")
Back to Journals » Cancer Management and Research » Volume 12
Oxymatrine Inhibits Colorectal Cancer Metastasis via Attenuating PKM2-Mediated Aerobic Glycolysis
Authors Li X, Sun J, Xu Q, Duan W, Yang L, Wu X, Lu G, Zhang L, Zheng Y
Received 15 June 2020
Accepted for publication 28 August 2020
Published 1 October 2020 Volume 2020:12 Pages 9503—9513
DOI https://doi.org/10.2147/CMAR.S267686
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
Xiaoping Li,1 Jie Sun,2 Qinghua Xu,1 Weiping Duan,2 Licheng Yang,1 Xing Wu,1 Guang Lu,1 Li Zhang,1 Yunfeng Zheng2
1Department of General Surgery, Liyang People′s Hospital, Jiangsu 213300, People’s Republic of China; 2School of Pharmacy, Nanjing University of Chinese Medicine, Nanjing 210023, People’s Republic of China
Correspondence: Xing Wu
Department of General Surgery, Liyang People′s Hospital, No. 70, JianShe West Road, Liyang City, Jiangsu Province, People’s Republic of China
Tel +86-519-6809-1103
Email [email protected]
Yunfeng Zheng
School of Pharmacy, Nanjing University of Chinese Medicine, No. 138, Xianlin Ave, Nanjing, Jiangsu Province, People’s Republic of China
Tel +86-25-8679-8186
Email [email protected]
Background: Colorectal cancer (CRC), a type of highly occurred intestinal cancer at present, is prone to metastasis at the later stage of chemotherapy. Looking for the anti-metastatic agents from natural compounds attracted much concern. Here, it aims to demonstrate whether oxymatrine, an anti-cancer natural compound, has anti-metastatic activity and its potential significance in clinic.
Materials and Methods: Wound healing assay and transwell assay were for evaluating the effect of oxymatrine on cell migration and invasion in vitro. Anti-metastatic action in vivo was determined by hepatic metastasis of colorectal cancer cells in mice.
Results: Oxymatrine can significantly inhibit cancer cell migration and invasion in vitro. The production of ATP, pyruvate, and lactate was suppressed in CRC cells under the treatment of oxymatrine, as well as the glucose consumption. Meantime, extracellular acidification rates (ECR) were evidently attenuated although the oxygen consumption rates (OCR) were not affected. Both clued that oxymatrine inhibition of metastasis is possibly related to blocking aerobic glycolysis. Subsequent results indicated that pyruvate kinase M2 (PKM2) not hexokinase (HK) and phosphofructokinase (PFK) were involved in oxymatrine blocking glycolysis as the PKM2 kinase activity and expression were inhibited by oxymatrine and the PKM2 activator, TEPP-46, can reverse in part the effect of oxymatrine induced in CRC cells. Furthermore, this process was also mediated by inhibition of glucose transporter 1 (GLUT1). Finally, the in vivo metastatic model in mice showed both 20 mg/kg and 40 mg/kg oxymatrine significantly inhibit liver metastasis of CRC cells in mice, and PKM2 and GLUT1 expression in liver of the oxymatrine-treated group is declined.
Conclusion: Oxymatrine exerted anti-metastatic activity dependent on inhibition of PKM2-mediated aerobic glycolysis. It is not only an anti-cancer agent but also a potential anti-metastatic compound with clinical application significance.
Keywords: oxymatrine, colorectal cancer, aerobic glycolysis, PKM2, GLUT1
Introduction
Nowadays, colorectal cancer (CRC) is a common cancer, with high incidence and mortality all over the world. In 2018, the number of patients diagnosed with colorectal cancer exceeded 2 million, and about 900,000 died. Global statistics indicate that the colorectal cancer patients are distributed widely in the world, especially in Europe, north America, Australia, and Japan.1 At present, following deep research on CRC and the invention of new drugs, a majority of CRC patients are receiving good treatment.2 However, CRC patients still have a high rate of metastasis. Although 5-year overall survival has tremendously increased in the past two decades, lots of colorectal cancer patients have died due to uncontrolled metastasis.3,4
Liver is the most common organ where colorectal cancer cells spread to. Approximately 20% of CRC patients have been probed with CRC cells in the liver at the first time of diagnosis,5 and another about 20% of cured CRC patients developed liver metastasis within 5 years from the first diagnosis.6 Thus, it is important to exactly evaluate the situation of CRC patients with metastasis. Generally, a multidisciplinary evaluation was made to choose the appropriate therapy dependent on the metastatic organ, the number of metastatic foci, and the potential for surgical resection.7 Nowadays, the rational therapy for colorectal liver metastases includes surgical resection, adjuvant chemotherapy, and neoadjuvant chemotherapy.8 Obviously, chemotherapy till now has been the major way to protect against metastasis in clinic. The chemotherapeutic drugs include 5-fu usually used before 1990s, oxaliplatin, capecitabine, and irinotecan after 1990s. At present, monoclonal antibodies such as bevacizumab have gradually become the favorite agents for colorectal cancer.9 However, most chemotherapeutic drugs could induce severe toxicity. So looking for highly effective but lowly toxic compounds from natural plants is always a research hotspot.
The root of Sophora flavescens Ait., named “kushen” in Chinese, was used to treat fever and infectious inflammation for a long time.10 This traditional Chinese medicine contains a lot of compounds mainly including alkaloids and flavonoids. Among alkaloids, matrine and oxymatrine, two tetracyclic quinazine compounds were demonstrated with a similar effect such as anti-inflammation, anti-virus, sedation and analgesia, cardiotonic effect, and hypotension.11 Recent decades, the anti-cancer activity of matrine and its derivatives, were gradually concerned.12 Oxymatrine as an analog of matrine also has been highly noted and its anti-cancer activity were focused on the liver, cervix, stomach carcinoma, and leukemia. The molecular mechanism research indicates that oxymatrine can inhibit cancer proliferation,13 induce cancer cell apoptosis,14 and differentiation, and downregulate the expression of epidermal growth factor receptor (EGFR).15 In the meantime, the study of oxymatrine affecting cancer metastasis is becoming a hotspot of searching anti-metastatic agents.
