")
Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 15
Pharmacogenetics of Breast Cancer Treatments: A Sub-Saharan Africa Perspective
Authors Nthontho KC , Ndlovu AK , Sharma K , Kasvosve I, Hertz DL , Paganotti GM
Received 24 February 2022
Accepted for publication 31 May 2022
Published 21 June 2022 Volume 2022:15 Pages 613—652
DOI https://doi.org/10.2147/PGPM.S308531
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Keneuoe Cecilia Nthontho,1,2 Andrew Khulekani Ndlovu,1 Kirthana Sharma,3 Ishmael Kasvosve,1 Daniel Louis Hertz,4 Giacomo Maria Paganotti2,5,6
1School of Allied Health Professions, Faculty of Health Sciences, University of Botswana, Gaborone, Botswana; 2Botswana-University of Pennsylvania Partnership, Gaborone, Botswana; 3Rutgers Global Health Institute, New Brunswick, NJ, USA; 4Department of Clinical Pharmacy, University of Michigan College of Pharmacy, Ann Arbor, MI, USA; 5Division of Infectious Diseases, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA; 6Department of Biomedical Sciences, Faculty of Medicine, University of Botswana, Gaborone, Botswana
Correspondence: Giacomo Maria Paganotti, Botswana-University of Pennsylvania Partnership, PO Box 45498, Riverwalk Gaborone, Botswana, Tel +267 3555375, Email [email protected]
Abstract: Breast cancer is the most frequent cause of cancer death in low- and middle-income countries, in particular among sub-Saharan African women, where response to available anticancer treatment therapy is often limited by the recurrent breast tumours and metastasis, ultimately resulting in decreased overall survival rate. This can also be attributed to African genomes that contain more variation than those from other parts of the world. The purpose of this review is to summarize published evidence on pharmacogenetic and pharmacokinetic aspects related to specific available treatments and the known genetic variabilities associated with metabolism and/or transport of breast cancer drugs, and treatment outcomes when possible. The emphasis is on the African genetic variation and focuses on the genes with the highest strength of evidence, with a close look on CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP3A4/5, CYP19A1, UGT1A4, UGT2B7, UGT2B15, SLC22A16, SLC38A7, FcγR, DPYD, ABCB1, and SULT1A1, which are the genes known to play major roles in the metabolism and/or elimination of the respective anti-breast cancer drugs given to the patients. The genetic variability of their metabolism could be associated with different metabolic phenotypes that may cause reduced patients’ adherence because of toxicity or sub-therapeutic doses. Finally, this knowledge enhances possible personalized treatment approaches, with the possibility of improving survival outcomes in patients with breast cancer.
Keywords: breast cancer, genetic variability, inter-ethnic differences, pharmacogenetics, sub-Saharan Africa, toxicity
Introduction
Pharmacogenetics evaluates the effect of inherited genomic variation on patient response (including resistance) to treatment. Genetic variability in germline DNA in the form of chromosomal alterations or DNA sequence variants can affect drug metabolism and/or toxicity, which can have important consequences on a patient’s response to breast cancer treatment.1 Conversely, genomic changes (DNA variants and gene expression profiles) in the breast tumour’s somatic genome may influence rates of apoptosis, cell proliferation, and DNA damage repair, which may also have effects on response to treatment and survival.2 This review will focus solely on human genetic variation in germline DNA underlying breast cancer drug metabolism and possible treatment outcomes, which may impact toxicity, recurrence, and survival. We will address, in particular, the drugs most widely used in Africa and the genetic variants identified in African individuals. Indeed, ethnic differences in anticancer drug disposition are important factors accounting for populations’ variation in treatment response and tolerability.3 It is also important to note that breast cancer incidence as well as cancer presentation in terms of age at onset and stage of cancer among African women tend to differ from that of more developed regions of the world. However, breast cancer types seem to show a similar pattern regardless of age, stage and ethnicities. Having said that, in the era of the development of personalized medicine few major problems arise for African populations: i) the socio-economic context may explain a particular epidemiology in terms of the age of onset and stage of cancer and access to treatment without excluding the genetic aspects predisposing to the disease and determining responses to treatments; ii) the great genetic diversity of African populations implies considering each one separately, taking the precaution of not generalizing data established for one ethnic group to all the others; iii) data on the frequencies of polymorphism in genes of interest in different populations are fragmentary; iv) the number of studies of association between the polymorphisms considered and the responses to breast cancer treatment is very limited.
Breast Cancer in Africa
Breast cancer is ranked as the most common cancer in women worldwide, increasing from 1.7 million incident cases in 2005 to 2.4 million cases in 2015.4 In the more developed regions of the world (Europe/USA/Australia and New Zealand) the age-standardized incidence rates (per 100,000 women per year) ranged between 74.5 and 91.6, whereas recent estimates for sub-Saharan Africa (SSA) were as follows: 30.4 in East Africa; 26.8 in Central Africa; 38.6 in West Africa; and 38.9 in Southern Africa.5 Although the incidence of breast cancer appears to be relatively low in SSA, survival from the disease is markedly poor in the region, with high mortality recorded in many settings.5,6 Sub-Saharan Africa has the highest age-standardized breast cancer mortality rate globally.5 Breast cancer is the most frequent cause of cancer death in less developed regions,7 causing one in five deaths in African women, described as a new “shift” from the previous decade.8 The poor survival of breast cancer patients in SSA has been attributed to several factors, inter alia: late presentation, poor healthcare infrastructure, reduced diagnostic capacity and delays, including substandard pathological data, together with lack of adequate funding, amidst other competing public health challenges.9 In addition, the steadfast adherence to some negative socio-cultural beliefs that delay presentation to health services may contribute to the observed lower survival rates compared to high-income countries. Breast tumours are diagnosed ten or twenty years earlier among Africans compared to developed countries and are at advanced stages at presentation. One possible explanation for this could be that breast cancer among young women in any population tends to be clinically and pathologically aggressive,10 which is also attributed to a consequence of the demographic structure of the population.11 Nevertheless, studies showed a comparable receptor status prevalence in SSA with that of the West.12,13 Further possible explanations for such high rate of aggressive phenotypes could be socio-economic aspects as well as susceptibility factors.
Given that African genomes contain more variation than those from any other continent,14,15 it is therefore of paramount importance to measure the extent of African pharmacogenetics in the context of disease treatment response, including breast cancer. In fact, African genetic variability, which has been well linked to disease resistance and susceptibility, also accounts for the variability in detoxifying pathways that are responsible for eliminating modern drugs.16 During evolution, different populations’ practices exposed individuals to xenobiotics that sometimes-imposed serious health and environmental risks, leading to the selection of specific adaptations linked to an efficient detoxification pathway. For example, significant differences in prevalence of acetylation phenotypes are found between hunter-gatherer and food-producing populations, both in SSA and worldwide, and between agriculturalists and pastoralists in Central Asia.17 This is likely an explanation for such large genetic variation in detoxifying pathways observed among African ethnic groups.
In view of all these factors, it is important to evaluate and discuss the specific pharmacogenetic landscape relevant to breast cancer treatments used in the SSA context, to enhance possible personalized treatment approaches, ultimately improving outcomes in patients with cancer.
Types of Breast Cancer
Breast cancer has a complex aetiology where susceptibility is influenced by both environmental and genetic factors, including increased estrogen exposure throughout a woman’s lifetime, age, family history, as well as modifiable risk factors, such as nutrition, exercise, and alcohol/tobacco use.18,19 Breast cancer may be more appropriately defined as a myriad of diseases characterized by variability in developmental pathways, propensity to metastasize, and response to treatment that can benefit from precision regimens targeted at individual patients.
Breast cancer types are either hormone receptor-positive (HR+) or hormone receptor-negative (HR-), based on tumour cells’ expression of the estrogen receptors (ER) and/or progesterone receptors (PR). ER+ breast cancer types are prognostic for improved survival outcomes and predict responsiveness to endocrine treatment (such as tamoxifen). By binding to ER (either ER-alpha or -beta), estrogen regulates a wide variety of cellular effects and physiological conditions including breast cancer cell proliferation and growth. Nearly two thirds of breast cancers are classified as ER+, and this is broadly true also for Africa, with regional variations.12,13 For example, HR+ breast cancer subtypes have an estimated prevalence of 80% in Nigeria.20 Other studies show a prevalence of ER+ breast cancer between 58% and 64% in South Africa.21 These subtypes have been associated with different prognoses, with patients with luminal A tumours having the best prognosis, and patients with triple negative subtype (ER-/PR-/HER2-) having the worst prognosis22 (Table 1). However, a significant minority of patients relapse despite adjuvant anti-estrogen therapy, with most patients with metastatic disease ultimately developing resistance to anti-estrogen therapies.
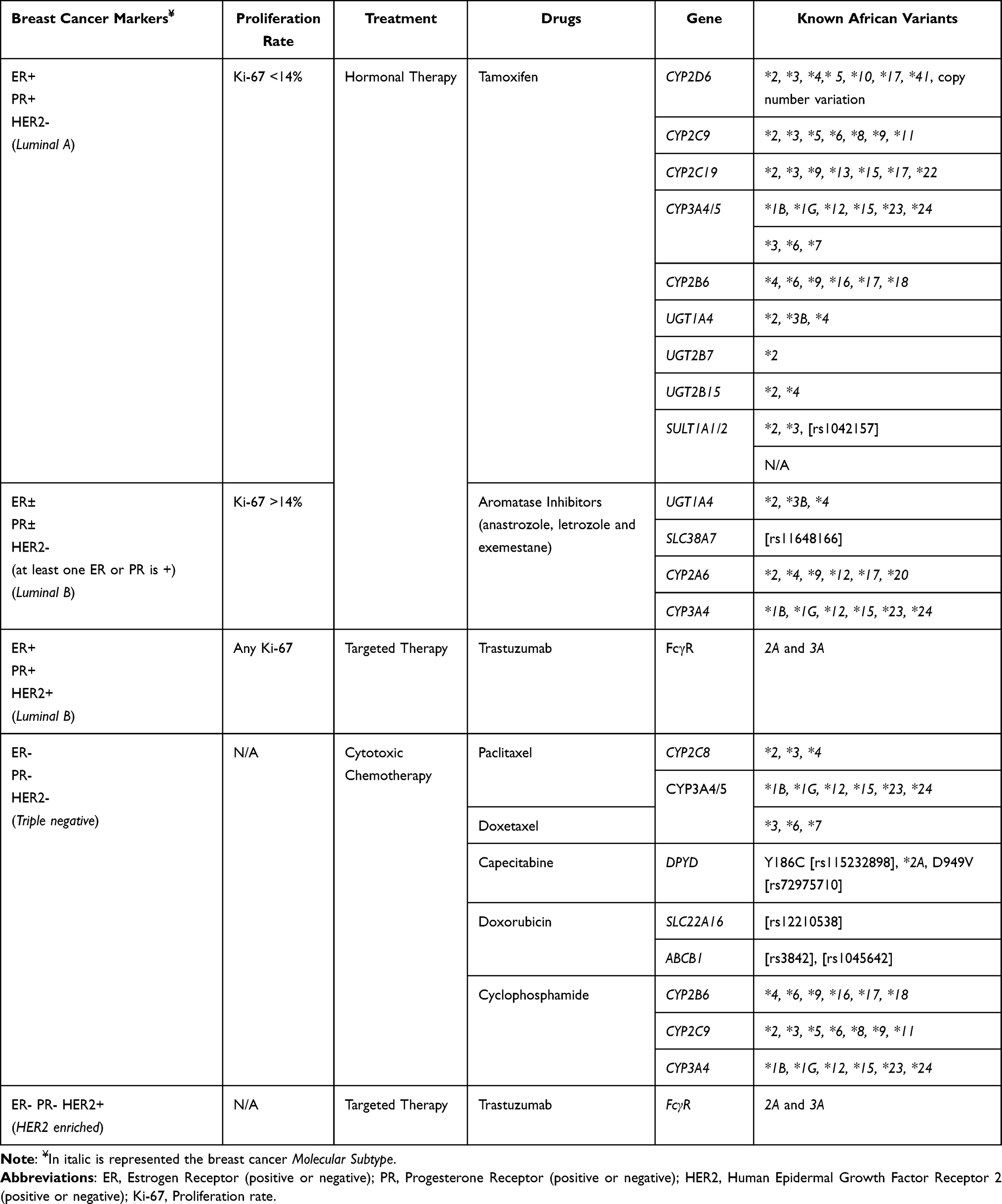 |
Table 1 Classification of Breast Cancer Subtypes Based on the Hormone Receptor (HR) Status, Treatment of Choice, and the Related Metabolizing Genes |
Breast cancer cells with higher than normal levels of the HER2 receptor protein are defined as HER2+. These cancers tend to grow and spread faster than other breast cancers but are much more likely to respond to treatment with HER2 targeting drugs (Table 1), such as monoclonal antibodies.
This narrative review aims to provide an overview of pharmacogenetic variation relevant to the treatment options for women with breast cancer in SSA. The focus will be on pharmacogenetic variants found in populations in SSA, where information is available. We will review the African genetic variation and focus on the genes with the highest strength of evidence. After identifying the most likely relevant pharmacogenes, a comment specifically on the patterns and frequencies of genetic variants in SSA populations will be given. The information will be used to comment on which drugs may be effective in Africans as compared to Caucasians and Asians, where most clinical trials and pharmacogenetic studies are conducted. For allelic frequency look-up and comparison several options, including Ensembl (http://useast.ensembl.org/index.html) and gnomAD (https://gnomad.broadinstitute.org/) are publicly available. Finally, helpful figures of the metabolism and pharmacology for each of the drugs described in the present work can be accessed via PharmGKB (https://www.pharmgkb.org).
Pharmacogenetics of Breast Cancer in Sub-Saharan Africa
Hormonal Therapy
Several hormonal agents have been approved for the prevention or treatment of breast cancer and prevention of recurrence, including the selective estrogen receptor modulator (SERM) tamoxifen, as well as the third-generation aromatase inhibitors (AIs) anastrozole, letrozole and exemestane. Hormonal therapy for breast cancer is one of the most available treatment options even in poorer countries. For example, many companies supply generic tamoxifen at a very low cost making it readily available and, in some countries it is available free of charge.8 Unlike tamoxifen, the access and availability of AIs are restricted in most of Africa.8 For example, recent data from Ghana, Nigeria and Kenya indicate that only 21–29% of eligible patients receive AI treatment.23
Tamoxifen
Tamoxifen is a potent antagonist of the ER with inhibitory effects on tumour growth that has become the gold standard for endocrine treatment of HR+ breast cancer in premenopausal and is also used in postmenopausal women when AIs are intolerable or unavailable.24 Nevertheless, its clinical effectiveness varies among individuals. Tamoxifen is a prodrug that undergoes considerable first-pass oxidative metabolism into more potent active metabolites, such as 4-hydroxytamoxifen (4-OH-tamoxifen) and 4-hydroxy-N-desmethyl-tamoxifen (endoxifen). These metabolites have a 30- to 100-fold higher affinity to ERs compared to tamoxifen. Because endoxifen reaches a steady-state plasma concentration 6- to 10-fold higher than 4-OH-tamoxifen, it is considered the most relevant metabolite in determining the parent drug’s clinical efficacy.25 CYP2D6 is the primary enzyme responsible for the activation of tamoxifen. However, other metabolic enzymes and transporters26 have been identified as possible contributors to tamoxifen plasma concentration variations.27
CYP2D6
Over the last few decades, a huge body of research first discovered and then elaborated on tamoxifen metabolism, identifying CYP2D6 as the main enzyme responsible for tamoxifen activation to endoxifen, the most potent antiestrogenic metabolite.28–30 CYP2D6 interindividual gene variation is the predominant predictor of plasma endoxifen level in a gene-dose dependent manner. In fact, CYP2D6 explains ~40–50% of endoxifen plasma concentration, while all the other known genes and clinical variables combined explain <10%.27 Globally, it has been challenging to demonstrate that endoxifen plasma concentration determines treatment efficacy, and a direct genotype–phenotype association has not yet been confirmed. Some studies have found that patients who carry reduced-function or non-functional CYP2D6 alleles derive inferior therapeutic benefits from tamoxifen,25 or have significantly shorter disease-free survival than non-carriers,31 while other studies did not find any association, or an inverse association.32–34 The clinical validity of this association has not been demonstrated, hence providing insufficient data for the clinical utility of CYP2D6 genotyping to guide tamoxifen treatment.34 It should be stressed that most of the studies to date have been conducted among Caucasians and/or Asians, and limited research has been conducted among Africans. From the few papers published on SSA breast cancer pharmacogenetics, a study conducted in Ethiopia26 found that an increase in CYP2D6 activity was associated with increased endoxifen concentration, confirming the notion of a linear relation between CYP2D6 and endoxifen plasma concentration. The authors also showed that all null or low activity genotypes (poor and intermediate metabolizers) in this Ethiopian cohort were associated with low endoxifen levels.26 In addition, other factors, such as environmental/dietary and regulatory mechanisms including other genetic polymorphisms, have been suggested as contributors to the generally low endoxifen concentration described in the study.26,35
Given the evolving data regarding the role of CYP2D6 for tamoxifen bioactivation and efficacy, it is critical to consider which CYP2D6 functional alleles may be found in African individuals. In general, more than 70 alleles of CYP2D6 have been identified with large interindividual and interethnic differences, and this is also true within SSA. For example, Africans have the second highest observed frequency of CYP2D6 poor metabolizer phenotypes globally (about 3%) after Caucasians (about 5%),36,37 and this reflects specific ethnic characteristics among different African populations. Importantly, the rate of CYP2D6 ultra-rapid metaboliser phenotypes among Africans is believed to be the highest worldwide.37 It should be noted that a huge range of variations could also be found among African ethnic groups concerning Single Nucleotide Polymorphisms (SNPs).
Alleles known to be unique to SSA populations include the reduced function CYP2D6*17 and CYP2D6*29 alleles38 (Table 2). However, other CYP2D6 low activity variants show a marked difference in distribution among African populations compared with other global populations: CYP2D6*3, CYP2D6*4, CYP2D6*9, CYP2D6*10 and CYP2D6*41, having higher (*10) or lower (*3, *4, *9 and *41) frequencies.39 Interestingly, the South African Cape Coloured population, which is a unique and genetically complex admixed group, shows distinctive allele frequencies for most of the genes analysed. For example, CYP2D6*5 (CYP2D6 gene deletion, that is a no activity variation) occurs more frequently in the South African Cape Coloured population40,41 than among other sympatric South African ethnic groups (Table 2).
