")
Back to Journals » Infection and Drug Resistance » Volume 15
Novel Pseudomonas aeruginosa Strains Co-Harbouring blaNDM-1 Metallo β-Lactamase and mcr-1 Isolated from Immunocompromised Paediatric Patients
Authors Chen H, Mai H, Lopes B, Wen F, Patil S
Received 29 March 2022
Accepted for publication 27 May 2022
Published 8 June 2022 Volume 2022:15 Pages 2929—2936
DOI https://doi.org/10.2147/IDR.S368566
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Video abstract presented by Sandip Patil.
Views: 799
Hongyu Chen,1 Huirong Mai,2 Bruno Lopes,3 Feiqiu Wen,2 Sandip Patil2,4
1Department of Laboratory Medicine, Shenzhen Children’s Hospital, Shenzhen, Guangdong, 518038, People’s Republic of China; 2Department of Haematology and Oncology, Shenzhen Children’s Hospital, Shenzhen, Guangdong, 518038, People’s Republic of China; 3Department of Microbiology, School of Medicine, Medical Sciences and Nutrition, University of Aberdeen, Aberdeen, UK; 4Paediatric Research Institute, Shenzhen Children’s Hospital, Shenzhen, Guangdong, 518038, People’s Republic of China
Correspondence: Sandip Patil, Department of Haematology and Oncology, Shenzhen Children’s Hospital, Shenzhen, Guangdong, 518038, People’s Republic of China, Tel +86-18813934471, Fax +86-755-83008283, Email [email protected] Feiqiu Wen, Department of Haematology and Oncology, Shenzhen Children’s Hospital, Shenzhen, Guangdong, 518038, People’s Republic of China, Tel +86-18938690333, Fax +86-755-83009888, Email [email protected]
Background: The rising resistance to carbapenems in Gram-negative bacteria worldwide poses a major clinical and public health risk. This study aimed to characterise carbapenem- and colistin-resistance genes, blaNDM-1 and mcr-1 located on IncX4 plasmid in MDR Pseudomonas aeruginosa, isolated from paediatric patients undergoing chemotherapy as a result of leukaemia.
Methods: In this study, six carbapenem-resistant strains of P. aeruginosa were isolated from two paediatric patients under chemotherapy treatment (1.8 years old female and 2.1 years male) from the Shenzhen Hospital, China, in the year 2019. Isolates were screened for conventional antibiotics such as tobramycin, cefepime, imipenem, and ciprofloxacin in additional colistin by using the broth dilution method. Furthermore, resistance determinants: mcr-1, blaNDM-1, blaKPC-1, and blaGES were screened using PCR and sequencing followed by multi-locus sequence typing. The horizontal gene transfer and location of mcr-1 and blaNDM-1 were determined by a liquid mating assay. In addition, Incompatibility type (Inc), PCR-based replicon type, and subgroup (MOB) of plasmid were studied.
Results: The screening for conventional antibiotics isolates showed 100% resistance to all the tested antibiotics except tobramycin. All isolates harboured carbapenemase encoding blaNDM-1, of which three also had mcr-1 located on a single IncX4 transferable plasmid. MLST typing revealed that four strains had a novel (new) STs type, while two belonged to ST1966.
Conclusion: This study identified for the first time colistin- and carbapenem-resistant MDR P. aeruginosa in paediatric patients with leukaemia in Shenzhen, China. It highlights the need for continuous surveillance in high-risk clones of MDR P. aeruginosa. Prudent use of antibiotics based on local antimicrobial susceptibility and clinical characteristics can help in reducing mortality in immunocompromised patients.