Metastasis can be induced by all kinds of factors which are hard to be completely elucidated till now. Among them, energy metabolism, specially the glucose metabolism, acts in a critical role in cancer metastasis.16 GLUTs, the proteins crossing the cell membrane, are responsible for importing the glucose from the extracellular circumstance to maintain cancer cell metastatic behavior. PK controls the transition from glucose to lactate in the process of glycolysis. Thus, both GLUTs and PK are indispensable during the whole aerobic glycolysis.17 Here, we found that oxymatrine exerts significant inhibition on colorectal cancer cells metastasis, and this process is dependent on PKM2/GLUT1-mediated aerobic glycolysis. It suggests oxymatrine possibly has the significance of anti-metastasis in clinic.
Materials and Methods
Chemicals and Bioreagents
Oxymatrine (purity 95%) was purchased from Chengdu Must Bio-Technology Co., Ltd (Chengdu, China). The medium including RPMI 1640, Modified Eagle Medium (DMEM), and fetal bovine serum (FBS), trypsin-EDTA were from Gibco (Grand Island, NY, USA). PKM2, GLUT1, primary antibodies were purchased from Abcam. Antibodies against LDHA, HK, and PFK were purchased from CST (Cell Signaling Technology), β-actin from ABclonal.
Cell Culture and Proliferation Assay
Human colorectal cancer cells (HT-29 and HCT-116) were purchased from Cell bank of Chinese Academy of Sciences (Shanghai, China). Both cells were qualified for test of mycoplasma contamination. A density of 104 cells/well were seeded into a 96‐well plate for overnight incubation. Then the solution with oxymatrine was added into the wells (oxymatrine final concentration=0–40 μM) for 48-hour treatment. Finally, 20 μL MTS reagent was piped into each well for chromogenic reaction. And the optical density was measured at 570 nm by a microplate reader (Biotek Corporation).
Wound Healing Assay
HT29 and HCT-116 cells (1.5×106 cells per well), seeded into 6-well plates and grown into full confluent monolayer cultures, were scraped. Then the basal medium was replaced with different concentrations of oxymatrine or DMSO for 24 hours. The ZEISS microscope was used to photograph the cell migration images at 0, 24, and 48 hours following scraping at three to four different locations. The image J was used to measure and analyze the wound closure.
Transwell Migration and Invasion Assay
A Transwell system (Corning, NY, USA) was used for evaluating cell migration and invasion. A transwell chamber precoated with or without 1–2 mg/mL Matrigel was used to determine cell invasion and migration, respectively. HT29 and HCT-116 (4×105 cells per transwell) cells were seeded into the upper chamber with serum-free medium. Then the cells were incubated with oxymatrine (1, 5, 10 μM), and 800 μL of medium with 30% FBS was added to the lower chamber. After 24-hour incubation at 37°C with 5% CO2, the medium in the upper chamber was removed and the non-migrating or non-invading cells were scraped by a cotton swab. After being fixed in 4% paraformaldehyde, the remaining cells were stained with 1% crystal violet solution for analysis. The cell photographs were taken by a Zeiss microscope, and the number of invaded and migrated cells were counted at 200× magnification.
Oxygen Consumption Rates and Extracellular Acidification Rates Assay
The Seahorse XF Glycolysis Stress Test Kit (Seahorse Biosciences, USA) and the Seahorse XFe24 Extracellular Flux Analyzer (Seahorse Biosciences, USA) were used to measure the glycolytic function in cells according to the manufacturer’s instructions. Seahorse XF Cell Mito Stress Test Kit (Seahorse Biosciences, USA) was used for determining the cellular mitochondrial capacity in accordance with the instructions. 1×104 cells were seeded into the 24-well plate for incubation overnight. Then the cells were treated with various oxymatrine for 24 hours before the test operation. After the cells were washed with Seahorse assay medium, automatic injection of 10 μM oligomycin, 2.5 μM FCCP, and 5 μM rotenone/antimycin A was to measure the oxygen consumption rates (OCR). And 100 mM glucose, 10 μM oligomycin, and 500 mM 2-deoxy-glucose (2-DG) were for determining the extracellular acidification rates (ECAR). Finally, the OCR and ECAR values were calculated by normalization to the cell number and the curve of OCR and ECAR plotted by the mean±standard deviation (SD).
Hepatic Metastasis Model of Colorectal Cancer Cells in Mice
The nude mice were anesthetized with 3% isoflurane and fixed on a mousepad. An incision was cut in the abdomen near the spleen and the spleen was exteriorized. Then 2×105 HT29 cells were injected into one side of the spleen. Ten minutes later, the injected side of the spleen was incised and the residual spleen was returned to the abdominal cavity followed with suture of the abdominal incision. The same operation but without injection of cancer cells was executed in the sham group. Seven days later, the mice with the injection of cancer cells were randomly divided into three groups, and two of them were administrated orally with oxymatrine (10 mg/kg and 20 mg/kg) but one, as the control group, with saline solution. After the 3-week administration, the mice were treated for metastatic analysis, HE staining, and immunohistochemistry analysis. The animal experiments were approved by the Ethics Committee of Nanjing University of Chinese Medicine, and the procedure strictly complies with the guidelines of the Administration of Affairs Concerning Experimental Animals approved by the State Council of People’s Republic of China.
Western Blot Analysis
Colorectal cancer cells were lysed in corresponding buffer with 1 mM phenylmethanesulfonyl fluoride (PMSF) and protein concentration was measured by bicinchoninic acid assay. Then cell lysates with 40 µg of protein were in turn loaded in SDS-PAGE (10%) for separation and transferred to polyvinylidene fluoride (PVDF) membranes. And the membrane containing the target protein was incubated with 5% non-fat milk solution for blocking nonspecific binding, with primary antibodies overnight at 4°C cold room for first immunoreaction, with appropriate secondary antibodies conjugated with horseradish peroxidase for second immunoreaction. Immunoreactive bands were visualized by enhanced chemiluminescence reagent for chromogenic reaction and photographed by Biorad gel imager.