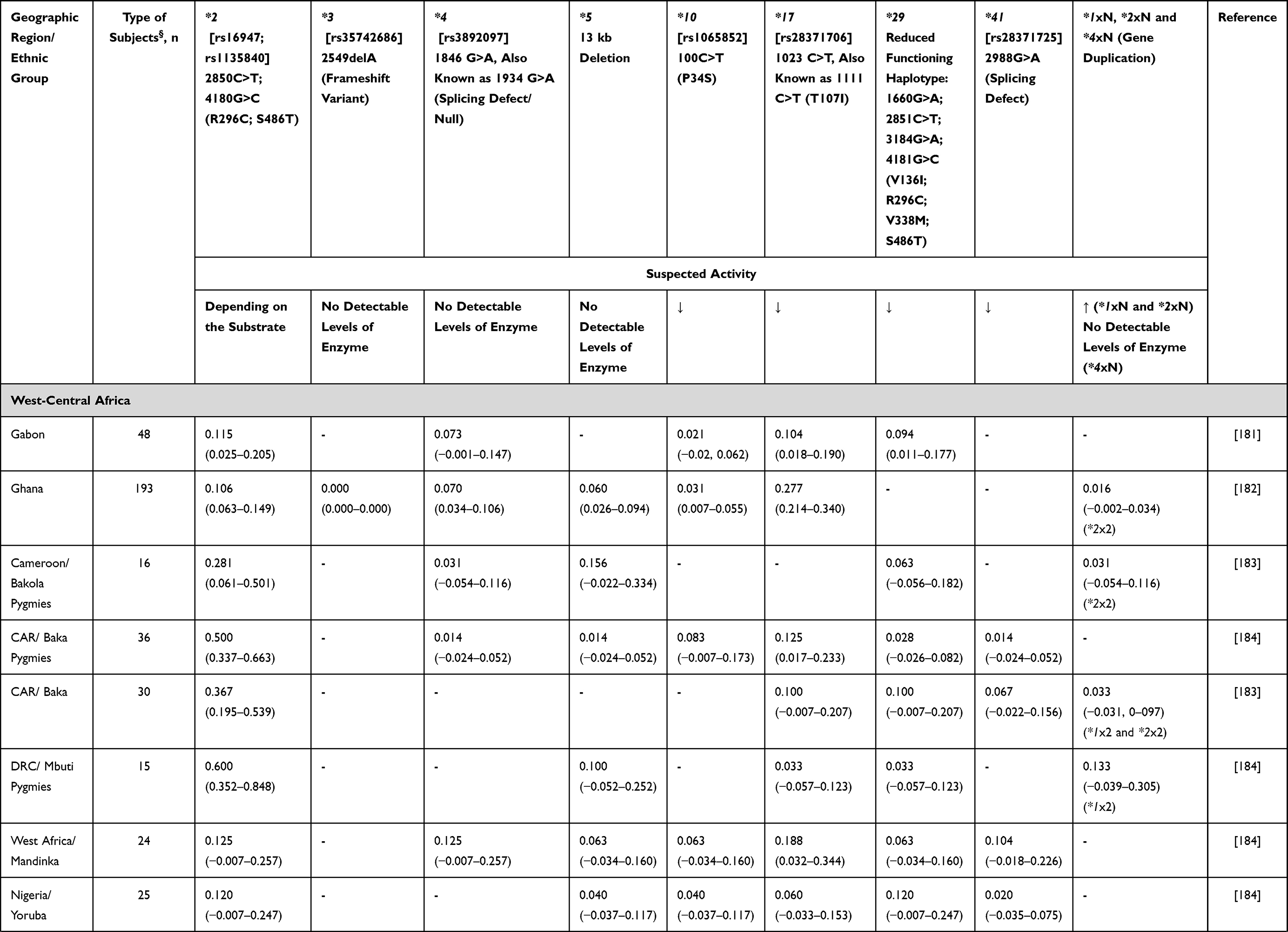 | 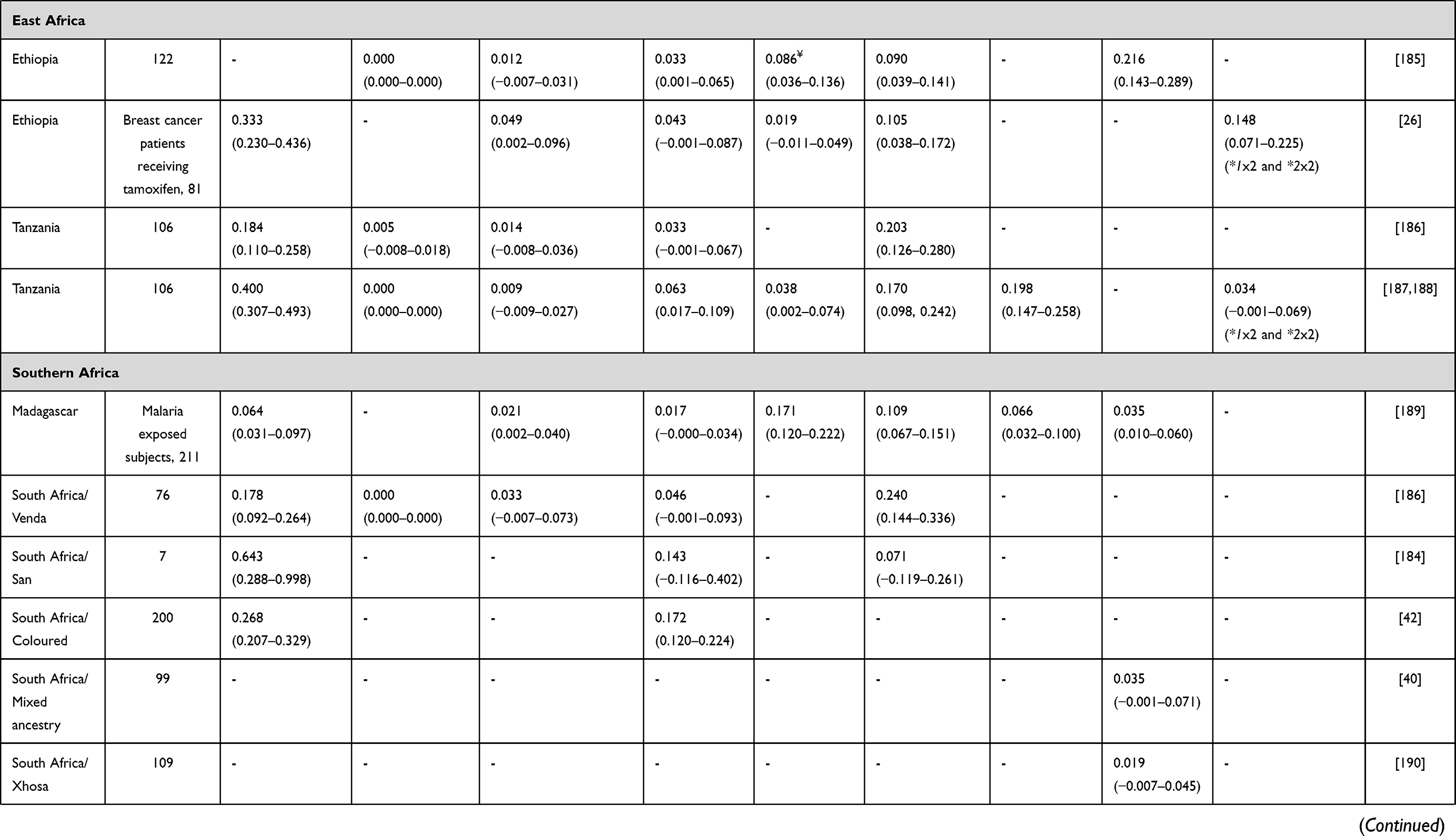 | 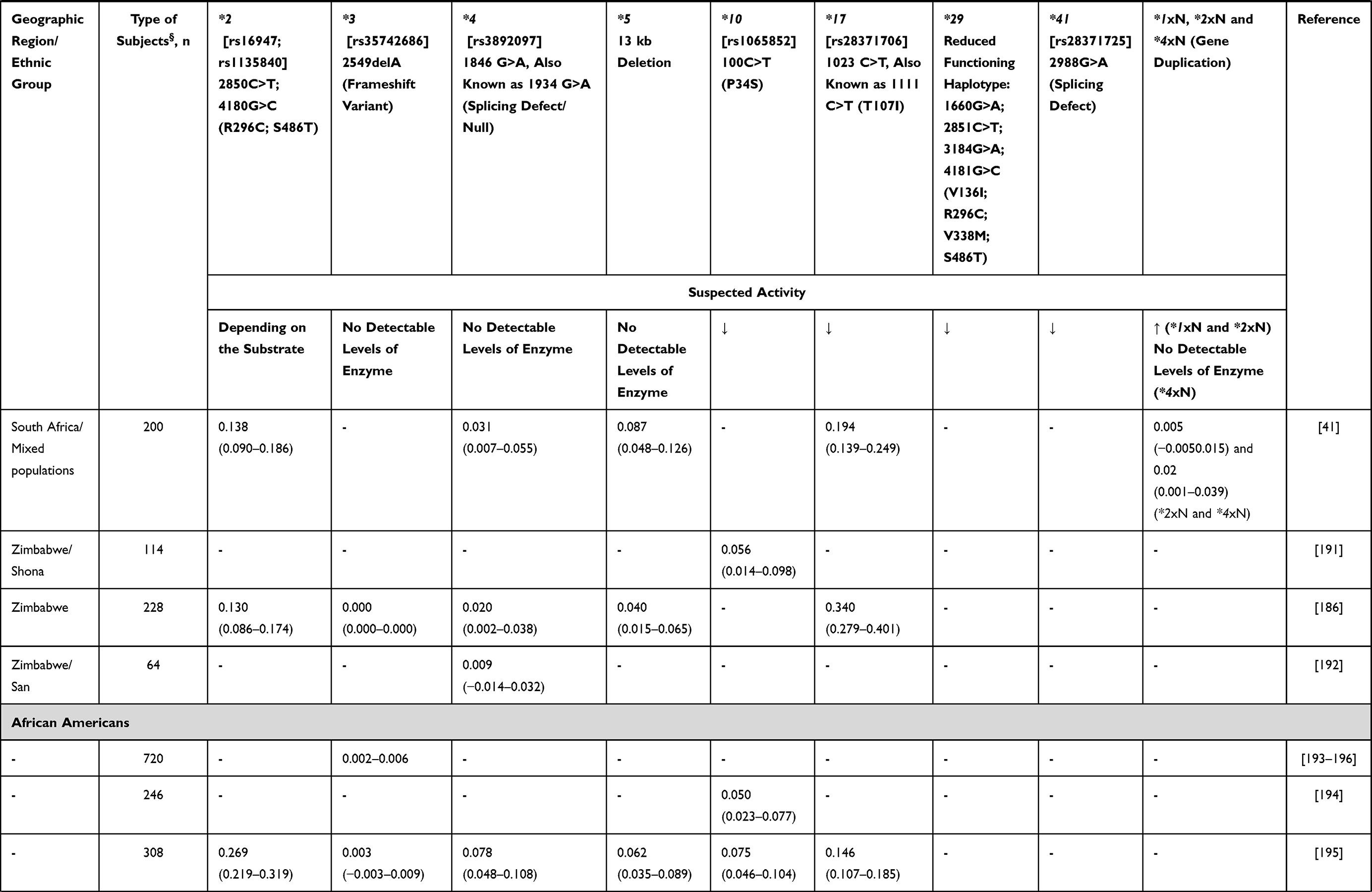 | 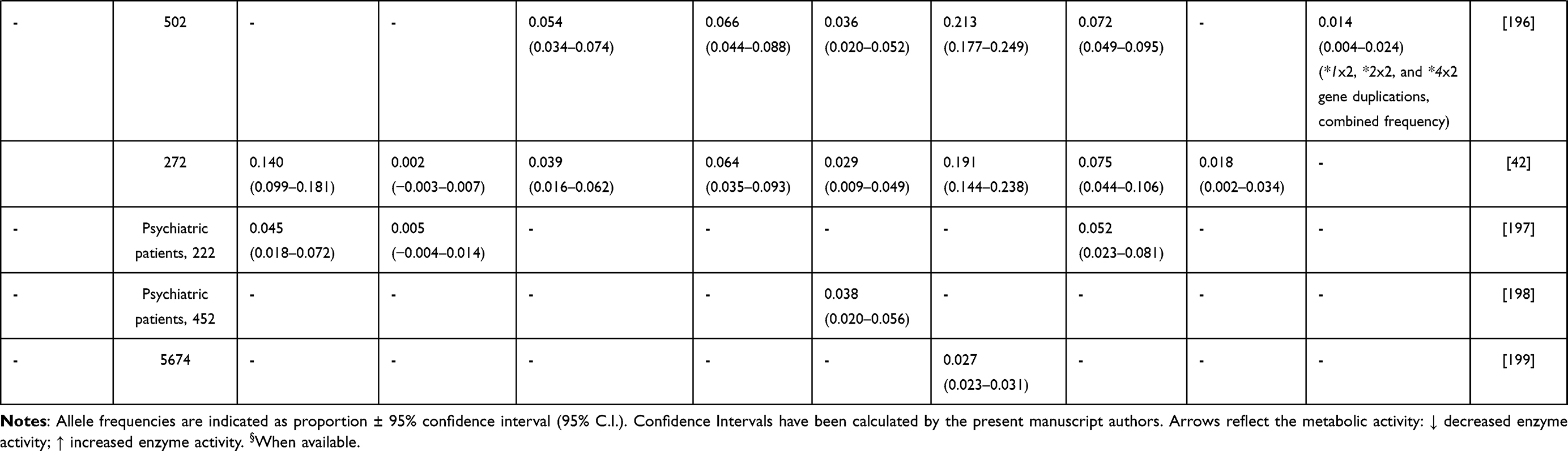 |
Table 2 CYP2D6 Allele Frequency in Sub-Saharan Africa |
In the last few years to simplify genotype interpretation and improve phenotype prediction, the utility of an “activity score” (AS) system was evaluated42 and subsequently improved.43 It spans from values 0 to 2, and >2, where AS is calculated based on an additive model counting individual alleles and haplotypes, similarly developed also for CYP2A6,44 CYP2B645,46 and for CYP2C19.47 The CYP2D6 AS distribution in world populations shows that Africans (in general) do not express particularly outlying metabolic phenotypes.48
Other Tamoxifen-Related Pharmacogenes
Tamoxifen is N-dealkylated and 4-hydroxylated to endoxifen by CYP2D6 but also by several other CYP enzymes that combined together may explain only a fraction (<10%) of tamoxifen metabolism.27 Among them, CYP2C9 and CYP2C19 play a major role, and CYP3A4, CYP3A5, and CYP2B6 a minor role.49 That could be due to the observation that CYP2C9/19 genes are more polymorphic than CYP3A4/5 genes. Furthermore, tamoxifen metabolites are inactivated prior to elimination by other non-CYP450 enzymes, through conjugation with a glucuronide or sulphate group (UGTs and SULT enzymes, respectively), specifically UGT1A4, UGT2B7, UGT2B15, and SULT1A1, SULT1A2 and UGT1A4.27
There is evidence that variation in CYP2C9 gene (together with CYP2D6) may also predict active metabolite concentrations and therefore may be useful to guide tamoxifen dosing.50–52 Lower concentrations of endoxifen and endoxifen/4-hydroxytamoxifen ratios were seen with impaired CYP2C9 activity if patients had the same CYP2D6 phenotype and were not taking CYP2D6 inhibitors.50 However, the only report from Africa (Ethiopia) concerning the possible effect of CYP2C9 polymorphisms (namely *2 and *3 alleles) did not show any association with tamoxifen and metabolite concentrations.26 Known CYP2C9 variants among Africans are as follows: *2, *3, *5, *6, *8, *9, *11, *31 and *32 (Table 3). There is no substantial difference in CYP2C9 frequencies between Africans and other world populations. In line with these observations, it is important to highlight that the influence of CYP2C9 genetic variants on the metabolism of the CYP2C9 probe drug warfarin has been shown to differ by “race”.53
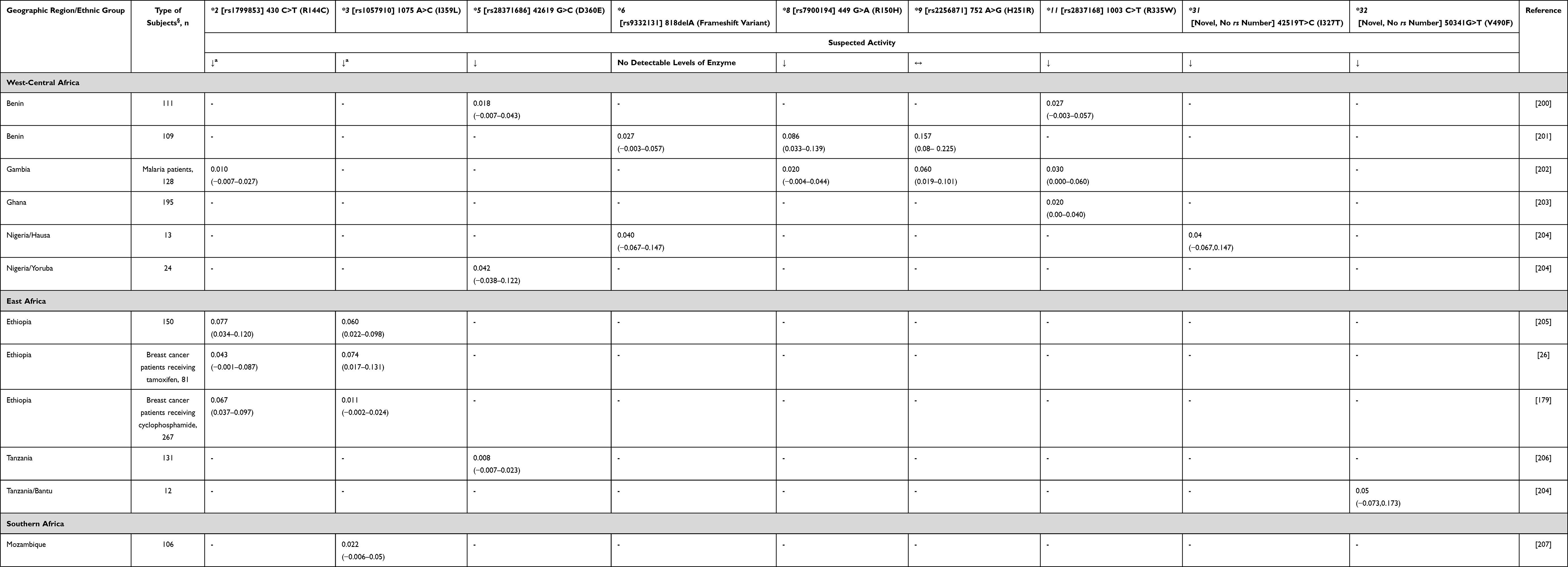 |
Table 3 CYP2C9 Allele Frequency in Sub-Saharan Africa |
CYP2C19 catalyzes the formation of several tamoxifen metabolites, including the conversion of 4-OH-tamoxifen to endoxifen. Some studies have detected an association between CYP2C19 status and clinical outcomes, while others have not. For example, Damkier et al49 and Sanchez-Spitman et al54 found no evidence to support the clinical role of CYP2C19 polymorphisms and response to tamoxifen in breast cancer patients. Conversely, one study found that CYP2C19*17 allele is linked to improved benefits with respect to lower breast cancer recurrence and relapse-free survival.55 This is also supported by another report which found that low activation of tamoxifen in patients with poor CYP2D6 activity and fast CYP2C19 (*17 allele) metabolism represents the fraction of patients that have the worst clinical outcome.56 The only report from Africa (Ethiopia) concerning the possible effect of CYP2C19 polymorphisms (namely *2 and *3 alleles) did not show any association with tamoxifen and metabolite concentrations,26 supporting the non-association relationship. Known CYP2C19 variants among Africans are as follows: *2, *3, *9, *13, *15, *17, *22 (Table 4). Global data comparison suggests that Africans have a high rate of fast metabolizers predicted phenotypes (around 20%, similar to Caucasians) and a relatively lower frequency of poor metabolizer phenotypes.37
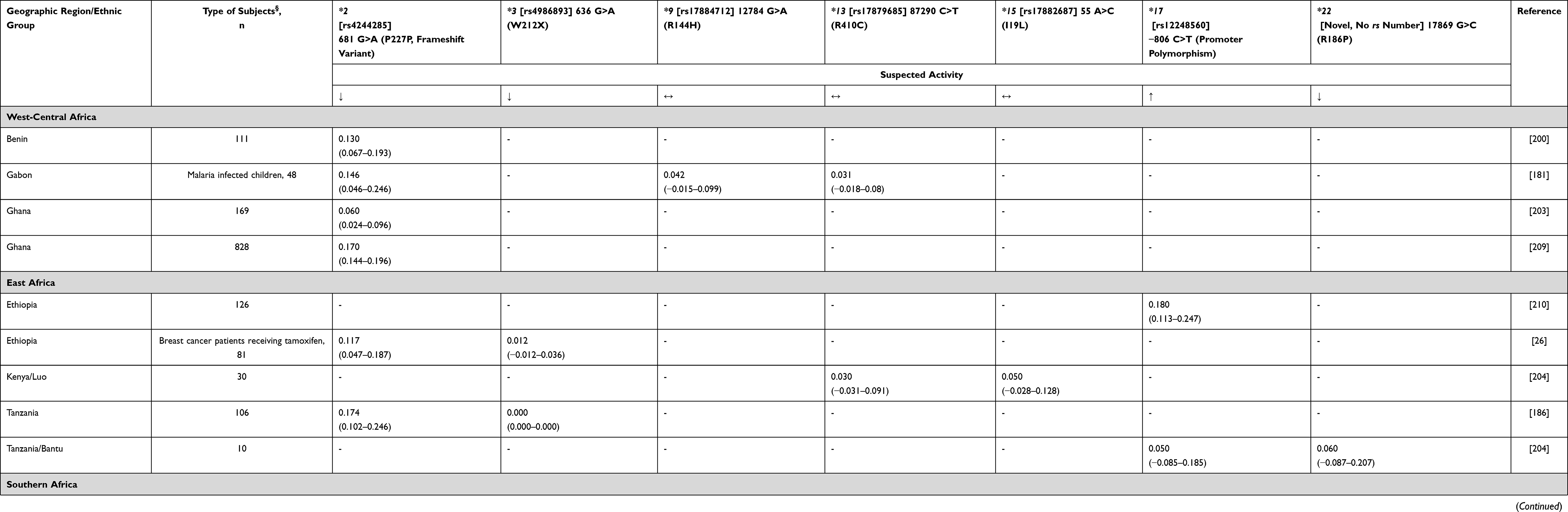 |
Table 4 CYP2C19 Allele Frequency in Sub-Saharan Africa |
As for the other contributors to tamoxifen metabolism, available data indicate the existence of a CYP3A4 main “African” polymorphism, CYP3A4*1B, with functional importance as to enzyme expression.16,57 The CYP3A4*1B slow metaboliser allele (rs2740574) is associated with a significantly reduced oxidation capacity compared to the CYP3A4*1 wild-type allele for several drugs.16,58 Importantly, there is a strong linkage disequilibrium (LD) between CYP3A4*1B and CYP3A5*3, which may also be responsible for the reported findings.59 In fact, high LD between haplotypes spanning CYP3A4 and CYP3A5 confounds the interpretation of the effects of polymorphisms in either gene on drug metabolism. However, despite CYP3A4*1B having been associated with a poor metabolizer phenotype and showing high to very high frequencies in African populations (Table 5), no data on CYP3A4*1B and tamoxifen metabolism has been reported to date. In addition, there is evidence that CYP3A4 is a very conserved gene with <10% of alleles harbouring identified variant haplotypes, and there is no substantial phenotype difference among global populations based on available studies.60
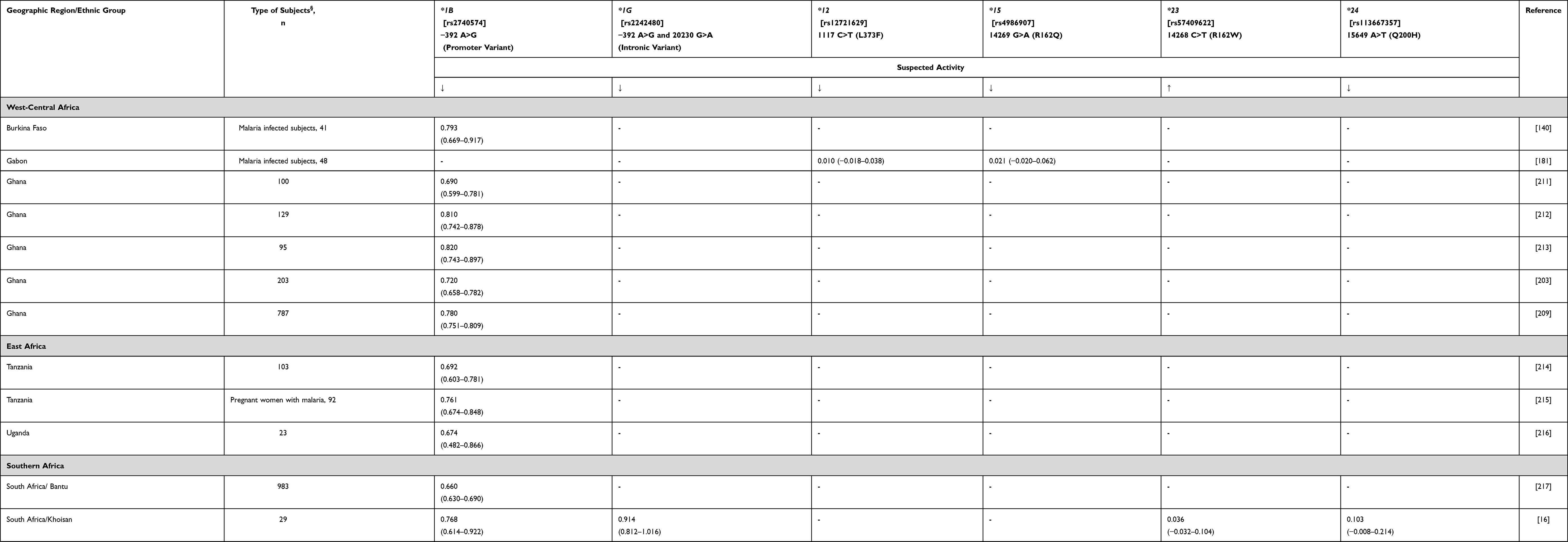 |
Table 5 CYP3A4 Allele Frequency in Sub-Saharan Africa |
Another important pharmacogene is CYP3A5. To our knowledge, the only report from Africa (Ethiopia) concerning the possible effect of CYP3A5 (but also CYP2C9 and CYP2C19) on altered tamoxifen metabolism and altered exposure to its metabolites, is that of Ahmed et al,26 where no evidence of association was observed. This confirms previous findings where CYP3A5*3 allele did not significantly contribute to explaining the inter-variability among patients treated with tamoxifen.61 CYP3A5*3 hampers enzyme expression and is dominant in Asian/Caucasian populations, whereas most Africans are CYP3A5*1 (so-called wild-type allele, associated with full enzyme expression). Conversely, a study from 2007 showed that the CYP3A5*3 allele was associated to higher 5-year recurrence-free survival (RFS) rate (but not 2-year RFS) in breast cancer patients from Sweden.62 Nevertheless, the association reported has not been validated, and it should be stressed that CYP3A5, which explains <2% of tamoxifen metabolism, would be extremely unlikely to be responsible for meaningful associations with tamoxifen efficacy and/or survival. Furthermore, although CYP3A5*3 shows its highest frequencies among Asians and Europeans (69–74% and 93–96%, respectively),63 the variant spectrum of CYP3A5 in Africans is distinctly different, with higher frequencies in CYP3A5*6 and *7 alleles, conferring a substantial homogeneity among world populations.64 The known CYP3A5 variants among Africans are *3, *6, and *7 (Table 6).
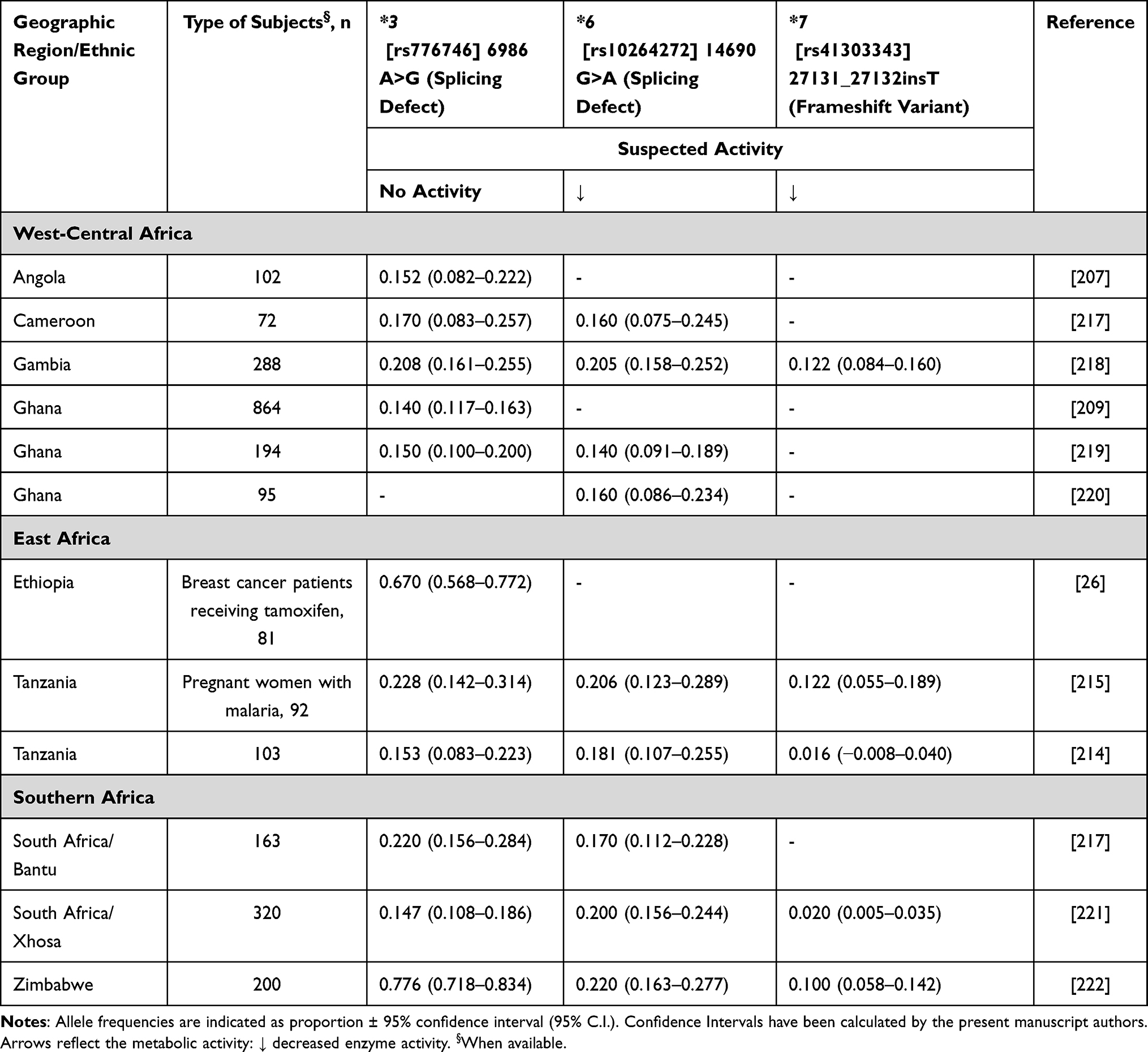 |
Table 6 CYP3A5 Allele Frequency in Sub-Saharan Africa |
The SULT1A1 and SULT1A2 are polymorphic genes that may affect endoxifen level. SULT1A1/2 catalyse the sulfation of endogenous and exogenous molecules, including endoxifen, thus contributing to their excretion.27,65 Two SNPs in the SULT1A1 gene, namely rs6839 (902 A>G) and rs1042157 (973 C>T), have been associated with decreased enzyme activity, and when carried together, showed higher levels of endoxifen plasma concentration.27 Conversely, a study made in Spain assessed that subjects who are wild-type for CYP2D6 and carry SULT1A2*2 or SULT1A2*3 showed significantly higher plasma levels of 4-OH-tamoxifen and endoxifen, than wild type.66 Similar to the inconsistent effects on endoxifen exposure, studies in tamoxifen treated women have reported that patients who carry SULT1A1*2 had better67 or poorer65,68 survival than non-carriers. These associations have not been validated, and no data are currently available from Africa in the context of breast cancer treatment. Known SULT1A1 variants assuming relevant frequencies in Africans are as follows: SULT1A1*2, SULT1A1*3 and rs1042157 (Table 7).
