")
Back to Journals » Cancer Management and Research » Volume 12
New Advances in Canonical Wnt/β-Catenin Signaling in Cancer
Authors Wen X, Wu Y , Awadasseid A, Tanaka Y , Zhang W
Received 21 April 2020
Accepted for publication 19 July 2020
Published 6 August 2020 Volume 2020:12 Pages 6987—6998
DOI https://doi.org/10.2147/CMAR.S258645
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Ahmet Emre Eşkazan
Xiaolan Wen,1,2 Yanling Wu,1,2 Annoor Awadasseid,1,3 Yoshimasa Tanaka,4 Wen Zhang1
1Laboratory of Chemical Biology and Molecular Drug Design, College of Pharmaceutical Science, Zhejiang University of Technology, Hangzhou 310014, People’s Republic of China; 2Laboratory of Molecular Immunology, Zhejiang Provincial Center for Disease Control and Prevention, Hangzhou 310051, People’s Republic of China; 3Center for Natural Products Research, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu, 610041, People’s Republic of China; 4Laboratory of Bioinformatics and Molecular Medicine, Center for Medical Innovation, Nagasaki University, Nagasaki, 852-8588, Japan
Correspondence: Yanling Wu
Laboratory of Molecular Immunology, Zhejiang Provincial Center for Disease Control and Prevention, 3399 Binsheng Road, Binjiang District, Hangzhou, Zhejiang 310051, People’s Republic of China
Tel +86 138 6715 2930
Email [email protected]
Wen Zhang
Laboratory of Chemical Biology and Molecular Drug Design, College of Pharmaceutical Science, Zhejiang University of Technology, 18 Chaowang Road, Xiacheng District, Hangzhou, Zhejiang 310014, People’s Republic of China
Tel +86 136 1671 0236
Email [email protected]
Abstract: Wnt/β-catenin-mediated signaling is a key pathway regulating tissue growth and development, and tumorigenesis, and has received increasing attention in recent years. In addition to participating in healthy tissue and organ development, ectopic activation of the pathway can cause a variety of tumors and other pathologies. The pathway plays a critical role in many processes such as proliferation, differentiation, apoptosis, migration, invasion, epithelial–mesenchymal transition and cancer cell stemness. The importance of the Wnt signal is self-evident. This review describes the underlying mechanism of Wnt signaling pathway and highlights the latest findings on the relationship between Wnt signaling pathway and tumorigenesis. In addition, the potential relationship between miRNAs and Wnt signaling is presented. Furthermore, we discuss the intrinsic link between Wnt signaling and cancer cell stemness, which shed light on the malignant progression of tumor cells. Finally, cancer treatment strategies based on the canonical Wnt signaling pathway are summarized, hoping to help clinical development.
Keywords: Wnt/β-catenin, tumor, miRNAs, cancer stem cell, target, therapeutic strategy
Introduction
Wnt signaling pathway plays a crucial role in embryonic development, adult tissue homeostasis, and cancer.1 Due to the role in cell fate and tissue development, increasing attention has been paid to Wnt signaling pathway in regenerative medicine.2 Moreover, the pathway is also involved in many pathological processes such as proliferation, differentiation, apoptosis, migration, invasion, epithelial-mesenchymal transition (EMT), and cancer cell stemness. Notably, the aberrant activation of Wnt/β-catenin signaling results in the expression of several modulators that antagonize the anti-tumor activity of T cells, which might lead to the failure of cancer immunotherapy.3 Since Wnt signaling plays a pivotal role in tumorigenesis, it is a promising target for the development of cancer therapeutics.
Since the discovery about 40 years ago, many reports on the relationship between Wnt signaling and malignant tumor progression have been published. The Wnt family consists of 19 cysteine-rich protein species, which deliver canonical Wnt signaling through the interaction with Frizzled (FZD) and co-receptor low-density lipoprotein-related receptors 5 and 6 (LRP5/6), thereby regulating cell fate, growth and tissue repair.4 After a series of posttranslational modifications, such as porcupine palmitoylation, lipid modification and glycosylation, in the endoplasmic reticulum, the Wnt precursor gradually maturates into the functional Wnt ligand. Then, the transmembrane protein Wntless (Wls) binds Wnt ligand, and reaches to the plasma membrane via the Golgi apparatus. Finally, Wnt is detached from the cell membrane as components of exosomes or lipid protein particles.1,5 As shown in Figure 1, Wnt ligand-binding membrane receptor complex induces constitutive activation of Wnt/β-catenin signaling, which delivers downstream signaling, LRP receptor phosphorylation recruits Axin to degrade; Dishevelled (DVL) protein activation leads to inactivation of destruction complex, which leads to the stability and accumulation of β-catenin.6 Subsequently, β-catenin is translocated into nucleus and interacts with the transcription coactivators like T-cell factor (TCF) and lymphoid enhancer factor (LEF) proteins to regulate the expression of Wnt target genes, such as AXIN2, cycling, and c-MYC.7 When Wnt signal is turned off, the scaffold protein Axin, adenomatous polyposis coli (APC), glycogen synthase kinase 3β (GSK3β) and casein kinase 1α (CK1α) form a multimeric destruction complex, which induces β-catenin phosphorylation and ubiquitination degradation.8
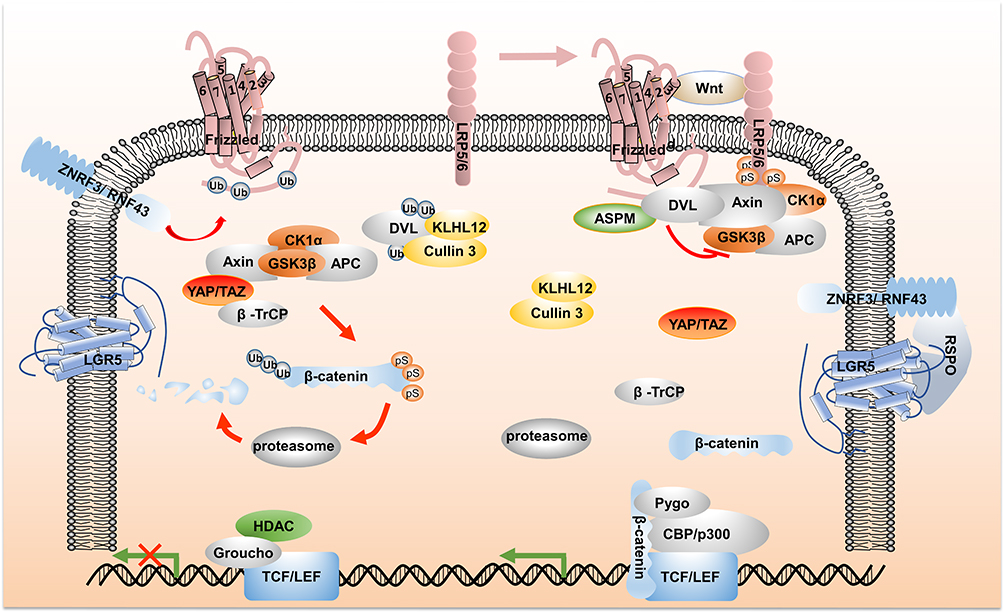 |
