")
Back to Archived Journals » Research and Reports in Biology » Volume 8
Photoperiodic control of diapause induction in the zygaenid moth Thyrassia penangae – involving day-length measurement
Authors He H, Chen C , Xiao H, Xue F
Received 1 November 2016
Accepted for publication 5 December 2016
Published 12 January 2017 Volume 2017:8 Pages 1—5
DOI https://doi.org/10.2147/RRB.S126279
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Muhammad Mukhtar
Hai-Min He, Chao Chen, Hai-Jun Xiao, Fang-Sen Xue
Institute of Entomology, Jiangxi Agricultural University, Nanchang, People’s Republic of China
Abstract: The zygaenid moth Thyrassia penangae is a typical long-day species, entering winter diapause as a prepupa in a cocoon in response to short-day conditions during autumn. The previous photoperiodic responses under non-24-h light–dark cycle and Nanda–Hamner experiments (greatly extended dark periods coupled with a short photophase or a long photophase) revealed that the moth measured day length rather than night length in the determination of diapause. In this study, we further extend non-24-h light–dark cycle experiments and use the Bünsow experiment (the extended scotophase or photophase interrupted by 1-h light pulse or 1-h dark interruption) to test the photoperiodism for diapause induction of this moth. The extended non-24-h light–dark cycle experiment showed that when the length of photophase exceeded critical day length, diapause incidence was quite low regardless of scotophase duration. No circadian rhythmicity was found in Bünsow experiments. The incidence of diapause was consistently low when preinterruption photophase was longer than the critical day length. The two experiments provide further evidence that this moth measures day length instead of night length.
Keywords: Thyrassia penangae, diapause, photoperiodism, day-length measurement, Bünsow experiment
Introduction
A wide diversity of organisms use photoperiod (day length) as an environmental cue to predict the changing seasons and to time their life-history events, such as dormancy and migration.1 Photoperiodic induction of diapause is widespread in insects.2 The relative importance of the light (L) and dark (D) components of the daily cycle has been investigated in a number of insects by independently varying L and D in overall cycle lengths close to 24 h in duration. In most reported cases, the duration of the night is more important than that of the day in diapause determination.3 In the European corn borer Ostrinia nubilalis (Guenée), for example, Beck4 studied the effects of 10, 12, and 14 h of light in combination with a wide range of scotophases and of 10, 12, and 14 h of dark in combination with a wide range of photophases. The incidence of larval diapause was the highest (>90%) in cycles containing 10–14 h of scotophase, indicating that the duration of the scotophase was far more important than that of the photophase. The central importance of night length was also seen in data for the flesh fly Sarcophaga argyrostoma (Robineau-Desvoidy)5 in which the incidence of pupal diapause was very low in cycles containing a short night (eg, LD 12:8 and LD 16:8) but approached 100% in cycles containing a long night (eg, LD 12:12 and LD 16:12), regardless of the duration of the accompanying light component. Experiments with non-24-h light–dark cycles in which photophases of 10, 12, 14, and 16 h were combined with different lengths of scotophase of 4–24 h have been carried out on the zygaenid moth Pseudopidorus fasciata (Felder & Felder, 1862), the pine caterpillars Dendrolimus punctatus (Walker), the rice stem borer Chilo suppressalis (Walker), the Asian corn borer Ostrinia furnacalis, and the fall webworm Hyphantria cunea in our laboratory.6–10 The photoperiodic response curves in these species all showed that the incidence of larval diapause was low or zero in cycles containing a short night (4 and 8 h), but high in cycles containing a long night (12, 16, 20, and 24 h), regardless of the length of the photophase, indicating that the length of the scotophase plays an essential role in the determination of diapause in these species.
In a minority of cases, experiments using independently varying durations of L and D during the daily photocycle have suggested the operation of a timer measuring duration of the light, rather than the dark, component of the cycle.3 In the linden bug, Pyrrhocoris apterus Linnaeus, Saunders11 studied the effects of L held constant with varying lengths of D, or with D held constant and varying lengths of L. The results showed that the incidence of diapause was consistently low when L was >16 h (above the critical day length) regardless of scotophase duration, indicating that day-length measurement is more important than night-length measurement in this bug.