Keywords: mcr-1, blaNDM-1, Pseudomonas aeruginosa, leukaemia patients
Introduction
Pseudomonas aeruginosa is a Gram-negative, rod-shaped, aerobic bacterium and one of the most common causes of infectious disease in paediatric patients, leading to high mortality.1 It is a very common opportunistic pathogen that can normally be present on the skin of healthy individuals but may cause serious infection in the case of immunocompromised patients with haematological malignancies.2,3 Carbapenem-resistant P. aeruginosa has been classified as a critically important pathogen by the WHO in the year 2017 because of its ability and resilience to develop antimicrobial resistance (AMR) to the most commonly used antimicrobial drugs required for life-threatening infections. Hence, new antibiotics are urgently required in order to tackle the silent pandemic of AMR.4 The acquisition of the resistance genes and genetic mutations enables the bacterium to resist antibiotics and survive in an antibiotic enriched environment where the selection pressure for the development of resistant strains is quite high.5 The resistant P. aeruginosa exposure to various antibiotics and the development of cross-resistance between antimicrobials may result in multidrug-resistant (MDR) strains which are not only difficult to treat but may also expand clonally and become endemic to a specific region such as a specific hospital. Such MDR P. aeruginosa strains may be transmitted from patient to patient and sometimes lead to outbreaks among patients attending/sharing the same ward.6 Recent data show that MDR P. aeruginosa has a high prevalence rate that varies from 15% to 30% globally in hospital settings7 In African countries alone, 13% of severe healthcare-associated infections are caused by MDR P. aeruginosa.8 The ST1966 high-risk clone of P. aeruginosa has a high capacity to acquire antibiotic resistance genes and is prevalent in many parts of China, especially in hospital settings (unpublished data). However, no study has reported the occurrence of ST1966 co-harbouring New Delhi Metallo-beta-lactamase-1 (NDM-1) and Klebsiella pneumoniae carbapenemase (KPC) in P. aeruginosa. A previous study has reported a co-existence of E. coli with NDM-1 and MCR-1.9 In addition, there has been no report on the co-existence of blaNDM-1 and plasmid-born colistin resistance mcr-1 in P. aeruginosa in paediatric patients. The occurrence and distribution of colistin resistance in P. aeruginosa clones is of huge concern, because of the fatality and limited treatment options and colistin being used as the last choice of drug to treat MDR pathogens.10 Due to the increased risk of infections caused by NDM-1 and KPC-producing pathogens and the lack of proven local information, the WHO recently pointed out the need to invest in resources to improve the surveillance of antimicrobial-resistant (AMR) clones of bacteria,11,12 using whole-genome sequencing technology.13 Hence, it is important to investigate and monitor the emergence of resistant clones and make prudent use of antibiotics in order to treat any life-threatening infections. The present study aimed to evaluate carbapenemase-producing genes and their location in P. aeruginosa strains isolated from paediatric patients undergoing chemotherapy for leukaemia in XX hospital in China.
Methods
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Ethics Committee, Shenzhen Children’s Hospital, Reference number: 2018 (013) dated 2018/09/03, which complies with international ethical standards. All experiments were conducted as per the hospital biosafety regulations act.
Isolation and Identification
In this report, six carbapenem-resistant strains of P. aeruginosa were isolated from two paediatric patients (1.8 years old female, and 2.1 years male). Both patients had a history of acute leukaemia and were admitted to our hospital for chemotherapy, from July to August 2019. The demographic and clinical data were recorded on a standardized case form. During treatment, patients developed symptoms such as fever, cough, diarrhoea, and tissue infection, which followed microbiological investigation (Table 1). To identify the pathogens, full blood, urine, peripheral blood, CSF, hydrothorax and stool were cultured on the blood agar and Macconkey’s agar. The initially isolated identification was done by using the VITEK®2 compact system (BioMerieux, France), followed by the API20 system and 16sRNA amplification. The sequencing of each purified PCR product was performed by Sangon Biotech, Shanghai. PCR assay was performed to amplify the full length of 16SRNA by using a set of universal primers: F-5’-AGAGTRTGATCMTYGCTWAC-3’; and R-5’-CGYTAMCTTWTTACGRCT-3’. PCR reaction volume was 20 µL and it contained 2µL (30ng) of genomic DNA extracted by QIAamp®DNA Kit from QIAGEN (Germany), 10 µL of 2X Master Mix, 0.4 µL (20 pmol) of each forward and reverse primer, and 8.2 µL of nuclease-free water. Thermocycler conditions were set at 94℃ for 5 min, followed by 35 cycles of denaturation at 94℃ for 30 sec, annealing at 55℃ for 30 sec and extension at 72℃ for 1min, followed by a final extension at 72℃ for 5 min. The PCR water was used as the control while P. aeruginosa ATCC 27853 DNA was used as the positive control. The PCR product was run on 1.8% agarose along with a DNA ladder (100Kb). The sequences were analyzed using NCBI BLAST N; https://blast.ncbi.nlm.nih.gov/Blast. This isolate was collected as a routine hospital investigation procedure. Only verbal consent was obtained because no personal information was used for research purposes, therefore written consent was not required.