Immunofluorescent Assay
Colorectal cancer cells were seeded on glass coverslips which were laid in a 6-well plate. After 24-hour treatment with oxymatrine, the cells were respectively fixed in 4% paraformaldehyde, permeabilized with 0.2% TritonX-100, and blocked in 1% BSA. Then the cells were in turn incubated with corresponding primary antibodies and secondary antibody obtained from Abcam and Hoechst nuclear dye. Finally, the photograph was taken and analyzed by fluorescent microscopy.
Immunohistochemistry Staining
The formalin-fixed, paraffin-embedded lung and liver tissues were cut into 5 μm sections for immunohistochemistry staining. Then the antigen was retrieved by citrate buffer (0.01 mL, pH 6.0), the sections were washed three times with PBS and incubated in endogenous peroxidase blockers for 10 minutes. The sections were then incubated overnight with antibodies at 4°C. After incubating with reaction enhancer before horseradish peroxidase-labeled anti-rabbit/mouse IgG antibody for 20 minutes at room temperature, the slides were prepared with DAB and analyzed by Olympus BX43 (×200).
Statistical Analysis
The results were determined using Student’s t-test and ANOVA test by GraphPad Prism software. P<0.05 was considered statistically significant.
Results
Oxymatrine Inhibits Colorectal Cancer Cells Metastatic Property in vitro
To understand the effect of oxymatrine on cancer metastasis, the concentrations of having no inhibition on cancer cells were screened firstly. The results indicated oxymatrine less than 10 μM rarely inhibits cell proliferation (Figure S1). Therefore, the concentrations of no less than 10 μM were chosen for the subsequent in vitro experiments.
Based on the data of wound healing assay, oxymatrine significantly inhibits the horizontal migration of HT29 under treatment of all concentrations (Figure 1A and B). The transwell assay without Matrigel showed oxymatrine can block the cell vertical migration after 24 hours of treatment (Figure 1C and D). Furthermore, the data from the transwell assay with Matrigel clued that the number of HT29 cells successfully penetrating the basement membrane-coated chambers in the control group is obviously more than in the oxymatrine-treated group (Figure 1E and F), demonstrating oxymatrine exerts an inhibitory effect in cell invasion. And similar results were repeated in human colorectal cancer HCT-116 cells (Figure S2).
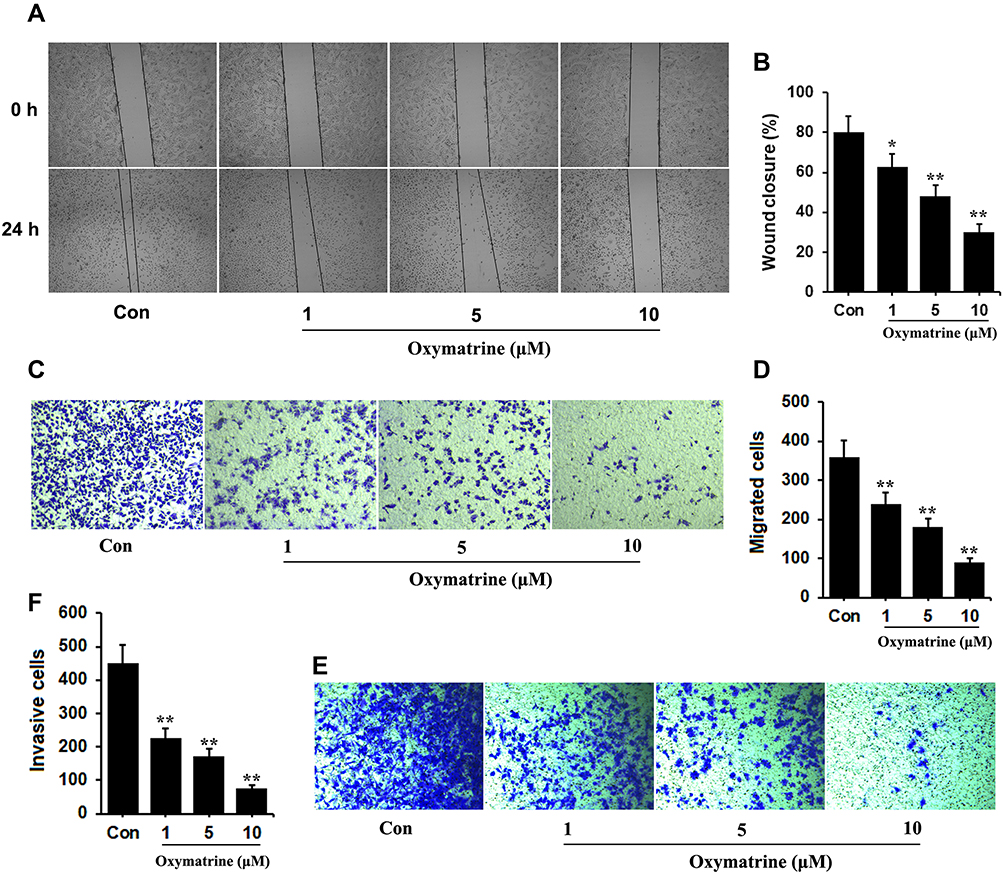 |
Figure 1 Oxymatrine inhibits migration and invasion of colorectal cancer cells in vitro. HT29 cells were seeded in six-well plates and treated with oxymatrine for 24 hours. (A) Representative images (40×) of oxymatrine-treated groups by wound healing assay. (B) The quantitative analysis of images of wound healing assay. (C and D) Representative images (200×) of oxymatrine-treated groups by transwell migration assay. The number of migrated cells was quantified. (E and F) Representative images (200×) of oxymatrine-treated groups by transwell invasion assay. The quantitative data is presented as mean±SD. Statistical difference was made by the one‐way ANOVA, *P<0.05, **P<0.01 (versus Control). |
Oxymatrine Blocks the Process of Aerobic Glycolysis in Colorectal Cancer Cells
Aerobic glycolysis is a characterized energy metabolism for cancer cells. And it has been tightly related to metastasis. Here, we tried to know whether oxymatrine inhibition of metastasis is associated with aerobic glycolysis. The data in Figure 2 show that ATP production of HT29 cells was concentration-dependently inhibited by oxymatrine (Figure 2A); the consumption of glucose and the production of pyruvate were decreased, as well as the production of lactate (Figure 2D–F). To further discover how oxymatrine affects glucose metabolism, the OCR and ECAR were determined using the XFe24 Extracellular Flux analyzer. It was found that oxymatrine can significantly inhibit ECAR in HT29 cells (Figure 2C), while the OCR has not been affected. This means that the energy metabolic form of HT29 cells was changed under the treatment of oxymatrine, but the transition to oxidative form was not observed. Furthermore, the same determination was duplicated in HCT-116 cells and similar data are shown in Figure S3. Totally, these results indicate that oxymatrine inhibits the glycolysis of colorectal cancer cells.