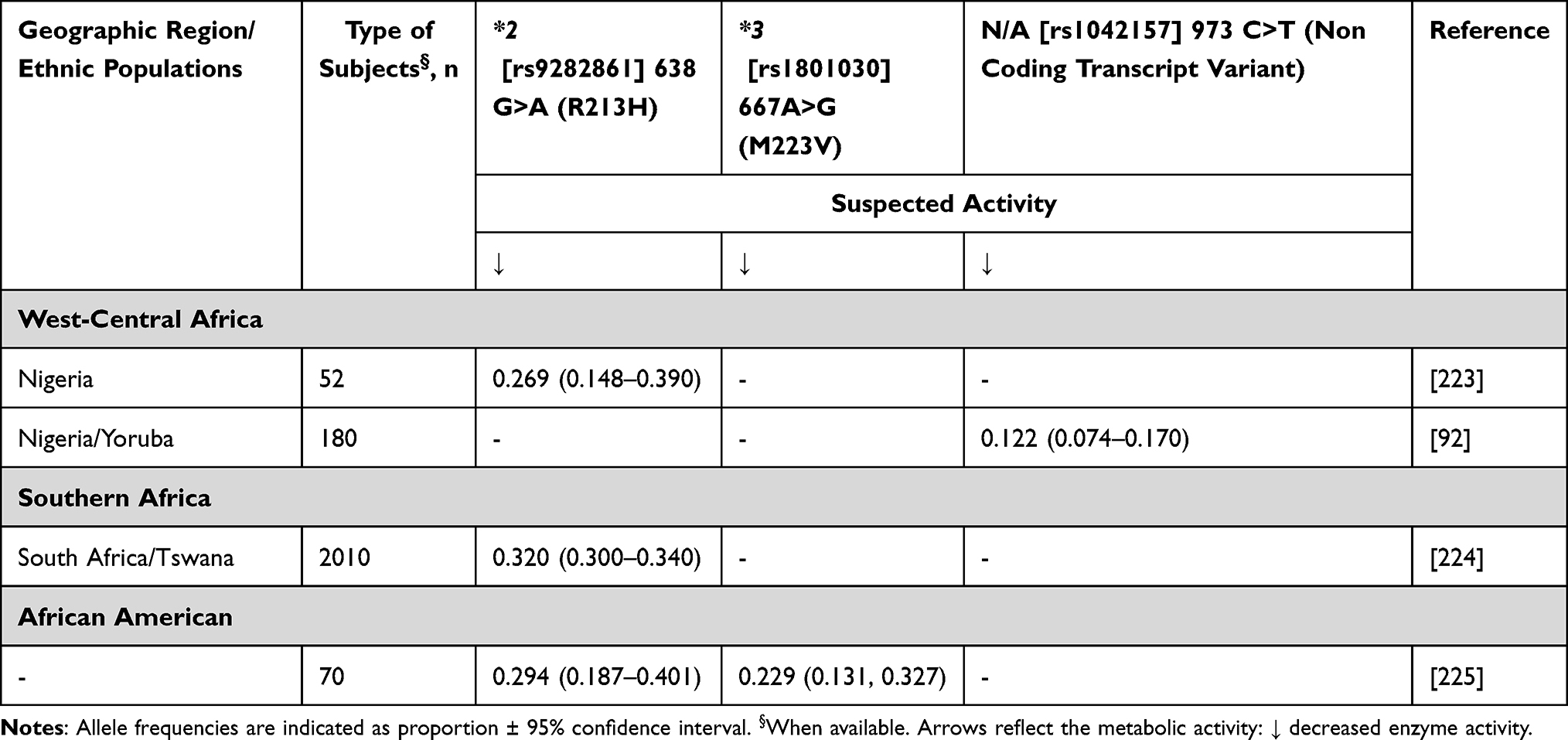 |
Table 7 SULT1A1 Allele Frequency in Sub-Saharan Africa |
Another route to tamoxifen metabolite elimination is via glucuronidation by UGT enzymes. These UGTs (1A4, 2B7, 2B15) have an almost negligible effect on inactive secondary metabolites. However, in a study conducted in Ethiopia, breast cancer patients carrying the UGT2B15*4 allele showed a lower plasma concentration of tamoxifen compared to those with wild-type genotype.26 Another set of data also suggests that in Caucasians, the co-presence of UGT1A4*2 and UGT1A4*3B may be associated with reduced concentrations of glucuronidated metabolites.69 Concerning UGT2B7, the UGT2B7*2 allele has been associated with decreasing activity against tamoxifen metabolites in vitro.70 No data are currently available from Africa in the context of breast cancer treatment. The African UGT frequencies are shown in Table 8.
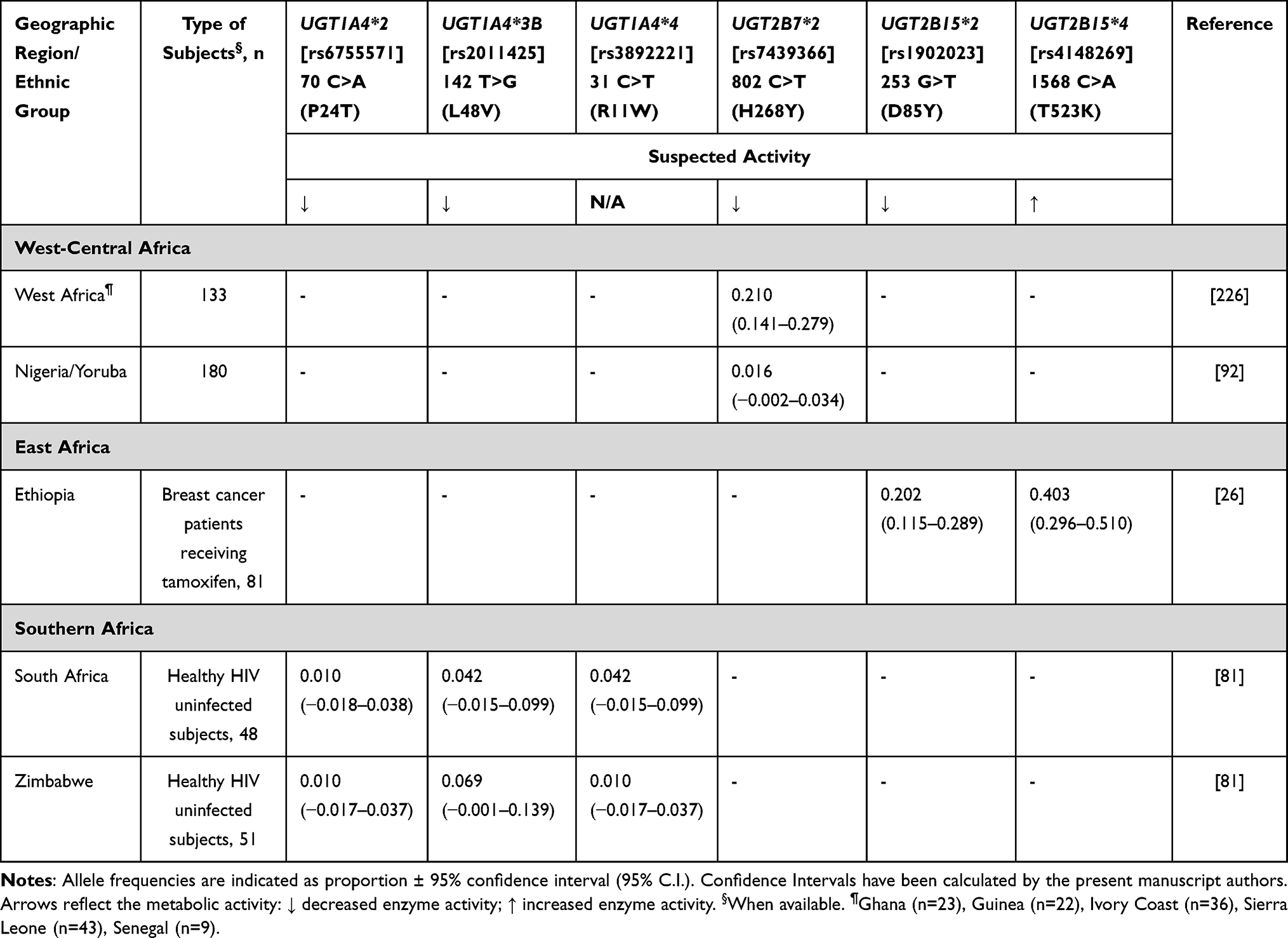 |
Table 8 UGTs Allele Frequency in Sub-Saharan Africa |
Aromatase Inhibitors
In contrast to tamoxifen, the third-generation aromatase inhibitors (AIs), anastrozole, letrozole and exemestane, are considered active in the parent form, and metabolism serves as a means of inactivation. Several large, randomized trials comparing AIs with tamoxifen as adjuvant hormonal therapy have demonstrated significant improvement in disease-free survival and reduction in breast cancer events.71 In general, AIs have demonstrated better efficacy than tamoxifen, but they are not easily available and are often too expensive for patients in developing countries.72 However, some studies have been conducted in South Africa where AIs are used as an alternative to tamoxifen.73,74
These drugs function by inhibiting the enzyme aromatase, which is encoded by the CYP19A1 gene. The aromatase enzyme is responsible for the conversion of androgens to estrogens that ultimately fuel ER+ breast cancer cells. Therefore, AI pharmacogenetics involves several genes encoding enzymes and transporters, while genetic variability of the CYP19A1 target enzyme may affect cancer susceptibility and/or AI treatment efficacy.75
Pharmacogenetics of AI Systemic Concentrations
Current knowledge of AI pharmacogenetics suggests that variability in anastrozole, letrozole and exemestane-metabolizing genes contributes to drug plasma concentrations, but there is much weaker evidence that drug concentrations have any meaningful effect on treatment toxicity76 or efficacy, including systemic estrogenic response.77 Anastrozole metabolism in vitro (using human liver microsomes and Baculovirus-insect cells expressing human P450s) is mediated by CYP3A4 and CYP3A5, with minor contribution by CYP2C8, CYP2D6 and CYP2B6.78 However, a recent genome-wide association study (GWAS) identified a polymorphism in the SLC38A7 gene (rs11648166) that was associated with higher systemic anastrozole concentration.79 This SNP may affect the expression of a glutamine plasma membrane influx transporter not previously known to transport drugs (including anastrozole). In vitro studies also implicate UGT1A4 in anastrozole metabolism and suggest that three promoter SNPs, rs3732219 (−219C>T), *1G allele (no rs number assigned, −217T>G) and rs3732218 (−163G>A), increase anastrozole glucuronidation.80 There are no known reports on the frequencies of these SNPs among Africans; however, three other low-activity UGT1A4 variants have been reported in a non-cancer context81 (Table 8).
Letrozole is pharmacologically similar to anastrozole, however its metabolism is dependent on CYP2A6, with a minor contribution by CYP3A4 and CYP3A5.82,83 A study by Borrie et al84 showed that the CYP2A6 reduced-function genotypes were significantly associated with increased plasma letrozole levels in Canadian patients. The relevance of CYP2A6 pharmacogenetics to letrozole pharmacokinetics was recently confirmed by GWAS.85 African frequencies for CYP2A6 alleles are shown in Table 9.
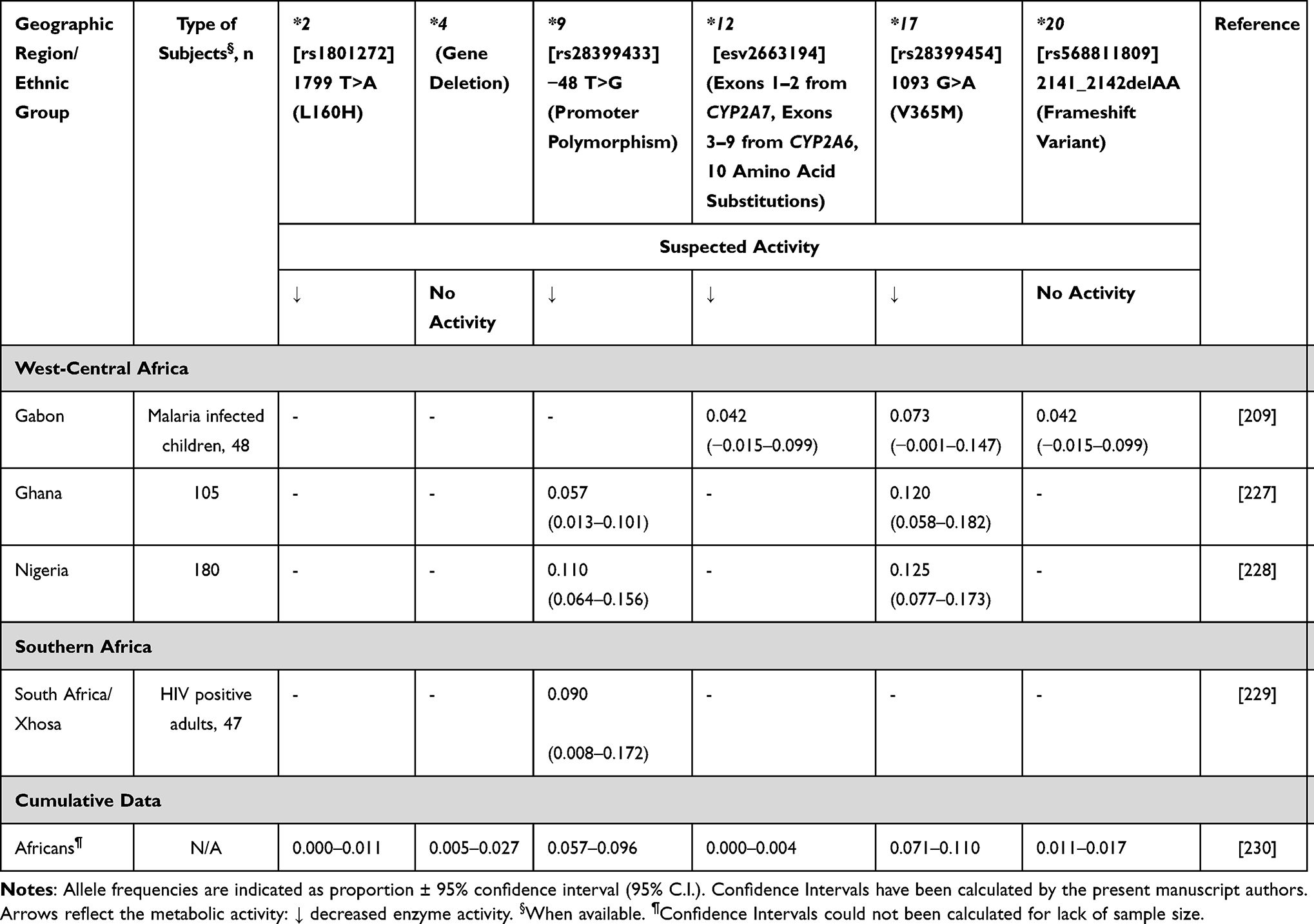 |
Table 9 CYP2A6 Allele Frequency in Sub-Saharan Africa |
Exemestane undergoes metabolism primarily through CYP3A4 with minor contribution from CYP1A1 and CYP4A11. The loss of function CYP3A4*22 allele is associated with a higher exemestane plasma concentration.86 CYP3A4*22 has an allelic frequency of 6–8% in Caucasians, whereas it has not been reported in African subjects.87 Nevertheless, it is well known that among Africans the CYP3A4*1B allele has a very high frequency (Table 5), but its possible contribution to exemestane metabolism has not yet been established.
Pharmacogenetics of AI Toxicity
Up to one quarter of patients discontinue AI therapy due to intolerable toxicities.88 The most common signs of toxicity are musculoskeletal, such as arthralgias, myalgias and tendinopathies, which can also be severe.76 One initial study assessed that a variant of estrogen receptor 1 signaling gene (ESR1), rs9322336 (151879295T>C), was associated with an increased risk of musculoskeletal toxicity during exemestane administration.88 A later GWAS implicated genetic variability in T-cell leukaemia protein 1 gene (TCL1A) that may increase AI-induced musculoskeletal toxicity, though other studies have been unable to replicate this finding.88–90 Recent work by Hertz et al91 suggests that carriers of osteoprotegerin gene (OPG) rs2073618 may be at increased risk of musculoskeletal adverse events. However, further validation of the role of ESR1, TCL1A and OPG genes awaits further research. ESR1 rs9322336 frequency in African Americans is 4.33%.92
Pharmacogenetics of Aromatase (CYP19A1)
CYP19A1 encodes for the aromatase enzyme that catalyses the conversion of androgens to estrogens. AIs target aromatase to reduce estrogen production, ultimately blocking replication of ER+ breast cancer cells. CYP19A1 polymorphisms therefore may impact estrogen production (that is a susceptibility factor for breast cancer) and also respond to AI treatment.
Sequencing of CYP19A1 reveals many variants, some of which have been shown to decrease aromatase activity in vitro.93 Alterations in aromatase expression have been implicated in the pathogenesis of estrogen-dependent diseases including breast cancer. For example, the data on CYP19A1 support an association between the number of (TTTA)n repeats in intron 4 and breast cancer risk, but the biological mechanism for this relationship is unknown.94 The same polymorphism has been associated with bone homeostasis in response to hormone replacement therapy.73 Studies of the putative association of CYP19A1 genetics and AI treatment outcomes have reported inconsistent results. For example, the SNP rs4646, in the context of letrozole treatment efficacy, was found to be associated with longer disease-free survival in metastatic breast cancer patients from Spain.95 Similarly, a study from China detected superior tumour response, clinical benefit, time to progression and overall survival in carriers of rs4646 receiving anastrozole treatment.96 Conversely, another study reported that patients rs4646 carriers were more likely to be non-responders to letrozole.97 Conflicting results for these and other CYP19A1 polymorphisms suggest null or negligible effects on AIs treatment outcomes.76 A recent report on CYP19A1 and bone loss with anastrozole in South Africa74 showed that genotyping for rs10046 (1531 G>A) is an additional tool for risk stratification in AI-related bone outcomes. Women with AA genotype were found to be about 10 times more likely to have an increased percentage of bone loss.74 Table 10 reports allele frequencies for relevant polymorphisms in African populations.
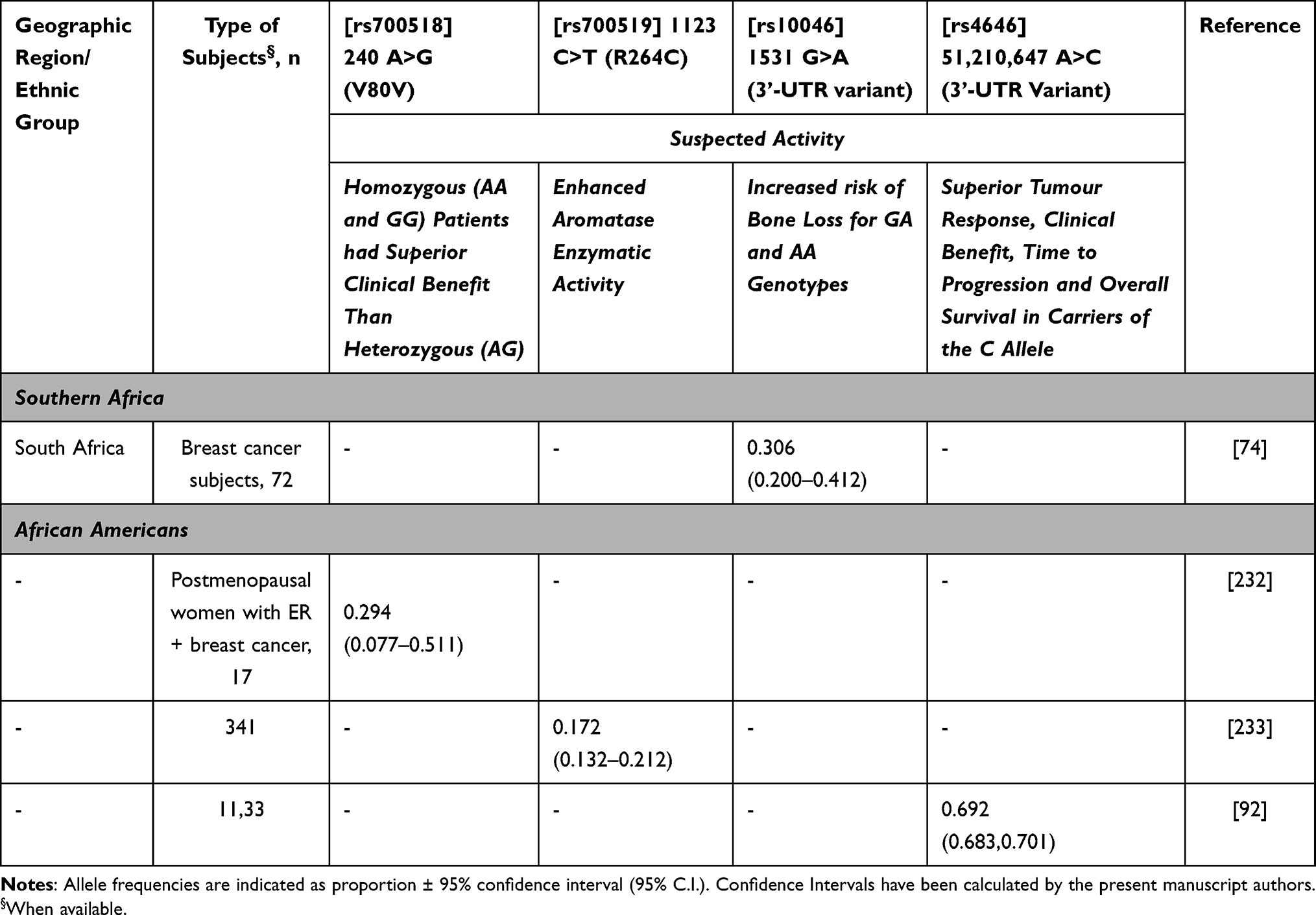 |
Table 10 CYP19A1 Allele Frequency in Sub–Saharan Africa |
Targeted Biological Therapy
Trastuzumab is a humanized monoclonal antibody that binds specifically to the HER2 receptor and suppresses cell proliferation that is driven by over-expression of the HER2 protein. The monoclonal antibody also binds Fc gamma (Fcγ) receptor on an effector cell, such as a natural killer cell, monocyte, or macrophage. The combination of trastuzumab with chemotherapy has led to a significant reduction in breast cancer recurrence and mortality in HER2 overexpressing or amplified tumours (HER2+) when used in the adjuvant setting.98–100 Despite substantial improvements in outcomes with the use of trastuzumab, with disease-free survival of more than 10 years in high-income countries, there are variations in response to adjuvant trastuzumab. Furthermore, in Africa, trastuzumab is increasingly used, but it is not widely available due to high drug pricing in many countries.101
Studies on the pharmacogenomics of trastuzumab have been mostly focused on polymorphisms of the Fcγ receptors 2a and 3a genes (FCGR2A and FCGR3A). There are two main candidate SNPs in the two genes thought to possibly influence trastuzumab treatment outcomes, namely FCGR2A H131R (rs1801274) and FCGR3A F158V (rs396991) (Table 11). These variations are also associated with auto-immune, auto-inflammatory and infectious diseases’ susceptibility,102 and with the efficacy of immunotherapy in cancer patients. However, genetic analysis of the variants at the locus is hampered by the genetic complexity deriving from a segmental duplication, inconsistent nomenclature, and a high degree of linkage disequilibrium.103 Nevertheless, the combination of the two wild-type genotypes (H/H and/or F/F, which are in linkage disequilibrium) has been associated with better response rate and progression-free survival compared with variant genotypes,104–106 According to the higher FCGR2A H131R and FCGR3A F158V frequencies among Africans (Table 11) than Caucasians,107 it can be hypothesized that African patients may not respond as well to trastuzumab treatment as non-African, though this has not been demonstrated and/or confirmed.
