")
Back to Journals » OncoTargets and Therapy » Volume 13
Pharmacological Inhibition of Necroptosis Promotes Human Breast Cancer Cell Proliferation and Metastasis
Authors Shen F, Pan X, Li M , Chen Y , Jiang Y , He J
Received 22 January 2020
Accepted for publication 25 March 2020
Published 16 April 2020 Volume 2020:13 Pages 3165—3176
DOI https://doi.org/10.2147/OTT.S246899
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Federico Perche
Feng Shen,1,* Xiangou Pan,2,* Min Li,3,* Yixing Chen,2 Ying Jiang,3 Jian He2
1Department of Medical Oncology, Zhongshan Hospital, Fudan University, Shanghai 200032, People’s Republic of China; 2Department of Radiation Oncology, Zhongshan Hospital, Fudan University, Shanghai 200032, People’s Republic of China; 3Department of General Surgery, Zhongshan Hospital, Fudan University, Shanghai 200032, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jian He
Department of Radiation Oncology, Zhongshan Hospital, Fudan University, Shanghai 200032, People’s Republic of China
Tel/ Fax +86-021- 64041990
Email [email protected]
Ying Jiang
Department of General Surgery, Zhongshan Hospital, Fudan University, Shanghai 200032, People’s Republic of China
Tel/Fax +86-21-64041990
Email [email protected]
Background: Breast cancer remains a great threat to females worldwide. As a recently defined programmed cell death pathway that associates with immune activation, RIP1/RIP3/MLKL necroptosis signaling has been implicated in a variety of diseases. The present study aimed to investigate the role of RIP1/RIP3/MLKL signaling in breast cancer cell proliferation and metastasis in vivo and in vitro.
Methods: Western blot and quantitative real-time PCR were performed to evaluate the activation of necroptosis signaling in clinical human breast cancer tissues. Correlation of necroptosis signaling markers with clinicopathological parameters was statistically assessed. Cell viability assay, colony formation assay, wound healing assay, and transwell migration and invasion assays were performed to investigate the effects of necroptosis inhibition on breast cancer cell proliferation and metastasis.
Results: Clinical breast cancer tissues showed significantly higher levels of tumor necrosis factor alpha (TNFα), RIP1, RIP3 and MLKL at both mRNA and protein levels as compared with their paired non-cancerous tissues. Phosphorylation of RIP3 and MLKL was also remarkably provoked. Statistics showed that both RIP1 and MLKL positively correlated with cancer parameters such as N-cadherin (p=0.002 for RIP1 and p=0.021 for MLKL) and Ki67 (p=0.031 for RIP1 and p=0.05 for MLKL). The MLKL expression level significantly correlated with tumor size (p=0.001) and the proliferation indicator Ki67 (p=0.018). In addition, pharmacological inhibition of the necroptosis signaling using necrostatin-1 promoted breast cancer cell proliferation and colony formation by approximately 50%. Blockade of necroptosis signaling also accelerated wound healing process and cell transmigration in breast cancer cells.
Conclusion: Our results suggested that pharmacological inhibition of necroptosis promoted breast cancer cell proliferation and metastasis. Modulation of tumor cell necroptosis might represent a novel strategy as to breast cancer treatment.
Keywords: breast cancer, necroptosis, RIP1, MLKL, proliferation, metastasis
Introduction
Breast cancer remains a great health threat to females worldwide with the incidence and mortality ranks only secondary to lung cancer.1 Over the last decades, the annual incidence of new breast cancer cases did not significantly slow down.1 Based on a recent statistic, approximately 252,710 new cases of invasive breast cancer and 40,610 breast cancer deaths have occurred among US women in 2017.2 A large-scale statistic showed that the death rate remained 40% higher in blacks (28.4 vs 20.3 deaths per 100,000) despite a lower incidence rate (126.7 vs 130.8); this disparity was magnified among black women aged <50 years, who had a death rate double that of whites. In the most recent 5-year period, the death rate declined in Hispanics (2.1% per year), blacks (1.5%), whites (1.0%), and Asians/Pacific Islanders (0.8%) but was stable in American Indians/Alaska Natives.3 Although various treatments, such as surgical resection,4 adjuvant endocrine therapy5 and radiation therapy,6 have improved patients’ survival and reduced tumor recurrence, the tendency to metastasis remains a great threat to patients’ health. Karyotypic and epidemiological analyses of human tumors at different stages suggest that breast cancer turns increasingly aggressive via a stepwise accumulation of genetic changes, such as transcriptional regulation,7,8 translational regulation and posttranslational (ie, epigenetic) regulation.9 The high morbidity and mortality as well as its great tendency to metastasize to other organs mandate new biomarkers for both early diagnosis and treatment of breast cancer.
Over the last few years, the developments around cancer immunotherapy have led to a paradigm shift in the treatment of many different cancers.10 The ability of the immune system to fight against cancer cells is largely dependent on the number and function of immune cells. One of the determinants in this context is the extent of lymphocytic infiltration (termed as tumour-infiltrating lymphocytes). Evidence has shown that tumour-infiltrating lymphocytes can be linked with an increased therapeutic response in HER2-positive breast cancer and triple-negative breast cancer.11,12 In this setting, release of inflammation factors and the subsequent recruitment of immune cells play critical roles in promoting cancer immunotherapy. However, mechanisms of inflammation factors release remain largely unknown. Should the switch of immune cell recruitment be manually controlled, it would be beneficial to monitor the therapeutic efficacy of breast cancer.
Necroptosis is a regulated form of necrosis, with the dying cells rupturing and releasing intracellular components that can trigger an innate immune response.13 Necroptosis signaling is modulated by the receptor-interacting serine/threonine-protein kinase 1 (RIP1) and subsequently activates the kinase RIP3 and the pseudokinase mixed lineage kinase domain-like protein (MLKL).14 Upon binding of tumor necrosis factor alpha (TNFα) to TNF receptor 1 (TNFR1) on the cell surface, RIP1 was recruited to form a complex with other proteins such as RIP3 and MLKL, known as necrosome. This complex then phosphorylates MLKL, leading to its cytoplasm translocation and causing eventual pore formation in cellular membrane.15
Despite the broad involvement of necroptosis signaling in immune response, the detailed role of necroptosis in breast cancer cell proliferation and metastasis remains largely unknown. In the present study, we collected clinical human tissues and found that breast cancer malignancy associated well with the activation of necroptosis signaling. The MLKL expression significantly correlated with pathological parameters and conferred prognostic values. Pharmacological inhibition of necroptosis accelerated breast cancer cell proliferation and migration. Our study represented the first one, to the best knowledge of us, to show direct evidence that the necroptosis signaling was activated upon breast cancer malignancy and pharmacological modulation of tumor cell necroptosis might be a novel strategy as to breast cancer treatment.