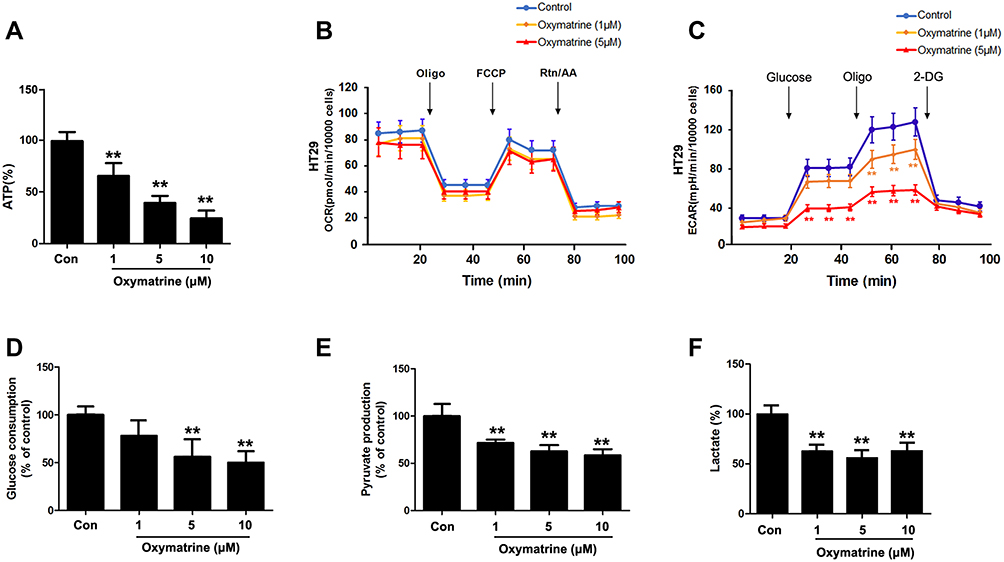 |
Figure 2 Oxymatrine inhibits aerobic glycolysis of colorectal cancer cells. HT29 cells were seeded in 24-well plates and treated with oxymatrine for 24 hours. Then the production of ATP (A), pyruvate (E), lactate (F), and the consumption of glucose (D) were measured by the corresponding determination kits. OCR (B) and ECAR (C) were determined in HT29 cells treated with oxymatrine. Quantitative data is presented as mean±SD, statistical difference was made by the one‐way ANOVA, vs control, **P<0.01. |
Oxymatrine Limits Rate-Limiting Enzyme PKM2
Three key rate-limiting enzymes HK, PFK, and PK are involved in controlling the glucose metabolism. To further understand the mechanism of oxymatrine inhibition of aerobic glycolysis, the kinase activity was measured. As shown in Figure 3, PK activity was obviously suppressed when the oxymatrine concentration is no less than 5 μM (Figure 3A), whereas the activity of HK and PFK keeps stable, suggesting that PK was the target of oxymatrine inhibition of glycolysis (Figure 3B and C). Additionally, the Western blot assay confirmed the same results as the kinase activity. The expression of PKM2, an isoform of PK, was declined, while HK and PFK expressed unchanged (Figure 3D and E). Furthermore, PKM2 mRNA expression was also decreased (Figure 3F). According to the above results, PKM2 is an important molecule mediating oxymatrine inhibition of metastasis.
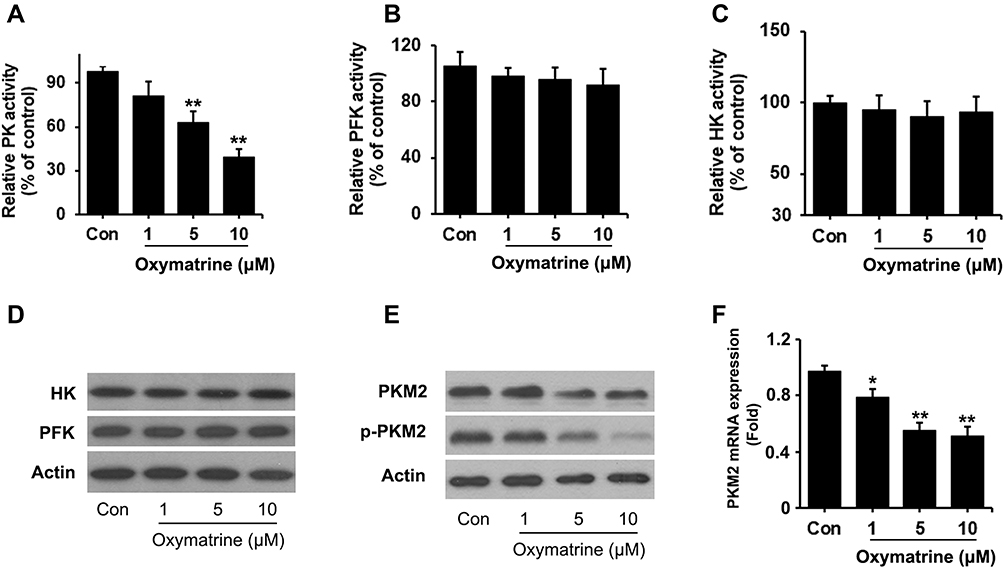 |
Figure 3 Oxymatrine inhibits kinase activity and expression of PKM2. HT29 cells were seeded in 6-well plates and treated with oxymatrine for 24 hours. Then the activity of PK (A), PFK (B), and HK (C) was measured using the corresponding kits. (D and E) Western blotting assay for HK, PFK, PKM2, and p-PKM2 was executed. (F) mRNA expression of PKM2 was detected. Quantitative data is presented as mean±SD, statistical difference was made by the one‐way ANOVA, versus control, *P<0.05, **P<0.01. |
Oxymatrine Inhibits Expression of GLUT1
Many other factors participated in the process of glycolysis. Glucose transporter 1 (GLUT1) and lactate dehydrogenase A (LDHA) were two proteins among them. GLUT1 responsible for importing the glucose from the extracellular side plays an initial role in glycolysis, while LDHA for transforming pyruvate to lactate acts as a last step to produce energy. As indicated in Figure 4, GLUT1 expression was inhibited in a concentration-dependent manner, but LDHA expression fluctuated slightly under the treatment of oxymatrine. And the further immunofluorescent data also justified GLUT1 was downregulated by 10 μM oxymatrine. Thus, GLUT1 mediates oxymatrine-induced aerobic glycolysis blocked.