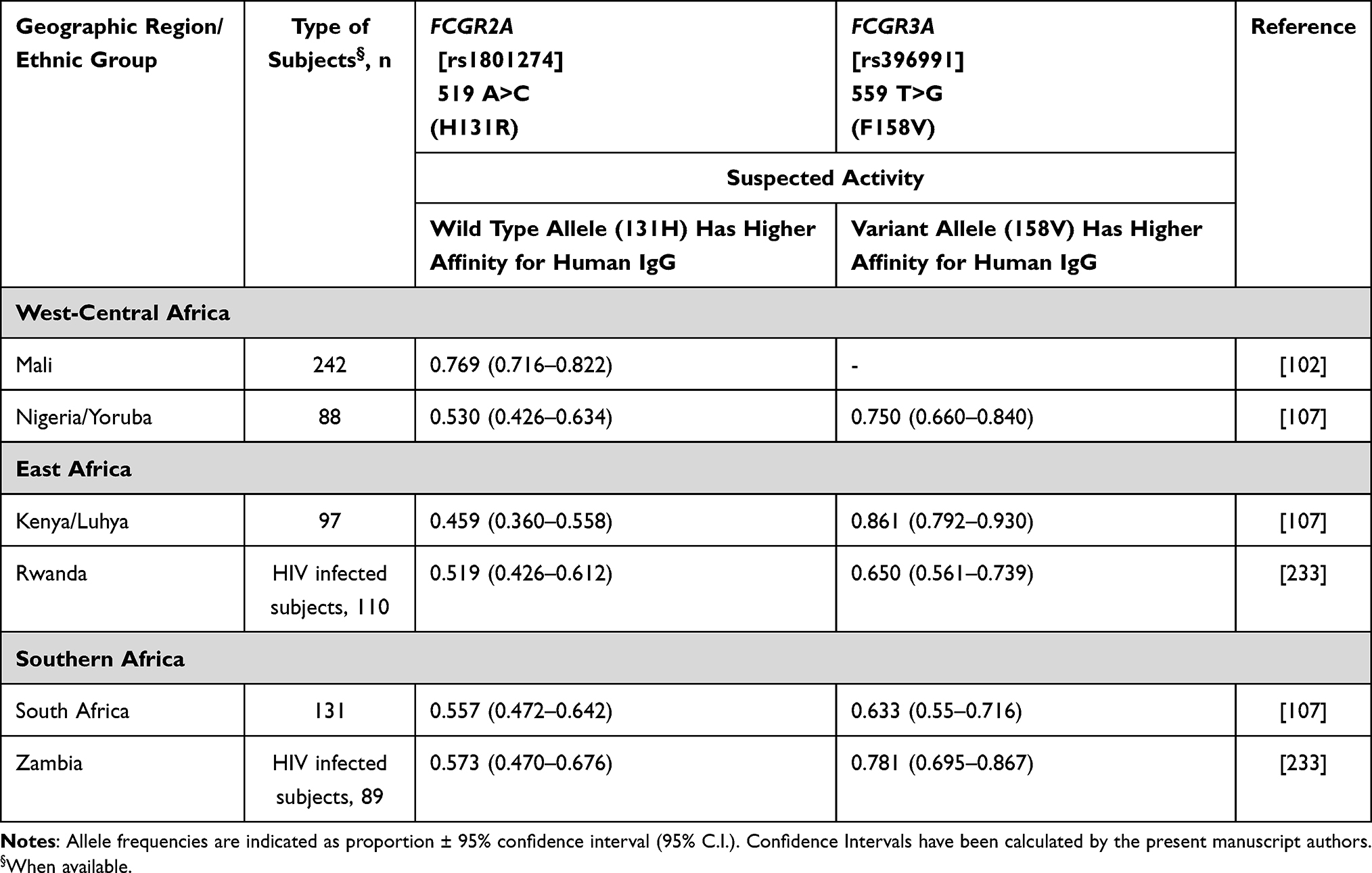 |
Table 11 FCG RECEPTORS Allele Frequency for Sub-Saharan Africa |
Another possible source of variation concerning trastuzumab and its targets, is the genetic variation at HER2 gene. Indeed, an association of rs1136201 (I655V, 1963 A>G) of HER2 with trastuzumab cardiotoxicity has been described108 where the allele G has been associated with response to trastuzumab. But in another study it has been found that there was no association between the aforementioned rs1136201 and toxicity; however, another polymorphism, rs1058808 (P1170A, 3418 C>G), was more likely to be found in cases with trastuzumab cardiotoxicity.109 In spite of this, the correlation between variations at the HER2 gene and cardiotoxicity risk have not been validated.110 Finally, no specific studies have been performed on SSA.
Cytotoxic Chemotherapy
HR+ tumours are not responsive to endocrine treatment and are therefore treated with chemotherapy. Cytotoxic drugs have a narrow therapeutic index, and there is substantial inter-patient variability in reaction to the administration of standard doses. As with the use of hormonal agents and other targeted therapies, pharmacogenetics may partly explain differences in the safety and efficacy of cytotoxic agents.
Antimetabolites
Capecitabine, an orally administered prodrug of the pyrimidine analog 5-fluorouracil (5-FU), is used frequently in the treatment of metastatic breast cancer.111 The rate of systemic 5-FU catabolism is the main determinant of capecitabine side effects, and it is dictated by the enzyme dihydropyrimidine-dehydrogenase (DPD), which is implicated in about 50% of cases of severe toxicity.112,113 Although capecitabine is not widely used in the African public health sector, reports show that it is being used in Nigeria and South Africa, at least.8,114,115
DPYD
DPD, encoded by DPYD gene, is the rate-limiting step in pyrimidine catabolism and deactivation of 5-FU.116 DPD is responsible for the degradation and removal of >80% of 5-FU administered to patients. Up to 7% of the overall populations carry a DPYD variant that reduces DPD enzyme activity, which causes excess drug accumulation and toxicity.116,117 Fluorouracil toxicity has a wide range of symptoms that include vomiting, nausea, kidney failure, and even death.118 Unlike the vast majority of other pharmacogenes described in this review, this association has been adequately confirmed, and there are prospective studies demonstrating improved clinical outcomes (decreased toxicity) in patients following a DPYD genotype-guided dose individualisation for fluoropyrimidine treatment.119,120 It is worth noting that DPD deficiency is a very rare condition (~1/200–1/1000) in which one has no DPD activity; however, a much larger percentage of patients carry a DPYD variant that reduces DPD activity (~7%). In fact, multiple DPYD variants have been identified, including well-known non-synonymous and splice site variations within the coding regions of the gene, and more novel variations within non-coding regions.121 Furthermore, reduced DPD activity may be partially due to epigenetic factors including gene promoter methylation.122 Among the variants associated with reduced DPD activity, DPYD*2A (rs3918290, intron mutation) displays similar allele frequencies across all ethnic groups tested, including Africans.123 DPYD*13 (rs55886062, 1679A>C) and 2846A>T are present in less than 2% of Caucasians.124 Importantly, in this review paper, a study125 searching for deleterious mutations in subjects of different ethnicities identified a non-synonymous variant, Y186C (rs115232898), that was found in heterozygosity in 6.4% of African Americans. The variant has been previously described among deficient African Americans122 and correlates with 5-FU toxicity.126,127 In vitro experiments demonstrated a 15–29% relative decrease in the activity of DYPD Y186C compared with wild-type DPYD.128 The DYPD Y186C appears to be nearly exclusive to African individuals,129 with an average frequency of 3% (Table 12).
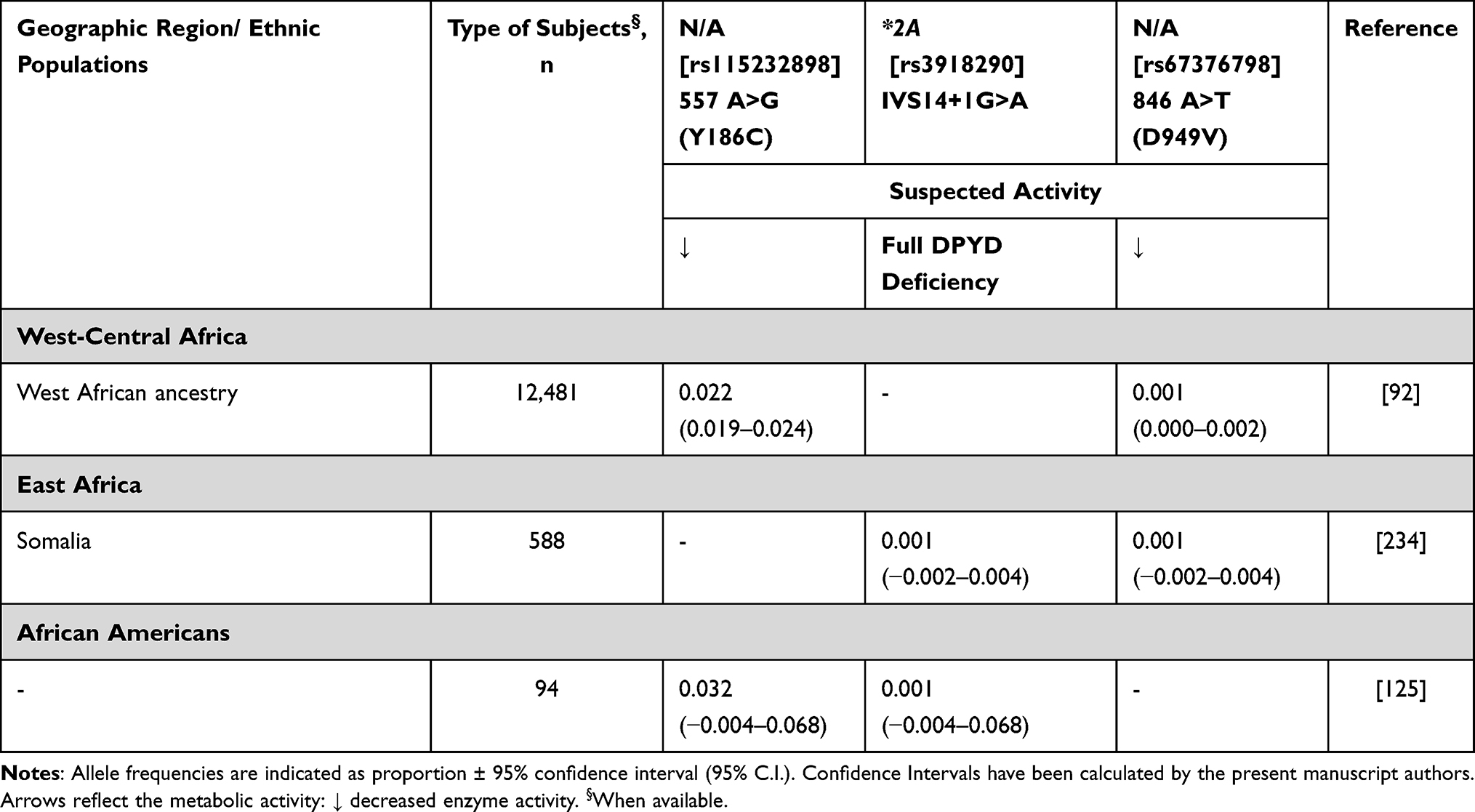 |
Table 12 DPYD Allele Frequency for Sub-Saharan Africa |
In summary, it is important to consider the pharmacogenomic risk of using fluoropyrimidine-based treatments in Africans. Variants with high predictive value for the onset of toxicity in Europeans were not observed in the African populations studied.130 African DPYD variants with predicted functional impacts, such as Y186C, should be validated and considered for inclusion in guidelines or testing strategies for African populations. Finally, additional research is needed to identify more variants that reduce DPD activity in Africans.
Antimicrotubules
The taxanes, paclitaxel and docetaxel, are some of the most effective chemotherapeutic agents against breast cancer and are indicated in both metastatic and adjuvant diseases. Taxanes disrupt microtubule depolymerization and spindle formation during cell replication, thereby causing cell death. Both paclitaxel and docetaxel are hydroxylated in the liver by CYP3A4/5, though paclitaxel is primarily metabolized by CYP2C8.131
Paclitaxel
Paclitaxel is a widely used drug for breast cancer treatment with an overall response rate of about 25%.132 The advanced paclitaxel formulation as albumin-embedded nanoparticles (nab paclitaxel) increased the initial response rate to 42%.132 However, resistance occurs frequently, and the evasion mechanisms remain unclear. Tumour recurrence occurs in 30% of node-negative, and up to 70% in node-positive breast cancer patients. Only 23% of relapsed patients survive 5 years after diagnosis, mainly due to metastasis to lymph nodes and distant organs.133 Paclitaxel is dosed according to body surface area and in most regimens infused for 3 hours. The dose limiting toxicities are neutropenia and neuropathy. However, there is a large interindividual variability in toxicity and therapeutic effects of paclitaxel, which remains a clinically relevant problem with implications on survival and quality of life of the patients. All this has a practical effect regarding the handling of dose delay, dose reduction or cessation of the treatment. Several possible causes of this variability have been suggested, including the possibility that SNPs in genes responsible for paclitaxel metabolism (ie, CYP2C8 and CYP3A4) could affect systemic exposure and toxicity.134 CYP2C8 is a polymorphic gene with several variant genotypes that may affect paclitaxel clearance.135 CYP2C8 comprises 7% of the total hepatic CYP450 content and plays an important role in the metabolism of a limited number of exogenous compounds. A clinical pharmacokinetic study demonstrated a reduction in paclitaxel metabolism associated with the CYP2C8*3 allele among Caucasians, and a modest reduction in paclitaxel metabolism with the CYP2C8*4 allele.136 Alternatively, another study reported that CYP2C8*3 may increase metabolic elimination of paclitaxel,137 as it does with most other substrates. Both of these alleles are found at lower frequencies in African populations, whereas CYP2C8*2 is found at higher frequencies.138–143 There is relatively little knowledge about the impact of CYP2C8*2 on paclitaxel metabolism. Some in vitro studies have found that CYP2C8*2 has a two-fold lower intrinsic clearance for paclitaxel than the wild-type,144–146 however, this has not been confirmed in a clinical pharmacokinetics study. Another study involving an African American breast cancer cohort found that the risk of paclitaxel-induced peripheral neuropathy was significantly greater in the CYP2C8 low-metabolizer group, which included all carriers of CYP2C8*2, CYP2C8*3, or CYP2C8*4.147 However, the influence of the CYP2C8*2 and CYP2C8*4 SNPs were not independently significant.147 Table 13 shows African frequency for CYP2C8 alleles, which does not suggest substantial high differences in metabolic activity phenotypes between Africans and non-Africans.
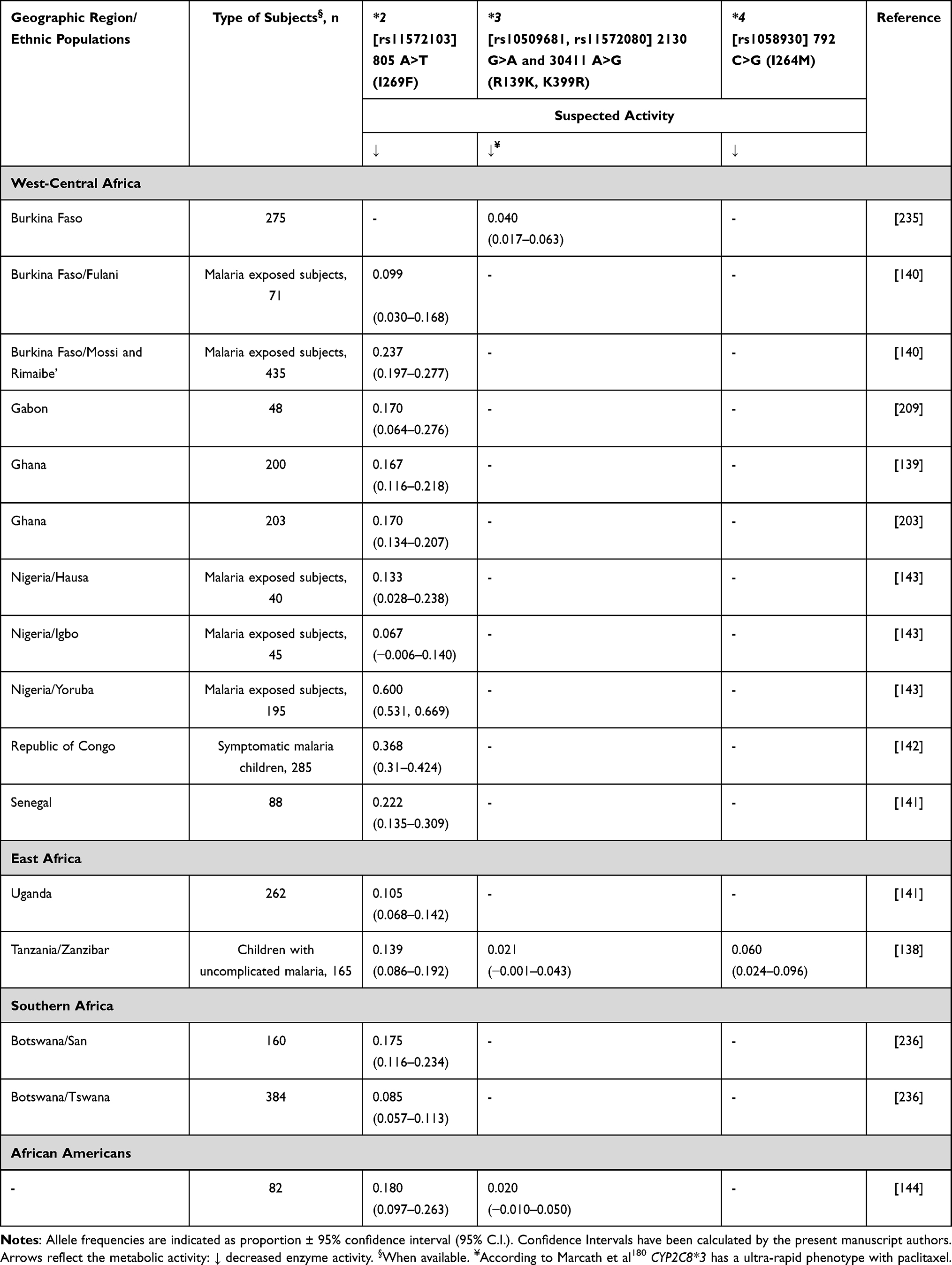 |
Table 13 CYP2C8 Allele Frequency in Sub-Saharan Africa |
CYP3A4 and CYP3A5 were previously discussed in the tamoxifen section. CYP3A4 alleles that affect enzyme activity, such as CYP3A4*22, may be associated with an increased risk of developing paclitaxel-induced peripheral neuropathy.148,149 However, no studies have been performed on African patient cohorts. Other non-pharmacogenes that have been reported to be associated with peripheral neuropathy have not been validated.150–152
Docetaxel
Genetics have not been demonstrated to have a meaningful effect docetaxel on pharmacokinetics or peripheral neuropathy.134 A GWAS reported a candidate polymorphism in VAC14 (rs875858),153 which is yet to be validated. To our knowledge, this SNP has not been described in African populations.
In the past decade, important new insights have also been obtained on polymorphic transporters involved in docetaxel elimination. Specifically, there is compelling evidence suggesting that hepatocellular uptake of taxanes from sinusoidal blood is regulated, at least in part, by the solute carrier OATP1B3 (encoded by SLCO1B3 gene) but without evidence of the impact of its genetic variation on docetaxel elimination.154,155 The secretion of taxanes from the liver into the bile, instead, depends on the ATP-binding cassette (ABC) transporters ABCB1 (P-glycoprotein) and ABCC2 (also called MRP2). There is some evidence of a possible effect of ABCB1 rs1045642 (3435C>T) on docetaxel plasma levels156 and toxicity,157,158 and of rs1202179 (287-4740G>A) on chemotherapy-induced alopecia.159 Although there are no association studies conducted in African cohorts, some of the variants of interest are found in African populations (Table 14).