Methods
Human Tissues
The use of human specimens was approved by the Ethic Review Board of Zhongshan Hospital, Fudan University (No. Y2019-010). For the histological study, slides were collected from a total of 50 breast cancer cases that were referred to Zhongshan Hospital from March 2019 to May 2019. For the investigation of molecular expression, a total of 10 breast cancer cases as well as their paired adjacent non-cancerous tissues were collected. All patients showed their full intention to participate in our study and their written informed consents were obtained. This study was performed in accordance with the Declaration of Helsinki. All efforts were made to preserve patients’ anonymity. The tumor tissues as well as their adjacent non-tumor tissues were frozen into liquid nitrogen as soon as dissection from patients during the surgeries for subsequent RNA and protein extracts or were fixed into 4% paraformaldehyde for histopathological assays. The clinical record from each patient was also obtained and the association of necroptosis with clinical characteristics was analyzed.
RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
Total RNAs from clinical patients were isolated by Trizol reagents (Invitrogen, NY, USA) according to the manufactures’ instructions. The quantity and quality of RNAs were assessed using Nanodrop 2000 (Thermo Scientific, NY, USA) by collecting the OD230nm and OD260nm. Afterwards, a total of 1μg RNA were reversely transcribed into cDNA with Reverse qPCR RT Master Mix (TaKaRa, Dalian, China). qRT-PCR assays were performed with SYBR Green qPCR Mix (TaKaRa) in an ABI 7900 Thermal Cycler System with the following procedure: 95°C for 2 min, followed by 35 cycles at 95°C for 10 s and 60°C for 30 s. Relative expressions of target genes were determined by the 2−ΔΔCT method. Target genes were normalized to GAPDH. The primers are listed in Table 1.
 |
Table 1 Primer Sequences Used in the Present Study |
Western Blot Analysis
Proteins extracted from human tissues and cultured MDA-MB-231 and MCF-7 cells were quantified by BCA methods (Thermo Scientifics, NY, USA). A total of 40 μg proteins were loaded onto a 10% SDS-PAGE gels. Electrophoresis was performed in the home-made running buffer, and afterwards, proteins were transferred into a 0.22 μm membrane (Millipore, USA) for 2 h with the voltage of 100 V. After blocking with 5% skim milk at room temperature for 1 h, the membranes were incubated with primary antibodies at 4°C overnight. The primary antibodies against RIP1 (ab106393, Abcam, NJ, USA), RIP3 (ab56164, Abcam), MLKL (ab184718, Abcam), β-actin (ab8227, Abcam) and GAPDH (ab181602, Abcam) were diluted into 1:1000 with tris-buffered saline containing 1‰ tween-20 (TBST). The secondary antibodies were purchased from Santa Cruz Biotech. (Santa Cruz, CA, USA). The chemiluminescence detection was performed with Supersignal West Femto Maximum Sensitivity Substrate (Thermo Scientific, CA, USA).
Hematoxylin and Eosin (H&E) Staining and Immunohistochemistry (IHC) Staining
Tissues from clinical patients were fixed, embedded in paraffin, sectioned at 4 μm slices and stained with H&E staining. For IHC staining, the slices were dewaxed with xylene and washed with 95% ethanol for 10 min. After washed with running water for 5 min at room temperature, the slices were put into 3% H2O2-methanol solution for 30 min at 37°C. For antigen retrieval, slides were microwaved in citrate buffer (PH=6.0) at 900 W for 2.5 min and then 150W for 9 min. After blocking, the slides were incubated with primary antibodies at 37°C for an hour and then at 4°C overnight. Afterwards, the secondary antibodies were incubated and chromogenic reaction was performed with an DAB reagent (ZSGB-Bio, Beijing, China). The images were captured under a Nikon microscope with five random fields of each slide. For the semiquantitative analysis, the staining score was calculated as staining intensity × positive percentage where staining intensity was classified as weak (score 1), mild (score 2) and intensive (score 3), and positive percentage was classified as very low positivity (<25% area being positive staining, score 1), low (26–50% area being positive area, score 2), medium (51–75% area being positive area, score 3) and high positivity (over 75% area being positive area, score 4). The final staining score was subtyped as low positivity (1≤final score ≤4), medium (5≤final score ≤8) and high positivity (9≤final score ≤12).
Cell Culture and siRNA Transfection
MDA-MB-231 and MCF-7 cells were purchased from American Type Culture Collection (ATCC, Massachusetts, USA) and cultured in dulbecco’s modified eagle medium (DMEM, Gibco, NY, USA) supplied with 10% fetal bovine serum (FBS, Gibco) without antibiotics. The culture medium was changed every other day, otherwise as stated. A specific siRNA against lysine demethylase 4A (KDM4A, also known as JMJD2A) was synthesized by GemoPharm Co. with the sequences as follows: sense, 5ʹ-GAGUUAUCAACUCAAGAUA-3ʹ and antisense, 5ʹ-UAUCUUGAGUUGAUAACUC-3ʹ. A scramble siRNA was used as negative control. Briefly, all siRNAs were diluted to 20 μmol/L with RNase-free water, and transiently transfected into cells with Lipofectamine 3000 (Invitrogen) as per the protocols. Transfection efficiency was verified prior to experiments.
Cell Viability Assays
MDA-MB-231 and MCF-7 cells in the presence or absence of KDM4A knockdown were seeded into 96-well plates and treated with Necrostatin-1 (Nec-1, 60 μM). Nec-1 was dissolved into DMSO at a concentration of 60 mM and the control cells were treated with the same dose of DMSO. Cells were then co-incubated with 10 µL CCK-8 reagent (Yisheng, Shanghai, China) for 1h in triplicate and the absorbance of each well was collected at 450nm. The experiments were continued for consecutive 4 days.
Colony Formation Assays
Both MDA-MB-231 and MCF-7 cells were seeded at 6-well plates in triplicate (100 cells/well) and pre-treated with or without Nec-1. After incubation for continuous 14 days at 37°C without replacing the culture medium, the plates were fixed with pre-iced methanol on ice and then stained with crystal violet (1%) for 5min at room temperature and washed with warm PBS twice. Colonies with more than 50 cells were considered as survivors and counted under a Nikon microscope (Japan).
Wound-Healing Assays
MDA-MB-231 and MCF-7 cells were seeded in six-well plates and treated with or without Nec-1 (60 μM). After 24 h of incubation, cells were washed with warm PBS twice and scraped a cross with 10µL sterile pipette tips in the center of every experimental well. The plates were washed with PBS again and photographed under a Nikon microscope at a magnification of 100×. The plates were put back into the 37°C incubator and cultured for additional 24 h. Afterwards, the cells were pictured again and the wound closure was calculated with five random fields.