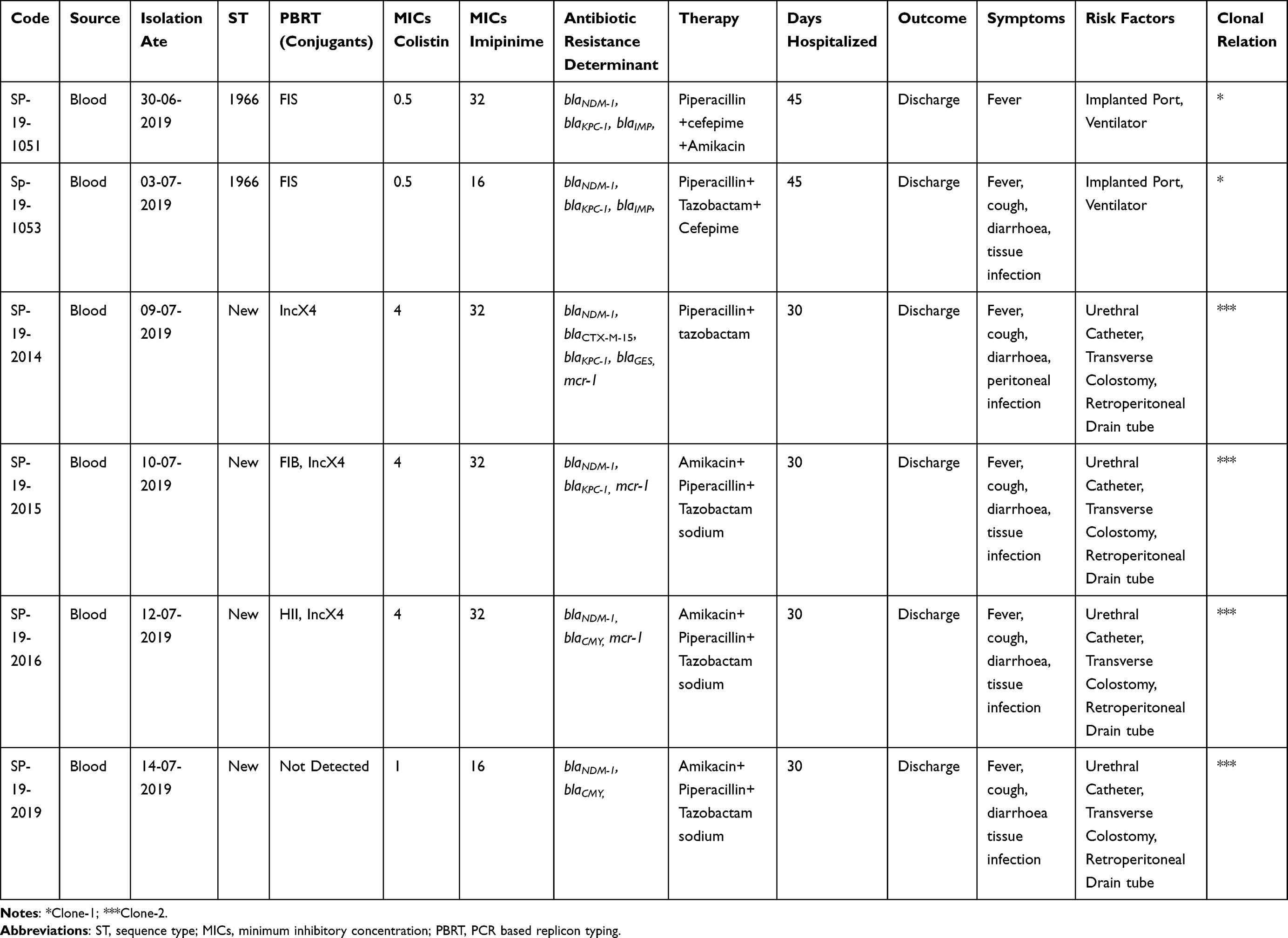 |
Table 1 Clinical and Molecular Characterization of Pseudomonas aeruginosa |
Antimicrobial Susceptibility
The antimicrobial susceptibility test (AST) was also performed using the VITEK®2 compact system (BioMerieux, France) with ASTGN09 card (software version 9.01) followed by the agar dilution method. Tested antibiotics were Amoxicillin, Tobramycin, Cefepime, Meropenem, Amikacin, Levofloxacin, Ciprofloxacin, Cefoperazone, Ticarcillin, and Colistin. The P. aeruginosa ATCC 27853 was used as a quality control strain for the AST. The carbapenemase production among carbapenem-resistant P. aeruginosa was confirmed using the modified Hodge test, while Metallo β-lactamase was determined by the MBL E-test.14 The protocol for both tests was well developed in our infection and drug resistance lab and E. coli ATCC 25992 was used as the control strain while characterized strain of E. coli from our lab was used as the positive control. Antimicrobial susceptibility excluding colistin results was interpreted according to CLSI guidelines, while colistin resistance was defined according to European Committee on Antimicrobial Susceptibility Testing (EUCAST) clinical breakpoints.15,16
Molecular Characterization
The PCR assays were performed for the detection of β-lactamase-encoding genes including blaCTX-M, blaTEM, blaSHV, blaGES, blaCARB, blaPER, blaIMP, blaCMY, blaSME, blaVIM, blaOXA, blaNDM, and blaKPC. The primers and protocol were adopted from our lab,17 whereas the PCR for colistin-resistant genes (mcr-1 to mcr-9) was performed by using primers described previously.18 The PCR conditions are well established in our laboratory. Thermocycler conditions were set at 94℃ for 5 min, followed by 35 cycles of denaturation at 94℃ for 30 sec, annealing at 55℃ for 30 sec and extension at 72℃ for 1min, followed by a final extension at 72℃ for 5 min. The amplified product was run on 1.5% agarose gel with ethidium bromide and extracted using a QIAquick® Gel Extraction Kit by QIAGEN (Germany) followed by sequencing. The Sequences were analysed by BlastN: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE= BlastSearch. The clonal relation and MLST typing were performed by amplifying conserved seven housekeeping genes including acsA, aroE, guaA, mutL, nuoD, ppsA, and trpE, according to the protocol mentioned on the P. aeruginosa MLST website (https://pubmlst.org/P.aeruginosa/). The primers for the above housekeeping genes were adopted from the same website. The amplified products were sequenced and Allelic profiles, as well as sequence types, were determined using https://pubmlst.org/bigsdb?db=pubmlstpaeruginosa_ seqdefandpage=profilesandscheme_id=1. The combination of alleles obtained at each locus defined its allelic profile or sequence type (ST). The combinations of alleles did not match with the locus database designated as a “new” ST. While the clonal relatedness was determined by the repetitive PCR (rep-PCR) (Diversilab, Biomerieux). All protocols were adapted from our laboratory.
Conjugation and Plasmid Characteristics