Figure 1 An overview of Wnt signaling pathway. As illustrated on the upper left side of Figure 1, in the absence of Wnt ligand, destruction complex induces the phosphorylation of β-catenin, and the phosphorylated β-catenin is recognized and ubiquitinated by YAP/TAZ-recruited β-TrCP, and subsequently subjected to proteasomal degradation. In addition, the E3 ubiquitin ligases Cullin 3/KLHL12 recognize DVL and induce its degradation. On the membrane, ZNRF3/RNF43 targets Frizzled receptor for ubiquitination and lysosomal degradation. In the nucleus, the inhibitory complex Groucho and histone deacetylases (HDAC) bind to TCF/LEF transcription factors, resulting in the inhibition of the Wnt signaling gene expression, as shown in the lower left side of Figure 1. On the upper right side of Figure 1, the phosphorylated LRP receptor recruits Axin and DVL protein to the plasma membrane. Activated DVL inactivates the destruction complex. In this process, the combination of DVL and ASPM inhibits the recognition of Cullin 3/KLHL12. β-catenin, with the assistance of histone modifying co-activators such as Pygopus (Pygo) and CREB binding protein (CBP)/p300, interacts with the transcription factors TCF/LEF to regulate target gene expression. Frizzled ubiquitination is inhibited through the interaction of RSPO with LGR4-6 and ZNRF3/RNF43. “Ub” stands for ubiquitination and “ps” stands for phosphorylation. Abbreviations: LRP5/6, low-density lipoprotein-related receptors 5 and 6; DVL, dishevelled; TCF, T-cell factor; LEF, lymphoid enhancer factor; APC, the adenomatous polyposis coli; GSK3β, the glycogen synthase kinase 3β; CK1α, casein kinase 1α; β-TrCP, β transducin repeat-containing protein; RSPO, R-spondin; LGR5, leucine-rich repeat-containing G-protein-coupled receptors 5; ASPM, abnormal spindle-like microcephaly associated; HDAC, histone deacetylases; Pygo, Pygopus; CBP, CREB binding protein. |
Other mechanisms have been identified downstream of the canonical Wnt signaling pathway, including the Hippo signaling cascade downstream nuclear effectors YAP/TAZ. Under Wnt OFF conditions, YAP/TAZ is an essential component of the destruction complex. It recruits E3 ubiquitin ligase β-transducin repeat-containing protein (β-TrCP) to promote β-catenin degradation, in which, YAP/TAZ is a negative regulator of Wnt signaling. However, when Wnt is ON, LRP5/6 induces the dissociation of YAP/TAZ from Axin, resulting in nuclear accumulation of YAP/TAZ and enhancement of the expression of pro-proliferative genes, in which YAP/TAZ is a positive regulator of Wnt signaling.9 Furthermore, members of the R-spondin (RSPO) ligand family were identified as positive effectors of Wnt signaling.10 In the absence of RSPO, the two homologues E3 ubiquitin ligases ZNRF3/RNF43 ubiquitinated Frizzled, inducing Fzd endocytosis. When RSPO forms complex with leucine-rich repeat-containing G-protein-coupled receptors (LGR) 4–6 and ZNRF3/RNF43, however, the Frizzled ubiquitination process is inhibited, thereby inducing the activation of the Wnt signaling cascade. Recently, the abnormal spindle-like microcephaly associated (ASPM)/Dishevelled signal axis was proposed to participate in the generation of canonical Wnt signaling. ASPM interacts with Wnt regulator DVL, blocking the recognition for DVL by the ubiquitin ligase complex Cullin 3/KLHL12, inhibiting the ubiquitination and subsequent degradation of DVL, and maintaining a high level of Wnt activity.11
Besides the canonical pathway, the Wnt signal also has an additional non-canonical activation pathway, which is independent of β-catenin. The non-canonical Wnt signaling pathway targets different receptors and activates multiple intracellular targets, among which the most widely studied are Wnt/planar cell polarity (PCP) and Wnt/Calcium (Ca2+) pathways. As shown in Figure 2, in the Wnt/PCP pathway, Wnt ligand binds to receptor proteins to recruit and activate DVL, which subsequently activates the small GTPases Rho or Rac, triggering downstream Rho-associated kinase (ROCK) and c-Jun N-terminal kinase (JNK) recruitment, allowing cytoskeleton reorganization.12 In the Wnt/Ca2+ pathway, Fzd receptor is activated, which induces the activation of DVL. Then, DVL recruits phospholipase C (PLC), which converts phosphatidylinositol 4,5-bisphosphate (PIP2) to diacylglycerol (DAG) and inositol 1,4,5-triphosphate (IP3). IP3 stimulates intracellular Ca2+ release and DAG and Ca2+ together activate downstream protein kinase C (PKC), calcineurin (CNA) and Ca2+/calmodulin-dependent protein kinase II (CaMKII), thereby regulating intracellular calcium fluxes and downstream calcium-dependent cytoskeletal and/or transcriptional responses.13,14 Although non-canonical Wnt signaling is also an indispensable factor in the development of cancer, this review mainly focuses on canonical Wnt signaling.
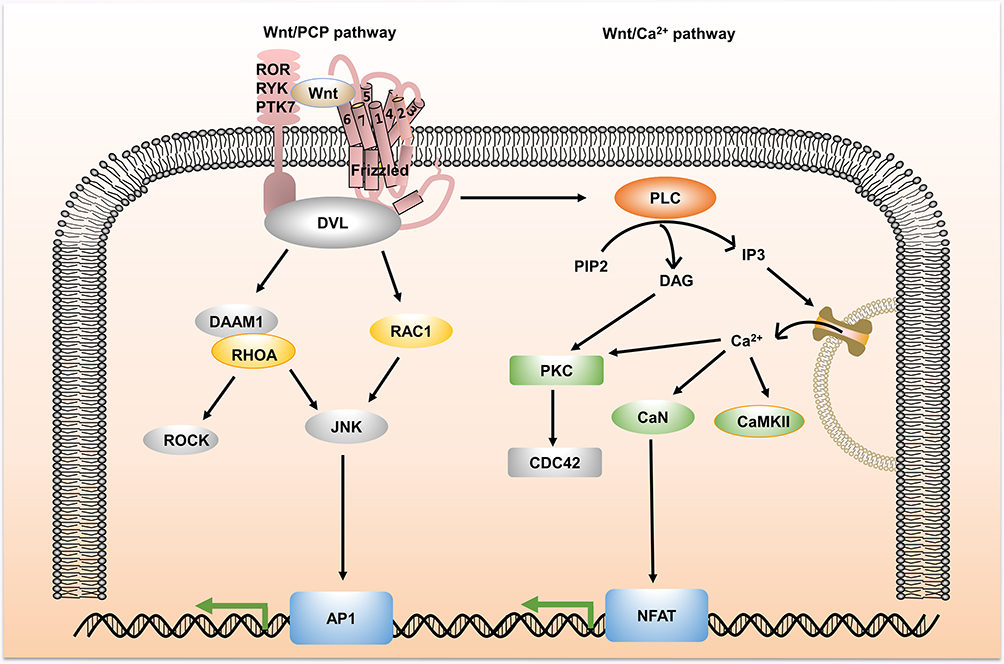 |
Figure 2 An overview of non-canonical Wnt signaling pathway. Non-canonical Wnt signaling is divided into Wnt/PCP and Wnt/Ca2+ pathways. In Wnt/PCP, Wnt ligand-binding receptor proteins trigger the activation of the ROCK and JNK downstream kinases, allowing cytoskeletal reorganization. In Wnt/Ca2+, non-canonical Wnt signaling increases the intracellular levels of DAG and IP3 by recruiting DVL, stimulates the intracellular Ca2+ release and activates downstream PKC, CaN and CaMKII, thereby regulating intracellular calcium fluxes and downstream calcium-dependent cytoskeletal and/or transcriptional responses. Abbreviations: ROCK, Rho-associated kinase; JNK, c-Jun N-terminal kinase; PLC, phospholipase C; PIP2, phosphatidylinositol 4,5-bisphosphate; DAG, diacylglycerol; IP3, inositol 1,4,5-triphosphate; PKC, protein kinase C; CAN, calcineurin; CaMKII, Ca2+/calmodulin-dependent protein kinase II; DAAM1, the formin protein dishevelled-associated activator of morphogenesis 1; RHOA, Ras homolog family member A; RAC1, Ras-related C3 botulinum toxin substrate 1; AP1, activator protein-1; CDC42, Cell division cycle protein 42; NFAT, nuclear factor of activated T cells. |
There is growing evidence that Wnt/β-catenin signaling is one of the main driving factors of some malignant tumors such as colorectal cancer, liver cancer and breast cancer. This review, therefore, focuses on the latest research progress on the correlation between the Wnt signal and tumors, and the potential relationship between the Wnt signal and miRNAs. In addition, the possible relationship between the Wnt signal and CSC is discussed. Finally, cancer treatment strategies based on the canonical Wnt signal are summarized.