The zygaenid moth Thyrassia penangae (Moor) (Lepidoptera: Zygaenidae), an important pest of the Japanese cayratia Cayratia japonica (Thunb.), overwinters as a prepupa in a cocoon. This insect is a typical long-day species with a critical day length of 13.5 h (a 50% response) at 25°C, showing that the light period is the decisive phase for diapause determination in photoperiodic responses under non-24-h light–dark cycle and Nanda–Hamner12 experiments (see Figures 5 and 7 in the study by He et al).13 In this study, we further test the photoperiodism of diapause induction of the zygaenid moth T. penangae by using extended non-24-h light–dark cycle experiments and Bünsow14 experiment and provide further evidence that the moth measures day length rather than night length.
Materials and methods
Experimental materials and insect rearing conditions
Full-grown larvae, prior to producing a cocoon, were collected on the Jiangxi Agricultural University campus (28°46′N, 115°50′E), Jiangxi Province, People’s Republic of China. They were transferred to plastic boxes (diameter 16 cm; height 10 cm) to form cocoons. When adults emerged, they were transferred to wooden cages with nectar for mating and oviposition. The eggs were collected in incubators. Newly hatched larvae were transferred to plastic boxes (14, 9, and 4.5 cm) containing tender growing terminals or squares of C. japonica. Each box contained at least 50 individuals unless otherwise noted. The larvae were exposed to a variety of experimental light regimes at 25°C±1°C until prepupation. All experiments were performed in illuminated incubators equipped with four fluorescent 30 W tubes with an irradiance of ~1.97 Wm−2. Prepupae that did not enter diapause generally emerged as adults within 9–10 days at 25°C. Thus, each prepupa that did not emerge with 15 days was considered to be in diapause. In fact, nondiapause prepupae can pupate within 2 days.
Photoperiodic responses under non-24-h light–dark cycles
A previous non-24-h light–dark cycle experiment was extended to a greater range to test whether the light component is more important than the dark in the determination of diapause, in which larvae were exposed to light cycles with constant scotophases of 6, 8, 10, 12, 14, and 16 h at 25°C and photophases varied from 4 to 24 h, or constant photophases of 8, 10, 12, 14, 16, and 18 h were combined with scotophases that varied from 4 to 24 h (see Figure 1, 8 panels from the earlier study by He et al).13