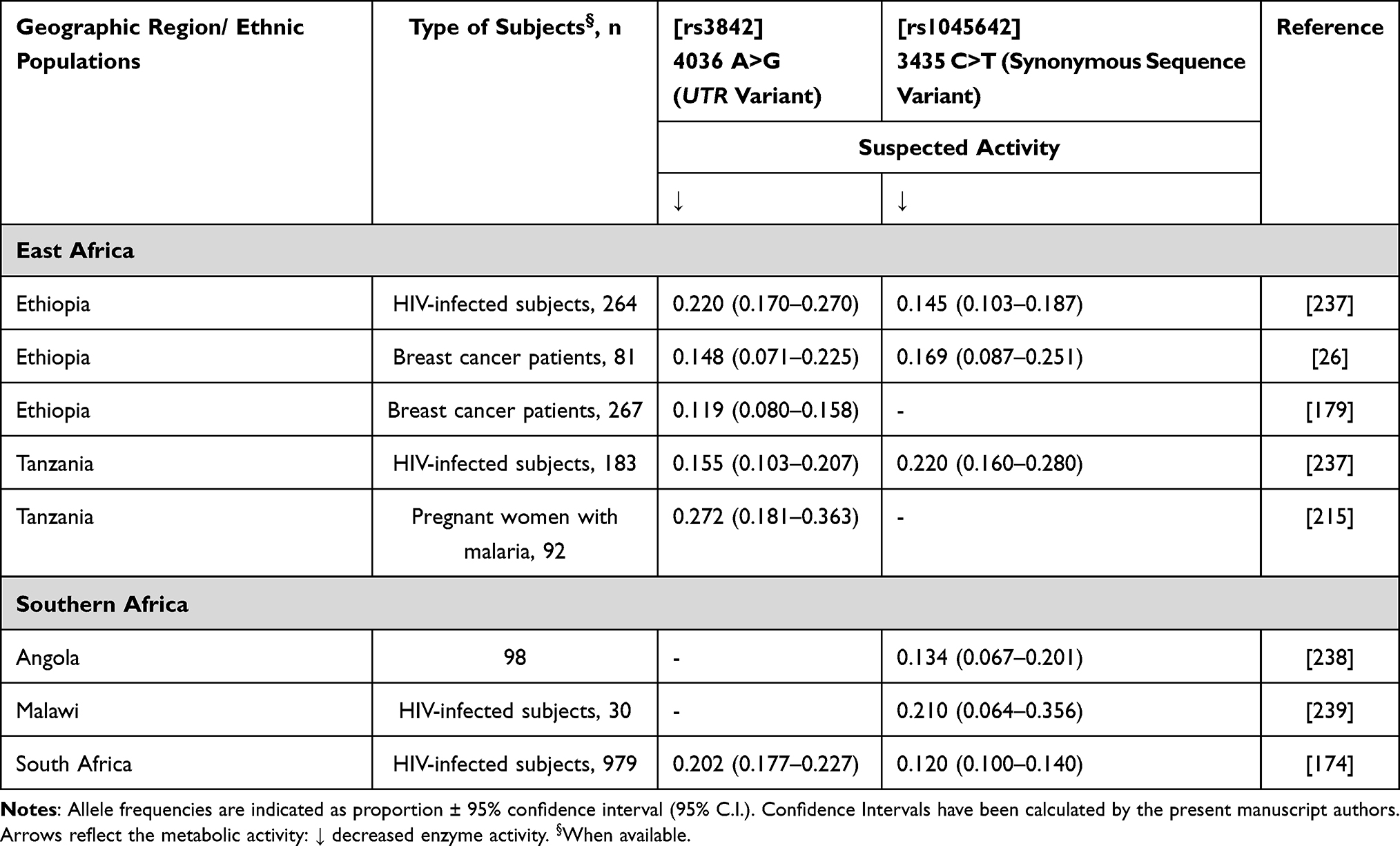 |
Table 14 ABCB1 Allele Frequency in Sub-Saharan Africa |
Anthracyclines
The anthracyclines doxorubicin and epirubicin have been widely used in breast cancer treatment for several decades, including within Africa.160–162 These drugs inhibit topoisomerase II and thereby induce cellular apoptosis. Pharmacogenetic variants have been observed in anthracycline-metabolizing enzymes, as well as transporters and proteins involved in oxidative stress and apoptosis.163–165
Approximately 50% of doxorubicin is eliminated from the body in its intact form, and the remainder through aldoketoreductase (AKR1A1) and carbonyl reductase (CBR1 and CBR3),166 with a minor contribution from NADH dehydrogenase (NQO1) and nitric oxide synthases (NOS1, NOS2 and NOS3).166 Variants in CBR1 and CBR3 have been associated with doxorubicin pharmacokinetics and clinical outcomes in paediatric cancer survivors, but these associations were not found in patients with breast cancer.167,168 Recent data suggest that CBR1 rs20572 and AKR1A1 rs2088102 might be protective factors for the hematologic toxicity during anthracycline-based chemotherapy in breast cancer patients.169 Unfortunately, rs20572 has a very low frequency among Africans, suggesting a possible, though not yet studied, explanation for the increased risk of hematologic toxicity in African patients.169 Data on AKR1A1 rs2088102 among Africans have been reported to be 0.514.92
SLC22A16
SLC22A16 encodes an organic zwitterion transporter protein that transports carnitine and some anticancer drugs, including anthracyclines.170 Carriers of the SLC22A16 rs12210538 (1226 T>C) allele have been reported to have a higher incidence of leucopenia and dose delay but have no difference in survival.171 SLC22A16 expression in cancer cells is associated with increased sensitivity to the cytotoxic effects of doxorubicin.163 Patients with the variant genotype may have greater uptake of doxorubicin into normal and tumour cells, leading to greater incidence of toxicity. Twenty–twenty five percent (20–25%) of Caucasians carry the variant,172 while allele frequency in Africans has been measured as 0.038.92
ABCB1
ABCB1 is a drug transporter that effluxes drugs from malignant cells. In a study investigating 68 white European women with locally advanced breast cancer, there was a greater likelihood of clinically complete response to neoadjuvant chemotherapy in patients carrying ABCB1 3435C>T.173 No data were retrieved concerning African patients using anthracyclines. See Table 14 for African ABCB1 allele frequencies. Importantly, ABCB1 3435C>T is higher in Caucasians and Asians (50–60%) than Africans.174 ABCB1 4036A>G frequencies seem to be comparable in Africans to Caucasians, which are somewhat lower than the frequency in Asians.174
Cyclophosphamide
Cyclophosphamide remains a stable component in many of the chemotherapy combinations used in breast cancer patients in SSA. Cyclophosphamide is a prodrug that undergoes metabolic activation, in the liver, which is primarily mediated by CYP2B6 and CYP2C9/CYP2C19, with a minor contribution from CYP3A4/5.175 The active metabolite aldophosphamide is subsequently inactivated by glutathione-s-transferases (GSTs).165
Xie et al176 showed that CYP2B6 516G>T has two-fold higher cyclophosphamide clearance in vitro compared to wild-type enzyme. CYP2B6 516G>T is a poor metabolizer polymorphism for the antiretroviral drugs efavirenz and nevirapine, with very high frequency in Africa,46 however showing a higher cyclophosphamide clearance rate than wild-type CYP2B6.176 However, several other studies have not found the effects of polymorphisms in CYP2B6 and CYP2C19 on cyclophosphamide pharmacokinetics.177,178 Data concerning African frequencies for CYP2B6 variants are shown in Table 15. In Ethiopia, it has been reported that carriers of CYP3A5*3 and CYP3A5*6 had lower cyclophosphamide elimination rate and longer half-life than subjects carrying the wild-type allele.179 Additionally, increased drug clearance has been reported in carriers of CYP2C9*2 and CYP2C9*3,179 similar to the increased activity for other substrates.180 Allele frequencies for these genes were previously reported in other sections.
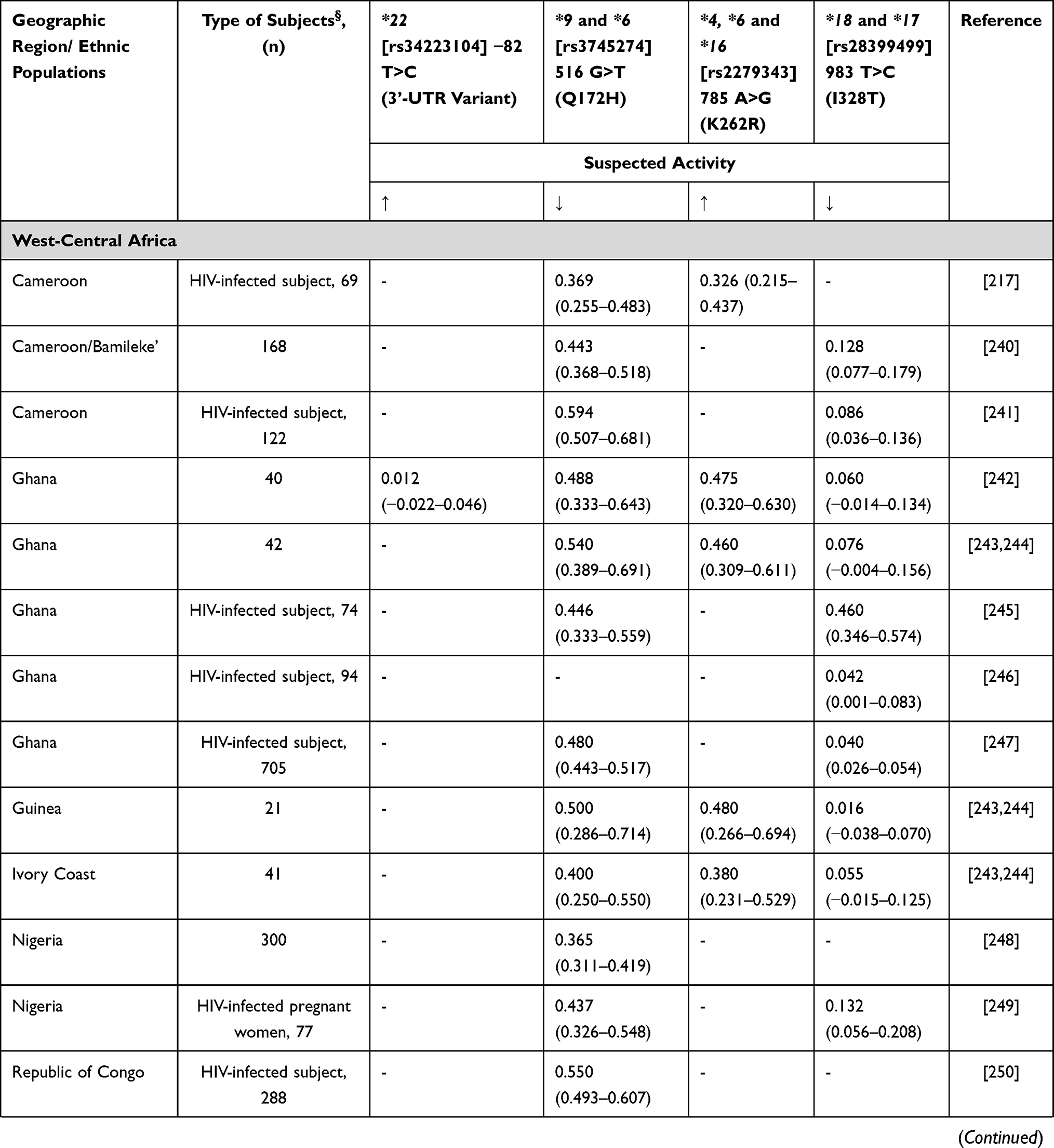 | 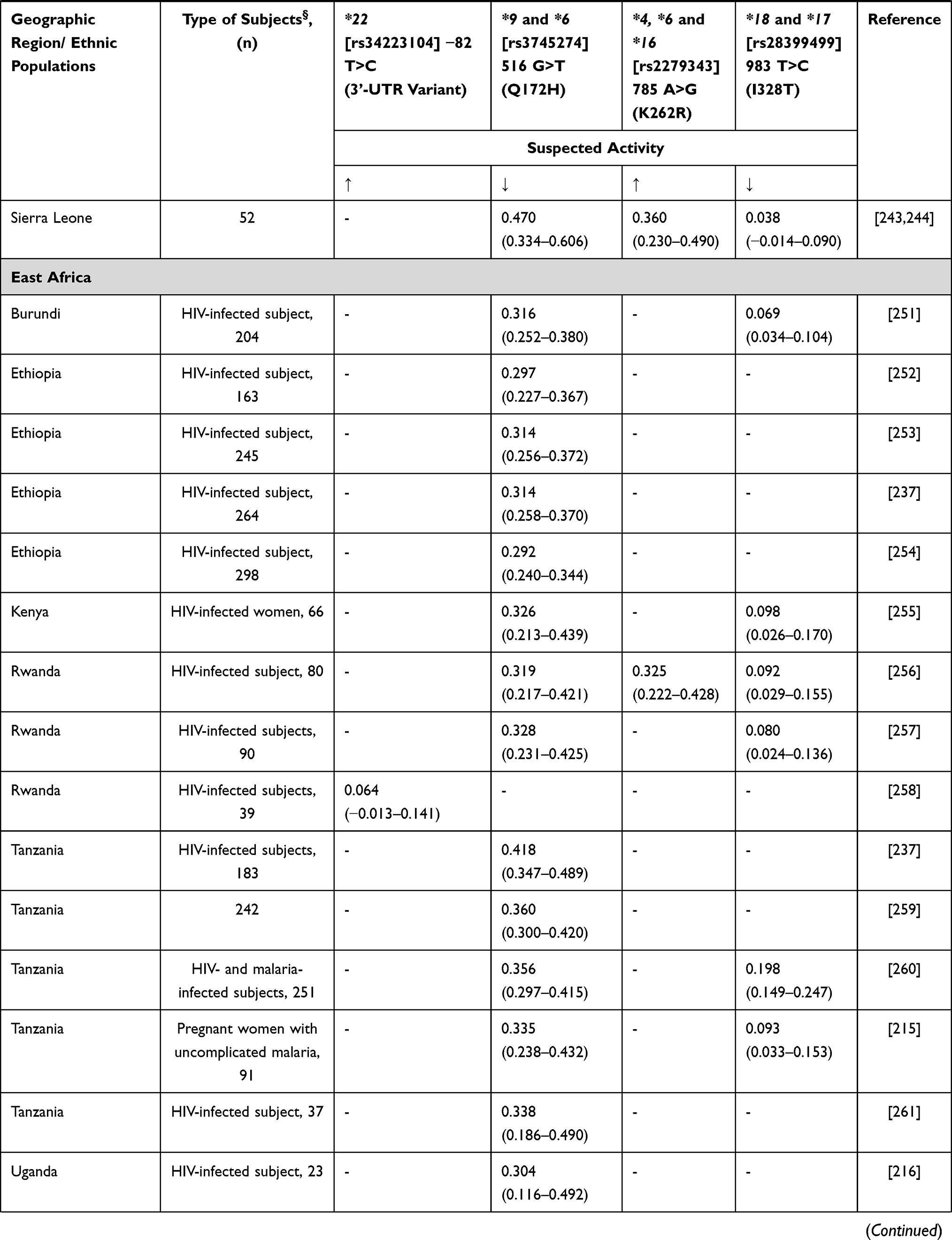 | 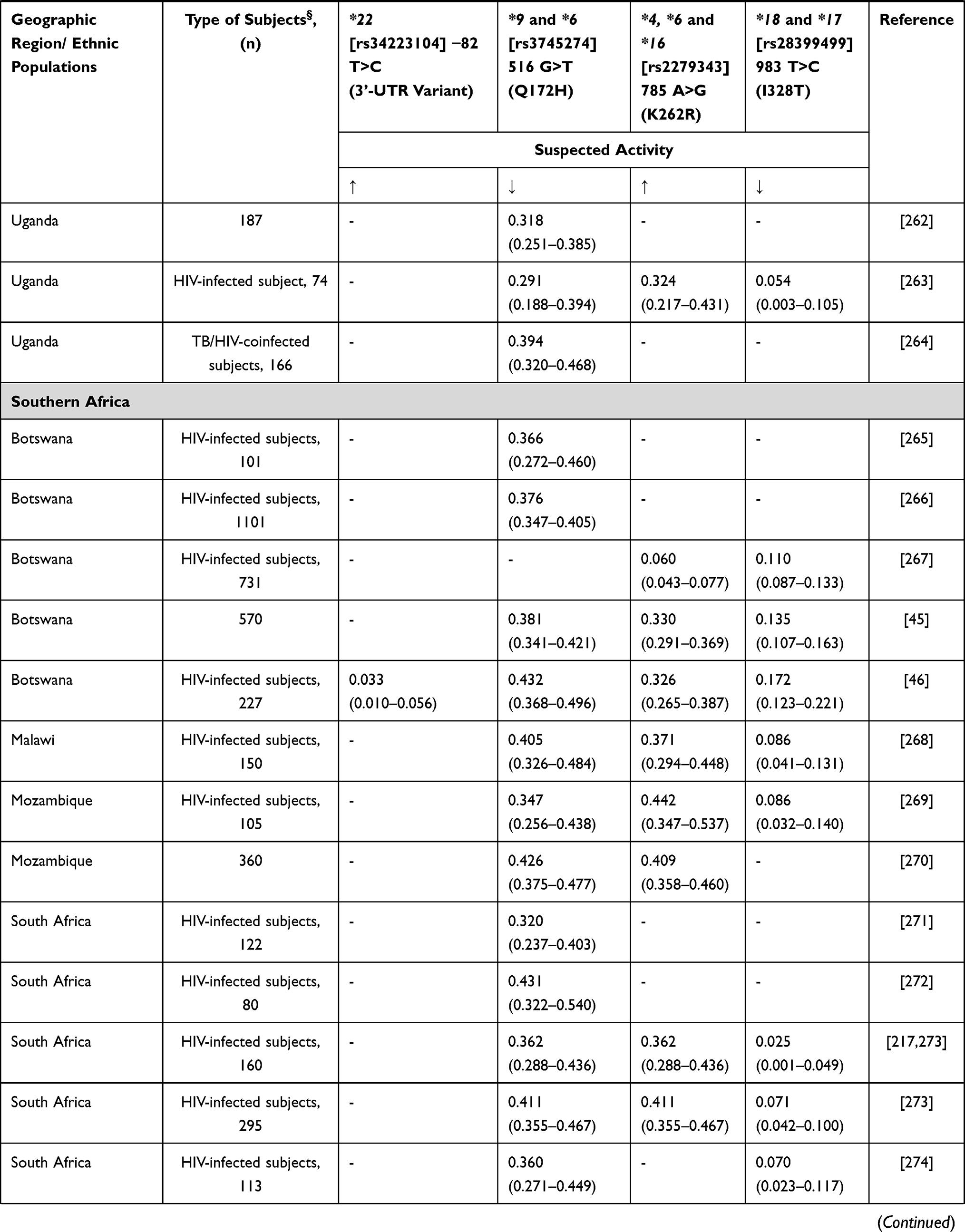 |  |
Table 15 CYP2B6 Allele Frequency in Sub-Saharan Africa |
Conclusion
This review paper summarized the findings with reference to the African variability of genes encoding for enzymes and transporters involved in the metabolism of drugs available to treat breast cancer in Africa. The high extent of diversification shown by African populations and ethnic groups is an example of adaptation and co-evolution between genetic loci deputed to the detoxification of exogenous molecules from the body, linked to different lifestyles in humans. This existing system is exploited by modern drugs, resulting in different pathways and rates of drug metabolism consistent with the extent of African genetic variability.
Most of the reported data for African alleles and/or SNP frequencies come from studies concerning malaria and HIV, the two main infectious diseases affecting the continent. This limits the availability of data for non-malaria and non-HIV treatment pharmacogenetics. Moreover, despite a growing interest and concern for non-communicable diseases, pharmacogenetics, and in particular breast cancer pharmacogenetics, is still a developing field in Africa. Indeed, some of the data reported in this paper are derived from studies conducted on African American subjects or patients, used as a proxy for Africans.
Only a small number of published studies have investigated the pharmacogenetics but also the pharmacokinetic/pharmacodynamic profile of drugs used to treat breast cancer in SSA. In particular, two studies, one on tamoxifen and another on cyclophosphamide among breast cancer patients, have been performed in Ethiopia.26,179 Another study was focused on AIs in South African women with breast cancer.74 We would like to stress that Ethiopia has a complex admixture component with a dynamic history of several Eurasian ancestries and some Nilotic and Semitic-Cushitic components. Similarly, South African ethnic diversity and admixture may hamper the transfer of these outputs to other African populations with different genetic backgrounds.
The data shown in this paper can be used to establish priorities in investigations but in no case be considered in the management of patients until pharmacogenetic studies have been carried out in the considered population. In fact, it is important to establish priority research in genetics of drug metabolism and transport because of the extent of breast cancer in all SSA. For example, because tamoxifen is used extensively in all settings, more research on this drug should be conducted in order to fill the gap in information about possible clinical outcomes among Africans, not yet demonstrated in other world populations. Additionally, DPYD genotyping for capecitabine may be clinically useful. More studies are needed to identify other low-activity alleles in African populations and to demonstrate the clinical benefit of pre-treatment DPYD genotyping to inform capecitabine treatment.
Several pharmacogenes (including transporters) show a non-negligible frequency for several African alleles, mostly poor metaboliser alleles. Among them, the CYP2/3 families show alleles with high to very high frequencies. There is a clear need for more studies to ascertain the possible risk and/or benefit of a specific treatment in that particular ethnic population, in an effort to maximise therapeutic output and survival rate for the women affected by breast cancer.
Funding
This work was supported through the Internal Research Round 39 Funding Award from the University of Botswana and by the Penn Center for AIDS Research [grant # P30AI045008].
Disclosure
Dr Daniel Louis Hertz is an unpaid scientific advisor to Advocates for Universal DPD/DPYD Testing (AUDT). The authors report no conflicts of interest in this work.
References
1. Hertz DL, McLeod HL. Use of pharmacogenetics for predicting cancer prognosis and treatment exposure, response and toxicity. J Hum Genet. 2013;58(6):346–352. doi:10.1038/jhg.2013.42
2. Pusztai L, Stec J, Ayers M, et al. Pharmacogenetics, pharmacogenomics, and predicting response to therapy. In: Ross JS, Hortobagyi GN, editors. Molecular Oncology of Breast Cancer. Sudbury: Jones and Bartlett Publishers; 2005:439–456.