Wnt Signaling in Colorectal Cancer
Colorectal cancer (CRC) is the third most common cancer in the world and the fourth most deadly cancer after lung, liver, and stomach cancer, with the highest incidence in developed countries. Constitutive activation of Wnt/β-catenin signaling is a functional marker of colorectal cancer, in which APC mutations occur in approximately 80% of CRCs. APC-based therapy may be, therefore, a promising strategy for the treatment of colorectal cancer.15 It was reported that APC silencing drives hyperproliferation in the intestine, and eventually forms colon adenomas. Restoring APC gene led to continuous regression without recurrence, whereas reestablished crypt homeostasis was found in tumor tissues carrying Kras and p53 mutations.8 Wnt signaling is thus a promising target for the development of cancer therapy. It was recently found that metastasis-associated in colon cancer 1 (MACC1) served as a transcriptional target of Wnt/β-catenin signaling. The DBC1 (Deleted in breast cancer-1) coactivator promotes self-renewal capacity and drug resistance in colon cancer by regulating LEF1/β-catenin-dependent enhancer in the MACC1 intron.16 Furthermore, Deptor (The DEP domain containing mTOR interacting), an mTOR inhibitor, promotes cancer cell proliferation and survival in CRC and is a potential target for novel cancer therapies. Deptor is a direct target gene for Wnt/β-catenin/c-Myc signaling.17 Silencing Deptor induces differentiation and inhibition of CRC cells by increasing ketone production and decreasing the expression of B lymphoma Mo-MLV insertion region 1 (Bmi1), while increasing mTOR activation. A combination of Akt/mTOR and Wnt/β-catenin inhibitors thus can exert a potent anti-tumor effect. In addition to inhibition of positive regulators for Wnt signaling, a negative regulator was also found as a possible target for Wnt signaling.18 V-set and transmembrane domain containing 2A (VSTM2A) is a secreted protein that inhibits Wnt/β-catenin signaling in colon cancer by directly inhibiting LRP6 activity and inducing LRP6 endocytosis and degradation.
Cancer metastasis is the main cause of death in CRC patients. Tocopherol alpha transfer protein-like (TTPAL) and chromatin organizer special AT-rich binding protein 1 (SATB1) are possibly involved in the invasion and metastasis of CRC. Gou et al found19 that the upregulation of TTPAL is associated with a poor prognosis of colon cancer, in which TTPAL directly interacts with thyroid receptor-interacting protein 6 (TRIP6) and inhibits the ubiquitination and degradation of TRIP6. TRIP6 competitively binds to the membrane-associated guanylate kinase with inverted domain structure-1 (MAGI1) tumor suppressor and dissociates β-catenin from MAGI1. Free β-catenin enters the nucleus to activate the oncogenic Wnt/β-catenin signal, increasing the ability of cancer cells to migrate and invade. In addition, over-activation of Wnt signaling promotes the interaction between TCF7L2/β-catenin complex and SATB1, leading to SATB1 expression.20 SATB1 regulates the expression of tumor growth- and metastasis-associated genes, regulating the interconversion of colon cancer in the invasion phenotype.
Wnt/β-catenin signaling affects the malignant progression of colon cancer by regulating tumor suppressor/activator. It is, therefore, pivotal to further explore Wnt signal-related targets, with a view to revealing a network of Wnt signaling in cancer and providing more options for cancer treatments. The roles of Wnt signaling in the development of colon cancer were summarized in this section, hoping to understand Wnt signaling from a new perspective.
Wnt Signaling in Liver Cancer
According to the survey by World Health Organization’s International Cancer Research Center in 2018, liver cancer is one of the world’s high-mortal cancers, and hepatocellular carcinoma (HCC) is the second leading cause of cancer death worldwide. β-catenin is expressed in the adult liver, which mediates Wnt signaling and regulates the transcription and translation of genes around liver cells, affecting various aspects of liver biology. Here, we mainly describe the roles of Wnt signaling in hepatic metabolic partitioning and liver regeneration. RSPO-LGR4/5-ZNRF3/RNF43 module is the main signaling complex in the Wnt/β-catenin signaling pathway to regulate liver metabolism, development and regeneration.21 Knocking down LGR4/5 in the mouse liver resulted in a loss of β-catenin signaling and a defect in hepatic metabolic partitioning. Supplementing RSPO1 or knocking down Znrf3/Rnf43 extends Wnt/β-catenin signal gradient in the liver in a reversible and LGR4/5-dependent manner, resuming the liver defect phenotype. In addition, liver regeneration is possibly regulated by type I transmembrane protein 9 (TMEM9). In the model of liver injury induced by CCl4, TMEM9 enhances the rapid assembly of the v-ATPase lysosomal proton pump, activates cathepsin activity in lysosomes, induces APC degradation, and activates Wnt/β-catenin pathway, leading to the promotion of liver regeneration.22 Wnt/β-catenin signaling is, therefore, essential for liver development, whereas the abnormal activation of the β-catenin signal is also a sign of promoting the development of hepatocellular carcinoma and other liver diseases. As shown in Figure 3, approximately 50% of HCC exhibited Wnt/β-catenin signaling activity, among them, about 30% of hepatocellular carcinomas had Catenin β1 (CTNNB1) mutations,23 which highlighted the importance of CTNNB1 in the progression of liver cancer. Together with the activation levels of other signals in HCC, we might hypothesize that the crosstalk between Wnt signaling and multiple signaling pathways promotes the development of liver cancer.24 In addition, in a mouse hepatocyte model, β-catenin activation per se is not sufficient to induce liver cancer, and other mutation events must be involved in the development of liver cancer, such as mutations in a telomerase reverse transcriptase gene (TERT)25 or those in the gene mesenchymal epithelial transition factor (MET).26 This phenomenon may be related to the weak β-catenin activity in the mouse hepatocyte model. We found that there are weak mutations in the CTNNB1 in HCC and HCA (hepatocellular adenoma), for example, the mutation takes place at K335/N387, S45, or T41 of the CTNNB1.27
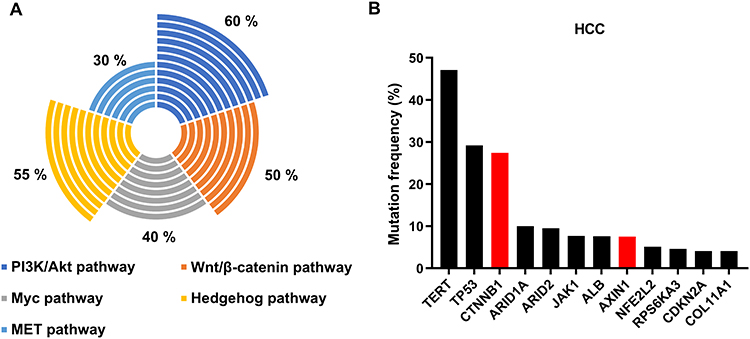 |
Figure 3 Status of Wnt/β-catenin signal activation in HCC. (A) Activation ratio of Wnt/β-catenin signaling in HCC. (B) Mutation frequency of common high-mutation genes in HCC. |
Wnt signaling plays a crucial role in the growth and development of liver. However, its aberrant activation may promote the occurrence of hepatocellular carcinoma and other liver lesions. It is noteworthy that a liver-specific Wnt/β-catenin signaling pathway exists in liver cells. Glypican-3 (GPC3) is a heparan sulfate proteoglycan specifically expressed on the surface of liver cancer cells. Li et al reported28 that GPC3 regulates the activation of Wnt signaling by dual mechanisms and promotes proliferation of hepatocellular carcinoma. When FZD is not abundant, GPC3 acts as an alternative Wnt receptor, and recruits Wnt at the cell surface through the N-terminal CRD domain, leading to the promotion of Wnt signaling. When FZD is abundant, GPC3 and FZD interact through their heparin sulfate (HS) chains, and the two molecules form a triple complex with Wnt to synergistically stimulate Wnt signaling. GPC3 might function as a bridge to stabilize the interaction between Wnt and FZD. Further studies promote the elucidation of the roles of Wnt signaling in the development of liver cancer and provide a theoretical basis for GPC3 targeted therapy.