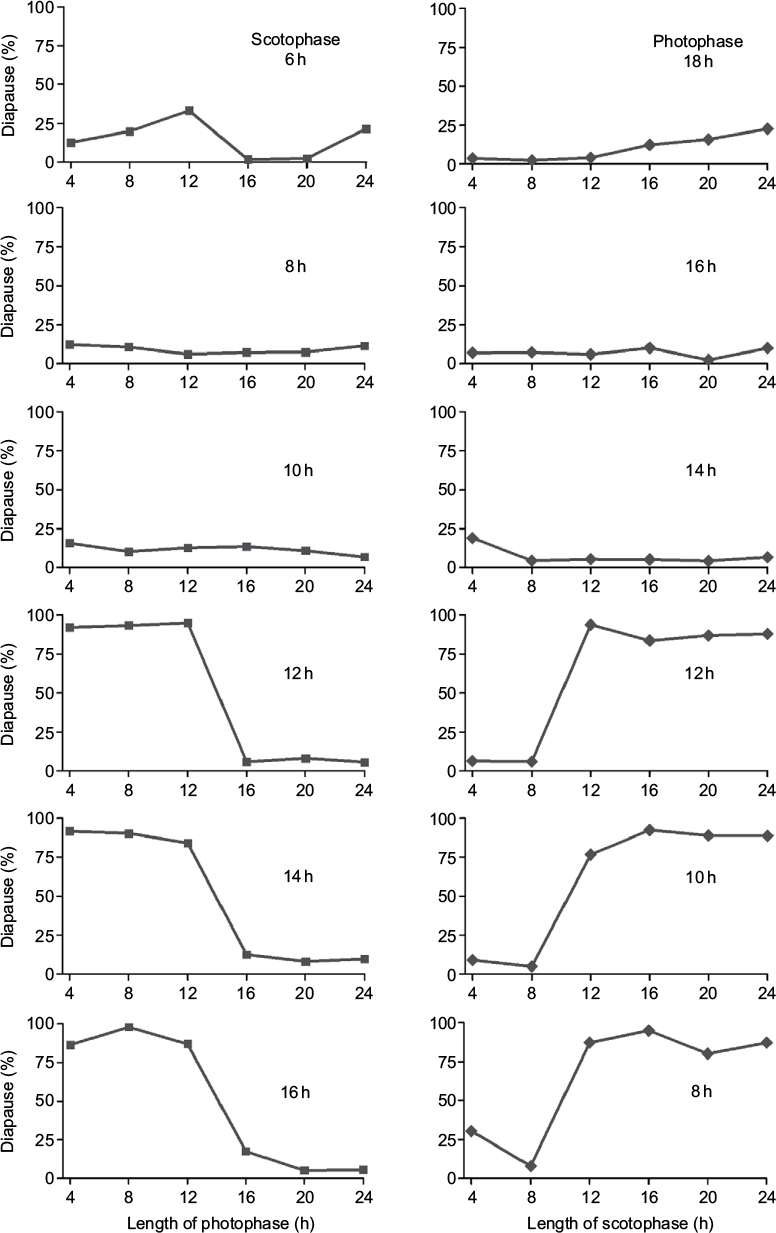 | Figure 1 Incidence of diapause in Thyrassia penangae under non-24 h photoperiodic cycles at 25°C. Left hand panels: scotophase held constant at 6, 8, 10, 12, 14, and 16 h, and photophase varied. Right hand panels: photophase held constant at 18, 16, 14, 12, 10, and 8 h and scotophase varied (8 panels from the earlier study by He et al13). n=32–67 for each point. |
Bünsow experiments
In Bünsow experiments, the constant photophase of 12 h was combined with a scotophase of 48 h, which was systematically interrupted by a 1-h light pulse, or the constant scotophase of 12 h was combined with a photophase of 48 h, which was systematically interrupted by a 1-h dark interruption.
Results
The experiment under non-24-h light–dark cycles shows that when D was held constant (Figure 1, left hand panels) diapause was consistently low regardless of scotophase duration when L exceeded the critical day length but the diapause incidence was consistently high when long scotophases of 12, 14, and 16 h were combined with short photophases of 4, 8, and 12 h. However, when L was held constant (Figure 1, right hand panels) at 14, 16, and 18 h (above the critical day length) diapause incidence was low (≤25%) regardless of scotophase duration. This experimental result indicates that day-length measurement is more important than night-length measurement in this moth. If night length was most relevant then high diapause would be expected when a long scotophase is combined with a long photophase. Interestingly, when scotophases were 4 and 8 h, almost all individuals developed without diapause, regardless of the length of the photophase. This may be explained that a short scotophase is read by the measurement system as a “long day” regardless of the length of the photophase.
No circadian rhythmicity was found in Bünsow experiments (Table 1).14 The incidence of diapause was high (>60%) when the preinterruption scotophase separated by a 1-h light pulse was longer than the critical night length (10.5 h); whereas the incidence of diapause was low (<18%) regardless of postinterruption scotophase when the preinterruption scotophase was shorter than the critical night length or close to the critical night length (Table 1, left). The incidence of diapause was low (<23%) when the preinterruption photophase separated by a 1-h dark interruption was longer than the critical day length; whereas diapause incidence was high (>65%) when the preinterruption photophase was shorter than the critical day length (13.5 h; Table 1, right). The result indicates that the length of the photophase plays a more important role in the determination of diapause than length of the scotophase.