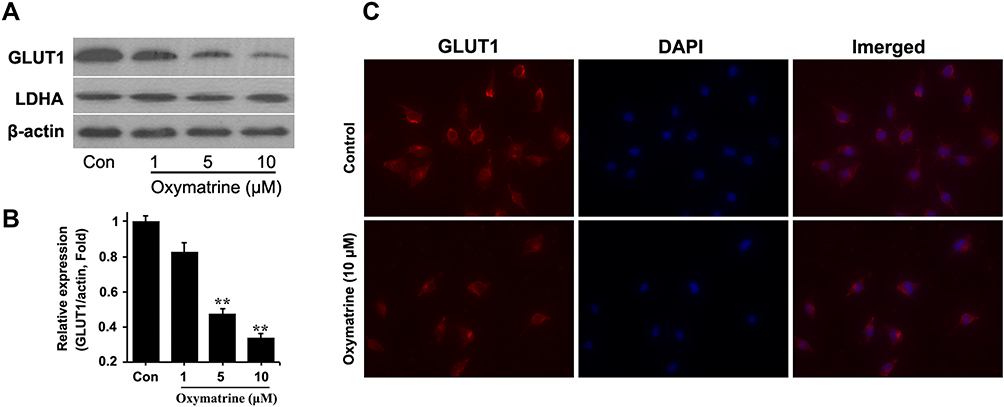 |
Figure 4 Oxymatrine inhibits expression of GLUT1. HT29 cells were seeded in 6-well plates and treated with oxymatrine for 24 hours. Then the Western blotting assay (A) and immunofluorescent staining (C) were used for determining the GLUT1 expression. The Western blot band was semi-quantified by image J (B). Quantitative data is presented as mean±SD, statistical difference was made by the one‐way ANOVA, versus control, **P<0.01. |
PKM2 Activator Reverses Oxymatrine Inhibition of Glycolysis
In order to further demonstrate the importance of PKM2 in oxymatrine inhibition of glycolysis-mediated metastasis, TEPP-46, a highly effective and selective PKM2 activator was chosen to conversely support it. As shown in Figure 5, oxymatrine inhibition of cancer cell migration at 10 μM was reversed when oxymatrine and TEPP-46 were together added for treatment of cells (Figure 5A). Similarly, the transwell assay indicated the same result as the wound healing assay (Figure 5B). In addition, the glycolytic intermediates downregulated by oxymatrine had a significant increase after TEPP-46 addition. Seen from Figure 5, the pyruvate production and the glucose consumption of HT29 cells were recovered significantly by TEPP-46 (Figure 5C and D), while lactate production also kept a low level (Figure 5E). Totally, oxymatrine inhibition of cancer metastasis via depressing PKM2 could be neutralized by PKM2 activator, further suggesting that PKM2 is a necessary factor mediating oxymatrine’s effect on metastasis.
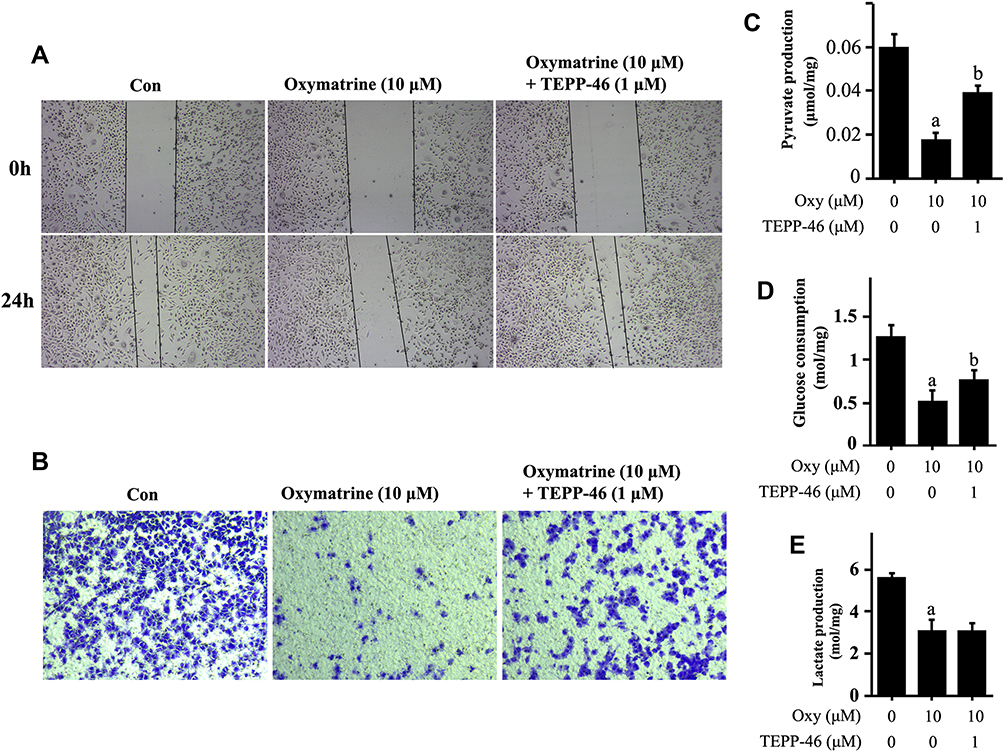 |
Figure 5 PKM2 activator, TEPP-46 could partially reverse oxymatrine’s effect on metastasis and glycolytic production. HT29 cells seeded in six-well plates were treated in turn with TEPP-46 and oxymatrine for 24 hours, then wound healing assay (A) and transwell assay (B) were used to confirm the response of HT29 cells. And the production of pyruvate (C) and lactate (E) and the glucose consumption (D) were tested. Quantitative data is presented as mean±SD, statistical difference was made by the one‐way ANOVA. Compared with the untreated group, aP<0.01; Compared with the oxymatrine-treated group, bP<0.01. |
Oxymatrine Inhibits Hepatic Metastasis of Colorectal Cancer Cells in vivo
According to Figure 6, the liver of control mice exists visible metastatic nodules, and the number of metastatic nodules in the oxymatrine-treated groups is significantly less than that in the control group (Figure 6A). HE staining indicated that the area of metastasis in liver tissue of untreated mice was obviously larger and formed a consecutive region, while in the treated group the metastatic area was limited in a small and dispersed region (Figure 6B). Based on the above results, it is concluded that oxymatrine can inhibit hepatic metastasis of colorectal cancer cells in vivo. Furthermore, the expression of PKM2 and GLUT1 was determined by immunohistochemistry. As indicated in Figure 6, compared with the control, the expression of PKM2 in the oxymatrine group was suppressed, as well as the expression of GLUT1 (Figure 6C). This in vivo result is consistent with the in vitro data, supporting the conclusion that oxymatrine inhibits the metastasis via impeding the GLUT1-mediated PKM2 signaling pathway.