Wnt Signaling in Breast Cancer
Wnt signaling is involved in the malignant progression of breast cancer, such as proliferation, invasion, metastasis, and drug resistance. In healthy breast tissue, inhibition of Wnt/β-catenin activity leads to developmental disorders and reduced cell proliferation during pregnancy.29 However, there is increasing evidence that aberrant activation of the Wnt signal promotes the development of breast cancer. It was demonstrated30 that bone marrow tyrosine kinase on chromosome X (BMX) is up-regulated in breast cancer. It inhibits the degradation of β-catenin by promoting GSK3β phosphorylation, thereby disrupting normal Wnt signaling and promoting the malignant progression of breast cancer. Rational regulation of Wnt/β-catenin signaling is, therefore, a key to the treatment of breast cancer, although it is difficult to specifically control the signaling. In theory, inhibition of Wnt/β-catenin signaling can prevent tumor progression. Surprisingly, it is likely that silencing canonical Wnt signaling drives cancer cells into dormancy, resulting in the hindrance of tumor cells from cancer immunoediting by the host immune system.31 It was reported that Dickkopf-1 (DKK1) inhibits lung metastasis by antagonizing non-canonical Wnt signal of breast cancer. DKK1, however, could promote breast-to-bone metastasis by regulating canonical Wnt signal in osteoblasts. Taken together, it is not straightforward to manipulate canonical Wnt signaling for the treatment of metastatic cancer.
Wnt Signaling and Micro-RNAs
In recent years, several studies have revealed the potential connection between non-coding RNA and Wnt signaling. Table 1 summarizes the effect of miRNAs on Wnt/β-catenin signaling in cancer.
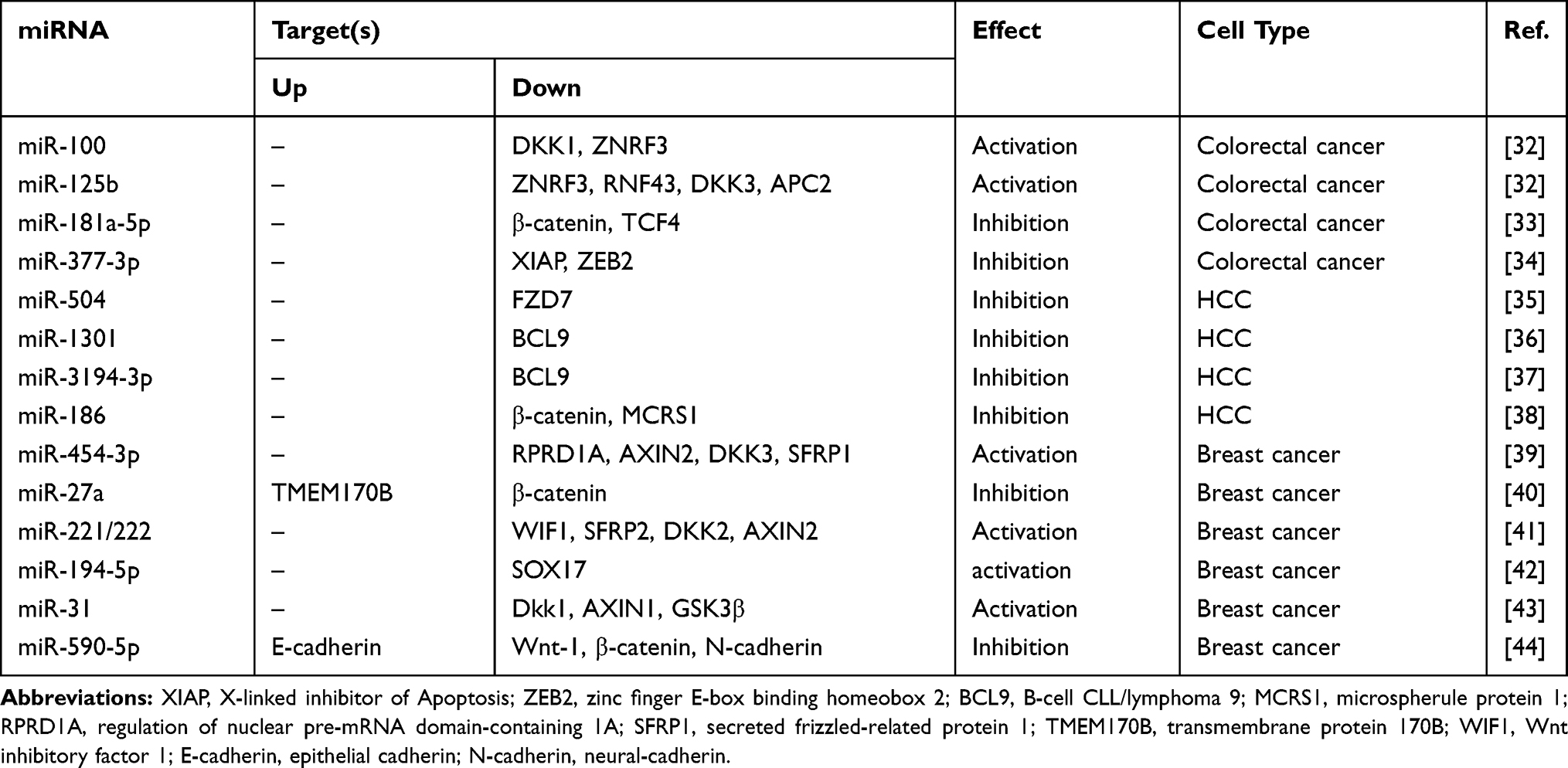 |
Table 1 Effect of miRNAs on Wnt/β-Catenin Signaling in Cancer |
It has been reported that miRNA expression is also controlled by Wnt signaling. In HCC, CTNNB1 binds to the miR-183/96/182 promoter region and increases the transcription of the miRNAs, thereby enhancing tumor cell invasion.45 It is noteworthy that there is a mutual feedback loop between some miRNAs and Wnt/β-catenin signaling. In colon cancer, miR-452 induces Wnt signaling by targeting GSK3β. Since T cell factor/lymphatic enhancement factor is an effective transcription factor in the miR-452 promoter, in turn, miR-452 transcription is promoted by Wnt signaling, forming a positive feedback loop.46 In addition, a negative feedback loop was found between the miRNA and Wnt signaling pathway. miR-145, a tumor suppressor miRNA, can interact with TCF4-β-catenin complex, recruit polycomb repressive complex 2 (PRC2) histone trimethylase, and suppress Wnt signal transduction. Recruitment of PRC2, however, results in a high level of methylation at the miR-145 promoter region, which down-regulates the expression of miR-145.47 It was further demonstrated that abnormally down-regulated miR-145 was inversely correlated with its inhibitory regulators (TCF4 and SUZ12 (Suppressor of Zeste 12 Protein Homolog)), indicating that miR-145 forms a double negative regulatory loop with the negative regulator of colorectal cancer. It is, therefore, imperative to further analyze the network and regulation of miRNA and Wnt/β-catenin signaling pathway, which would provide insights into the development of miRNA-based drugs.