Transwell Assays
MDA-MB-231 and MCF-7 cells were stimulated with or without Nec-1 for 24 h. Afterwards, cells were washed with PBS, trypsinized and a total of approximately 5,000 cells were re-suspended in FBS-free DMEM, seeded into the upper chamber (pore size: 8µm, Corning, NY, USA) and then 60 µM Nec-1 was mixed with the cells. Meanwhile, 600µL complete DMEM (10% FBS) were added into the lower chamber to avoid of bubbles. After incubation for 12 h, the chamber was washed with PBS, fixed with ice-cold methanol and stained with crystal violet (1%) at room temperature for 5min. Cells on the top surface were scraped with cotton swab and those attached on the lower surface of the chamber were photographed under the microscope with five random sights. As for the invasion assays, the chamber was pre-incubated with Matrigel (Corning, NY, USA) for 6h prior to experiments.
Statistical Analysis
Data were shown as mean ± standard deviation (SD). GraphPad 5.0 software (La Giolla, CA, USA) was involved for the analysis. The Student’s t-test was included to compare the differences between groups and one-way or two-way ANOVA were used for difference comparison among three groups when necessary. For correlation analysis of molecular expression levels, the linear regression models were performed using GraphPad Prism 5.0 and the goodness of fit for regression models was qualified using r2 values. Any value of p<0.05 was considered as statistical significance.
Results
Necroptosis Signaling Is Activated in Breast Cancer Tissues
Initially, to assess the expression of RIP1/RIP3/MLKL necroptosis signaling in breast cancer, we collected 10 surgical dissected breast cancer tissues. In 6 of the 10 breast cancer cases, the mRNA level of TNFα was more than 2-fold increased as compared with their adjacent non-cancerous tissues (Figure 1A), indicating the remarkable release of pro-inflammatory factors in breast cancer tissues. Moreover, RIP1 and MLKL, two core components of necroptosis signaling were consistently promoted at their mRNA levels (Figure 1B and C) and protein levels (Figure 1D) in cancerous tissues. In fact, the protein levels of RIP1 and MLKL were both remarkably elevated in cancerous tissues as compared with the paired adjacent tissues in all the tested 6 cases (Figure 1D). RIP3, a downstream of RIP1 also increased at both the total protein form and phosphorylated form. The phosphorylated level of MLKL (pMLKL) which is the executor of cell membrane rupture also significantly increased in the cancerous tissues relative to their counterparts (Figure 1D). To confirm the link between necroptosis and subsequent release of inflammation factor, we isolated the total RNAs from the 10 cases of breast cancer and it was found that the level of RIP1 significantly correlated with that of TNFα (p<0.0001, r2=0.899, Figure 1E). Likewise, the level of MLKL showed a good linear correlation with that of TNFα (p<0.0001, r2=0.884, Figure 1F). All these clinical investigations demonstrated the activation of necroptosis signaling in breast cancers.
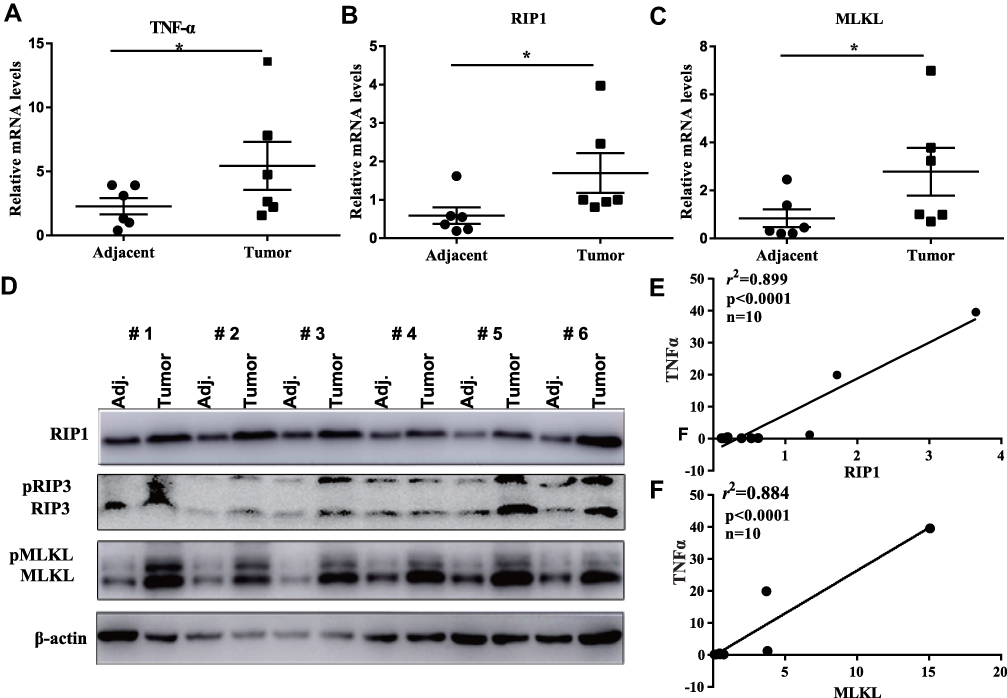 |
Figure 1 Necroptosis signaling is activated in breast cancer tissues. (A–C) qRT-PCR analysis of the pro-inflammatory factor TNFα and necroptosis signaling key components RIP1 and MLKL in 6 breast cancer tissues and their adjacent non-cancerous tissues. (D) In the 6 breast cancer cases, Western blotting was performed to detect the protein levels of necroptosis signaling and their phosphorylated levels. (E, F) after qRT-PCR analysis of RIP1, MLKL and TNFα, linear regression was performed to investigate the correlation of necroptosis signaling with the subsequent inflammation factors. Note: *p<0.05 as indicated. Abbreviations: TNFα, tumor necrosis factor alpha; RIP1, receptor-interacting serine/threonine-protein kinase 1; RIP3, receptor-interacting serine/threonine-protein kinase 3; MLKL, mixed lineage kinase domain-like protein; Adj, adjacent. |
Necroptosis Correlates with Tumor Malignancy Parameters
Cancer cell proliferation and epithelial-to-mesenchymal transition are markers of tumor aggressiveness. Next, the association of necroptosis with breast cancer aggressive markers was assessed. In the collected breast cancer tissues, it was found that MLKL correlated well with the mesenchymal marker N-cadherin (p=0.021, r2=0.617, Figure 2A) and with the tumor cell proliferation marker Ki67 (p=0.05, r2=0.443, Figure 2B). Another cell proliferation marker proliferative cell nuclear antigen (PCNA) also tended to correlate with MLKL (p=0.305, r2=0.149, Figure 2C) and the epithelial marker E-cadherin tended to negatively correlate with MLKL (p=0.589, r2=0.044, Figure 2D). Likewise, the expression levels of RIP1 showed significant positive correlation with that of N-cadherin (p=0.002, r2=0.754, Figure 2E) and Ki67 (p=0.031, r2=0.505, Figure 2F), though it failed to yield significant correlation with PCNA (p=0.290, r2=0.158, Figure 2G) and E-cadherin (p=0.845, r2=0.006, Figure 2H). These analyses suggested that the extent of necroptosis correlated well with breast cancer aggressiveness.