A mating assay was performed to analyse the horizontal gene transfer of blaNDM-1 and mcr-1 by using streptomycin-resistant E. coli C600 as the recipient strain. The Luria broth medium for both isolates was mixed and incubated for meeting at 37℃ for 24hrs. The successful transconjugants were selected on streptomycin (2000 µg/mL) and colistin (4 µg/mL) seeded Muller Hinton agar plate. While blaNDM-1 conjugants were selected on streptomycin (2000 µg/mL) and meropenem (1 μg/mL) seeded Muller Hinton agar plate. Furthermore, transconjugants were tested by PCR assay and sequencing for the presence of mcr-1 and co-existence of other genes including blaNDM-1, blaKPC, and blaCTX-M-15. The transconjugant frequency was determined as the number of transconjugants divided by the total colony-forming unit (CFU). In addition, the plasmid incompatibility (Inc) group was determined based on PCR-based replicon typing (PBRT). The primer pairs adopted based on the EMBL Gene Databank, for HI1, HI2, I1-Iγ, X, L/M, N, FIA, FIB, W, Y, P, FIC, A/C, T, FIIAS, F, K and B/O replicons results analysed as per described.19 Degenerate primer MOB typing (DPMT) means sub-groups were determined by using specific primers which were encoded in 33 references release as previously introduced by Alvarado A et al,20 while protocol was standardised in our laboratory.
Result
All P. aeruginosa isolates were confirmed by API-20 and 16s RNA sequencing. The modified Hodge test shows the inactivation of the carbapenem resulting in positive for carbapenemase production while MBL-E-test results confirmed the production of Metallo-beta-lactamase for all tested strains. The AST results revealed that in all the tested carbapenemase-producing P. aeruginosa isolates 6/6,100% were resistant to amoxicillin, amikacin and ticarcillin; while 60% of these isolates were resistant to other tested antibiotics (Figure 1). About 3/6, 50% were resistant to colistin which is the last resort of choice to treat infection caused by MDR strains. The resistant determinant results showed that all isolates harboured blaNDM-1 among which four co-harboured mcr-1. In addition, carbapenemase-producing P. aeruginosa were carrying blaKPC-1 (4/6), followed by blaIMP (3/6), blaGES (1/6), and blaCMY (1/6). Colistin use is one of the major factors responsible for the development of colistin resistance but in this study, patients were not treated with colistin (Table 1). In our study, MLST results indicate that 2/6 isolates belonged to P. aeruginosa ST1966 high-risk clone, the other four isolates allocated alleles combination were not matched with reported STs in PubMLST database so reported as new STs (Table 2). A total of three successful mcr-1 carrying transconjugants were obtained while one-SP-19-2019 failed to transfer the mcr-1 gene (Table 1). Our isolate SP-19-2019 conjugation failure may be due to the location of mcr-1 on the chromosome. The horizontal gene transfer frequency ranged from 10−3 to 10−5. Successful three conjugants co-existence of blaNDM-1 and mcr-1 which were located in IncX4 plasmid group based on the PBRT results while MOB revealed that plasmid belongs to the P-31 subgroup (Table 1). Moreover, two blaNDM-1 plasmids of transconjugants belong to IncFIS and subgroup P-12. In genotyping, we found that SP-19-1051, 1054 and 2019 were clonally related (>95%). The other three isolates (Sp-19-2014, 2015, 2016) were of the same sequence type. Both patients had invasive infections with colistin-resistant P. aeruginosa but were able to recover as a result of antimicrobial therapy and were discharged.