Wnt Signaling in Cancer Stemness
It is hypothesized that malignant tumors grow in a layered manner and cancer stem cells (CSCs) having self-renewal characteristics and multi-directional differentiation ability play important roles in the tumor development and progression. CSCs continuously maintain the replacement of new CSC tumorigenic subpopulations and differentiate into non-CSC progenies with high proliferative ability.48 It is assumed that cancer originates in part from CSCs. de Sousa e Melo et al demonstrated that, in colorectal cancer, Lgr5-GFP+ tumor cells had higher tumorigenicity than Lgr5-GFP− tumor cells, showing that Lgr5+ cells had tumor-initiating ability.49 In addition, depletion of Lgr5+ cells triggers differentiated cancer cells to continuously reverse to cells in a tumor-propagating state and replenish the Lgr5+ CSC pool. In another study, a similar conclusion was provided. Mature leukemia cells could reacquire clonogenic and leukemogenic properties through de-differentiation. It is worthy of note that CSC plasticity contributes to the interconversion of leukemia status through interfering with endogenous PU.1, an ETS family pioneer transcription factor required for myelopoiesis.50 However, CSCs that transformed from differentiated cells require additional factors (mutations, inflammation, or changes in the microenvironment) for their stemness features.51 For example, after hepatotoxin induction, hepatocytes produce tumor nodules and express progenitor cell markers.52 Targeting CSCs and preventing the generation of CSCs from non-CSCs are thus a key for the development of cancer therapy.
There is a wealth of evidence that CSCs are closely related to tumor invasion, metastasis and drug resistance. The differentiation gradient between CSCs and their non-CSC progenies results in tumor heterogeneity, which indicates that cells of different genotypes can exist in the same tumor, including subgroups with transferability. Owing to the unique nature of tumors, CSCs have become the leading cause of drug resistance in cancer treatment. Wnt/β-catenin signaling plays an essential role in the reprogramming and cancer cell stemness and is an indispensable cytokine network for promoting the progression of CSCs. Reilein et al found that the cell types of CSC-derivatives depend in part on the magnitude of spatially graded Wnt pathway activity.53 The latest report54 proves the remarkable heterogeneity in Wnt activity in HCC cells, among which Wnt-activityhighALDH1+EpCAM+ triple-positive cells are the most stem-like and highly tumorigenic cells in all CSC populations. This type of cells is called “superpotent CSCs” (spCSC). The ASPM/Dishevelled signal axis is highly activated in spCSCs and is heterogeneous in HCC tissues. Future studies would reveal the underlying mechanism of Wnt and stem cell heterogeneity in liver cancer, and provide new insights into CSC-based tumor treatment strategy.
Wnt signaling contributes to reprogramming and maintenance of the CSC status, such as the epithelial–mesenchymal transition. Chang et al found that in epithelial cells, the SRY-related high-mobility-group box gene 15 (Sox15) tumor suppressor interacted with the β-catenin/E-cadherin complex and then interacted with the proximal promoter region of caspase-3 (CASP3). Caspase-3 inactivates the Twist family bHLH transcription factor (Twist), which inhibits Wnt signaling for CSC phenotype acquisition. In interstitial cells, Twist1 forms a complex with β-catenin/TCF4 and binds to the proximal promoter region of ATP-binding cassette G2 (ABCG2), which induces the expression of stem cell-related genes and promotes Wnt signaling to induce the CSC phenotype. In the presence of Wnt signaling, EMT regulates the CSC phenotype by inducing the conversion of two types of complexes.55 In addition to activated Wnt signaling, inactivated Wnt signaling can also promote malignant progression of CSCs, especially tumor recurrence. Autocrine inhibition of WNT signaling induces the resting state of CSCs and tumor dormancy. After the primary tumor undergoes periodic proliferation and cancer immunoediting, a small fraction of the progenies are transformed into latency competent cancer (LCC) cells, and these cells exhibit Sox-dependent stem phenotype. LCC cells actively silence WNT signaling to enter into a quiescent state by expressing Sox2 target gene DKK1. The cancer cells then gain long-term viability through immune escape. When the function of cancer immunoediting is somehow hampered, LCC cells randomly enter cell cycle, triggering the next metastatic explosion.56
CSC-based therapy may be a viable therapeutic intervention for cancer. For example, retinoids trigger the upregulation of homeobox A5 (HOXA5) expression. HOXA5 antagonizes Wnt signaling in colon cancer by inducing Wnt signaling inhibitors, such as adenomatosis polyposis coli down-regulated 1 (APCDD1), CXXC finger protein 4 (CXXC4), and naked cuticle homolog 1 (NKD1), and promotes differentiation of intestinal cancer stem cells.57,58 However, we need to pay attention to the negative role of Wnt signaling in tumor recurrence. Since cancer immunoediting can effectively eliminate latent cells, cancer immunotherapy is a powerful means to suppress LCC cell activation.
Targeting Strategy for Wnt Signaling
Wnt/β-catenin signaling is involved in the regulation of cell transduction in tumors, which is a multi-faceted and multi-level complex process. Due to its complexity, Wnt/β-catenin signaling is still being explored as a target for the treatment of cancer. Currently, Wnt signaling has shown some promising targets: Fzd protein, β-catenin destruction complex, nuclear β-catenin, and enzymes Porcupine and tankyrase (TNKS). Small molecule drugs targeting Wnt signaling can be obtained via high throughput compound library screening, some of whose structures are shown in Figure 4.