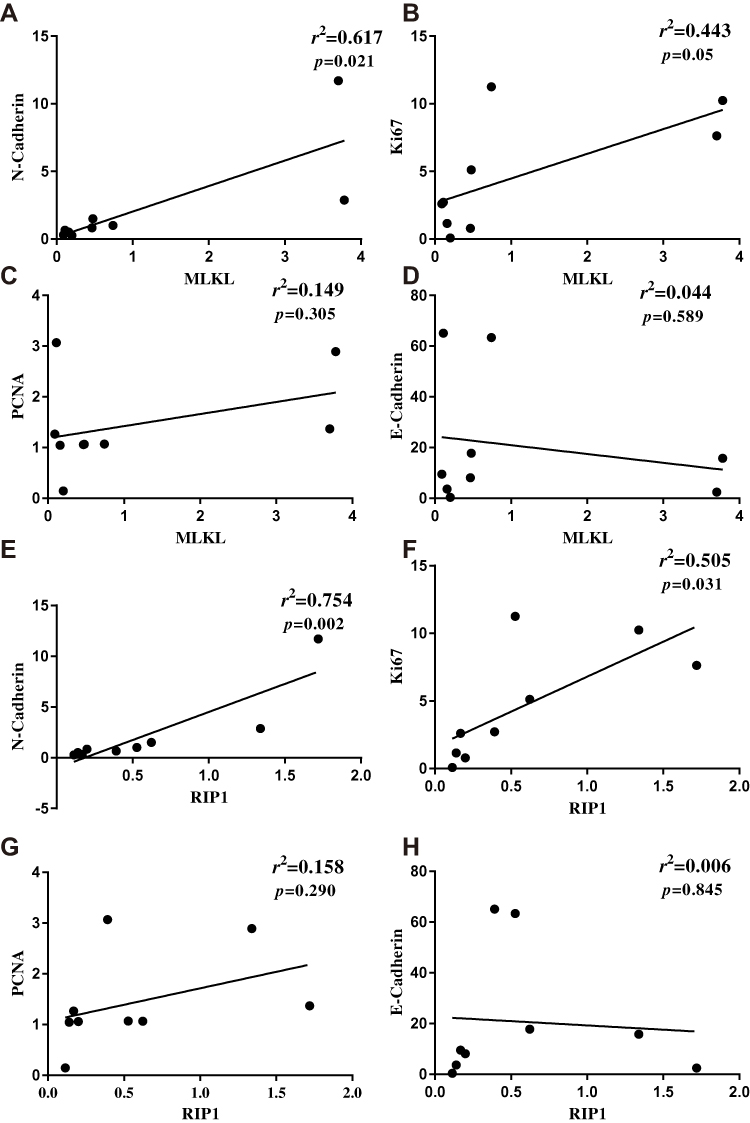 |
Figure 2 Extent of necroptosis significantly correlated with breast cancer aggressiveness. (A–D) After qRT-PCR analysis of MLKL and breast cancer pathological parameters N-cadherin, Ki67, PCNA and E-cadherin, linear regression analysis was performed to assess the association of necroptosis with cancer pathological parameters. (E–H) After qRT-PCR analysis of RIP1 and N-cadherin, Ki67, PCNA and E-cadherin, linear regression analysis was performed to assess the association of necroptosis with cancer pathological parameters. Abbreviations: RIP1, receptor-interacting serine/threonine-protein kinase 1; MLKL, mixed lineage kinase domain-like protein; PCNA, proliferative cell nuclear antigen. |
Necroptosis Activation Correlated Well with Tumor Size and Breast Cancer Malignancy Markers
Furthermore, 50 specimens from independent breast cancer patients were collected and subject to histopathological analysis. H&E analysis confirmed the aggressive property of these collected breast cancer specimens as compared with the adjacent normal tissues (Figure 3A). Immunohistochemistry analysis revealed the strong membrane-location of MLKL in breast cancer tissues (Figure 3B), suggesting the activation of necroptosis. Interestingly, unlike the fringe effect which shows intensive staining of molecules at the borderline of a slide, the staining of MLKL gradually intensified from the slide margin to the central zones (Figure 3B). This finding mirrored the stronger activation of necroptosis in central areas of a breast neoplasm. We then semiquantitatively analyzed the staining results of MLKL based on positive percentage and staining intensity. The representative images of MLKL staining in each category are shown in Figure 3C. It turned out that approximately one-third (32%) cases were MLKL-high staining (Figure 3D). Interestingly, the staining of MLKL did not correlate with patients’ age (p=0.615), TNM staging (p=0.296) or lymph node metastasis (p=0.091) but significantly associated with tumor size (p=0.001) (Table 2). The staining of MLKL also did not correlate with pathological parameters such as estrogen receptor alpha (ERα) (p=0.094), prostaglandin receptor (PR) (p=0.125) or human epithelial receptor 2 (HER2) (p=0.193) but significantly associated with Ki67 (p=0.018) (Table 3). In fact, the staining of MLKL showed comparable intensity between early stage and late stage of breast cancers (p=0.17, Figure 3E) and significantly positively correlated with Ki67 by linear regression analysis (p=0.0001, r2=0.27, Figure 3F). These data suggested that necroptosis activation correlated well with tumor size and breast cancer malignancy markers.