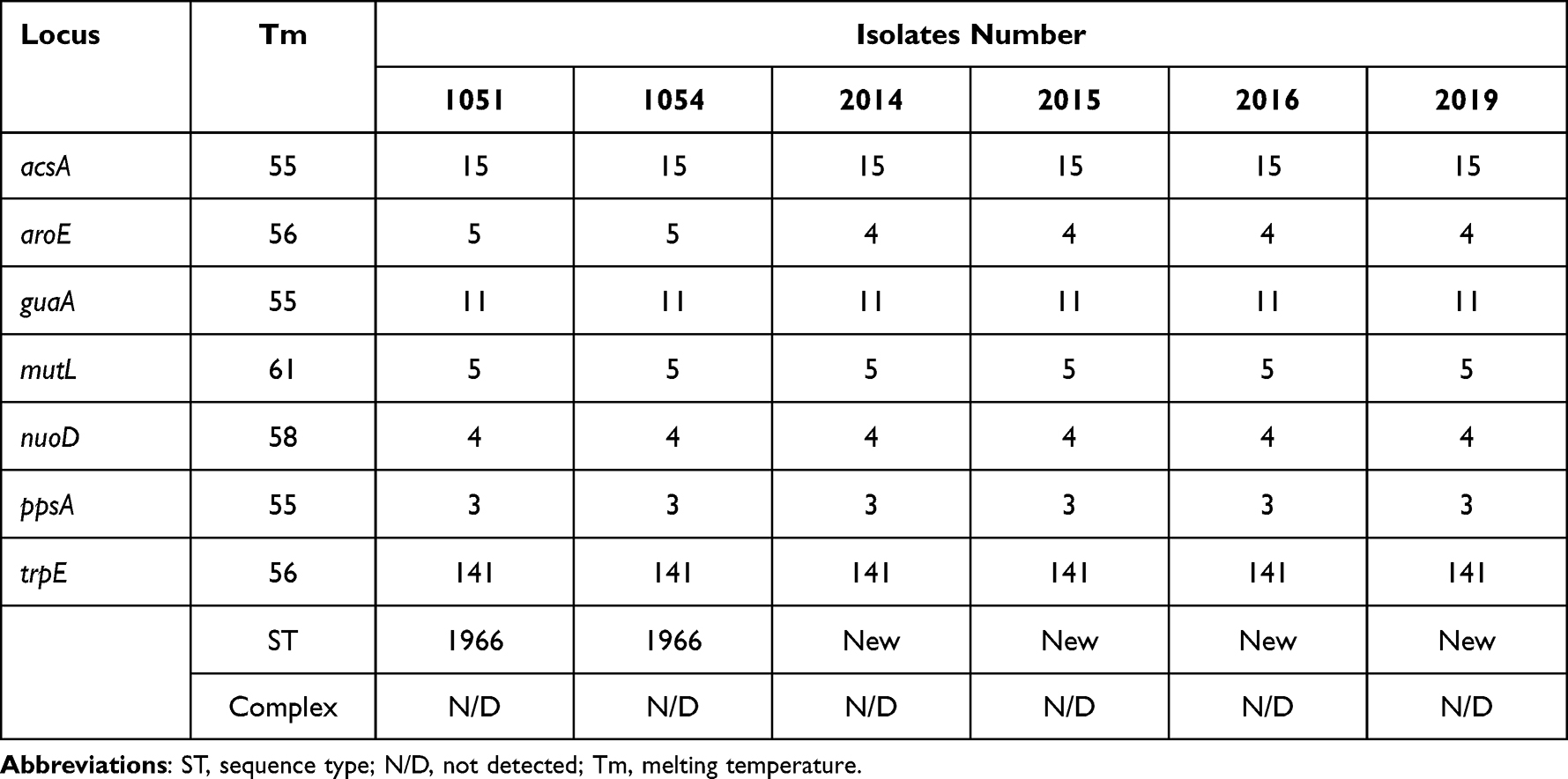 |
Table 2 Combination of Alleles with Sequences Type |
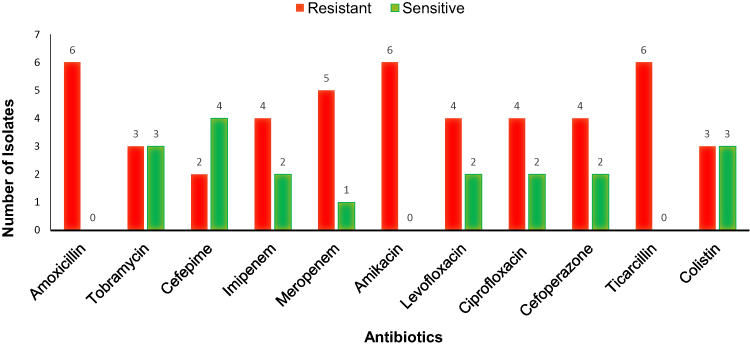 |
Figure 1 Antibiotic susceptibility testing of clinical isolates Pseudomonas aeruginosa. |
Discussion
Infections caused by P. aeruginosa are difficult to treat since this bacterium has several mechanisms of resistance and virulence, such as the acquisition of plasmids or integrons.21 The presence of colistin resistance plasmid-mediated mcr-1 in P. aeruginosa bacteria has drawn global attention because colistin last resort to treat infection caused by MDR pathogens.22 Therefore, the an urgent requirement for novel antibiotics with new modes of action or alternative approaches to identifying molecules that can interfere with the process of efflux such as P-glycoprotein.23 Our previous studies describe the co-production of MCR-1 and NDM-1 in Extended-Spectrum β-Lactamase-Producing Escherichia coli ST648 Isolated from a Colonized Patient in China.9 In the present study, the isolates were resistant to the commonly used antimicrobials to treat Gram-negative infections. Interestingly, we observed that three among six were resistant to colistin, which is the last choice of drug to treat infection caused by MDR Gram-negative bacteria.24 These antimicrobial susceptibility results are in agreement with other studies that demonstrated the high levels of resistance in P. aeruginosa recovered from bloodstream infections.25 We reported carbapenemase encoding blaNDM-1 from six strains which were the first detected in P. aeruginosa in Serbia in the year 2011.26 In the present study, three strains co-harbouring colistin resistance mcr-1 and carbapenemase encoding blaNDM-1, to the best of our knowledge; this has not yet been reported. Nevertheless, the co-existence of mcr-1 and blaCTX-M has already been reported earlier.27 In our study, we reported 2/6 P. aeruginosa that belong to ST1966, which is similar to a Chinese strain reported in 2011 from the Peking Hospital, Beijing (unpublished data). In addition, previous studies have reported that the novel sequence type strains are evolving, and increasing the mortality rate of P. aeruginosa bacteraemia,28,29 this highlights the need to further investigate this strain by the application of whole-genome sequencing. We have 3/4 mcr-1 conjugants while one failed, we suppose the location of a gene on the chromosome. The