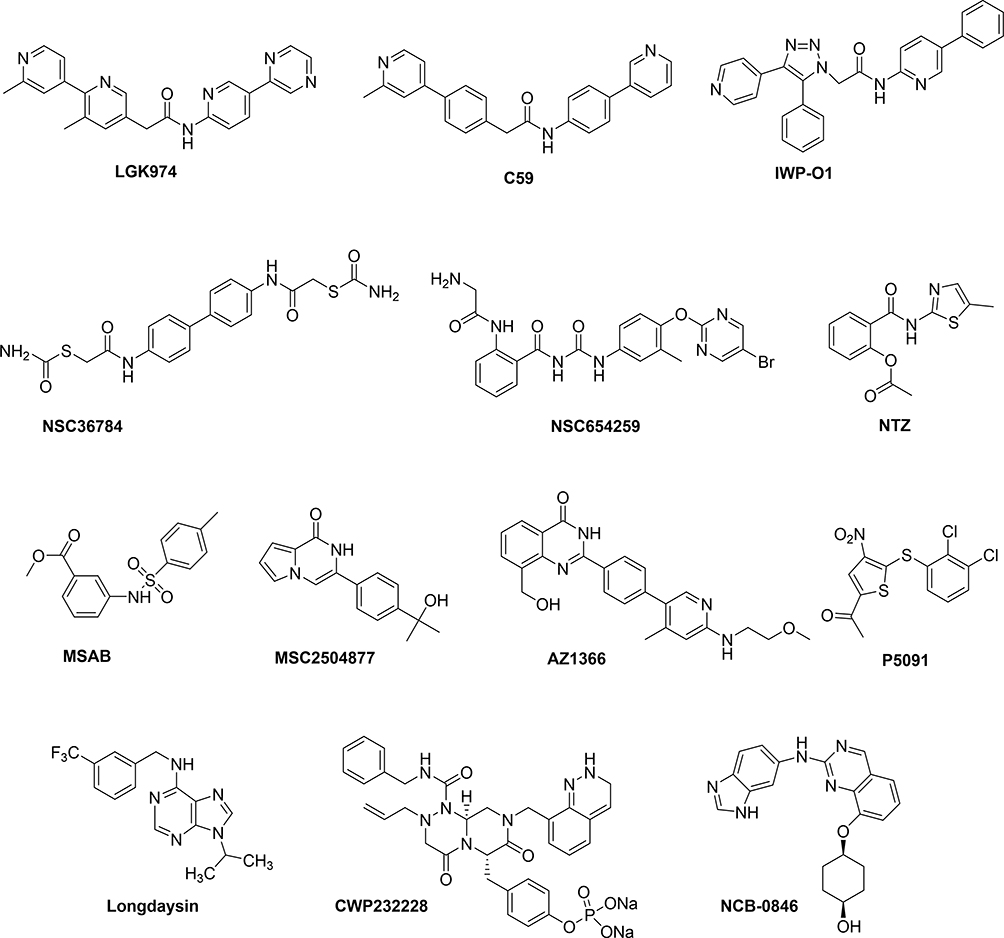 |
Figure 4 The structures of small molecules that downregulate Wnt signaling. |
Porcupine is a membrane-bound O-acyltransferase, which has been the focus of attention as a key enzyme for Wnt ligand synthesis. Small molecule inhibitors that specifically target Porcupine, such as LGK974 and C59 have been obtained, some of which can inhibit Wnt-driven cancers at effective therapeutic doses. A non-clinical study revealed that these drugs were not effective. Funck-Brentano et al demonstrated that Porcupine inhibitors have a harmful effect on bone mass and strength, which may increase the risk of fractures.59 Li et al encapsulated the drugs in liposomes and examined for the safety and efficacy of the drugs. In this method, the drug release was minimized in circulation, thereby reducing the cytotoxic effect of Porcupine inhibitors CGX1321. It is worthy of note that this drug delivery system yielded favorable effect on cancers.60 In addition, You et al reported that IWP-O1 was a highly effective Porcupine inhibitor with a triazole backbone. The inhibitory activity of IWP-O1 was 2.5 times more potent than that of the positive control drug LGK974, with significantly improved metabolic stability, and suitable for the study of mouse models.61
The FZD protein acts as the main receptor for Wnt signaling and usually harnesses its cysteine-rich domain (CRD) to interact with Wnt ligand to transduce the signal. Compounds that compete with Wnt ligand by binding FZD CRD with high affinity will thus attenuate Wnt-β-catenin signaling. Li et al screened a series of FZD8 CRD small molecule inhibitors through in silico molecular modeling and biophysical methods, among which NSC36784 and NSC654259 were the most effective.62 These compounds specifically bind to the Wnt binding site at submicromolar concentrations, and effectively inhibit Wnt signaling-induced LRP6 phosphorylation, and inhibit Wnt3a-induced signaling in a dose-dependent manner.
β-catenin is the core regulator of Wnt signaling, and its importance is palpable. Due to the unsatisfactory effectiveness of previously developed synthetic inhibitors against β-catenin (XAV939 and IWP-2) in pharmacokinetics and pharmacodynamics, however, in vivo preclinical studies were terminated. It is thus imperative to find other β-catenin targeted therapies. Latest research demonstrated that the small molecule nitazoxanide (NTZ) stabilizes the peptide arginine deiminase 2 (PAD2), promotes citrullination of β-catenin to induce its degradation and effectively inhibits the growth of colorectal tumors driven by APC or CTNNB1 mutations.63 So-Young Hwang et al reported that MSAB, a β-catenin inhibitor, which bound to β-catenin, promoted its degradation and down-regulated the expression of Wnt/β-catenin target genes c-MYC and Cyclin D1.64 Unfortunately, preclinical studies of these small molecules have not been conducted yet, probably because the optimization of the compounds has not been completed.
In addition to directly targeting β-catenin to inhibit Wnt signaling, compounds with different modes of action are also available, some of which indirectly promote degradation of β-catenin. Tankyrases 1/2 are coupled with Axin poly ADP-ribosylation (PARsylation). The complex is ubiquitinated by the RNF146 (The RING finger protein) E3 ubiquitin ligase and used for proteasome degradation, greatly reducing the level of Axin in tumor cells.65,66 Inhibiting TNKS can stabilize the level of intracellular Axin and indirectly down-regulate β-catenin expression. MSC2504877, a small molecule TNKS inhibitor, can inhibit APC mutant colorectal tumor cells and effectively suppress the intestinal stem cell proliferation phenotype;67 Another inhibitor AZ1366 significantly inhibited non-small cell lung cancer. When combined with the epidermal growth factor receptor (EGFR) inhibitors, the drug suppressed tumor growth and improved the survival of Wnt-responsive lung cancer.68 It was, however, reported69 that the clinical application of tankyrase inhibitors is limited by intestinal toxicity and the therapeutic index of mice was <1. In addition, Ubiquitin-specific protease 7 (USP7), a deubiquitinating enzyme, that is overexpressed in colorectal cancer cell lines and tissues, is associated with poor prognosis of the disease. Compound P5091 enhances β-catenin ubiquitination and degradation by inhibiting USP7, thereby inducing tumor apoptosis and inhibiting tumor growth.70 CK1 is a family of serine/threonine-specific protein kinases, and it exists as a positive/negative regulator of Wnt/β-catenin signaling. In Wnt OFF, CK1α phosphorylates β-catenin to promote its degradation. When Wnt signal is on, however, CK1 kinase phosphorylates LRP6 and DVL, resulting in the accumulation of β-catenin. Recently, it has been reported71 that Longdaysin, a small molecule drug, can significantly reduce the phosphorylation of LRP6 and DVL2 in breast cancer by inhibiting CK1δ/ε, thereby down-regulating Wnt signal transduction to exert anticancer effects.