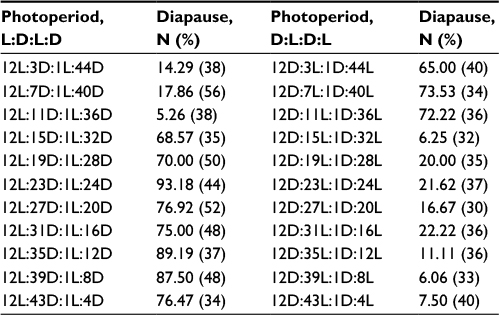 | Table 1 Diapause incidence of Thyrassia penangae in Bünsow experiments at 25°C, in which a constant photophase of 12 h was combined with a scotophase of 48 h which was systematically scanned by a 1-h light pulse or in which a constant scotophase of 12 h was combined with a photophase of 48 h which was systematically scanned by a 1-h dark interruption Abbreviations: D, dark; L, light. |
It is worth mentioning that diapause incidences were quite low (5.26%) when the moths were exposed to a short-day 12-h photoperiod, followed by an 11-h scotophase (which exceeds the critical night length). This may be because the 1-h light pulse just falls into a photoinducible phase (the late in the scotophase within a 24-h cycle), and thus results in a low incidence of diapause. According to the previous night-interruption experiment in 24 photoperiods, 1-h light pulse placed 3 h before lights-on inhibited diapause most effectively (92.7% development; Figure 6 in the study by He et al).13
Discussion
To our knowledge, this zygaenid moth provides the best example that the photoperiodic clock in some species measures duration of the light component of the light–dark cycle rather than the night. Although Saunders11 initially reported that the photoperiodic clock of P. apterus also measures day length rather than night length, he more recently suggested that night length could likely be measured if brighter light were used in his experiment because the linden bug is a diurnal species and may have a low sensitivity to light.3 In fact, the zygaenid moth is also a diurnal species with strong activity in full sunlight.13 However, the light intensity used in our experiments is ~1.97 Wm−2 and is sufficiently high to start photoperiodic response in this moth. If the system for measuring photoperiodic time differs between diurnal and nocturnal species, the Bünning’s hypothesis,15 suggesting a causal connection between circadian rhythms and photoperiodism, is possibly not suitable to explain photoperiodic induction of diapause in this zygaenid moth.
Whether the ability of animals to use day length as a cue for the timing of seasonal events (photoperiodism) is related to the endogenous circadian clock that regulates the timing of daily events has been debated for over 70 years.3,16 There remains no conclusive evidence defining the photoperiodic control of diapause in insects; in fact, there is an intense controversy regarding Bünning’s hypothesis.17–20 The genetic and metabolic components of photoperiodic responses vary among insect species, and diapause appears to have evolved independently numerous times, thus, we anticipate that not all photoperiodic clocks of insects will interact with circadian clocks in the same fashion.21
Our previous experiments on photoperiodic responses under non-24-h light–dark cycle revealed that when the length of photophase exceeded the critical day length (13.5 h), diapause was inhibited effectively regardless of the duration of the accompanying dark component (Figure 5 in the study by He et al)13. The previous Nanda–Hamner experiments showed that all light–dark cycles from LD 12:12 to LD 12:72 resulted in a short-day response and induced diapause, while all light–dark cycles from LD 14:4 to LD 14:72 resulted in a long day response and induced development (Figure 7 in the study by He et al).13 The present extended non-24-h light–dark cycle experiment further implicates day-length measurement in the determination of diapause. In the present Bünsow experiments, diapause induction mainly depended on whether the length of the preinterruption scotophase or the length of preinterruption photophase exceeded the critical night length or the critical day length. If the preinterruption scotophase was longer than the critical night length, diapause was induced but if the preinterruption photophase was longer than the critical day length, diapause was prevented. Therefore, these results may rule out a circadian basis to diapause initiation in this species and strongly suggest that this species measures day length instead of night length.
Acknowledgments
We thank Professor David L Denlinger of Ohio State University for critical reading the first version of the manuscript and for his valuable and helpful comments. This work was financially supported by National Natural Science Foundation of People’s Republic of China (31560608).