incidence of chromosomally encoded mcr-1 is rare although it has been reported in several cases.30 Here, we demonstrate that the mcr-1 gene is located on IncX4 plasmids, the plasmids that have been reported to play a significant role in the spread of the mcr-1 gene.31,32 Although there are limitations to this study, these findings help fill the knowledge gap that has hindered the mechanism of resistance and surveillance for this strain and can be used for further investigation. In addition, we need to consider the resistance mechanisms in MDR Pseudomonas as well as the related virulence factor and biofilm production and develop new correlations among these which may help for both diagnostic and therapeutic purposes.33
Conclusion
This is the first report to detect blaNDM-1 and colistin resistance mcr-1 in MDR P. aeruginosa isolates recovered from bloodstream infection in immunocompromised patients during leukaemia treatment. Colistin resistance is emerging in P. aeruginosa in a novel sequence type and may have a high-risk clone. The spread of carbapenemases in clinical isolates highlights the need for continuous surveillance for high-risk clones of P. aeruginosa, particularly in paediatric patients.
Abbreviations
NDM, New Delhi Metallo-beta-lactamase; AMR, Antimicrobial-resistant; EUCAST, European Committee on Antimicrobial Susceptibility Testing; PCR, Polymerase Chain Reaction; PBRT, PCR-based replicon typing; MLST, Multi-locus sequence typing (MLST) method and (PFGE); Inc, Incompatibility; STs, Sequence types..
Ethics Approval and Consent to Participate
The study was approved by the Shenzhen Children’s Hospital (Research) ethics committee (Reference number: 2018 (013).
Consent to Publish
The clinical isolate samples used in this research were part of the routine Hospital Laboratory procedure. Because we did not use the patient’s name or personal information, therefore no written informed consent was required. In addition, verbal consent was taken from the parents’ patients but not written.
Author Contributions
All authors made a significant contribution to the work including conceptualization, study design, execution, acquisition of data, analysis and interpretation, and were involved in drafting, revising and critically reviewing the manuscript. The final version of this manuscript has been approved by all authors and agreed to be accountable for all aspects of the work.