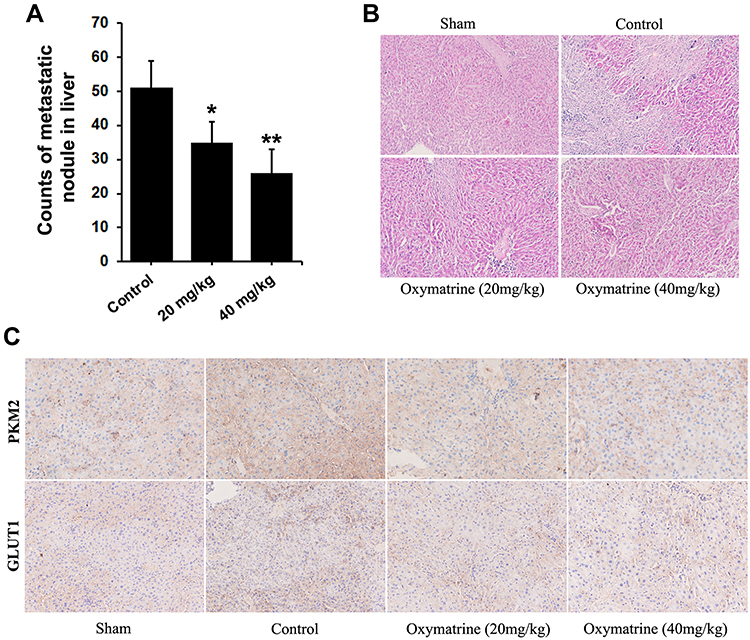 |
Figure 6 Oxymatrine inhibits hepatic metastasis of colorectal cancer cells in vivo. (A) The counts of metastatic module in liver of mice; (B) H&E staining of the liver tissues of mice xenografted with HT29 cells; (C) The immunochemistry staining for PKM2 and GLUT1 in liver. Quantitative data is presented as mean±SD, statistical difference was made by the one‐way ANOVA, versus control, *P<0.05, **P<0.01. |
Discussion
Oxymatrine has been proved with inhibitory effect on a series of cancer cells. Oxymatrine can induce pancreatic cancer cells apoptosis through affecting the ratio of Bax/Bcl2 and release of mitochondrial cytochrome c,18 and cause the human bladder cancer cell apoptosis and cell cycle arrest.19 Oxymatrine inhibits cell proliferation via attenuation of the RAS/ERK pathway,20 abolishing the PI3K expression,21,22 suppression of EGFR signaling,15 and deactivating STAT5.23 Furthermore, oxymatrine combined with chemotherapeutic agents could enhance their anti-cancer effect. Oxymatrine synergistically enhances the antitumor effects of cisplatin in human gastric cancer cells,24 non-small cell lung cancer,25 and anti-cancer effects of bevacizumab against triple-negative breast cancer via impeding expression of Wnt/β-Catenin.26 For an aspect of resistance, oxymatrine can inhibit epithelial-mesenchymal transition and NF-κB signaling, leading to the reverse of 5-fluorouracil resistance in colon cancer cells.27 The above results indicated the anti-cancer research on oxymatrine is considerably focused. However, the understanding of oxymatrine on metastasis is rarely elucidated. Chen et al28 reported oxymatrine can block the metastasis of hepatocellular carcinoma cells in vitro by reducing p38-mediated MMP-2/-9 expression. Here, not only oxymatrine inhibition of metastasis in colorectal cancer cells, but also the molecular mechanism was demonstrated.
The glucose metabolism of cancer cells is completely different than normal cells. Warburg,29 in 1924, found that cancer cells consume plenty of glucose by a high-rate but lowly efficient way to provide ATP and produce lactate even in the presence of oxygen, which was named the Warburg effect. This is also called aerobic glycolysis because the glycolysis occurred even if the oxygen is sufficient.30 Actually, aerobic glycolysis is independent of oxygen supply and produces large amounts of lactate.31 The accumulation of lactate can decrease pH value of the local tumor region, and stimulate the obvious change of tumor microenvironment.32 This enhances the motility, migration, and invasion of cancer cells, and causes cancer cells to penetrate the vessel to metastasize to the remote tissues.32 Thus, aerobic glycolysis can promote cancer metastasis.33 In this article, we found that oxymatrine exerts a significant inhibition on colorectal cancer cell migration and invasion, and the ATP production was decreased, as well as the production of lactate, suggesting that oxymatrine inhibition of metastasis is related with suppression of aerobic glycolysis.