3. O’Donnell PH, Dolan ME. Cancer pharmacoethnicity: ethnic differences in susceptibility to the effects of chemotherapy. Clin Cancer Res. 2009;15(15):4806–4814. doi:10.1158/1078-0432.CCR-09-0344
4. Allen C, Allen C, Allen C, et al.; Global Burden of Disease Cancer Collaboration. Global, regional, and national cancer incidence, mortality, years of life lost, years lived with disability, and disability-adjusted life-years for 32 cancer groups, 1990 to 2015: a systematic analysis for the global burden of disease study [published correction appears in JAMA Oncol. 2017 Mar 1;3(3):418]. JAMA Oncol. 2017;3(4):524–548. doi:10.1001/jamaoncol.2016.5688
5. Azubuike SO, Muirhead C, Hayes L, McNally R. Rising global burden of breast cancer: the case of sub-Saharan Africa (with emphasis on Nigeria) and implications for regional development: a review. World J Surg Oncol. 2018;16(1):63. doi:10.1186/s12957-018-1345-2
6. Jedy-Agba E, McCormack V, Adebamowo C, et al. Stage at diagnosis of breast cancer in sub-Saharan Africa: a systematic review and meta-analysis. Lancet Glob Health. 2016;4(12):e923–e935. doi:10.1016/S2214-109X(16)30259-5
7. Martinez-Cannon BA, Barragan-Carrillo R, Villarreal-Garza C. Young women with breast cancer in resource-limited settings: what we know and what we need to do better. Breast Cancer. 2021;13:641–650. doi:10.2147/BCTT.S303047
8. Vanderpuye V, Grover S, Hammad N, et al. An update on the management of breast cancer in Africa. Infect Agent Cancer. 2017;12:13. doi:10.1186/s13027-017-0124-y
9. Pace LE, Shulman LN. Breast Cancer in Sub-Saharan Africa: challenges and Opportunities to Reduce Mortality. Oncologist. 2016;21(6):739–744. doi:10.1634/theoncologist.2015-0429
10. Murphy BL, Day CN, Hoskin TL, Habermann EB, Boughey JC. Adolescents and young adults with breast cancer have more aggressive disease and treatment than patients in their forties. Ann Surg Oncol. 2019;26(12):3920–3930. doi:10.1245/s10434-019-07653-9
11. Akarolo-Anthony SN, Ogundiran TO, Adebamowo CA. Emerging breast cancer epidemic: evidence from Africa. Breast Cancer Res. 2010;12(Suppl4):S8. doi:10.1186/bcr2737
12. Kantelhardt EJ, Mathewos A, Aynalem A, et al. The prevalence of estrogen receptor-negative breast cancer in Ethiopia. BMC Cancer. 2014;14:895. doi:10.1186/1471-2407-14-895
13. Sayed S, Moloo Z, Wasike R, et al. Is breast cancer from Sub Saharan Africa truly receptor poor? Prevalence of ER/PR/HER2 in breast cancer from Kenya. Breast. 2014;23(5):591–596. doi:10.1016/j.breast.2014.06.006
14. Gomez F, Hirbo J, Tishkoff SA. Genetic variation and adaptation in Africa: implications for human evolution and disease. Cold Spring Harb Perspect Biol. 2014;6(7):a008524. doi:10.1101/cshperspect.a008524
15. Choudhury A, Aron S, Botigué LR, et al. High-depth African genomes inform human migration and health [published correction appears in Nature. 2021 Apr;592(7856):E26]. Nature. 2020;586(7831):741–748. doi:10.1038/s41586-020-2859-7
16. Drögemöller B, Plummer M, Korkie L, et al. Characterization of the genetic variation present in CYP3A4 in three South African populations. Front Genet. 2013;4:17. doi:10.3389/fgene.2013.00017
17. Podgorná E, Diallo I, Vangenot C, et al. Variation in NAT2 acetylation phenotypes is associated with differences in food-producing subsistence modes and ecoregions in Africa. BMC Evol Biol. 2015;15:263. doi:10.1186/s12862-015-0543-6
18. Bissonauth V, Shatenstein B, Ghadirian P. Nutrition and breast cancer among sporadic cases and gene mutation carriers: an overview. Cancer Detect Prev. 2008;32(1):52–64. doi:10.1016/j.cdp.2008.01.005
19. Rim A, Chellman-Jeffers M, Fanning A. Trends in breast cancer screening and diagnosis. Cleve Clin J Med. 2008;75(Suppl 1):S2–S9. doi:10.3949/ccjm.75.suppl_1.s2
20. Adebamowo CA, Famooto A, Ogundiran TO, Aniagwu T, Nkwodimmah C, Akang EE. Immunohistochemical and molecular subtypes of breast cancer in Nigeria. Breast Cancer Res Treat. 2008;110(1):183–188. doi:10.1007/s10549-007-9694-5
21. Cubasch H, Joffe M, Hanisch R, et al. Breast cancer characteristics and HIV among 1092 women in Soweto, South Africa. Breast Cancer Res Treat. 2013;140(1):177–186. doi:10.1007/s10549-013-2606-y
22. Kakudji BK, Mwila PK, Burger JR, du Plessis JM, Naidu K. Breast cancer molecular subtypes and receptor status among women at Potchefstroom Hospital: a cross-sectional study. Pan Afr Med J. 2021;38:85. doi:10.11604/pamj.2021.38.85.23039
23. Twahir M, Oyesegun R, Yarney J, et al. Real-world challenges for patients with breast cancer in sub-Saharan Africa: a retrospective observational study of access to care in Ghana, Kenya and Nigeria. BMJ Open. 2021;11(3):e041900. doi:10.1136/bmjopen-2020-041900
24. Jordan VC. Tamoxifen: a most unlikely pioneering medicine. Nat Rev Drug Discov. 2003;2(3):205–213. doi:10.1038/nrd1031
25. Schroth W, Goetz MP, Hamann U, et al. Association between CYP2D6 polymorphisms and outcomes among women with early stage breast cancer treated with tamoxifen. JAMA. 2009;302(13):1429–1436. doi:10.1001/jama.2009.1420
26. Ahmed JH, Makonnen E, Fotoohi A, Aseffa A, Howe R, Aklillu E. CYP2D6 genotype predicts plasma concentrations of tamoxifen metabolites in Ethiopian breast cancer patients. Cancers. 2019;11(9):1353. doi:10.3390/cancers11091353
27. Helland T, Alsomairy S, Lin C, Søiland H, Mellgren G, Hertz DL. Generating a precision endoxifen prediction algorithm to advance personalized tamoxifen treatment in patients with breast cancer. J Pers Med. 2021;11(3):201. doi:10.3390/jpm11030201
28. Crewe HK, Ellis SW, Lennard MS, Tucker GT. Variable contribution of cytochromes P450 2D6, 2C9 and 3A4 to the 4-hydroxylation of tamoxifen by human liver microsomes. Biochem Pharmacol. 1997;53(2):171–178. doi:10.1016/s0006-2952(96)00650-8
29. de Groot MJ, Bijloo GJ, van Acker FA, Fonseca Guerra C, Snijders JG, Vermeulen NP. Extension of a predictive substrate model for human cytochrome P4502D6. Xenobiotica. 1997;27(4):357–368. doi:10.1080/004982597240514
30. Ingelman-Sundberg M. Genetic polymorphisms of cytochrome P450 2D6 (CYP2D6): clinical consequences, evolutionary aspects and functional diversity. Pharmacogenomics J. 2005;5(1):6–13. doi:10.1038/sj.tpj.6500285
31. Goetz MP, Rae JM, Suman VJ, et al. Pharmacogenetics of tamoxifen biotransformation is associated with clinical outcomes of efficacy and hot flashes. J Clin Oncol. 2005;23(36):9312–9318. doi:10.1200/JCO.2005.03.3266
32. Hertz DL, McLeod HL, Irvin WJ. Tamoxifen and CYP2D6: a contradiction of data. Oncologist. 2012;17(5):620–630. doi:10.1634/theoncologist.2011-0418
33. Regan MM, Leyland-Jones B, Bouzyk M, et al. CYP2D6 genotype and tamoxifen response in postmenopausal women with endocrine-responsive breast cancer: the breast international group 1–98 trial. J Natl Cancer Inst. 2012;104(6):441–451. doi:10.1093/jnci/djs125
34. Hertz DL, Kidwell KM, Hilsenbeck SG, et al. CYP2D6 genotype is not associated with survival in breast cancer patients treated with tamoxifen: results from a population-based study. Breast Cancer Res Treat. 2017;166(1):277–287. doi:10.1007/s10549-017-4400-8
35. Aklillu E, Herrlin K, Gustafsson LL, Bertilsson L, Ingelman-Sundberg M. Evidence for environmental influence on CYP2D6-catalysed debrisoquine hydroxylation as demonstrated by phenotyping and genotyping of Ethiopians living in Ethiopia or in Sweden. Pharmacogenetics. 2002;12(5):375–383. doi:10.1097/00008571-200207000-00005
36. Gaedigk A, Sangkuhl K, Whirl-Carrillo M, Klein T, Leeder JS. Prediction of CYP2D6 phenotype from genotype across world populations [published correction appears in Genet Med. 2016;18(11):1167]. Genet Med. 2017;19(1):69–76. doi:10.1038/gim.2016.80
37. Koopmans AB, Braakman MH, Vinkers DJ, Hoek HW, van Harten PN. Meta-analysis of probability estimates of worldwide variation of CYP2D6 and CYP2C19. Transl Psychiatry. 2021;11(1):141. doi:10.1038/s41398-020-01129-1
38. Alessandrini M, Asfaha S, Dodgen TM, Warnich L, Pepper MS. Cytochrome P450 pharmacogenetics in African populations. Drug Metab Rev. 2013;45(2):253–275. doi:10.3109/03602532.2013.783062
39. Rajman I, Knapp L, Morgan T, Masimirembwa C. African genetic diversity: implications for cytochrome p450-mediated drug metabolism and drug development. EBioMedicine. 2017;17:67–74. doi:10.1016/j.ebiom.2017.02.017
40. Gaedigk A, Coetsee C. The CYP2D6 gene locus in South African Coloureds: unique allele distributions, novel alleles and gene arrangements. Eur J Clin Pharmacol. 2008;64(5):465–475. doi:10.1007/s00228-007-0445-7
41. Dodgen TM, Labuschagne CJ, van Schalkwyk A, et al. Pharmacogenetic comparison of CYP2D6 predictive and measured phenotypes in a South African cohort [published correction appears in Pharmacogenomics J. 2017 Jul;17 (4): 393]. Pharmacogenomics J. 2016;16(6):566–572. doi:10.1038/tpj.2015.76
42. Gaedigk A, Simon SD, Pearce RE, Bradford LD, Kennedy MJ, Leeder JS. The CYP2D6 activity score: translating genotype information into a qualitative measure of phenotype. Clin Pharmacol Ther. 2008;83(2):234–242. doi:10.1038/sj.clpt.6100406
43. Caudle KE, Sangkuhl K, Whirl-Carrillo M, et al. Standardizing CYP2D6 genotype to phenotype translation: consensus recommendations from the clinical pharmacogenetics implementation consortium and Dutch Pharmacogenetics Working Group. Clin Transl Sci. 2020;13(1):116–124. doi:10.1111/cts.12692
44. Kumondai M, Hosono H, Orikasa K, et al. Genetic polymorphisms of CYP2A6 in a case-control study on bladder cancer in Japanese smokers. Biol Pharm Bull. 2016;39(1):84–89. doi:10.1248/bpb.b15-00604
45. Tawe L, Motshoge T, Ramatlho P, et al. Human cytochrome P450 2B6 genetic variability in Botswana: a case of haplotype diversity and convergent phenotypes. Sci Rep. 2018;8(1):4912. doi:10.1038/s41598-018-23350-1
46. Maseng MJ, Tawe L, Thami PK, et al. Association of CYP2B6 genetic variation with efavirenz and nevirapine drug resistance in HIV-1 patients from Botswana [published correction appears in Pharmgenomics Pers Med. 2021 Mar 31;14:395]. Pharmgenomics Pers Med. 2021;14:335–347. doi:10.2147/PGPM.S289471
47. Dodgen TM, Drögemöller BI, Wright GE, et al. Evaluation of predictive CYP2C19 genotyping assays relative to measured phenotype in a South African cohort. Pharmacogenomics. 2015;16(12):1343–1354. doi:10.2217/pgs.15.80
48. LLerena A, Naranjo ME, Rodrigues-Soares F, Penas-LLedó EM, Fariñas H, Tarazona-Santos E. Interethnic variability of CYP2D6 alleles and of predicted and measured metabolic phenotypes across world populations. Expert Opin Drug Metab Toxicol. 2014;10(11):1569–1583. doi:10.1517/17425255.2014.964204
49. Damkier P, Kjærsgaard A, Barker KA, et al. CYP2C19*2 and CYP2C19*17 variants and effect of tamoxifen on breast cancer recurrence: analysis of the International Tamoxifen Pharmacogenomics Consortium dataset. Sci Rep. 2017;7(1):7727. doi:10.1038/s41598-017-08091-x
50. Powers JL, Buys SS, Fletcher D, et al. Multigene and drug interaction approach for tamoxifen metabolite patterns reveals possible involvement of CYP2C9, CYP2C19, and ABCB1. J Clin Pharmacol. 2016;56(12):1570–1581. doi:10.1002/jcph.771
51. Hansten PD. The underrated risks of tamoxifen drug interactions. Eur J Drug Metab Pharmacokinet. 2018;43(5):495–508. doi:10.1007/s13318-018-0475-9
52. Chen Y, Marcath LA, Eliassen FM, et al. Effect of genetic variability in 20 pharmacogenes on concentrations of tamoxifen and its metabolites. J Pers Med. 2021;11(6):507. doi:10.3390/jpm11060507
53. Limdi NA, Brown TM, Yan Q, et al. Race influences warfarin dose changes associated with genetic factors. Blood. 2015;126(4):539–545. doi:10.1182/blood-2015-02-627042
54. Sanchez-Spitman AB, Swen JJ, Dezentjé VO, Moes DJ, Gelderblom H, Guchelaar HJ. Effect of CYP2C19 genotypes on tamoxifen metabolism and early-breast cancer relapse. Sci Rep. 2021;11(1):415. doi:10.1038/s41598-020-79972-x
55. Sim SC, Risinger C, Dahl ML, et al. A common novel CYP2C19 gene variant causes ultrarapid drug metabolism relevant for the drug response to proton pump inhibitors and antidepressants. Clin Pharmacol Ther. 2006;79(1):103–113. doi:10.1016/j.clpt.2005.10.002
56. Sim S, Lövrot J, Lindh JD, Bergh J, Xie H. Effect of CYP2C19 and CYP2D6 genotype on tamoxifen treatment outcome indicates endogenous and exogenous interplay. Pharmacogenomics. 2018;19(13):1027–1037. doi:10.2217/pgs-2018-0089
57. Cavaco I, Reis R, Gil JP, Ribeiro V. CYP3A4*1B and NAT2*14 alleles in a native African population. Clin Chem Lab Med. 2003;41(4):606–609. doi:10.1515/CCLM.2003.091
58. Lamba JK, Lin YS, Schuetz EG, Thummel KE. Genetic contribution to variable human CYP3A-mediated metabolism. Adv Drug Deliv Rev. 2002;54(10):1271–1294. doi:10.1016/s0169-409x(02)00066-2
59. Wojnowski L, Hustert E, Klein K, et al. Re: modification of clinical presentation of prostate tumors by a novel genetic variant in CYP3A4. J Natl Cancer Inst. 2002;94:
60. Zhou Y, Ingelman-Sundberg M, Lauschke VM. Worldwide distribution of cytochrome P450 alleles: a meta-analysis of population-scale sequencing projects. Clin Pharmacol Ther. 2017;102(4):688–700. doi:10.1002/cpt.690
61. Sanchez-Spitman AB, Moes DJ, Gelderblom H, Dezentje VO, Swen JJ, Guchelaar HJ. Effect of CYP3A4*22, CYP3A5*3, and CYP3A combined genotypes on tamoxifen metabolism. Eur J Clin Pharmacol. 2017;73(12):1589–1598. doi:10.1007/s00228-017-2323-2
62. Wegman P, Elingarami S, Carstensen J, Stål O, Nordenskjöld B, Wingren S. Genetic variants of CYP3A5, CYP2D6, SULT1A1, UGT2B15 and tamoxifen response in postmenopausal patients with breast cancer. Breast Cancer Res. 2007;9(1):R7. doi:10.1186/bcr1640
63. Bains RK. African variation at cytochrome P450 genes: evolutionary aspects and the implications for the treatment of infectious diseases. Evol Med Public Health. 2013;2013(1):118–134. doi:10.1093/emph/eot010
64. Lee JS, Cheong HS, Kim LH, et al. Screening of genetic polymorphisms of CYP3A4 and CYP3A5 genes. Korean J Physiol Pharmacol. 2013;17(6):479–484. doi:10.4196/kjpp.2013.17.6.479
65. Nowell S, Sweeney C, Winters M, et al. Association between sulfotransferase 1A1 genotype and survival of breast cancer patients receiving tamoxifen therapy. J Natl Cancer Inst. 2002;94(21):1635–1640. doi:10.1093/jnci/94.21.1635
66. Fernández-Santander A, Gaibar M, Novillo A, et al. Relationship between genotypes Sult1a2 and Cyp2d6 and tamoxifen metabolism in breast cancer patients. PLoS One. 2013;8(7):e70183. doi:10.1371/journal.pone.0070183
67. Sanchez-Spitman AB, Dezentjé VO, Swen JJ, Moes DJ, Gelderblom H, Guchelaar HJ. Genetic polymorphisms of 3’-untranslated region of SULT1A1 and their impact on tamoxifen metabolism and efficacy. Breast Cancer Res Treat. 2018;172(2):401–411. doi:10.1007/s10549-018-4923-7
68. Yang J, Shen J, Tang S, et al. The outcomes of tamoxifen therapy in breast cancer patients and genotypes of SULT1A1 and glucuronosyltransferase. Int J Clin Exp Med. 2017;10(4):6273–6282.
69. Romero-Lorca A, Novillo A, Gaibar M, Bandrés F, Fernández-Santander A. Impacts of the glucuronidase genotypes UGT1A4, UGT2B7, UGT2B15 and UGT2B17 on tamoxifen metabolism in breast cancer patients [published correction appears in PLoS One. 2015;10(10):e0140921]. PLoS One. 2015;10(7):e0132269. doi:10.1371/journal.pone.0132269
70. Blevins-Primeau AS, Sun D, Chen G, et al. Functional significance of UDP-glucuronosyltransferase variants in the metabolism of active tamoxifen metabolites. Cancer Res. 2009;69(5):1892–1900. doi:10.1158/0008-5472.CAN-08-3708
71. Dowsett M, Cuzick J, Ingle J, et al. Meta-analysis of breast cancer outcomes in adjuvant trials of aromatase inhibitors versus tamoxifen. J Clin Oncol. 2010;28(3):509–518. doi:10.1200/JCO.2009.23.1274
72. Kemfang Ngowa JD, Yomi J, Kasia JM, Mawamba Y, Ekortarh AC, Vlastos G. Breast cancer profile in a group of patients followed up at the radiation therapy unit of the Yaounde General Hospital, Cameroon. Obstet Gynecol Int. 2011;2011:143506. doi:10.1155/2011/143506
73. Baatjes KJ, Conradie M, Apffelstaedt JP, Kotze MJ. Pharmacogenetics of aromatase inhibitors in endocrine responsive breast cancer: lessons learnt from tamoxifen and CYP2D6 genotyping. Anticancer Agents Med Chem. 2017;17(13):1805–1813. doi:10.2174/1871521409666170412124226
74. Baatjes K, Peeters A, McCaul M, et al. CYP19A1 rs10046 pharmacogenetics in postmenopausal breast cancer patients treated with aromatase inhibitors: one-year follow-up. Curr Pharm Des. 2020;26(46):6007–6012. doi:10.2174/1381612826666200908141858
75. Glubb DM, O’Mara TA, Shamsani J, Spurdle AB. The association of CYP19A1 variation with circulating estradiol and aromatase inhibitor outcome: can CYP19A1 variants be used to predict treatment efficacy? Front Pharmacol. 2017;8:218. doi:10.3389/fphar.2017.00218
76. Hertz DL, Henry NL, Rae JM. Germline genetic predictors of aromatase inhibitor concentrations, estrogen suppression and drug efficacy and toxicity in breast cancer patients. Pharmacogenomics. 2017;18(5):481–499. doi:10.2217/pgs-2016-0205
77. Hertz DL, Speth KA, Kidwell KM, et al. Variable aromatase inhibitor plasma concentrations do not correlate with circulating estrogen concentrations in post-menopausal breast cancer patients. Breast Cancer Res Treat. 2017;165(3):659–668. doi:10.1007/s10549-017-4346-x
78. Kamdem LK, Liu Y, Stearns V, et al. In vitro and in vivo oxidative metabolism and glucuronidation of anastrozole. Br J Clin Pharmacol. 2010;70(6):854–869. doi:10.1111/j.1365-2125.2010.03791.x
79. Dudenkov TM, Liu D, Cairns J, et al. Anastrozole aromatase inhibitor plasma drug concentration genome-wide association study: functional epistatic interaction between SLC38A7 and ALPPL2. Clin Pharmacol Ther. 2019;106(1):219–227. doi:10.1002/cpt.1359
80. Edavana VK, Dhakal IB, Williams S, et al. Potential role of UGT1A4 promoter SNPs in anastrozole pharmacogenomics. Drug Metab Dispos. 2013;41(4):870–877. doi:10.1124/dmd.112.048157
81. Seneviratne HK, Tillotson J, Lade JM, et al. Metabolism of long-acting rilpivirine after intramuscular injection: HIV prevention trials network study 076 (HPTN 076). AIDS Res Hum Retroviruses. 2021;37(3):173–183. doi:10.1089/AID.2020.0155
82. Murai K, Yamazaki H, Nakagawa K, Kawai R, Kamataki T. Deactivation of anti-cancer drug letrozole to a carbinol metabolite by polymorphic cytochrome P450 2A6 in human liver microsomes. Xenobiotica. 2009;39(11):795–802. doi:10.3109/00498250903171395
83. Desta Z, Kreutz Y, Nguyen AT, et al. Plasma letrozole concentrations in postmenopausal women with breast cancer are associated with CYP2A6 genetic variants, body mass index, and age. Clin Pharmacol Ther. 2011;90(5):693–700. doi:10.1038/clpt.2011.174
84. Borrie AE, Rose RV, Choi YH, et al. Letrozole concentration is associated with CYP2A6 variation but not with arthralgia in patients with breast cancer. Breast Cancer Res Treat. 2018;172(2):371–379. doi:10.1007/s10549-018-4910-z
85. Hertz DL, Douglas JA, Kidwell KM, et al. Genome-wide association study of letrozole plasma concentrations identifies non-exonic variants that may affect CYP2A6 metabolic activity. Pharmacogenet Genomics. 2021;31(5):116–123. doi:10.1097/FPC.0000000000000429
86. Hertz DL, Kidwell KM, Seewald NJ, et al. Polymorphisms in drug-metabolizing enzymes and steady-state exemestane concentration in postmenopausal patients with breast cancer. Pharmacogenomics J. 2017;17(6):521–527. doi:10.1038/tpj.2016.60
87. de Jonge H, Elens L, de Loor H, van Schaik RH, Kuypers DR. The CYP3A4*22 C>T single nucleotide polymorphism is associated with reduced midazolam and tacrolimus clearance in stable renal allograft recipients. Pharmacogenomics J. 2015;15(2):144–152. doi:10.1038/tpj.2014.49
88. Henry NL, Skaar TC, Dantzer J, et al. Genetic associations with toxicity-related discontinuation of aromatase inhibitor therapy for breast cancer. Breast Cancer Res Treat. 2013;138(3):807–816. doi:10.1007/s10549-013-2504-3
89. Ingle JN, Schaid DJ, Goss PE, et al. Genome-wide associations and functional genomic studies of musculoskeletal adverse events in women receiving aromatase inhibitors. J Clin Oncol. 2010;28(31):4674–4682. doi:10.1200/JCO.2010.28.5064
90. Leyland-Jones B, Gray KP, Abramovitz M, et al. ESR1 and ESR2 polymorphisms in the BIG 1–98 trial comparing adjuvant letrozole versus tamoxifen or their sequence for early breast cancer. Breast Cancer Res Treat. 2015;154(3):543–555. doi:10.1007/s10549-015-3634-6
91. Hertz DL, Smith KL, Zong Y, et al. Further evidence that OPG rs2073618 is associated with increased risk of musculoskeletal symptoms in patients receiving aromatase inhibitors for early breast cancer. Front Genet. 2021;12:662734. doi:10.3389/fgene.2021.662734
92. Genome aggregation database (gnomAD v2.1.1). Available from: https://gnomad.broadinstitute.org/.
93. Ma CX, Adjei AA, Salavaggione OE, et al. Human aromatase: gene resequencing and functional genomics. Cancer Res. 2005;65(23):11071–11082. doi:10.1158/0008-5472.CAN-05-1218
94. Blackburn HL, Ellsworth DL, Shriver CD, Ellsworth RE. Role of cytochrome P450 genes in breast cancer etiology and treatment: effects on estrogen biosynthesis, metabolism, and response to endocrine therapy. Cancer Causes Control. 2015;26(3):319–332. doi:10.1007/s10552-014-0519-7
95. Colomer R, Monzo M, Tusquets I, et al. A single-nucleotide polymorphism in the aromatase gene is associated with the efficacy of the aromatase inhibitor letrozole in advanced breast carcinoma. Clin Cancer Res. 2008;14(3):811–816. doi:10.1158/1078-0432.CCR-07-1923
96. Liu L, Bai YX, Zhou JH, et al. A polymorphism at the 3’-UTR region of the aromatase gene is associated with the efficacy of the aromatase inhibitor, anastrozole, in metastatic breast carcinoma. Int J Mol Sci. 2013;14(9):18973–18988. doi:10.3390/ijms140918973
97. Garcia-Casado Z, Guerrero-Zotano A, Llombart-Cussac A, et al. A polymorphism at the 3’-UTR region of the aromatase gene defines a subgroup of postmenopausal breast cancer patients with poor response to neoadjuvant letrozole. BMC Cancer. 2010;10:36. doi:10.1186/1471-2407-10-36
98. Piccart-Gebhart MJ, Procter M, Leyland-Jones B, et al. Trastuzumab after adjuvant chemotherapy in HER2-positive breast cancer. N Engl J Med. 2005;353(16):1659–1672. doi:10.1056/NEJMoa052306
99. Romond EH, Perez EA, Bryant J, et al. Trastuzumab plus adjuvant chemotherapy for operable HER2-positive breast cancer. N Engl J Med. 2005;353(16):1673–1684. doi:10.1056/NEJMoa052122
100. Slamon D, Eiermann W, Robert N, et al. Adjuvant trastuzumab in HER2-positive breast cancer. N Engl J Med. 2011;365(14):1273–1283. doi:10.1056/NEJMoa0910383
101. Gershon N, Berchenko Y, Hall PS, Goldstein DA. Cost effectiveness and affordability of trastuzumab in sub-Saharan Africa for early stage HER2-positive breast cancer. Cost Eff Resour Alloc. 2019;17:5. doi:10.1186/s12962-019-0174-7
102. Cherif M, Amoako-Sakyi D, Dolo A, et al. Distribution of FcγR gene polymorphisms among two sympatric populations in Mali: differing allele frequencies, associations with malariometric indices and implications for genetic susceptibility to malaria. Malar J. 2016;15:29. doi:10.1186/s12936-015-1082-8
103. Mellor JD, Brown MP, Irving HR, Zalcberg JR, Dobrovic A. A critical review of the role of Fc gamma receptor polymorphisms in the response to monoclonal antibodies in cancer. J Hematol Oncol. 2013;6:1. doi:10.1186/1756-8722-6-1
104. Musolino A, Naldi N, Bortesi B, et al. Immunoglobulin G fragment C receptor polymorphisms and clinical efficacy of trastuzumab-based therapy in patients with HER-2/neu-positive metastatic breast cancer. J Clin Oncol. 2008;26(11):1789–1796. doi:10.1200/JCO.2007.14.8957
105. Hertz DL, McLeod HL, Hoskins JM. Pharmacogenetics of breast cancer therapies. Breast. 2009;18(Suppl 3):S59–S63. doi:10.1016/S0960-9776(09)70275-9
106. Gavin PG, Song N, Kim SR, et al. Association of polymorphisms in FCGR2A and FCGR3A with degree of trastuzumab benefit in the adjuvant treatment of ERBB2/HER2-positive breast cancer: analysis of the NSABP B-31 trial. JAMA Oncol. 2017;3(3):335–341. doi:10.1001/jamaoncol.2016.4884
107. Lassaunière R, Tiemessen CT. Variability at the FCGR locus: characterization in Black South Africans and evidence for ethnic variation in and out of Africa. Genes Immun. 2016;17(2):93–104. doi:10.1038/gene.2015.60
108. Beauclair S, Formento P, Fischel JL, et al. Role of the HER2 [Ile655Val] genetic polymorphism in tumorogenesis and in the risk of trastuzumab-related cardiotoxicity. Ann Oncol. 2007;18(8):1335–1341. doi:10.1093/annonc/mdm181
109. Stanton SE, Ward MM, Christos P, et al. Pro1170 Ala polymorphism in HER2-neu is associated with risk of trastuzumab cardiotoxicity. BMC Cancer. 2015;15:267. doi:10.1186/s12885-015-1298-6
110. Peddi PF, Fasching PA, Liu D, et al. Genetic polymorphisms and correlation with treatment induced cardiotoxicity and prognosis in breast cancer patients [published online ahead of print, 2022 Feb 2]. Clin Cancer Res. 2022;28:1854–1862. doi:10.1158/1078-0432.CCR-21-1762
111. Longley DB, Harkin DP, Johnston PG. 5-fluorouracil: mechanisms of action and clinical strategies. Nat Rev Cancer. 2003;3(5):330–338. doi:10.1038/nrc1074
112. Lévy E, Piedbois P, Buyse M; Meta-Analysis Group in Cancer. Toxicity of fluorouracil in patients with advanced colorectal cancer: effect of administration schedule and prognostic factors. J Clin Oncol. 1998;16(11):3537–3541. doi:10.1200/JCO.1998.16.11.3537
113. Schwab M, Zanger UM, Marx C, et al. Role of genetic and nongenetic factors for fluorouracil treatment-related severe toxicity: a prospective clinical trial by the German 5-FU Toxicity Study Group. J Clin Oncol. 2008;26(13):2131–2138. doi:10.1200/JCO.2006.10.4182
114. Arowolo OA, Njiaju UO, Ogundiran TO, et al. Neo-adjuvant capecitabine chemotherapy in women with newly diagnosed locally advanced breast cancer in a resource-poor setting (Nigeria): efficacy and safety in a Phase II feasibility study. Breast J. 2013;19(5):470–477. doi:10.1111/tbj.12149
115. Herbst CL, Miot JK, Moch SL, Ruff P. Colorectal Cancer (CRC) treatment and associated costs in the public sector compared to the private sector in Johannesburg, South Africa. BMC Health Serv Res. 2020;20(1):290. doi:10.1186/s12913-020-05112-w
116. Lee A, Ezzeldin H, Fourie J, Diasio R. Dihydropyrimidine dehydrogenase deficiency: impact of pharmacogenetics on 5-fluorouracil therapy. Clin Adv Hematol Oncol. 2004;2(8):527–532.