Funding
Shenzhen Fund for Guangdong Provincial High-Level Clinical Key Specialties (No. SZGSP012); and Shenzhen Key Medical Discipline Construction Fund (No. SZXK034).
Disclosure
The authors declare no competing conflicts of interest in this work.
References
1. Winfried K, Jan R, Hartmut B, et al.; Hospital Infection Surveillance System for Patients With Hematologic/Oncologic Malignancies Study Group (ONKO-KISS). Contribution of specific pathogens to bloodstream infection mortality in neutropenic patients with hematologic malignancies: results from a multicentric surveillance cohort study. Transpl Infect Dis. 2019;21:e13186. doi:10.1111/tid.13186
2. Reynolds D, Kollef M. The epidemiology and pathogenesis and treatment of pseudomonas aeruginosa infections: an update. Drugs. 2021;81(18):2117–2131. doi:10.1007/s40265-021-01635-6
3. Tofas P, Samarkos M, Piperaki ET, et al. Pseudomonas aeruginosa bacteraemia in patients with hematologic malignancies: risk factors, treatment and outcome. Diagn Microbiol Infect Dis. 2017;88(4):335–341. doi:10.1016/j.diagmicrobio.2017.05.003
4. Available from: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed.
5. Bottery MJ, Pitchford JW, Friman VP. Ecology and evolution of antimicrobial resistance in bacterial communities. ISME J. 2021;15(4):939–948. doi:10.1038/s41396-020-00832-7
6. Aloush V, Navon-Venezia S, Seigman-Igra Y, Cabili S, Carmeli Y. Multidrug-resistant Pseudomonas aeruginosa: risk factors and clinical impact. Antimicrob Agents Chemother. 2006;50(1):43–48. doi:10.1128/AAC.50.1.43-48.2006
7. Horcajada JP, Montero M, Oliver A, et al. Epidemiology and treatment of multidrug-resistant and extensively drug-resistant pseudomonas aeruginosa infections. Clin Microbiol Rev. 2019;32(4):e00031–e00019. doi:10.1128/CMR.00031-19
8. Mekonnen H, Seid A, Molla Fenta G, Gebrecherkos T. Antimicrobial resistance profiles and associated factors of Acinetobacter and Pseudomonas aeruginosa nosocomial infection among patients admitted at Dessie comprehensive specialized Hospital, North-East Ethiopia. A cross-sectional study. PLoS One. 2021;16(11):e0257272. doi:10.1371/journal.pone.0257272
9. Sandip P, Jiang M, Wen F. Molecular characterization of co-existence of MCR-1 and NDM-1 in extended-spectrum β-lactamase-producing Escherichia coli ST648 isolated from a colonized patient in China. Jundishapur J Microbiol. 2019;12(7):e91272.
10. Patil S, Liu X, Chen H, Francisco NM, Wen F, Chen Y. Genetic characterization of colistin-resistant Salmonella enterica ST34 Co-harbouring plasmid-Borne mcr-1, blaCTX-M-15 and blaKPC-2 recovered from a paediatric patient in Shenzhen, China. Infect Drug Resist. 2022;15:757–763. doi:10.2147/IDR.S349585
11. World Health Organization. Global antimicrobial resistance and use surveillance system (GLASS): whole-genome sequencing for surveillance of antimicrobial resistance. Geneva: World Health Organization; 2020.
12. World Health Organization. Global antimicrobial resistance surveillance system (GLASS) report. Geneva: World Health Organization; 2019.
13. Hendriksen RS, Bortolaia V, Tate H, Tyson GH, Aarestrup FM, McDermott PF. Using genomics to track global antimicrobial resistance. Front Public Health. 2019;7:242. doi:10.3389/fpubh.2019.00242
14. Picão R, Andrade S, Nicoletti A, et al. Metallo-beta-lactamase detection: comparative evaluation of double-disk synergy versus combined disk tests for IMP-, GIM-, SIM-, SPM-, or VIM-producing isolates. J Clin Microbiol. 2008;46:2028–2037. doi:10.1128/JCM.00818-07
15. Patel JB, Cockerill FR, Bradford PA. M100-S25 performance standards for antimicrobial susceptibility testing; Twenty-fifth informational supplement; 2015.