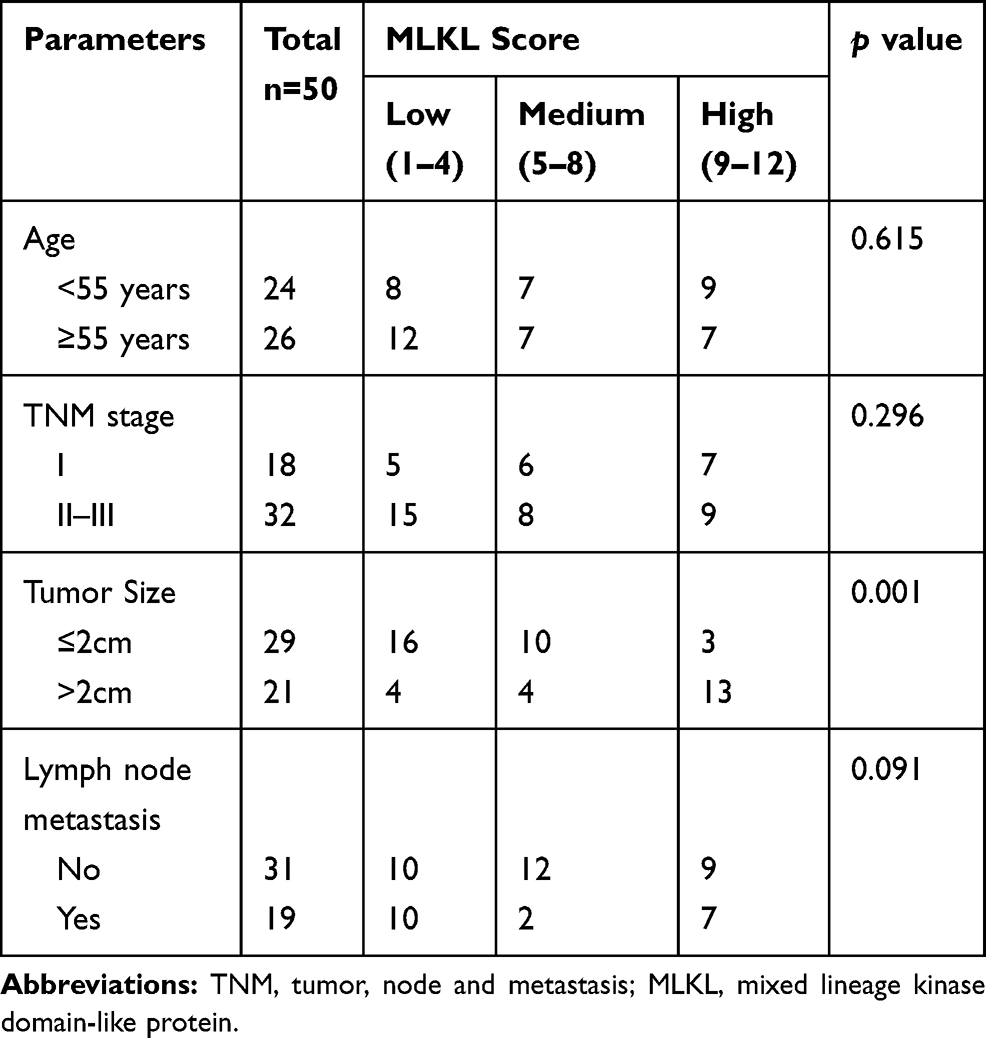 |
Table 2 Association Between Necroptosis Signaling with Clinical Parameters |
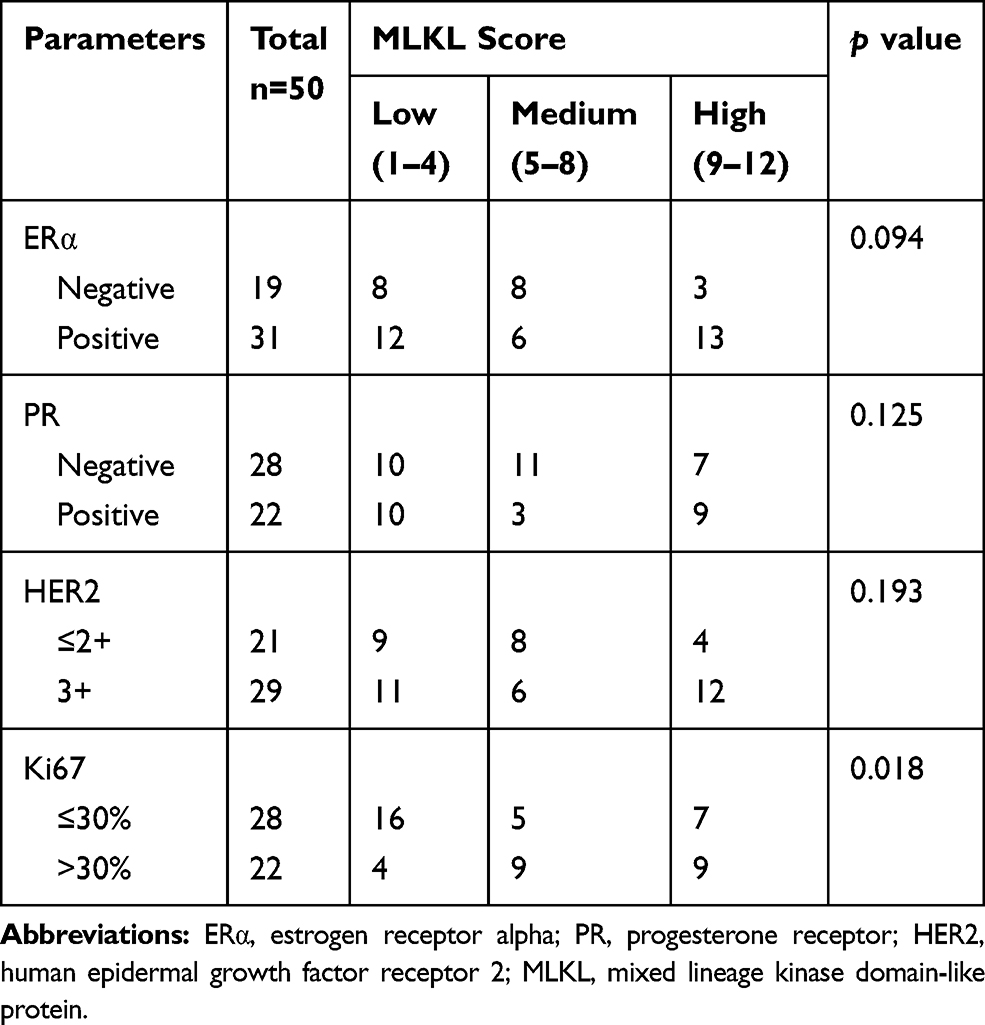 |
Table 3 Association Between Necroptosis Signaling with Pathological Parameters |
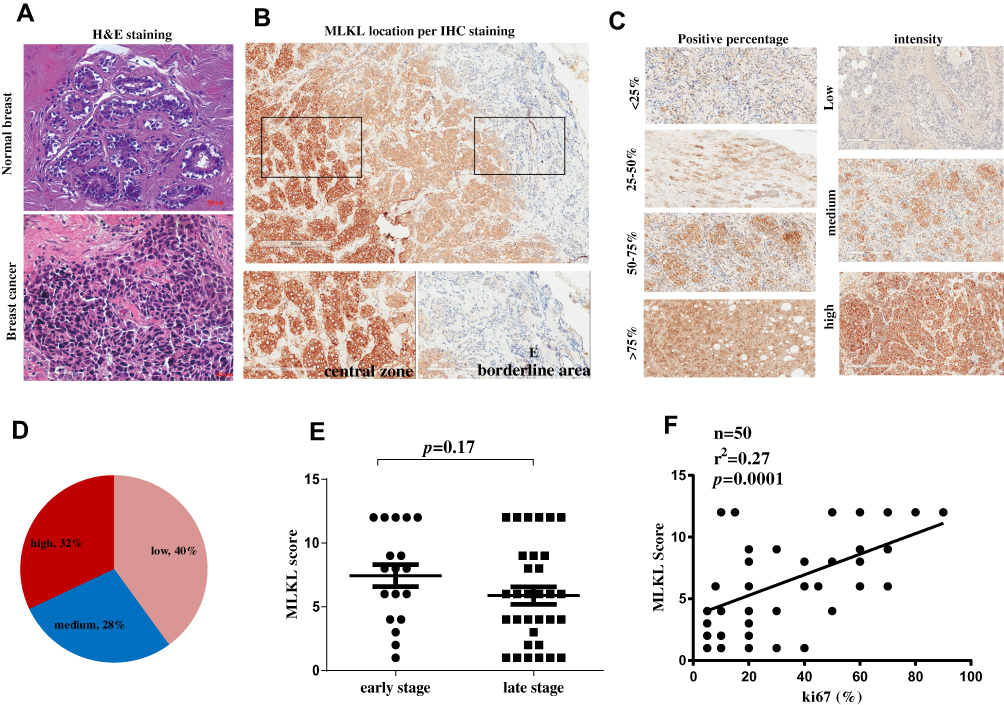 |
Figure 3 Necroptosis activation is an independent predictive factor for breast cancer malignancy. (A) H&E staining of a breast cancer tissue and a normal breast tissue. (B) immunohistochemistry analysis of MLKL in breast cancer tissues. The subcellular location of MLKL was visualized and its staining intensity in central zone and borderline areas were outlined and magnified, respectively. (C) Representative images of MLKL staining scores. (D) After scoring of MLKL staining, MLKL was classified as low expression, medium expression and high expression. (E, F) The correlation of MLKL expression scores with breast cancer stages and Ki67. Abbreviations: MLKL, mixed lineage kinase domain-like protein; H&E, hematoxylin and eosin staining; IHC, immunohistochemistry. |
Blockade of Necroptosis Signaling Promotes Breast Cancer Cell Proliferation and Colony Formation
Effects of necroptosis signaling blockade on breast cancer cell malignant capacities were then assessed. It was found that pharmacological inhibition of necroptosis using necrostatin-1 (Nec-1) significantly promoted cell proliferation in both ERα-negative MDA-MB-231 cells (p=0.03, Figure 4A) ERα-positive MCF-7 cells (p<0.001, Figure 4B), further validating that necroptosis signaling is a critical controller of tumor cell fate irrespective of ERα subtype. In the colony formation assay, the clonogenic capacities were visibly enhanced after 24 hours’ treatment of Nec-1 (Figure 4C) and counting of the colonies showed that Nec-1 increased the colonies by up to 50% in both cell lines (p=0.006 for MDA-MB-231 cells and p=0.043 for MCF-7 cells, Figure 4D). KDM4A is a well-documented factor that contributes to breast cancer cell proliferation16,17 and knockdown of KDM4A confers tumor protection.18 To further verify the promotion effect by Nec-1, we