117. Saif MW, Syrigos K, Mehra R, Mattison LK, Diasio RB. Dihydropyrimidine dehydrogenase deficiency (DPD) in GI malignancies: experience of 4-years. Pak J Med Sci. 2007;23(6):832–839.
118. Rai K, Batukbhai BDO, Brooks GA. Risk of treatment-related death in carriers of pathogenic DPYD polymorphisms treated with fluoropyrimidine chemotherapy: a systematic review and patient-level analysis. J Clin Oncol. 2019;37(15 Suppl):e15132–e15132. doi:10.1200/JCO.2019.37.15suppl.e15132
119. Deenen MJ, Meulendijks D, Cats A, et al. Upfront genotyping of DPYD*2A to individualize fluoropyrimidine therapy: a safety and cost analysis. J Clin Oncol. 2016;34(3):227–234. doi:10.1200/JCO.2015.63.1325
120. Henricks LM, Lunenburg CA, de Man FM, et al. DPYD genotype-guided dose individualisation of fluoropyrimidine therapy in patients with cancer: a prospective safety analysis. Lancet Oncol. 2018;19(11):1459–1467. doi:10.1016/S1470-2045(18)30686-7
121. Amstutz U, Froehlich TK, Largiadèr CR. Dihydropyrimidine dehydrogenase gene as a major predictor of severe 5-fluorouracil toxicity. Pharmacogenomics. 2011;12(9):1321–1336. doi:10.2217/pgs.11.72
122. Ezzeldin HH, Lee AM, Mattison LK, Diasio RB. Methylation of the DPYD promoter: an alternative mechanism for dihydropyrimidine dehydrogenase deficiency in cancer patients. Clin Cancer Res. 2005;11(24 Pt 1):8699–8705. doi:10.1158/1078-0432.CCR-05-1520
123. Ramos E, Doumatey A, Elkahloun AG, et al. Pharmacogenomics, ancestry and clinical decision making for global populations. Pharmacogenomics J. 2014;14(3):217–222. doi:10.1038/tpj.2013.24
124. Dean L, Kane M. Fluorouracil therapy and DPYD genotype. In: Pratt VM, Scott SA, Pirmohamed M, editors. Medical Genetics Summaries. Bethesda: National Center for Biotechnology Information (US); 2016:1–17.
125. Offer SM, Lee AM, Mattison LK, Fossum C, Wegner NJ, Diasio RB. A DPYD variant (Y186C) in individuals of African ancestry is associated with reduced DPD enzyme activity. Clin Pharmacol Ther. 2013;94(1):158–166. doi:10.1038/clpt.2013.69
126. Saif MW, Lee AM, Offer SM, McConnell K, Relias V, Diasio RB. A DPYD variant (Y186C) specific to individuals of African descent in a patient with life-threatening 5-FU toxic effects: potential for an individualized medicine approach. Mayo Clin Proc. 2014;89(1):131–136. doi:10.1016/j.mayocp.2013.09.008
127. Zaanan A, Dumont LM, Loriot MA, Taieb J, Narjoz C. A case of 5-FU-related severe toxicity associated with the p.Y186C DPYD variant. Clin Pharmacol Ther. 2014;95(2):136. doi:10.1038/clpt.2013.183
128. Offer SM, Fossum CC, Wegner NJ, Stuflesser AJ, Butterfield GL, Diasio RB. Comparative functional analysis of DPYD variants of potential clinical relevance to dihydropyrimidine dehydrogenase activity. Cancer Res. 2014;74(9):2545–2554. doi:10.1158/0008-5472.CAN-13-2482
129. Offer SM, Diasio RB. Response to “A case of 5-FU-related severe toxicity associated with the P.Y186C DPYD variant”. Clin Pharmacol Ther. 2014;95(2):137. doi:10.1038/clpt.2013.207
130. da Rocha JEB, Lombard Z, Ramsay M. Potential impact of DPYD variation on fluoropyrimidine drug response in sub-saharan African populations. Front Genet. 2021;12:626954. doi:10.3389/fgene.2021.626954
131. Cresteil T, Monsarrat B, Dubois J, Sonnier M, Alvinerie P, Gueritte F. Regioselective metabolism of taxoids by human CYP3A4 and 2C8: structure-activity relationship. Drug Metab Dispos. 2002;30(4):438–445. doi:10.1124/dmd.30.4.438
132. O’Shaughnessy J, Gradishar WJ, Bhar P, Iglesias J. Nab-paclitaxel for first-line treatment of patients with metastatic breast cancer and poor prognostic factors: a retrospective analysis. Breast Cancer Res Treat. 2013;138(3):829–837. doi:10.1007/s10549-013-2447-8
133. Gonzalez-Angulo AM, Morales-Vasquez F, Hortobagyi GN. Overview of resistance to systemic therapy in patients with breast cancer. Adv Exp Med Biol. 2007;608:1–22. doi:10.1007/978-0-387-74039-3_1
134. Hertz DL. Exploring pharmacogenetics of paclitaxel- and docetaxel-induced peripheral neuropathy by evaluating the direct pharmacogenetic-pharmacokinetic and pharmacokinetic-neuropathy relationships. Expert Opin Drug Metab Toxicol. 2021;17(2):227–239. doi:10.1080/17425255.2021.1856367
135. Bergmann TK, Gréen H, Brasch-Andersen C, et al. Retrospective study of the impact of pharmacogenetic variants on paclitaxel toxicity and survival in patients with ovarian cancer. Eur J Clin Pharmacol. 2011;67(7):693–700. doi:10.1007/s00228-011-1007-6
136. Bergmann TK, Brasch-Andersen C, Gréen H, et al. Impact of CYP2C8*3 on paclitaxel clearance: a population pharmacokinetic and pharmacogenomic study in 93 patients with ovarian cancer. Pharmacogenomics J. 2011;11(2):113–120. doi:10.1038/tpj.2010.19
137. Marcath LA, Pasternak AL, Hertz DL. Challenges to assess substrate-dependent allelic effects in CYP450 enzymes and the potential clinical implications. Pharmacogenomics J. 2019;19(6):501–515. doi:10.1038/s41397-019-0105-1
138. Cavaco I, Strömberg-Nörklit J, Kaneko A, et al. CYP2C8 polymorphism frequencies among malaria patients in Zanzibar. Eur J Clin Pharmacol. 2005;61(1):15–18. doi:10.1007/s00228-004-0871-8
139. Röwer S, Bienzle U, Weise A, et al. Short communication: high prevalence of the cytochrome P450 2C8*2 mutation in Northern Ghana. Trop Med Int Health. 2005;10(12):1271–1273. doi:10.1111/j.1365-3156.2005.01525.x
140. Paganotti GM, Gallo BC, Verra F, et al. Human genetic variation is associated with Plasmodium falciparum drug resistance. J Infect Dis. 2011;204(11):1772–1778. doi:10.1093/infdis/jir629
141. Paganotti GM, Gramolelli S, Tabacchi F, et al. Distribution of human CYP2C8*2 allele in three different African populations. Malar J. 2012;11:125. doi:10.1186/1475-2875-11-125
142. Peko SM, Ntoumi F, Vouvoungui C, et al. Distribution of the cytochrome P450 CYP2C8*2 allele in Brazzaville, Republic of Congo. Int J Infect Dis. 2019;85:49–53. doi:10.1016/j.ijid.2019.04.035
143. Shittu O, Opeyemi OA, Omotesho OB, et al. Clinical probe of Cyp2C8*2 mutants in a malaria hyperendemic zone: evidence from North-Central, Nigeria. Acta Medica. 2020;63(3):119–123. doi:10.14712/18059694.2020.29
144. Dai D, Zeldin DC, Blaisdell JA, et al. Polymorphisms in human CYP2C8 decrease metabolism of the anticancer drug paclitaxel and arachidonic acid. Pharmacogenetics. 2001;11(7):597–607. doi:10.1097/00008571-200110000-00006
145. Gao Y, Liu D, Wang H, Zhu J, Chen C. Functional characterization of five CYP2C8 variants and prediction of CYP2C8 genotype-dependent effects on in vitro and in vivo drug-drug interactions. Xenobiotica. 2010;40(7):467–475. doi:10.3109/00498254.2010.487163
146. Yu L, Shi D, Ma L, Zhou Q, Zeng S. Influence of CYP2C8 polymorphisms on the hydroxylation metabolism of paclitaxel, repaglinide and ibuprofen enantiomers in vitro. Biopharm Drug Dispos. 2013;34(5):278–287. doi:10.1002/bdd.1842
147. Hertz DL, Roy S, Jack J, et al. Genetic heterogeneity beyond CYP2C8*3 does not explain differential sensitivity to paclitaxel-induced neuropathy. Breast Cancer Res Treat. 2014;145(1):245–254. doi:10.1007/s10549-014-2910-1
148. de Graan AJ, Elens L, Sprowl JA, et al. CYP3A4*22 genotype and systemic exposure affect paclitaxel-induced neurotoxicity. Clin Cancer Res. 2013;19(12):3316–3324. doi:10.1158/1078-0432.CCR-12-3786
149. Apellániz-Ruiz M, Lee MY, Sánchez-Barroso L, et al. Whole-exome sequencing reveals defective CYP3A4 variants predictive of paclitaxel dose-limiting neuropathy. Clin Cancer Res. 2015;21(2):322–328. doi:10.1158/1078-0432.CCR-14-1758
150. Cliff J, Jorgensen AL, Lord R, et al. The molecular genetics of chemotherapy-induced peripheral neuropathy: a systematic review and meta-analysis. Crit Rev Oncol Hematol. 2017;120:127–140. doi:10.1016/j.critrevonc.2017.09.009
151. Staff NP, Fehrenbacher JC, Caillaud M, Damaj MI, Segal RA, Rieger S. Pathogenesis of paclitaxel-induced peripheral neuropathy: a current review of in vitro and in vivo findings using rodent and human model systems. Exp Neurol. 2020;324:113121. doi:10.1016/j.expneurol.2019.113121
152. Argyriou AA, Bruna J, Genazzani AA, Cavaletti G. Chemotherapy-induced peripheral neurotoxicity: management informed by pharmacogenetics. Nat Rev Neurol. 2017;13(8):492–504. doi:10.1038/nrneurol.2017.88
153. Hertz DL, Owzar K, Lessans S, et al. Pharmacogenetic discovery in CALGB (Alliance) 90401 and mechanistic validation of a VAC14 polymorphism that increases risk of docetaxel-induced neuropathy. Clin Cancer Res. 2016;22(19):4890–4900. doi:10.1158/1078-0432.CCR-15-2823
154. Baker SD, Verweij J, Cusatis GA, et al. Pharmacogenetic pathway analysis of docetaxel elimination. Clin Pharmacol Ther. 2009;85(2):155–163. doi:10.1038/clpt.2008.95
155. de Graan AJ, Lancaster CS, Obaidat A, et al. Influence of polymorphic OATP1B-type carriers on the disposition of docetaxel. Clin Cancer Res. 2012;18(16):4433–4440. doi:10.1158/1078-0432.CCR-12-0761
156. Priyadarshini R, Raj GM, Sundaram R, Kayal S, Ramesh A, Shewade DG. Association of plasma docetaxel levels with ABCB1 gene polymorphisms and tumour response in locally advanced breast cancer patients of South India on neo-adjuvant chemotherapy. Breast Cancer. 2021;28(2):414–423. doi:10.1007/s12282-020-01177-x
157. Kim HJ, Im SA, Keam B, et al. ABCB1 polymorphism as prognostic factor in breast cancer patients treated with docetaxel and doxorubicin neoadjuvant chemotherapy. Cancer Sci. 2015;106(1):86–93. doi:10.1111/cas.12560
158. Kus T, Aktas G, Kalender ME, et al. Polymorphism of CYP3A4 and ABCB1 genes increase the risk of neuropathy in breast cancer patients treated with paclitaxel and docetaxel. Onco Targets Ther. 2016;9:5073–5080. doi:10.2147/OTT.S106574
159. Núñez-Torres R, Martín M, García-Sáenz JÁ, et al. Association between ABCB1 genetic variants and persistent chemotherapy-induced alopecia in women with breast cancer. JAMA Dermatol. 2020;156(9):987–991. doi:10.1001/jamadermatol.2020.1867
160. Kiire CF, Gombe-Mbalawa C, Tsega E, et al. Multicentre study of the treatment of primary liver cancer in Africa with two anthracycline drugs. Cent Afr J Med. 1992;38(11):428–431.
161. Naidoo H. The Genetics of Anthracycline-Induced Cardiotoxicity in Cancer Patients [dissertation]. Cape Town: University of Cape Town; 2018.
162. Zhang WZ, Azibani F, Sliwa K. Subclinical anthracycline therapy-related cardiac Dysfunction: an ignored stage B heart failure in an African population. Cardiovasc J Afr. 2020;31(5):262–266. doi:10.5830/CVJA-2020-013
163. Okabe M, Unno M, Harigae H, et al. Characterization of the organic cation transporter SLC22A16: a doxorubicin importer. Biochem Biophys Res Commun. 2005;333(3):754–762. doi:10.1016/j.bbrc.2005.05.174
164. Lal S, Wong ZW, Jada SR, et al. Novel SLC22A16 polymorphisms and influence on doxorubicin pharmacokinetics in Asian breast cancer patients. Pharmacogenomics. 2007;8(6):567–575. doi:10.2217/14622416.8.6.567
165. Westbrook K, Stearns V. Pharmacogenomics of breast cancer therapy: an update. Pharmacol Ther. 2013;139(1):1–11. doi:10.1016/j.pharmthera.2013.03.001
166. Yao S, Maghsoudlou D, Ambrosone CB. Breast cancer pharmacogenetics in the era of personalized medicine. Curr Breast Cancer Rep. 2012;4:271–281. doi:10.1007/s12609-012-0088-2
167. Ambrosone CB, Barlow WE, Reynolds W, et al. Myeloperoxidase genotypes and enhanced efficacy of chemotherapy for early-stage breast cancer in SWOG-8897. J Clin Oncol. 2009;27(30):4973–4979. doi:10.1200/JCO.2009.21.8669
168. Choi JY, Barlow WE, Albain KS, et al. Nitric oxide synthase variants and disease-free survival among treated and untreated breast cancer patients in a Southwest Oncology Group clinical trial. Clin Cancer Res. 2009;15(16):5258–5266. doi:10.1158/1078-0432.CCR-09-0685
169. Cui L, Huang J, Zhan Y, et al. Association between the genetic polymorphisms of the pharmacokinetics of anthracycline drug and myelosuppression in a patient with breast cancer with anthracycline-based chemotherapy. Life Sci. 2021;276:119392. doi:10.1016/j.lfs.2021.119392
170. Aouida M, Poulin R, Ramotar D. The human carnitine transporter SLC22A16 mediates high affinity uptake of the anticancer polyamine analogue bleomycin-A5. J Biol Chem. 2010;285(9):6275–6284. doi:10.1074/jbc.M109.046151
171. Faraji A, Dehghan Manshadi HR, Mobaraki M, Zare M, Houshmand M. Association of ABCB1 and SLC22A16 gene polymorphisms with incidence of doxorubicin-induced febrile neutropenia: a survey of Iranian breast cancer patients. PLoS One. 2016;11(12):e0168519. doi:10.1371/journal.pone.0168519
172. Bray J, Sludden J, Griffin MJ, et al. Influence of pharmacogenetics on response and toxicity in breast cancer patients treated with doxorubicin and cyclophosphamide. Br J Cancer. 2010;102(6):1003–1009. doi:10.1038/sj.bjc.6605587
173. Kafka A, Sauer G, Jaeger C, et al. Polymorphism C3435T of the MDR-1 gene predicts response to preoperative chemotherapy in locally advanced breast cancer. Int J Oncol. 2003;22(5):1117–1121.
174. Swart M, Ren Y, Smith P, Dandara C. ABCB1 4036A>G and 1236C>T polymorphisms affect plasma efavirenz levels in South African HIV/AIDS patients. Front Genet. 2012;3:236. doi:10.3389/fgene.2012.00236
175. Helsby NA, Yong M, van Kan M, de Zoysa JR, Burns KE. The importance of both CYP2C19 and CYP2B6 germline variations in cyclophosphamide pharmacokinetics and clinical outcomes. Br J Clin Pharmacol. 2019;85(9):1925–1934. doi:10.1111/bcp.14031
176. Xie HJ, Yasar U, Lundgren S, et al. Role of polymorphic human CYP2B6 in cyclophosphamide bioactivation. Pharmacogenomics J. 2003;3(1):53–61. doi:10.1038/sj.tpj.6500157
177. Ekhart C, Doodeman VD, Rodenhuis S, Smits PH, Beijnen JH, Huitema AD. Influence of polymorphisms of drug metabolizing enzymes (CYP2B6, CYP2C9, CYP2C19, CYP3A4, CYP3A5, GSTA1, GSTP1, ALDH1A1 and ALDH3A1) on the pharmacokinetics of cyclophosphamide and 4-hydroxycyclophosphamide. Pharmacogenet Genomics. 2008;18(6):515–523. doi:10.1097/FPC.0b013e3282fc9766
178. Afsar NA, Ufer M, Haenisch S, et al. Relationship of drug metabolizing enzyme genotype to plasma levels as well as myelotoxicity of cyclophosphamide in breast cancer patients. Eur J Clin Pharmacol. 2012;68(4):389–395. doi:10.1007/s00228-011-1134-0
179. Ahmed JH, Makonnen E, Bisaso RK, et al. Population pharmacokinetic, pharmacogenetic, and pharmacodynamic analysis of cyclophosphamide in Ethiopian breast cancer patients. Front Pharmacol. 2020;11:406. doi:10.3389/fphar.2020.00406
180. Marcath LA, Kidwell KM, Robinson AC, et al. Patients carrying CYP2C8*3 have shorter systemic paclitaxel exposure. Pharmacogenomics. 2019;20(2):95–104. doi:10.2217/pgs-2018-0162
181. Pernaute-Lau L, Adegnika AA, Zhou Y, et al. Pharmacogene sequencing of a Gabonese population with severe plasmodium falciparum malaria reveals multiple novel variants with putative relevance for antimalarial treatment. Antimicrob Agents Chemother. 2021;65(7):e0027521. doi:10.1128/AAC.00275-21
182. Griese EU, Asante-Poku S, Ofori-Adjei D, Mikus G, Eichelbaum M. Analysis of the CYP2D6 gene mutations and their consequences for enzyme function in a West African population. Pharmacogenetics. 1999;9(6):715–723. doi:10.1097/00008571-199912000-00006
183. Fuselli S, de Filippo C, Mona S, et al. Evolution of detoxifying systems: the role of environment and population history in shaping genetic diversity at human CYP2D6 locus. Pharmacogenet Genomics. 2010;20(8):485–499. doi:10.1097/FPC.0b013e32833bba25
184. Sistonen J, Sajantila A, Lao O, Corander J, Barbujani G, Fuselli S. CYP2D6 worldwide genetic variation shows high frequency of altered activity variants and no continental structure. Pharmacogenet Genomics. 2007;17(2):93–101. doi:10.1097/01.fpc.0000239974.69464.f2
185. Aklillu E, Persson I, Bertilsson L, Johansson I, Rodrigues F, Ingelman-Sundberg M. Frequent distribution of ultrarapid metabolizers of debrisoquine in an Ethiopian population carrying duplicated and multiduplicated functional CYP2D6 alleles. J Pharmacol Exp Ther. 1996;278(1):441–446.