Functional Wnt signaling requires the interaction between β-catenin and T cell factor in the nucleus, so antagonizing their binding could inhibit Wnt signaling. The CWP232228 small molecule inhibitor suppresses the growth of stem cell-like cells (BCSC) in breast cancer tumor mass by inhibiting β-catenin-mediated transcription. It is a new BCSC-targeted drug for the treatment of breast cancer.72 In addition, the regulators essentially involved in β-catenin and TCF4 transcription complexes are also targets for the development of cancer therapeutics, such as Traf2- and Nck-interacting protein kinase (TNIK). NCB-0846 inhibits Wnt signaling by disrupting the formation of TCF/β-catenin transcription complex, and ultimately inhibits Wnt-driven intestinal tumorigenesis and colorectal cancer stem cell activity in Apc (min/+) mice.73
Regulating cancer epigenetics can also be a potential therapeutic strategy. Ten eleven translocation 1 (TET1) is a tumor suppressor, which binds to the Wnt endogenous inhibitors DKK1 and SFRP2 gene promoters to maintain their hypomethylation. TET1 inhibitors thus promote the transcription of DKK1 and SFRP2 and exert effective tumor-suppressive effects.74,75 Therefore, TET1 can be considered as a new target for anti-cancer therapy. The Wnt signaling transcription factor OVO-like 2 (OVOL2) maintains the hypermethylation of Wnt target genes by promoting histone deacetylase 1 recruitment to the TCF4-β-catenin complex. Expressed high level of OVOL2 can inhibit Wnt activity and the metastasis of colorectal tumors.76 Recently, Zhang et al reported that targeting histone methyltransferase G9a restored the expression of Wnt signaling silencing gene APC2 through promoter demethylation, and then down-regulated Wnt signaling to inhibit the growth of lung cancer in vivo and in vitro.77
Conclusion
Wnt/β-catenin signaling is involved in the malignant progression of tumors. Targeting Wnt signaling thus has a great potential in the development of cancer therapy. However, careful attention has to be paid to the difference in the effector molecules of Wnt among cancers of different origins. Identification of the cancer-specific regulation of the Wnt signaling pathway is prerequisite for the development of cancer therapeutics based on the Wnt signaling therapy. In addition, we should carefully examine whether the therapy targeting Wnt/β-catenin signaling is a correct strategy under the disease background, and avoid obtaining opposite therapeutic effects. It has been shown that miRNAs directly and indirectly regulate Wnt signaling. The inhibition of Wnt signaling using siRNA, miRNA and/or pharmacological agents might also be promising interventions of cancer. In this review, we focused on the potential connection between Wnt signaling and stem cell heterogeneity, and how CSCs use Wnt and EMT to promote tumor invasion. It highlights the cardinal role of CSCs in cancer progression, hoping to provide new insights into CSC-based treatment. Finally, we summarized Wnt signal modulators, and mentioned the possible side effects of Wnt inhibitors. We believe that designing more efficient drug delivery systems is required for the future development of cancer treatment. We should continue to delineate the relationship between Wnt signaling and cancer, by which we will be able to harness the information on Wnt signaling pathways to clinical studies.
Acknowledgments
We gratefully acknowledge the financial support from National Natural Science Foundation of China (project nos. 21572207 (Wen Zhang), 21877101 (Wen Zhang)) and the Science Foundation of National Health Commission of China (Project No.: WKJ-ZJ-1713 (Yanling Wu)). We also thank Prof. Mingdong Huang and Dr. Cai Yuan polishing the manuscript.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Zhan T, Rindtorff N, Boutros M. Wnt signaling in cancer. Oncogene. 2017;36(11):1461–1473. doi:10.1038/onc.2016.304
2. Majidinia M, Aghazadeh J, Jahanban-Esfahlani R, Yousefi B. The roles of Wnt/β-catenin pathway in tissue development and regenerative medicine. J Cell Physiol. 2018;233(8):5598–5612. doi:10.1002/jcp.26265
3. Wang B, Tian T, Kalland KH, Ke X, Qu Y. Targeting Wnt/β-catenin signaling for cancer immunotherapy. Trends Pharmacol Sci. 2018;39(7):648–658. doi:10.1016/j.tips.2018.03.008
4. Clevers H, Nusse R. Wnt/β-catenin signaling and disease. Cell. 2012;149(6):1192–1205. doi:10.1016/j.cell.2012.05.012
5. Banziger C, Soldini D, Schutt C, Zipperlen P, Hausmann G, Basler K. Wntless, a conserved membrane protein dedicated to the secretion of Wnt proteins from signaling cells. Cell. 2006;125(3):509–522. doi:10.1016/j.cell.2006.02.049
6. Nusse R, Clevers H. Wnt/β-catenin signaling, disease, and emerging therapeutic modalities. Cell. 2017;169(6):985–999. doi:10.1016/j.cell.2017.05.016
7. Cadigan KM, Waterman ML. TCF/LEFs and Wnt signaling in the nucleus. Cold Spring Harb Perspect Biol. 2012;4(11):a007906. doi:10.1101/cshperspect.a007906
8. Dow LE, O’Rourke KP, Simon J, et al. Apc restoration promotes cellular differentiation and reestablishes crypt homeostasis in colorectal cancer. Cell. 2015;161(7):1539–1552. doi:10.1016/j.cell.2015.05.033
9. Liu H, Du S, Lei T, et al. Multifaceted regulation and functions of YAP/TAZ in tumors (Review). Oncol Rep. 2018;40(1):16–28. doi:10.3892/or.2018.6423
10. Hao H-X, Jiang X, Cong F. Control of Wnt receptor turnover by R-spondin-ZNRF3/RNF43 signaling module and its dysregulation in cancer. Cancers. 2016;8(6):54. doi:10.3390/cancers8060054
11. Pai VC, Hsu -C-C, Chan T-S, et al. ASPM promotes prostate cancer stemness and progression by augmenting Wnt-Dvl-3-β-catenin signaling. Oncogene. 2019;38(8):1340–1353. doi:10.1038/s41388-018-0497-4
12. Bengoa-Vergniory N, Kypta RM. Canonical and noncanonical Wnt signaling in neural stem/progenitor cells. Cell Mol Life Sci. 2015;72(21):4157–4172. doi:10.1007/s00018-015-2028-6
13. Lang CMR, Chan CK, Veltri A, Lien WH. Wnt signaling pathways in Keratinocyte Carcinomas. Cancers. 2019;11(9):1216. doi:10.3390/cancers11091216
14. Corda G, Sala A. Non-canonical WNT/PCP signalling in cancer: Fzd6 takes centre stage. Oncogenesis. 2017;6(7):e364. doi:10.1038/oncsis.2017.69
15. Koveitypour Z, Panahi F, Vakilian M, et al. Signaling pathways involved in colorectal cancer progression. Cell Biosci. 2019;9:97. doi:10.1186/s13578-019-0361-4