depleted the expression of KDM4A in breast cancer cells and found that the KDM4A-mediated promotion of RIP1 and MLKL was blunted by Nec-1 treatments (Figure 4E). More importantly, KDM4A siRNA-conferred retardation of cell proliferation was restored by Nec-1 treatments in MDA-MB-231 cells (p=0.03, Figure 4F) and MCF-7 cells (p=0.016, Figure 4G). These results suggested that inactivation of necroptosis signaling promoted breast cancer proliferation and colony formation.
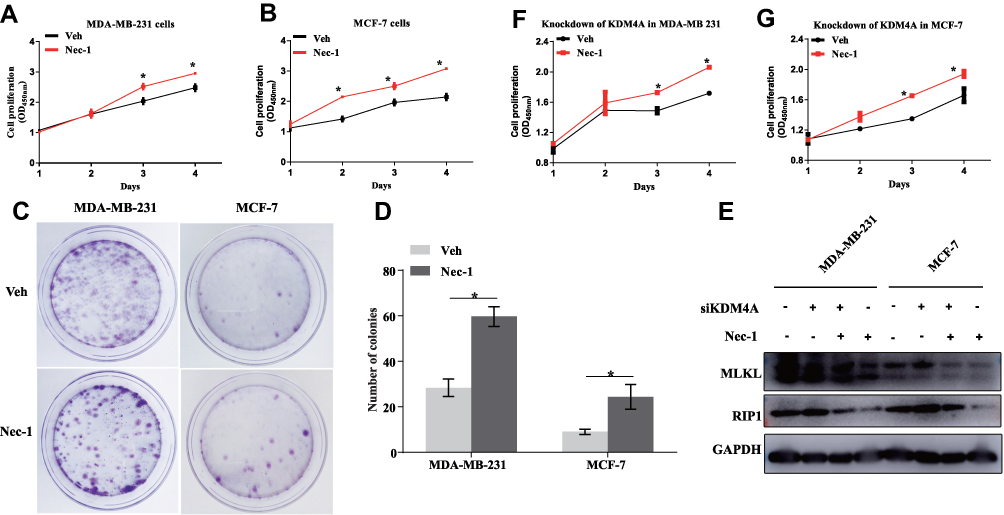 |
Figure 4 Inactivation of necroptosis signaling promotes breast cancer proliferation and colony formation. (A, B) MDA-MB-231 cells and MCF-7 cells with or without Nec-1 treatments were subject to Cell counting kit-8 (cck-8) assay. (C, D) MDA-MB-231 cells and MCF-7 cells with or without Nec-1 treatments were subject to colony formation assay. (E) Western blotting detection of RIP1 and MLKL protein expression at indicated treatments. (F, G) KDM4A-depleted MDA-MB-231 cells and MCF-7 cells were subject to Cell counting kit-8 (cck-8) assay after pretreatments with Nec-1 or a vehicle control. Note: *p<0.05 as indicated. Abbreviations: KDM4A, lysine demethylase 4A; Nec-1, necrostatin-1; RIP1, receptor-interacting serine/threonine-protein kinase 1; MLKL, mixed lineage kinase domain-like protein; OD 450nm, optical density at the wavelength of 450 nm; Veh, vehicle; GAPDH, glyceraldehyde-3-phosphate dehydrogenase. |
Inactivation of Necroptosis Signaling Promotes Wound Recovery in Breast Cancer Cell Lines
Next, the effects of necroptosis signaling on cell recovery of wounds were assessed. In MCF-7 cells, after 36 hours of healing, control MCF-7 cells only recovered 42% of the wound which was in great contrast to Nec-1 treated cells that closed over 70% of the wound (p=0.001, Figure 5A). In MDA-MB-231 cells, after 24-hour free recovery, vehicle-treated cells recovered approximately 62% of the wound but Nec-1-treated cells almost fully closed the wound (p=0.001, Figure 5B). These data suggested that inhibition of necroptosis signaling in breast cancer cells favored high pervasion.