Aerobic glycolysis is controlled by many kinases including HK, PFK, and PKM2. HK and PFK were justified not to be affected by oxymatrine, while PK activity was inhibited. PKM2, an isoform of pyruvate kinase, regulates the glycolytic intermediates and shifts the glucose metabolism in favor of cancer cells.34,35 The majority of tumor tissues and cancer cell lines express high PKM2.35,36 Nowadays, targeting metabolism of cancer cells is regarded as a new way to against cancer. PKM2 has been considered a potential target for cancer therapy, boosting the research of PKM2 inhibitors.34 There are three ways to affect PKM2: inhibition of PKM2 expression, decreasing kinase activity of PKM2, and activation of tetrameric PKM2 form.34 Silencing the PKM2 via siRNA attenuates proliferation and induces apoptosis in many cancer cell lines.37 Several compounds have demonstrated that inhibition of PKM2 activity results in cancer cell death and apoptosis, indicating PKM2 activity is also responsible for cell growth.38 In addition, the researchers also thought activation of highly active tetrameric PKM2 can inhibit cell proliferation as the precursors for the synthesis of the cell were deficient.39 Oxymatrine can not only attenuate the activity of PKM2 but also inhibits its expression, meaning that the characteristic of oxymatrine on PKM2 is dual inhibition. In this study, whether oxymatrine may activate PKM2 dimer to tetramer is not confirmed.
Membrane protein GLUT family starts the first step of glucose metabolism. The high expression of GLUTs has been confirmed in a series of cancer cells. GLUT1, a subunit of 14 GLUT proteins, shares an important role in GLUTs family and tumor environment and glycolysis.40 GLUT1 overexpression responds to many signals such as HIF-1a, VEGF receptor, and calcium channel transactivation.41 But as a transcriptional factor, GLUT1 regulation of glycolysis is under control by a lot of molecules. Particularly, in colorectal cancer cells, KRAS or BRAF mutation is tightly related with GLUT1 expression.42 Oxymatrine inhibition of GLUT1 may be dependent on the mutant KRAS or BRAF in HT29 cells. This also illustrated that oxymatrine inhibition of PKM2-controlled glycolysis is dependent on GLUT1. However, whether oxymatrine inhibits PKM2 leading to GLUT1 downregulation or downregulation of GLUT1 results in PKM2 inactivation is unclear, and this needs to be further investigated.
Totally, a potent anti-cancer compound oxymatrine in this study was demonstrated with potential anti-metastatic activity in colorectal cancer cells, and this depends on the dual inhibition of PKM2 and downregulation of GLUT1 expression.
Funding
This work was supported by National Natural Science Foundation of China (NSFC) (No. 81973482).
Disclosure
The authors declare that they have no conflict of interest.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. doi:10.3322/caac.21492
2. Xie YH, Chen YX, Fang JY. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct Target Ther. 2020;5:22.
3. Kopetz S, Chang GJ, Overman MJ, et al. Improved survival in metastatic colorectal cancer is associated with adoption of hepatic resection and improved chemotherapy. J Clin Oncol. 2009;27:3677–3683. doi:10.1200/JCO.2008.20.5278
4. Huang D, Sun W, Zhou Y, et al. Mutations of key driver genes in colorectal cancer progression and metastasis. Cancer Metastasis Rev. 2018;37:173–187.
5. van der Geest LG, Lam-Boer J, Koopman M, Verhoef C, Elferink MA, de Wilt JH. Nationwide trends in incidence, treatment and survival of colorectal cancer patients with synchronous metastases. Clin Exp Metastasis. 2015;32:457–465. doi:10.1007/s10585-015-9719-0
6. van Gestel YR, de Hingh IH, van Herk-Sukel MP, et al. Patterns of metachronous metastases after curative treatment of colorectal cancer. Cancer Epidemiol. 2014;38:448–454.
7. De Andrade JP, Warner SG, Fong Y. Treatment of metastatic colorectal cancer: innovations in surgical techniques. J Surg Oncol. 2019;119:653–659. doi:10.1002/jso.25418
8. Piawah S, Venook AP. Targeted therapy for colorectal cancer metastases: a review of current methods of molecularly targeted therapy and the use of tumor biomarkers in the treatment of metastatic colorectal cancer. Cancer. 2019;125:4139–4147. doi:10.1002/cncr.32163
9. Sveen A, Kopetz S, Lothe RA. Biomarker-guided therapy for colorectal cancer: strength in complexity. Nat Rev Clin Oncol. 2020;17:11–32.
10. Wang W, You R-L, Qin W-J, et al. Anti-tumor activities of active ingredients in Compound Kushen Injection. Acta Pharmacol Sin. 2015;36:676–679. doi:10.1038/aps.2015.24