186. Dandara C, Masimirembwa CM, Magimba A, et al. Genetic polymorphism of CYP2D6 and CYP2C19 in east- and Southern African populations including psychiatric patients. Eur J Clin Pharmacol. 2001;57(1):11–17. doi:10.1007/s002280100282
187. Wennerholm A, Johansson I, Massele AY, et al. Decreased capacity for debrisoquine metabolism among black Tanzanians: analyses of the CYP2D6 genotype and phenotype. Pharmacogenetics. 1999;9(6):707–714. doi:10.1097/00008571-199912000-00005
188. Wennerholm A, Johansson I, Hidestrand M, Bertilsson L, Gustafsson LL, Ingelman-Sundberg M. Characterization of the CYP2D6*29 allele commonly present in a black Tanzanian population causing reduced catalytic activity [published correction appears in Pharmacogenetics 2001 Nov;11(8):743]. Pharmacogenetics. 2001;11(5):417–427. doi:10.1097/00008571-200107000-00005
189. Mehlotra RK, Gaedigk A, Howes RE, Rakotomanga TA, Ratsimbasoa AC, Zimmerman PA. CYP2D6 genetic variation and its implication for vivax malaria treatment in Madagascar. Front Pharmacol. 2021;12:654054. doi:10.3389/fphar.2021.654054
190. Wright GE, Niehaus DJ, Drögemöller BI, Koen L, Gaedigk A, Warnich L. Elucidation of CYP2D6 genetic diversity in a unique African population: implications for the future application of pharmacogenetics in the Xhosa population. Ann Hum Genet. 2010;74(4):340–350. doi:10.1111/j.1469-1809.2010.00585.x
191. Masimirembwa CM, Johansson I, Hasler JA, Ingelman-Sundberg M. Genetic polymorphism of cytochrome P450 CYP2D6 in Zimbabwean population. Pharmacogenetics. 1993;3(6):275–280. doi:10.1097/00008571-199312000-00001
192. Matimba A, Oluka MN, Ebeshi BU, et al. Establishment of a biobank and pharmacogenetics database of African populations. Eur J Hum Genet. 2008;16(7):780–783. doi:10.1038/ejhg.2008.49
193. Evans WE, Relling MV, Rahman A, McLeod HL, Scott EP, Lin JS. Genetic basis for a lower prevalence of deficient CYP2D6 oxidative drug metabolism phenotypes in black Americans. J Clin Invest. 1993;91(5):2150–2154. doi:10.1172/JCI116441
194. Leathart JB, London SJ, Steward A, Adams JD, Idle JR, Daly AK. CYP2D6 phenotype-genotype relationships in African-Americans and Caucasians in Los Angeles. Pharmacogenetics. 1998;8(6):529–541. doi:10.1097/00008571-199812000-00010
195. Wan YJ, Poland RE, Han G, et al. Analysis of the CYP2D6 gene polymorphism and enzyme activity in African-Americans in Southern California. Pharmacogenetics. 2001;11(6):489–499. doi:10.1097/00008571-200108000-00004
196. Gaedigk A, Bradford LD, Marcucci KA, Leeder JS. Unique CYP2D6 activity distribution and genotype-phenotype discordance in black Americans. Clin Pharmacol Ther. 2002;72(1):76–89. doi:10.1067/mcp.2002.125783
197. Cai WM, Nikoloff DM, Pan RM, et al. CYP2D6 genetic variation in healthy adults and psychiatric African-American subjects: implications for clinical practice and genetic testing. Pharmacogenomics J. 2006;6(5):343–350. doi:10.1038/sj.tpj.6500378
198. de Leon J, Susce MT, Johnson M, et al. DNA microarray technology in the clinical environment: the AmpliChip CYP450 test for CYP2D6 and CYP2C19 genotyping. CNS Spectr. 2009;14(1):19–34. doi:10.1017/s1092852900020022
199. Del Tredici AL, Malhotra A, Dedek M, et al. Frequency of CYP2D6 alleles including structural variants in the United States. Front Pharmacol. 2018;9:305. doi:10.3389/fphar.2018.00305
200. Allabi AC, Gala JL, Desager JP, Heusterspreute M, Horsmans Y. Genetic polymorphisms of CYP2C9 and CYP2C19 in the Beninese and Belgian populations. Br J Clin Pharmacol. 2003;56(6):653–657. doi:10.1046/j.1365-2125.2003.01937.x
201. Allabi AC, Gala JL, Horsmans Y. CYP2C9, CYP2C19, ABCB1 (MDR1) genetic polymorphisms and phenytoin metabolism in a Black Beninese population. Pharmacogenet Genomics. 2005;15(11):779–786. doi:10.1097/01.fpc.0000174787.92861.91
202. Janha RE, Sisay-Joof F, Hamid-Adiamoh M, et al. Effects of genetic variation at the CYP2C19/CYP2C9 locus on pharmacokinetics of chlorcycloguanil in adult Gambians. Pharmacogenomics. 2009;10(9):1423–1431. doi:10.2217/pgs.09.72
203. Kudzi W, Dodoo AN, Mills JJ. Characterisation of CYP2C8, CYP2C9 and CYP2C19 polymorphisms in a Ghanaian population. BMC Med Genet. 2009;10:124. doi:10.1186/1471-2350-10-124
204. Matimba A, Del-Favero J, Van Broeckhoven C, Masimirembwa C. Novel variants of major drug-metabolising enzyme genes in diverse African populations and their predicted functional effects. Hum Genomics. 2009;3(2):169–190. doi:10.1186/1479-7364-3-2-169
205. Scordo MG, Aklillu E, Yasar U, Dahl ML, Spina E, Ingelman-Sundberg M. Genetic polymorphism of cytochrome P450 2C9 in a Caucasian and a black African population. Br J Clin Pharmacol. 2001;52(4):447–450. doi:10.1046/j.0306-5251.2001.01460.x
206. Staehli Hodel EM, Csajka C, Ariey F, et al. Effect of single nucleotide polymorphisms in cytochrome P450 isoenzyme and N-acetyltransferase 2 genes on the metabolism of artemisinin-based combination therapies in malaria patients from Cambodia and Tanzania. Antimicrob Agents Chemother. 2013;57(2):950–958. doi:10.1128/AAC.01700-12
207. Oliveira E, Pereira R, Amorim A, McLeod H, Prata MJ. Patterns of pharmacogenetic diversity in African populations: role of ancient and recent history. Pharmacogenomics. 2009;10(9):1413–1422. doi:10.2217/pgs.09.74
208. Mitchell C, Gregersen N, Krause A. Novel CYP2C9 and VKORC1 gene variants associated with warfarin dosage variability in the South African black population. Pharmacogenomics. 2011;12(7):953–963. doi:10.2217/pgs.11.36
209. Yen-Revollo JL, Van Booven DJ, Peters EJ, et al. Influence of ethnicity on pharmacogenetic variation in the Ghanaian population. Pharmacogenomics J. 2009;9(6):373–379. doi:10.1038/tpj.2009.28
210. Li-Wan-Po A, Girard T, Farndon P, Cooley C, Lithgow J. Pharmacogenetics of CYP2C19: functional and clinical implications of a new variant CYP2C19*17. Br J Clin Pharmacol. 2010;69(3):222–230. doi:10.1111/j.1365-2125.2009.03578.x
211. Tayeb MT, Clark C, Ameyaw MM, et al. CYP3A4 promoter variant in Saudi, Ghanaian and Scottish Caucasian populations. Pharmacogenetics. 2000;10(8):753–756. doi:10.1097/00008571-200011000-00009
212. Zeigler-Johnson CM, Walker AH, Mancke B, et al. Ethnic differences in the frequency of prostate cancer susceptibility alleles at SRD5A2 and CYP3A4. Hum Hered. 2002;54(1):13–21. doi:10.1159/000066695
213. Garsa AA, McLeod HL, Marsh S. CYP3A4 and CYP3A5 genotyping by pyrosequencing. BMC Med Genet. 2005;6:19. doi:10.1186/1471-2350-6-19
214. Ferreira PE, Veiga MI, Cavaco I, et al. Polymorphism of antimalaria drug metabolizing, nuclear receptor, and drug transport genes among malaria patients in Zanzibar, East Africa. Ther Drug Monit. 2008;30(1):10–15. doi:10.1097/FTD.0b013e31815e93c6
215. Mutagonda RF, Kamuhabwa AAR, Minzi OMS, et al. Effect of pharmacogenetics on plasma lumefantrine pharmacokinetics and malaria treatment outcome in pregnant women. Malar J. 2017;16(1):267. doi:10.1186/s12936-017-1914-9
216. Penzak SR, Kabuye G, Mugyenyi P, et al. Cytochrome P450 2B6 (CYP2B6) G516T influences nevirapine plasma concentrations in HIV-infected patients in Uganda. HIV Med. 2007;8(2):86–91. doi:10.1111/j.1468-1293.2007.00432.x
217. Swart M, Skelton M, Wonkam A, Kannemyer L, Chin’ombe N, Dandara C. CYP1A2, CYP2A6, CYP2B6, CYP3A4 and CYP3A5 polymorphisms in two bantu-speaking populations from Cameroon and South Africa: implications of global pharmacogenetics. Curr Pharmacogenomics Person Med. 2012;10(1):43–53. doi:10.2174/1875692111201010043
218. Wojnowski L, Turner PC, Pedersen B, et al. Increased levels of aflatoxin-albumin adducts are associated with CYP3A5 polymorphisms in The Gambia, West Africa. Pharmacogenetics. 2004;14(10):691–700. doi:10.1097/00008571-200410000-00007
219. Kudzi W, Dodoo AN, Mills JJ. Genetic polymorphisms in MDR1, CYP3A4 and CYP3A5 genes in a Ghanaian population: a plausible explanation for altered metabolism of ivermectin in humans? BMC Med Genet. 2010;11:111. doi:10.1186/1471-2350
220. Kudzi W, Adjei GO, Ofori-Adjei D, Dodoo AN. Pharmacogenetics in Ghana: reviewing the evidence. Ghana Med J. 2011;45(2):73–80. doi:10.4314/gmj.v45i2.68932
221. Dandara C, Ballo R, Parker MI. CYP3A5 genotypes and risk of oesophageal cancer in two South African populations. Cancer Lett. 2005;225(2):275–282. doi:10.1016/j.canlet.2004.11.004
222. Roy JN, Lajoie J, Zijenah LS, et al. CYP3A5 genetic polymorphisms in different ethnic populations. Drug Metab Dispos. 2005;33(7):884–887. doi:10.1124/dmd.105.003822
223. Coughtrie MW, Gilissen RA, Shek B, et al. Phenol sulphotransferase SULT1A1 polymorphism: molecular diagnosis and allele frequencies in Caucasian and African populations. Biochem J. 1999;337(Pt 1):45–49. doi:10.1042/bj3370045
224. Mbongwa HP, Pretorius PJ, Kruger A, Koekemoer G, Reinecke CJ. Single nucleotide and copy number polymorphisms of the SULT1A1 gene in a South African Tswana population group. S Afr J Sci. 2011;107(11/12):1–6. doi:10.4102/sajs.v107i11/12.395
225. Carlini EJ, Raftogianis RB, Wood TC, et al. Sulfation pharmacogenetics: SULT1A1 and SULT1A2 allele frequencies in Caucasian, Chinese and African-American subjects. Pharmacogenetics. 2001;11(1):57–68. doi:10.1097/00008571-200102000-00007
226. Mehlotra RK, Bockarie MJ, Zimmerman PA. Prevalence of UGT1A9 and UGT2B7 nonsynonymous single nucleotide polymorphisms in West African, Papua New Guinean, and North American populations. Eur J Clin Pharmacol. 2007;63(1):1–8. doi:10.1007/s00228-006-0206-z
227. Gyamfi MA, Fujieda M, Kiyotani K, Yamazaki H, Kamataki T. High prevalence of cytochrome P450 2A6*1A alleles in a black African population of Ghana. Eur J Clin Pharmacol. 2005;60(12):855–857. doi:10.1007/s00228-004-0854-9
228. Adehin A, Bolaji OO, Maggo S, Kennedy MA. Relationship between metabolic phenotypes and genotypes of CYP1A2 and CYP2A6 in the Nigerian population. Drug Metab Pers Ther. 2017;32(1):39–47. doi:10.1515/dmpt-2016-0041
229. Decloedt EH, Sinxadi PZ, van Zyl GU, et al. Pharmacogenetics and pharmacokinetics of CNS penetration of efavirenz and its metabolites. J Antimicrob Chemother. 2019;74(3):699–709. doi:10.1093/jac/dky481
230. Tanner JA, Tyndale RF. Variation in CYP2A6 activity and personalized medicine. J Pers Med. 2017;7(4):18. doi:10.3390/jpm7040018
231. Napoli N, Rastelli A, Ma C, et al. Genetic polymorphism at Val80 (rs700518) of the CYP19A1 gene is associated with aromatase inhibitor associated bone loss in women with ER + breast cancer. Bone. 2013;55(2):309–314. doi:10.1016/j.bone.2013.04.021
232. Guo L, Liu Y, Liu L, et al. The CYP19A1 (TTTA)n repeat polymorphism may affect the prostate cancer risk: evidence from a meta-analysis. Am J Mens Health. 2021;15(3):15579883211017033. doi:10.1177/15579883211017033
233. Connolly S, Wall KM, Tang J, et al. Fc-gamma receptor IIA and IIIA variants in two African cohorts: lack of consistent impact on heterosexual HIV acquisition, viral control, and disease progression. Virology. 2018;525:132–142. doi:10.1016/j.virol.2018.09.015
234. Elraiyah T, Jerde CR, Shrestha S, et al. Novel deleterious dihydropyrimidine dehydrogenase variants may contribute to 5-fluorouracil sensitivity in an East African population. Clin Pharmacol Ther. 2017;101(3):382–390. doi:10.1002/cpt.531
235. Parikh S, Ouedraogo JB, Goldstein JA, Rosenthal PJ, Kroetz DL. Amodiaquine metabolism is impaired by common polymorphisms in CYP2C8: implications for malaria treatment in Africa. Clin Pharmacol Ther. 2007;82(2):197–203. doi:10.1038/sj.clpt.6100122
236. Motshoge T, Tawe L, Muthoga CW, et al. Cytochrome P450 2C8*2 allele in Botswana: human genetic diversity and public health implications. Acta Trop. 2016;157:54–58. doi:10.1016/j.actatropica.2016.01.028
237. Ngaimisi E, Habtewold A, Minzi O, et al. Importance of ethnicity, CYP2B6 and ABCB1 genotype for efavirenz pharmacokinetics and treatment outcomes: a parallel-group prospective cohort study in two sub-Saharan Africa populations. PLoS One. 2013;8(7):e67946. doi:10.1371/journal.pone.0067946
238. Kiaco K, Rodrigues AS, Do Rosário V, Gil JP, Lopes D. The drug transporter ABCB1 c.3435C>T SNP influences artemether-lumefantrine treatment outcome. Malar J. 2017;16(1):383. doi:10.1186/s12936-017-2006-6
239. Brown KC, Hosseinipour MC, Hoskins JM, et al. Exploration of CYP450 and drug transporter genotypes and correlations with nevirapine exposure in Malawians. Pharmacogenomics. 2012;13(1):113–121. doi:10.2217/pgs.11.132
240. Paganotti GM, Russo G, Sobze MS, et al. CYP2B6 poor metaboliser alleles involved in efavirenz and nevirapine metabolism: CYP2B6*9 and CYP2B6*18 distribution in HIV-exposed subjects from Dschang, Western Cameroon. Infect Genet Evol. 2015;35:122–126. doi:10.1016/j.meegid.2015.08.003
241. Nguefeu Nkenfou C, Atogho Tiedeu B, Nguefeu Nkenfou C, et al. Adverse drug reactions associated with CYP 2B6 polymorphisms in HIV/ AIDS-treated patients in Yaoundé, Cameroon. Appl Clin Genet. 2019;12:261–268. doi:10.2147/TACG.S226318
242. Klein K, Lang T, Saussele T, et al. Genetic variability of CYP2B6 in populations of African and Asian origin: allele frequencies, novel functional variants, and possible implications for anti-HIV therapy with efavirenz. Pharmacogenet Genomics. 2005;15(12):861–873. doi:10.1097/01213011-200512000-00004
243. Mehlotra RK, Ziats MN, Bockarie MJ, Zimmerman PA. Prevalence of CYP2B6 alleles in malaria-endemic populations of West Africa and Papua New Guinea. Eur J Clin Pharmacol. 2006;62(4):267–275. doi:10.1007/s00228-005-0092-972
244. Mehlotra RK, Bockarie MJ, Zimmerman PA. CYP2B6 983T>C polymorphism is prevalent in West Africa but absent in Papua New Guinea: implications for HIV/AIDS treatment. Br J Clin Pharmacol. 2007;64(3):391–395. doi:10.1111/j.1365-2125.2007.02884.x
245. Kwara A, Lartey M, Sagoe KW, Rzek NL, Court MH. CYP2B6 (c.516G–>T) and CYP2A6 (*9B and/or *17) polymorphisms are independent predictors of efavirenz plasma concentrations in HIV- infected patients. Br J Clin Pharmacol. 2009;67(4):427–436. doi:10.1111/j.1365-2125.2009.03368.x
246. Kwara A, Lartey M, Sagoe KW, Kenu E, Court MH. CYP2B6, CYP2A6 and UGT2B7 genetic polymorphisms are predictors of efavirenz mid-dose concentration in HIV-infected patients. AIDS. 2009;23(16):2101–2106. doi:10.1097/QAD.0b013e3283319908
247. Sarfo FS, Zhang Y, Egan D, et al. Pharmacogenetic associations with plasma efavirenz concentrations and clinical correlates in a retrospective cohort of Ghanaian HIV-infected patients. J Antimicrob Chemother. 2014;69(2):491–499. doi:10.1093/jac/dkt372
248. Sa’ad Toyin A, Julius Olugbenga S, Rahman Ayodele B, Olusola Joseph O, Taibat Moji BO. Allele and genotype frequencies of cytochrome P450 2B6 516G>T single nucleotide polymorphism in HIV- negative and HIV-infected adult Nigerian Populations. Am J Ther. 2016;23(6):e1715–e1719. doi:10.1097/MJT.0000000000000340
249. Olagunju A, Schipani A, Bolaji O, Khoo S, Owen A. Evaluation of universal versus genotype-guided efavirenz dose reduction in pregnant women using population pharmacokinetic modelling. J Antimicrob Chemother. 2018;73(1):165–172. doi:10.1093/jac/dkx334
250. Peko SM, Gueye NSG, Vouvoungui C, et al. Cytochrome P450 CYP2B6*6 distribution among Congolese individuals with HIV, Tuberculosis and Malaria infection. Int J Infect Dis. 2019;82:111–116. doi:10.1016/j.ijid.2019.02.025
251. Calcagno A, D’Avolio A, Simiele M, et al. Influence of CYP2B6 and ABCB1 SNPs on nevirapine plasma concentrations in burundese HIV-positive patients using dried sample spot devices. Br J Clin Pharmacol. 2012;74(1):134–140. doi:10.1111/j.1365-2125.2012.04163.x
252. Habtewold A, Amogne W, Makonnen E, et al. Long-term effect of efavirenz autoinduction on plasma/peripheral blood mononuclear cell drug exposure and CD4 count is influenced by UGT2B7 and CYP2B6 genotypes among HIV patients. J Antimicrob Chemother. 2011;66(10):2350–2361. doi:10.1093/jac/dkr304
253. Yimer G, Amogne W, Habtewold A, et al. High plasma efavirenz level and CYP2B6*6 are associated with efavirenz-based HAART- induced liver injury in the treatment of naïve HIV patients from Ethiopia: a prospective cohort study. Pharmacogenomics J. 2012;12(6):499–506. doi:10.1038/tpj.2011.34
254. Habtewold A, Aklillu E, Makonnen E, et al. Population pharmacokinetic model linking plasma and peripheral blood mononuclear cell concentrations of efavirenz and its metabolite, 8-hydroxy-efavirenz, in HIV patients. Antimicrob Agents Chemother. 2017;61(8):e00207– e00217. doi:10.1128/AAC.00207-17
255. Oluka MN, Okalebo FA, Guantai AN, McClelland RS, Graham SM. Cytochrome P450 2B6 genetic variants are associated with plasma nevirapine levels and clinical response in HIV-1 infected Kenyan women: a prospective cohort study. AIDS Res Ther. 2015;12:10. doi:10.1186/s12981-015-0052-0
256. Mutwa PR, Fillekes Q, Malgaz M, et al. Mid-dosing interval efavirenz plasma concentrations in HIV-1-infected children in Rwanda: treatment efficacy, tolerability, adherence, and the influence of CYP2B6 polymorphisms. J Acquir Immune Defic Syndr. 2012;60(4):400–404. doi:10.1097/QAI.0b013e3182569f57
257. Bienvenu E, Swart M, Dandara C, Ashton M. The role of genetic polymorphisms in cytochrome P450 and effects of tuberculosis co-treatment on the predictive value of CYP2B6 SNPs and on efavirenz plasma levels in adult HIV patients. Antiviral Res. 2014;102:44–53. doi:10.1016/j.antiviral.2013.11.011
258. Radloff R, Gras A, Zanger UM, et al. Novel CYP2B6 enzyme variants in a Rwandese population: functional characterization and assessment of in silico prediction tools. Hum Mutat. 2013;34(5):725–734. doi:10.1002/humu.22295
259. Marwa KJ, Schmidt T, Sjögren M, Minzi OM, Kamugisha E, Swedberg G. Cytochrome P450 single nucleotide polymorphisms in an indigenous Tanzanian population: a concern about the metabolism of artemisinin-based combinations. Malar J. 2014;13:420. doi:10.1186/1475-2875-13-420
260. Maganda BA, Minzi OM, Ngaimisi E, Kamuhabwa AA, Aklillu E. CYP2B6*6 genotype and high efavirenz plasma concentration but not nevirapine are associated with low lumefantrine plasma exposure and poor treatment response in HIV-malaria-coinfected patients. Pharmacogenomics J. 2016;16(1):88–95. doi:10.1038/tpj.2015.37
261. Kitabi EN, Minzi OMS, Mugusi S, et al. Long-term efavirenz pharmacokinetics is comparable between Tanzanian HIV and HIV/tuberculosis patients with the same CYP2B6*6 genotype. Sci Rep. 2018;8(1):16316. doi:10.1038/s41598-018-34674-3
262. Mukonzo JK, Röshammar D, Waako P, et al. A novel polymorphism in ABCB1 gene, CYP2B6*6 and sex predict single-dose efavirenz population pharmacokinetics in Ugandans. Br J Clin Pharmacol. 2009;68(5):690–699. doi:10.1111/j.1365-2125.2009.03516.x
263. Jamshidi Y, Moreton M, McKeown DA, et al. Tribal ethnicity and CYP2B6 genetics in Ugandan and Zimbabwean populations in the UK: implications for efavirenz dosing in HIV infection. J Antimicrob Chemother. 2010;65(12):2614–2619. doi:10.1093/jac/dkq369
264. von Braun A, Castelnuovo B, Ledergerber B, et al. High efavirenz serum concentrations in TB/HIV-coinfected Ugandan adults with a CYP2B6 516 TT genotype on anti-TB treatment. J Antimicrob Chemother. 2019;74(1):135–138. doi:10.1093/jac/dky379
265. Gross R, Aplenc R, Tenhave T, et al. Slow efavirenz metabolism genotype is common in Botswana. J Acquir Immune Defic Syndr. 2008;49(3):336–337. doi:10.1097/QAI.0b013e31817c1ed0
266. Vujkovic M, Bellamy SL, Zuppa AF, et al. Brief report: CYP2B6 516G>T minor allele protective of late virologic failure in efavirenz- treated HIV-infected patients in Botswana. J Acquir Immune Defic Syndr. 2017;75(4):488–491. doi:10.1097/QAI.0000000000001442
267. Vujkovic M, Bellamy SL, Zuppa AF, et al. Polymorphisms in cytochrome P450 are associated with extensive efavirenz pharmacokinetics and CNS toxicities in an HIV cohort in Botswana. Pharmacogenomics J. 2018;18(5):678–688. doi:10.1038/s41397-018-0028-2
268. Dickinson L, Chaponda M, Carr DF, et al. Population pharmacokinetic and pharmacogenetic analysis of nevirapine in hypersensitive and tolerant HIV-infected patients from Malawi. Antimicrob Agents Chemother. 2014;58(2):706–712. doi:10.1128/AAC.02069-13
269. Ciccacci C, Di Fusco D, Marazzi MC, et al. Association between CYP2B6 polymorphisms and nevirapine-induced SJS/TEN: a pharmacogenetics study. Eur J Clin Pharmacol. 2013;69(11):1909–1916. doi:10.1007/s00228-013-1549-x
270. Arnaldo P, Thompson RE, Lopes MQ, Suffys PN, Santos AR. Frequencies of cytochrome P450 2B6 and 2C8 allelic variants in the Mozambican Population. Malays J Med Sci. 2013;20(4):13–23.
271. Cohen K, Grant A, Dandara C, et al. Effect of rifampicin-based antitubercular therapy and the cytochrome P450 2B6 516G>T polymorphism on efavirenz concentrations in adults in South Africa. Antivir Ther. 2009;14(5):687–695. doi:10.1177/135965350901400502
272. Gounden V, van Niekerk C, Snyman T, George JA. Presence of the CYP2B6 516G> T polymorphism, increased plasma efavirenz concentrations and early neuropsychiatric side effects in South African HIV-infected patients. AIDS Res Ther. 2010;7:32. doi:10.1186/1742-6405-7-32
273. Swart M, Skelton M, Ren Y, Smith P, Takuva S, Dandara C. High predictive value of CYP2B6 SNPs for steady-state plasma efavirenz levels in South African HIV/AIDS patients. Pharmacogenet Genomics. 2013;23(8):415–427. doi:10.1097/FPC.0b013e328363176f
274. Sinxadi PZ, Leger PD, McIlleron HM, et al. Pharmacogenetics of plasma efavirenz exposure in HIV-infected adults and children in South Africa. Br J Clin Pharmacol. 2015;80(1):146–156. doi:10.1111/bcp.12590
275. Röhrich CR, Drögemöller BI, Ikediobi O, et al. CYP2B6*6 and CYP2B6*18 predict long-term efavirenz exposure measured in hair samples in HIV-positive South African women. AIDS Res Hum Retroviruses. 2016;32(6):529–538. doi:10.1089/AID.2015.0048
276. Reay R, Dandara C, Viljoen M, Rheeders M. CYP2B6 haplotype predicts efavirenz plasma concentration in Black South African HIV-1-infected children: a Longitudinal Pediatric Pharmacogenomic Study. OMICS. 2017;21(8):465–473. doi:10.1089/omi.2017.0078
277. Nyakutira C, Röshammar D, Chigutsa E, et al. High prevalence of the CYP2B6 516G–>T(*6) variant and effect on the population pharmacokinetics of efavirenz in HIV/ AIDS outpatients in Zimbabwe. Eur J Clin Pharmacol. 2008;64(4):357–365. doi:10.1007/s00228-007-0412-3
278. Maimbo M, Kiyotani K, Mushiroda T, Masimirembwa C, Nakamura Y. CYP2B6 genotype is a strong predictor of systemic exposure to efavirenz in HIV-infected Zimbabweans. Eur J Clin Pharmacol. 2012;68(3):267–271. doi:10.1007/s00228-011-1118-0
279. Dhoro M, Zvada S, Ngara B, et al. CYP2B6*6, CYP2B6*18, body weight and sex are predictors of efavirenz pharmacokinetics and treatment response: population pharmacokinetic modeling in an HIV/ AIDS and TB cohort in Zimbabwe. BMC Pharmacol Toxicol. 2015;16(4). doi:10.1186/s40360-015-0004-2
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.