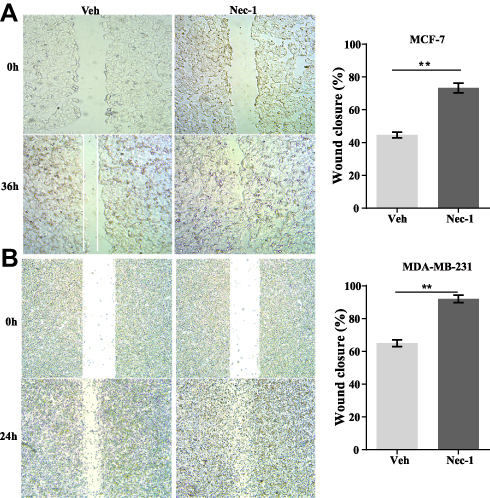 |
Figure 5 Inactivation of necroptosis signaling promotes wound recovery in breast cancer cell lines. (A) MCF-7 cells with or without Nec-1 treatments were subject to wound healing assay. After 36-hour free recovery, the wound was visualized (left panel) and the percentage of wound closure was quantified (right panel). (B) MDA-MB-231 cells with or without Nec-1 treatments were subject to wound healing assay. After 24-hour free recovery, the wound was visualized (left panel) and the percentage of wound closure was quantified (right panel). Note: **p<0.01 as indicated. Abbreviations: Veh, vehicle; NEC-1, necrostatin-1. |
Inhibition of Necroptosis Signaling Enhances Cell Migration Capacities in Breast Cancer Cells
Effects of necroptosis signaling inhibition on cell migration were further assessed. In the transwell migration assay (Figure 6A, left panel), after 12 hours, an average of 222 Nec-1-treated MDA-MB-231 cells migrated to the underneath surface, which contradicted to only 173 control MDA-MB-231 cells (p=0.009, Figure 6A, right panel). An average of 19 Nec-1-treated MCF-7 cells migrated to the underneath surface which was significantly higher than that in the vehicle-treated MCF-7 cells (p=0.004, Figure 6A, right panel). In the transwell invasion assay (Figure 6B, left panel), Nec-1 pretreatments led to an average of 175-invaded MDA-MB-231 cells which was significantly higher than vehicle-treated cells (only 120 cells) (p=0.001, Figure 6B right panel). Nec-1 treatments also increased the invasion capacities of MCF-7 cells by over 90% (p=0.02, Figure 6B, right panel). These findings suggested that inhibition of necroptosis signaling enhanced cell migration capacities in breast cancer cells.
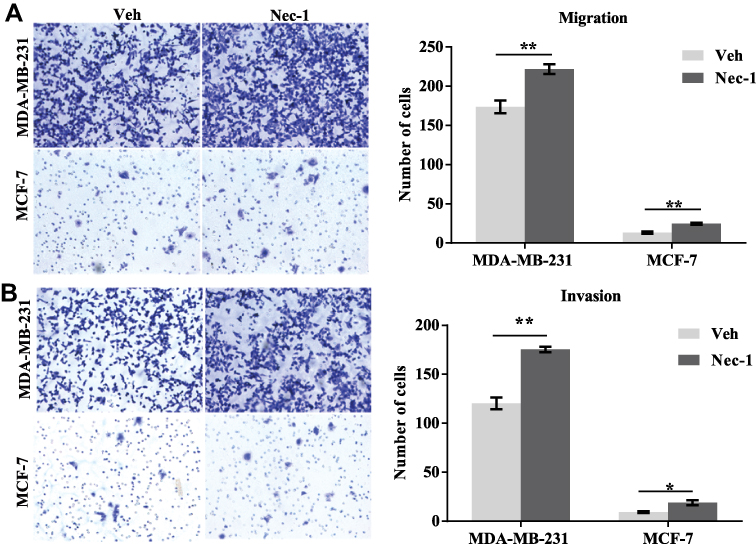 |
Figure 6 Inactivation of necroptosis signaling promotes wound recovery in breast cancer cell lines. (A) MCF-7 and MDA-MB-231 cells with or without Nec-1 treatments were subject to transwell migration assay (left panel). After 12-hour free migration, the numbers of cells on the underneath surface of the membrane were counted (right panel). (B) MCF-7 and MDA-MB-231 cells with or without Nec-1 treatments were subject to transwell invasion assay (left panel). After 12-hour free migration, the numbers of cells on the underneath surface of the membrane were counted (right panel). Notes: *p<0.05; **p<0.01 as indicated. Abbreviations: Veh, vehicle; NEC-1, necrostatin-1. |
Discussion
Breast cancer is one of the most frequently diagnosed cancer types in the world, and breast cancer alone is expected to occupy approximately 30% of all cancers newly diagnosed in women.19 For the past few years, the discovery of cancer immunotherapy has received great attention due to its rationale that innate immune cells help fight against tumor cells.10 Previously immunotherapy was not considered for the treatment of breast cancers due to its so-called property of not being immunogenic.20 Nowadays evidence has indicated that breast cancer patients show an adaptive immune response against their tumors, detectable either in the peripheral blood or in the tumor.20 Hence, breast cancer immunotherapy has been considered to be possible.
As one critical process that releases intracellular components and thereby triggering innate immune responses, necroptosis has been implicated in a variety of inflammatory diseases. The present study investigated the roles of necroptosis signaling in breast cancer cell proliferation and migration. Initially, it was found that key components of necroptosis signaling significantly correlated with breast cancer clinical and pathological parameters. These correlations were independent of neither TNM staging nor pathological subtype, suggesting that necroptosis activation is a widely existed phenomenon along with breast cancer development. Patients at any age, stage or pathological subtype may undergo a comparable extent of necroptosis activation, particularly in the central zone of the neoplasm. In addition, the extent of necroptosis activation correlated well with TNFα, a key pro-inflammatory factor that recruits immune cells to cancer tissues. Our findings suggested the controllability of breast cancer immunotherapy via the target of necroptosis signaling21 and mirrored the simultaneous activation of tumor cell necroptosis with breast cancer malignancy.
To probe the detailed roles of the necroptosis signaling in breast cancer progression, we used a mature pharmacologic inhibitor (Nec-1) of the necroptosis signaling. In the in vitro studies, it was found that pretreatments of breast cancer cells with Nec-1, the specific inhibitor of necroptosis signaling, caused significant increases in cell proliferation, colony formation, wound closure and transmigration capacities. Nec-1 treatments also blunted KDM4A siRNA-conferred protection of breast cancer cell proliferation. Taken together, this evidence was conclusive that necroptosis signaling negatively regulated breast cancer malignant properties. Necroptosis signaling was an endogenous protective signaling for human systems that was potently activated against tumor malignancy and might represent a potent target for therapy of breast cancer.
Necroptosis has been implicated in development, inflammation and disease.22 Previous studies have reported necroptosis as a desperate programmed suicide pathway in breast cancer MDA-MB-468 cells.23 In fact, necroptosis signaling has been reported to serve as a potent target, and extracts from traditional Chinese medicine have been shown to confer therapeutic efficacy against breast cancer by specifically inducing necroptotic cell death.24,25 A Smac mimetic also protected against breast cancer via inducing caspase-independent necroptosis.26 All these pioneer studies were consistent with our present study and altogether suggested necroptosis as a critical programmed suicide pathway that might serve as a potential target for designing therapeutic drugs against breast cancer.