11. Yamazaki M. The pharmacological studies on matrine and oxymatrine. Yakugaku Zasshi. 2000;120:1025–1033.
12. Yong J, Wu X, Lu C. Anticancer advances of matrine and its derivatives. Curr Pharm Des. 2015;21:3673–3680. doi:10.2174/1381612821666150122123748
13. Wu C, Huang W, Guo Y, et al. Oxymatrine inhibits the proliferation of prostate cancer cells in vitro and in vivo. Mol Med Rep. 2015;11:4129–4134. doi:10.3892/mmr.2015.3338
14. Li M, Su B-S, Chang L-H, et al. Oxymatrine induces apoptosis in human cervical cancer cells through guanine nucleotide depletion. Anticancer Drugs. 2014;25:161–173. doi:10.1097/CAD.0000000000000012
15. Li W, Yu X, Tan S, Liu W, Zhou L, Liu H. Oxymatrine inhibits non-small cell lung cancer via suppression of EGFR signaling pathway. Cancer Med. 2018;7:208–218. doi:10.1002/cam4.1269
16. McDonald OG, Li X, Saunders T, et al. Epigenomic reprogramming during pancreatic cancer progression links anabolic glucose metabolism to distant metastasis. Nat Genet. 2017;49(3):367–376. doi:10.1038/ng.3753
17. Sheng H, Tang W. Glycolysis inhibitors for anticancer therapy: a review of recent patents. Recent Pat Anticancer Drug Discov. 2016;11(3):297–308. doi:10.2174/1574892811666160415160104
18. Ling Q, Xu X, Wei X. Oxymatrine induces human pancreatic cancer PANC-1 cells apoptosis via regulating expression of Bcl-2 and IAP families, and releasing of cytochrome c. J Exp Clin Cancer Res. 2011;30:66. doi:10.1186/1756-9966-30-66
19. Li S, Zhang Y, Liu Q, et al. Oxymatrine inhibits proliferation of human bladder cancer T24 cells by inducing apoptosis and cell cycle arrest. Oncol Lett. 2017;13:4453–4458. doi:10.3892/ol.2017.6013
20. Wang Y, Yang S, Zhang S, Wu X. Oxymatrine inhibits proliferation and migration of vulvar squamous cell carcinoma cells via attenuation of the RAS/RAF/MEK/ERK pathway. Cancer Manag Res. 2020;12:2057–2067. doi:10.2147/CMAR.S245696
21. Guo L, Yang T. Oxymatrine inhibits the proliferation and invasion of breast cancer cells via the PI3K pathway. Cancer Manag Res. 2019;11:10499–10508. doi:10.2147/CMAR.S221950
22. Ni Z, Yi J. Oxymatrine induces nasopharyngeal cancer cell death through inhibition of PI3K/AKT and NF‑κB pathways. Mol Med Rep. 2017;16:9701–9706. doi:10.3892/mmr.2017.7822
23. Jung YY, Shanmugam MK, Narula AS, et al. Oxymatrine attenuates tumor growth and deactivates STAT5 signaling in a lung cancer xenograft model. Cancers (Basel). 2019;11:49. doi:10.3390/cancers11010049
24. Liu Y, Qin L, Bi T, et al. Oxymatrine synergistically potentiates the antitumor effects of cisplatin in human gastric cancer cells. J Cancer. 2018;9:4527–4535. doi:10.7150/jca.28532
25. Ye J, Zou -M-M, Li P, et al. Oxymatrine and cisplatin synergistically enhance anti-tumor immunity of CD8+ T cells in non-small cell lung cancer. Front Oncol. 2018;8:631. doi:10.3389/fonc.2018.00631
26. Xie W, Zhang Y, Zhang S, et al. Oxymatrine enhanced anti-tumor effects of Bevacizumab against triple-negative breast cancer via abating Wnt/β-Catenin signaling pathway. Am J Cancer Res. 2019;9:1796–1814.
27. Liang L, Wu J, Luo J, et al. Oxymatrine reverses 5-fluorouracil resistance by inhibition of colon cancer cell epithelial-mesenchymal transition and NF-κB signaling in vitro. Oncol Lett. 2020;19:519–526. doi:10.3892/ol.2019.11090
28. Chen K, Zhu P, Ye J, et al. Oxymatrine inhibits the migration and invasion of hepatocellular carcinoma cells by reducing the activity of MMP-2/-9 via regulating p38 signaling pathway. J Cancer. 2019;10:5397–5403. doi:10.7150/jca.32875
29. Warburg O. On the origin of cancer cells. Science. 1956;123:309–314. doi:10.1126/science.123.3191.309
30. Dang CV, Semenza GL. Oncogenic alterations of metabolism. Trends Biochem Sci. 1999;24:68–72. doi:10.1016/S0968-0004(98)01344-9
31. Hsu PP, Sabatini DM. Cancer cell metabolism: warburg and beyond. Cell. 2008;134:703–707. doi:10.1016/j.cell.2008.08.021
32. Chen C, Bai L, Cao F, et al. Targeting LIN28B reprograms tumor glucose metabolism and acidic microenvironment to suppress cancer stemness and metastasis. Oncogene. 2019;38:4527–4539. doi:10.1038/s41388-019-0735-4
33. Vander HMG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009;324:1029–1033. doi:10.1126/science.1160809
34. Iqbal MA, Gupta V, Gopinath P, Mazurek S, Bamezai RN. Pyruvate kinase M2 and cancer: an updated assessment. FEBS Lett. 2014;588:2685–2692. doi:10.1016/j.febslet.2014.04.011
35. Yang W, Xia Y, Ji H, et al. Nuclear PKM2 regulates β-catenin transactivation upon EGFR activation. Nature. 2011;480:118–122. doi:10.1038/nature10598
36. Jiang Y, Li X, Yang W, et al. PKM2 regulates chromosome segregation and mitosis progression of tumor cells. Mol Cell. 2014;53:75–87. doi:10.1016/j.molcel.2013.11.001
37. Goldberg MS, Sharp PA. Pyruvate kinase M2-specific siRNA induces apoptosis and tumor regression. J Exp Med. 2012;209:217–224. doi:10.1084/jem.20111487
38. Vander HMG, Christofk HR, Schuman E, et al. Identification of small molecule inhibitors of pyruvate kinase M2. Biochem Pharmacol. 2010;79:1118–1124. doi:10.1016/j.bcp.2009.12.003
39. Boxer MB, Jiang J-K, Vander Heiden MG, et al. Evaluation of substituted N, N′-Diarylsulfonamides as activators of the tumor cell specific M2 isoform of pyruvate kinase. J Med Chem. 2010;53:1048–1055. doi:10.1021/jm901577g
40. Ancey PB, Contat C, Meylan E. Glucose transporters in cancer - from tumor cells to the tumor microenvironment. FEBS J. 2018;285:2926–2943. doi:10.1111/febs.14577
41. Suh HN, Han HJ. Fibronectin-induced VEGF receptor and calcium channel transactivation stimulate GLUT-1 synthesis and trafficking through PPARγ and TC10 in mouse embryonic stem cells. Stem Cell Res. 2013;10:371–386. doi:10.1016/j.scr.2013.01.008
42. Yun J, Rago C, Cheong I, et al. Glucose deprivation contributes to the development of KRAS pathway mutations in tumor cells. Science. 2009;325:1555–1559. doi:10.1126/science.1174229
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.