Disclosure
The authors report no conflicts of interest in this work.
References
Bradshaw WE, Holzapfel CM. Evolution of animal photoperiodism. Ann Rev Ecol Evol S. 2007;38:1–25. | ||
Saunders DS. Insect Clocks. 3rd ed. Amsterdam, The Netherlands: Elsevier;2002. | ||
Saunders DS. Insect photoperiodism: measuring the night. J Insect Physiol. 2013;59(1):1–10. | ||
Beck SD. Photoperiodic induction of diapause in an insect. Biol Bull. 1962;122(1):1–12. | ||
Saunders DS. The photoperiodic clock in the flesh-fly, Sarcophaga argyrostoma. J Insect Physiol. 1973;19(10):1941–1954. | ||
Wei X, Xue F, Li AQ. Photoperiodic clock of diapause induction in Pseudopidorus fasciata (Lepidoptera: Zygaenidae). J Insect Physiol. 2001;47(12):1367–1375. | ||
Huang LL, Xue FS, Wang GH, Han RD, Ge F. Photoperiodic response of diapause induction in the pine caterpillar, Dendrolimus punctatus. Entomol Exp Appl. 2005;117(2):127–133. | ||
Xiao HJ, Mou FC, Zhu XF, Xue FS. Diapause induction, maintenance and termination in the rice stem borer Chilo suppressalis (Walker). J Insect Physiol. 2010;56(11):1558–1564. | ||
Yang HZ, Tu XY, Xia QW, He HM, Chen C, Xue FS. Photoperiodism of diapause induction and diapause termination in Ostrinia furnacalis (Lepidoptera: Crambidae). Entomol. Exp Appl. 2014;153(1):34–46. | ||
Chen C, Wei X, Xiao H, He H, Xia Q, Xue F. Diapause Induction and Termination in Hyphantria cunea (Drury) (Lepidoptera: Arctiinae). PLoS One. 2014;9(5):e98145. | ||
Saunders DS. Insect photoperiodism: the linden bug, Pyrrhocoris apterus, a species that measures daylength rather than nightlength. Experientia. 1987;43(8):935–937. | ||
Nanda K, Hamner KC. Studies on the nature of the endogenous rhythm affecting photoperiodic response of Biloxi soybean. Bot Gaz. 1958;120(1):14–25. | ||
He HM, Xian ZH, Huang F, Liu XP, Xue FS. Photoperiodism of diapause induction in Thyrassia penangae (Lepidoptera: Zygaenidae). J Insect Physiol. 2009;55(11):1003–1008. | ||
Bünsow RC. The circadian rhythm of photoperiodic responsiveness in Kalanchoe. Cold Spring Harb Symp Quant Biol. 1960;25:257–260. | ||
Bünning E. Die endogene Tagesrhythmik als Grundlage der Photoperiodischen Reaktion. Ber Deut Bot Ges. 1936;54:590–607. German. | ||
Koštál V. Insect photoperiodic calendar and circadian clock: independence, cooperation, or unity? J Insect Physiol. 2011;57(5):538–556. | ||
Veerman A. Photoperiodic time measurement in insects and mites: a critical evaluation of the oscillator-clock hypothesis. J Insect Physiol. 2001;47(10):1097–1109. | ||
Danks HV. How similar are daily and seasonal biological clocks? J Insect Physiol. 2005;51(6):609–619. | ||
Bradshaw WE, Holzapfel CM. What season is it anyway? Circadian tracking vs. photoperiodic anticipation in insects. J Biol Rhythms. 2010;25(3):155–165. | ||
Bradshaw WE, Emerson KJ, Holzapfel CM. Genetic correlations and the evolution of photoperiodic time measurement within a local population of the pitcher-plant mosquito, Wyeomyia smithii. Heredity (Edinb). 2012;108(5):473–479. | ||
Meuti ME, Denlinger DL. Evolutionary links between circadian clocks and photoperiodic diapause in insects. Integr Comp Biol. 2013;53(1):131–143. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.