Conclusion
In all, the present study identified necroptosis as a common phenomenon that was simultaneously activated when breast cancer became aggressive. Necroptosis activation correlated well with tumor size and breast cancer malignancy markers. Necroptosis might represent a suicide pathway for breast cancer cells and serve as an endogenous protective signaling for human systems. Approaches enhancing tumor cell necroptosis might be promising therapeutic strategies against breast cancer.
Ethics and Consent Statement
The use of human specimens was approved by the Ethics Review Board of Zhongshan Hospital, Fudan University (No. Y2019-010). All patients showed their full intention to participate in this study and their written informed contents were obtained. This study was performed in accordance with the Declaration of Helsinki.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
This work was financially supported by the Shen Kang Hospital Development Center Foundation (grant No. SHDC12014207).
Disclosure
The authors have declared that no competing interest exists in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020;70(1):7–30. doi:10.3322/caac.21590
2. DeSantis CE, Ma J, Goding SA, Newman LA, Jemal A. Breast cancer statistics, 2017, racial disparity in mortality by state. CA Cancer J Clin. 2017;67(6):439–448. doi:10.3322/caac.21412
3. DeSantis CE, Ma J, Gaudet MM, et al. Breast cancer statistics, 2019. CA Cancer J Clin. 2019;69(6):438–451. doi:10.3322/caac.21583
4. Nguyen T, Hattery E, Khatri VP. Radiofrequency ablation and breast cancer: a review. Gland Surg. 2014;3(2):128–135. doi:10.3978/j.issn.2227-684X.2014.03.05
5. Freedman OC, Fletcher GG, Gandhi S, et al. Adjuvant endocrine therapy for early breast cancer: a systematic review of the evidence for the 2014 cancer care ontario systemic therapy guideline. Curr Oncol. 2015;22:95–113. doi:10.3747/co.22.2326
6. Rosenberg SM, Partridge AH. Management of breast cancer in very young women. Breast. 2015;24(Suppl 2):S154–8. doi:10.1016/j.breast.2015.07.036
7. Han X, Guo X, Zhang W, Cong Q. MicroRNA-937 inhibits the malignant phenotypes of breast cancer by directly targeting and downregulating forkhead box Q1. Onco Targets Ther. 2019;12:4813–4824. doi:10.2147/OTT.S207593
8. Li L, Gao P, Li Y, et al. JMJD2A-dependent silencing of Sp1 in advanced breast cancer promotes metastasis by downregulation of DIRAS3. Breast Cancer Res Treat. 2014;147(3):487–500. doi:10.1007/s10549-014-3083-7
9. Fifield BA, Qemo I, Kirou E, Cardiff RD, Porter LA. The atypical cyclin-like protein Spy1 overrides p53-mediated tumour suppression and promotes susceptibility to breast tumourigenesis. Breast Cancer Res. 2019;21(1):140. doi:10.1186/s13058-019-1211-3
10. Nathan MR, Schmid P. The emerging world of breast cancer immunotherapy. Breast. 2018;37:200–206. doi:10.1016/j.breast.2017.05.013
11. Luen SJ, Salgado R, Fox S, et al. Tumour-infiltrating lymphocytes in advanced HER2-positive breast cancer treated with pertuzumab or placebo in addition to trastuzumab and docetaxel: a retrospective analysis of the CLEOPATRA study. Lancet Oncol. 2017;18(1):52–62. doi:10.1016/S1470-2045(16)30631-3
12. Tung NM, Winer EP. Tumor-infiltrating lymphocytes and response to platinum in triple-negative breast cancer. J Clin Oncol. 2015;33(9):969–971. doi:10.1200/JCO.2014.59.6031
13. Newton K, Manning G. Necroptosis and inflammation. Annu Rev Biochem. 2016;85(1):743–763. doi:10.1146/annurev-biochem-060815-014830
14. Lin J, Kumari S, Kim C, et al. RIPK1 counteracts ZBP1-mediated necroptosis to inhibit inflammation. Nature. 2016;540(7631):124–128. doi:10.1038/nature20558
15. Luo Q, Yang D, Qi Q, et al. Role of the death receptor and endoplasmic reticulum stress signaling pathways in polyphyllin I-regulated apoptosis of human hepatocellular carcinoma HepG2 cells. Biomed Res Int. 2018;2018:5241941. doi:10.1155/2018/5241941
16. Li L, Xue A, Li B, et al. JMJD2A contributes to breast cancer progression through transcriptional repression of the tumor suppressor ARHI. Breast Cancer Res. 2014;16(3):R56. doi:10.1186/bcr3667
17. Metzger E, Stepputtis SS, Strietz J, et al. KDM4 inhibition targets breast cancer stem-like cells. Cancer Res. 2017;77(21):5900–5912. doi:10.1158/0008-5472.CAN-17-1754
18. Berry WL, Shin S, Lightfoot SA, Janknecht R. Oncogenic features of the JMJD2A histone demethylase in breast cancer. Int J Oncol. 2012;41(5):1701–1706. doi:10.3892/ijo.2012.1618
19. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. doi:10.3322/caac.21387
20. Disis ML, Stanton SE. Immunotherapy in breast cancer: an introduction. Breast. 2018;37:196–199. doi:10.1016/j.breast.2017.01.013
21. Garg AD, Agostinis P. Cell death and immunity in cancer: from danger signals to mimicry of pathogen defense responses. Immunol Rev. 2017;280(1):126–148. doi:10.1111/imr.12574
22. Weinlich R, Oberst A, Beere HM, Green DR. Necroptosis in development, inflammation and disease. Nat Rev Mol Cell Biol. 2017;18(2):127–136. doi:10.1038/nrm.2016.149
23. Shahsavari Z, Karami-Tehrani F, Salami S, Ghasemzadeh M. RIP1K and RIP3K provoked by shikonin induce cell cycle arrest in the triple negative breast cancer cell line, MDA-MB-468: necroptosis as a desperate programmed suicide pathway. Tumour Biol. 2016;37(4):4479–4491. doi:10.1007/s13277-015-4258-5
24. Shahsavari Z, Karami-Tehrani F, Salami S. Shikonin induced necroptosis via reactive oxygen species in the T-47D breast cancer cell line. Asian Pac J Cancer Prev. 2015;16(16):7261–7266. doi:10.7314/APJCP.2015.16.16.7261
25. Khorsandi L, Orazizadeh M, Niazvand F, Abbaspour MR, Mansouri E, Khodadadi A. Quercetin induces apoptosis and necroptosis in MCF-7 breast cancer cells. Bratisl Lek Listy. 2017;118(2):123–128. doi:10.4149/BLL_2017_025
26. Jin G, Lan Y, Han F, et al. Smac mimetic-induced caspase-independent necroptosis requires RIP1 in breast cancer. Mol Med Rep. 2016;13(1):359–366. doi:10.3892/mmr.2015.4542
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.