterol. 2016;22:5228–5236. doi:10.3748/wjg.v22.i22.5228
20. Zheng Y, Cantley LC. Toward a better understanding of folate metabolism in health and disease. J Exp Med. 2019;216:253–266. doi:10.1084/jem.20181965
21. Kim W, Woo HD, Lee J, et al. Dietary folate, one-carbon metabolism-related genes, and gastric cancer risk in Korea. Mol Nutr Food Res. 2016;60:337–345. doi:10.1002/mnfr.201500384
22. Moazzen S, Dolatkhah R, Tabrizi JS, et al. Folic acid intake and folate status and colorectal cancer risk: a systematic review and meta-analysis. Clin Nutr. 2018;37:1926–1934. doi:10.1016/j.clnu.2017.10.010
23. Tio M, Andrici J, Cox MR, Eslick GD. Folate intake and the risk of prostate cancer: a systematic review and meta-analysis. Prostate Cancer Prostatic Dis. 2014;17:213–219. doi:10.1038/pcan.2014.16
24. Ni Y, Du J, Yin X, Lu M. Folate intake, serum folate, and risk of esophageal cancer: a systematic review and dose-response meta-analysis. Eur J Cancer Prev. 2019;28:173–180. doi:10.1097/CEJ.0000000000000441
25. Rabender CS, Alam A, Sundaresan G, et al. The role of nitric oxide synthase uncoupling in tumor progression. Mol Cancer Res. 2015;13:1034–1043. doi:10.1158/1541-7786.MCR-15-0057-T
26. Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev. 2007;87:315–424.
27. Chittiboyina S, Chen Z, Chiorean EG, Kamendulis LM, Hocevar BA, Batra SK. The role of the folate pathway in pancreatic cancer risk. PLoS One. 2018;13:e0193298. doi:10.1371/journal.pone.0193298
28. Cronin SJF, Seehus C, Weidinger A, et al. The metabolite BH4 controls T cell proliferation in autoimmunity and cancer. Nature. 2018;563:564–568. doi:10.1038/s41586-018-0701-2
29. Latini A, de Bortoli da Silva L, da Luz Scheffer D, et al. Tetrahydrobiopterin improves hippocampal nitric oxide-linked long-term memory. Mol Genet Metab. 2018;125:104–111. doi:10.1016/j.ymgme.2018.06.003
30. Hirayama A, Kami K, Sugimoto M, et al. Quantitative metabolome profiling of colon and stomach cancer microenvironment by capillary electrophoresis time-of-flight mass spectrometry. Cancer Res. 2009;69(11):4918–4925. doi:10.1158/0008-5472.CAN-08-4806
31. Wu H, Xue R, Tang Z, et al. Metabolomic investigation of gastric cancer tissue using gas chromatography/mass spectrometry. Anal Bioanal Chem. 2010;396:1385–1395. doi:10.1007/s00216-009-3317-4
32. Wu G, Fang Y-Z, Yang S, Lupton JR, Turner ND. Glutathione metabolism and its implications for health. J Nutr. 2004;134:489–492. doi:10.1093/jn/134.3.489
33. Lv H, Zhen C, Liu J, Yang P, Hu L, Shang P. Unraveling the potential role of glutathione in multiple forms of cell death in cancer therapy. Oxid Med Cell Longev. 2019;2019:3150145. doi:10.1155/2019/3150145
34. Go Y-M, Jones DP. Redox theory of aging: implications for health and disease. Clin Sci (Lond). 2017;131:1669–1688. doi:10.1042/CS20160897
35. Dvash E, Har-Tal M, Barak S, Meir O, Rubinstein M. Leukotriene C4 is the major trigger of stress-induced oxidative DNA damage. Nat Commun. 2015;6:10112. doi:10.1038/ncomms10112
36. Veiga-da-Cunha M, Sokolova T, Opperdoes F, Van Schaftingen E. Evolution of vertebrate glucokinase regulatory protein from a bacterial N-acetylmuramate 6-phosphate etherase. Biochem J. 2009;423:323–332. doi:10.1042/BJ20090986
37. Chang JY, Chang HC. Improvements in the quality and shelf life of kimchi by fermentation with the induced bacteriocin-producing strain, Leuconostoc citreum GJ7 as a starter. J Food Sci. 2010;75:M103–110. doi:10.1111/j.1750-3841.2009.01486.x
38. Nam Y-D, Chang H-W, Kim K-H, Roh SW, Bae J-W. Metatranscriptome analysis of lactic acid bacteria during kimchi fermentation with genome-probing microarrays. Int J Food Microbiol. 2009;130:140–146. doi:10.1016/j.ijfoodmicro.2009.01.007
39. Asquith M, Davin S, Stauffer P, et al. Intestinal metabolites are profoundly altered in the context of HLA-B27 expression and functionally modulate disease in a rat model of spondyloarthritis. Arthritis Rheumatol. 2017;69:1984–1995. doi:10.1002/art.40183
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.