16. Matuschek E, Åhman J, Webster C, Kahlmeter G. Antimicrobial susceptibility testing of colistin - evaluation of seven commercial MIC products against standard broth microdilution for Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter spp. Clin Microbiol Infect. 2018;24(8):865–870. doi:10.1016/j.cmi.2017.11.020
17. Patil S, Chen H, Zhang X, Lian M, Ren PG, Wen F. Antimicrobial resistance and resistance determinant insights into multi-drug resistant gram-negative bacteria isolates from paediatric patients in China. Infect Drug Resist. 2019;12:3625–3634. doi:10.2147/IDR.S223736
18. Zhuoren L, Wenjuan Y, Zhangqi S, Yang W, Jianzhong S, Timothy W. Epidemiology of mobile colistin resistance genes mcr-1 to mcr-9. J Antimicrob Chemother. 2020;75(11):3087–3095. doi:10.1093/jac/dkaa205
19. Carattoli A, Bertini A, Villa L, Falbo V, Hopkins KL, Threlfall EJ. Identification of plasmids by PCR-based replicon typing. J Microbiol Methods. 2005;63(3):219–228. doi:10.1016/j.mimet.2005.03.018
20. Alvarado A, Garcillán-Barcia MP, de la Cruz F. A degenerate primer MOB typing (DPMT) method to classify gamma-proteobacterial plasmids in clinical and environmental settings. PLoS One. 2012;7(7):e40438. doi:10.1371/journal.pone.0040438
21. Raúl R, Jennifer V, Esther V, María Á, Jaime L, Fernando C. Bacteraemia due to extensively drug-resistant Pseudomonas aeruginosa sequence type 235 high-risk clone: facing the perfect storm. Int J Antimicrob Agents. 2018;52(2):172–179. doi:10.1016/j.ijantimicag.2018.03.018
22. Caselli E, D’Accolti M, Soffritti I, Piffanelli M, Mazzacane S. Spread of mcr-1-driven colistin resistance on hospital surfaces, Italy. Emerg Infect Dis. 2018;24(9):1752–1753. doi:10.3201/eid2409.171386
23. Usai D, Donadu M, Bua A, et al. Enhancement of antimicrobial activity of pump inhibitors associating drugs. J Infect Dev Ctries. 2019;13(2):162–164. doi:10.3855/jidc.11102
24. Zheng P, Renee R, Bernard R, Tong-Jun L, Zhenyu C. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnol Adv. 2019;37(1):177–192. doi:10.1016/j.biotechadv.2018.11.013
25. Hamed T, Sanaz D, Mohammad Reza A. Co-harboring of mcr-1 and β-lactamase genes in Pseudomonas aeruginosa by high-resolution melting curve analysis (HRMA): molecular typing of superbug strains in bloodstream infections (BSI). Infect Genet Evol. 2020;85:104518. doi:10.1016/j.meegid.2020.104518
26. Jovcic B, Zorica L, Vesna S, et al. Emergence of NDM-1 metallo-beta-lactamase in Pseudomonas aeruginosa clinical isolates from Serbia. Antimicrob Agents Chemother. 2011;55(8):3929–3931. doi:10.1128/AAC.00226-11
27. Cecilia Q, Marcela N, JoséDi C. Current scenario of plasmid-mediated colistin resistance in Latin America. Rev Argent Microbio. 2019;51(1):93–100.
28. Lidia R, Alba B, Jessica B, et al. Pseudomonas aeruginosa Isolates from Spanish Children: occurrence in Faecal Samples, Antimicrobial Resistance, Virulence, and Molecular Typing. Biomed Res Int. 2018;2018:1–8.
29. Schwab F, Geffers C, Behnke M, Petra J. ICU mortality following ICU-acquired primary bloodstream infections according to the type of pathogen: a prospective cohort study in 937 Germany ICUs (2006–2015). PLoS One. 2018;13(3):1–13. doi:10.1371/journal.pone.0194210
30. Liu Z, Liu Y, Xi W, et al. Genetic features of plasmid- and chromosome-mediated mcr-1 in Escherichia coli isolates from animal organs with lesions. Front Microbiol. 2021;12:707332. doi:10.3389/fmicb.2021.707332
31. Michel D, Gauri G, Philip A, et al. Detection of the plasmid-mediated mcr-1 gene conferring colistin resistance in human and food isolates of Salmonella enterica and Escherichia coli in England and Wales. J Antimicrob Chemother. 2016;71(8):2300–2305. doi:10.1093/jac/dkw093
32. Michael M, Laura M, James R, et al. Dissemination of the mcr-1 colistin resistance gene. Lancet Infect Dis. 2016;16(3):289–290. doi:10.1016/S1473-3099(16)00067-0
33. Gajdács M, Baráth Z, Kárpáti K, et al. No correlation between biofilm formation, virulence factors, and antibiotic resistance in Pseudomonas aeruginosa: results from a laboratory-based in vitro study. Antibiotics. 2021;10(9):1134. doi:10.3390/antibiotics
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.