16. Kim HJ, Moon SJ, Kim SH, Heo K, Kim JH. DBC1 regulates Wnt/β-catenin-mediated expression of MACC1, a key regulator of cancer progression, in colon cancer. Cell Death Dis. 2018;9(8):831. doi:10.1038/s41419-018-0899-9
17. Wang Q, Zhou Y, Rychahou P, et al. Deptor is a novel target of Wnt/β-catenin/c-Myc and contributes to colorectal cancer cell growth. Cancer Res. 2018;78(12):3163–3175. doi:10.1158/0008-5472.CAN-17-3107
18. Dong Y, Zhang Y, Kang W, et al. VSTM2A suppresses colorectal cancer and antagonizes Wnt signaling receptor LRP6. Theranostics. 2019;9(22):6517–6531. doi:10.7150/thno.34989
19. Gou H, Liang JQ, Zhang L, et al. TTPAL promotes colorectal tumorigenesis by stabilizing TRIP6 to activate Wnt/β-catenin signaling. Cancer Res. 2019;79(13):3332–3346. doi:10.1158/0008-5472.CAN-18-2986
20. Mir R, Pradhan SJ, Patil P, Mulherkar R, Galande S. Wnt/β-catenin signaling regulated SATB1 promotes colorectal cancer tumorigenesis and progression. Oncogene. 2016;35(13):1679–1691. doi:10.1038/onc.2015.232
21. Planas-Paz L, Orsini V, Boulter L, et al. The RSPO–LGR4/5–ZNRF3/RNF43 module controls liver zonation and size. Nat Cell Biol. 2016;18(5):467–479. doi:10.1038/ncb3337
22. Jung Y-S, Stratton SA, Lee SH, et al. TMEM9-v-ATPase activates Wnt/β-catenin signaling via APC lysosomal degradation for liver regeneration and tumorigenesis. Hepatology. 2020. doi:10.1002/hep.31305
23. Lee J-S. The mutational landscape of hepatocellular carcinoma. Clin Mol Hepatol. 2015;21(3):220–229. doi:10.3350/cmh.2015.21.3.220
24. Khalaf AM, Fuentes D, Morshid AI, et al. Role of Wnt/β-catenin signaling in hepatocellular carcinoma, pathogenesis, and clinical significance. J Hepatocell Carcinoma. 2018;5:61–73. doi:10.2147/JHC.S156701
25. Nault J-C, Zucman-Rossi J. TERT promoter mutations in primary liver tumors. Clin Res Hepatol Gastroenterol. 2016;40(1):9–14. doi:10.1016/j.clinre.2015.07.006
26. Tao J, Xu E, Zhao Y, et al. Modeling a human hepatocellular carcinoma subset in mice through coexpression of met and point-mutant β-catenin. Hepatology. 2016;64(5):1587–1605. doi:10.1002/hep.28601
27. Colnot S. Focusing on beta-catenin activating mutations to refine liver tumor profiling. Hepatology. 2016;64(6):1850–1852. doi:10.1002/hep.28761
28. Li N, Wei L, Liu X, et al. A frizzled-like cysteine-rich domain in Glypican-3 mediates Wnt binding and regulates hepatocellular carcinoma tumor growth in mice. Hepatology. 2019;70(4):1231–1245. doi:10.1002/hep.30646
29. Angeloni V, Tiberio P, Appierto V, Daidone MG. Implications of stemness-related signaling pathways in breast cancer response to therapy. Semin Cancer Biol. 2015;31:43–51. doi:10.1016/j.semcancer.2014.08.004
30. Li K, Pan W-T, Ma Y-B, et al. BMX activates Wnt/β-catenin signaling pathway to promote cell proliferation and migration in breast cancer. Breast Cancer. 2020;27(3):363–371. doi:10.1007/s12282-019-01024-8
31. Zhuang X, Zhang H, Li X, et al. Differential effects on lung and bone metastasis of breast cancer by Wnt signalling inhibitor DKK1. Nat Cell Biol. 2017;19(10):1274–1285. doi:10.1038/ncb3613
32. Lu Y, Zhao X, Liu Q, et al. lncRNA MIR100HG-derived miR-100 and miR-125b mediate cetuximab resistance via Wnt/β-catenin signaling. Nat Med. 2017;23(11):1331–1341. doi:10.1038/nm.4424
33. Han P, Li J-W, Zhang B-M, et al. The lncRNA CRNDE promotes colorectal cancer cell proliferation and chemoresistance via miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol Cancer. 2017;16(1):9. doi:10.1186/s12943-017-0583-1
34. Huang L, Liu Z, Hu J, et al. MiR-377-3p suppresses colorectal cancer through negative regulation on Wnt/β-catenin signaling by targeting XIAP and ZEB2. Pharmacol Res. 2020;156:104774. doi:10.1016/j.phrs.2020.104774
35. Quan H, Li B, Yang J. MicroRNA-504 functions as a tumor suppressor in hepatocellular carcinoma through inhibiting Frizzled-7-mediated-Wnt/β-catenin signaling. Biomed Pharmacother. 2018;107:754–762. doi:10.1016/j.biopha.2018.07.150
36. Yang C, Xu Y, Cheng F, et al. miR-1301 inhibits hepatocellular carcinoma cell migration, invasion, and angiogenesis by decreasing Wnt/β-catenin signaling through targeting BCL9. Cell Death Dis. 2017;8(8):e2999. doi:10.1038/cddis.2017.356
37. Yao B, Li Y, Wang L, et al. MicroRNA-3194-3p inhibits metastasis and epithelial-mesenchymal transition of hepatocellular carcinoma by decreasing Wnt/β-catenin signaling through targeting BCL9. Artif Cells Nanomed Biotechnol. 2019;47(1):3885–3895. doi:10.1080/21691401.2019.1670190
38. Wang H, Ou J, Jian Z, Ou Y. miR-186 modulates hepatocellular carcinoma cell proliferation and mobility via targeting MCRS1-mediated Wnt/β-catenin signaling. J Cell Physiol. 2019;234(12):23135–23145. doi:10.1002/jcp.28878
39. Ren L, Chen H, Song J, et al. MiR-454-3p-mediated Wnt/β-catenin signaling antagonists suppression promotes breast cancer metastasis. Theranostics. 2019;9(2):449–465. doi:10.7150/thno.29055
40. Li M, Han Y, Zhou H, et al. Transmembrane protein 170B is a novel breast tumorigenesis suppressor gene that inhibits the Wnt/β-catenin pathway. Cell Death Dis. 2018;9(2):91. doi:10.1038/s41419-017-0128-y
41. Liu S, Wang Z, Liu Z, et al. miR-221/222 activate the Wnt/β-catenin signaling to promote triple-negative breast cancer. J Mol Cell Biol. 2018;10(4):302–315. doi:10.1093/jmcb/mjy041
42. Yang F, Xiao Z, Zhang S. Knockdown of miR-194-5p inhibits cell proliferation, migration and invasion in breast cancer by regulating the Wnt/β-catenin signaling pathway. Int J Mol Med. 2018;42(6):3355–3363. doi:10.3892/ijmm.2018.3897
43. Lv C, Li F, Li X, et al. MiR-31 promotes mammary stem cell expansion and breast tumorigenesis by suppressing Wnt signaling antagonists. Nat Commun. 2017;8(1):1036. doi:10.1038/s41467-017-01059-5
44. Gao J, Yu S-R, Yuan Y, et al. MicroRNA-590-5p functions as a tumor suppressor in breast cancer conferring inhibitory effects on cell migration, invasion, and epithelial-mesenchymal transition by downregulating the Wnt-β-catenin signaling pathway. J Cell Physiol. 2019;234(2):1827–1841. doi:10.1002/jcp.27056
45. Leung WKC, He M, Chan AWH, Law PTY, Wong N. Wnt/β-Catenin activates MiR-183/96/182 expression in hepatocellular carcinoma that promotes cell invasion. Cancer Lett. 2015;362(1):97–105. doi:10.1016/j.canlet.2015.03.023
46. Li T, Jian X, He H, et al. MiR-452 promotes an aggressive colorectal cancer phenotype by regulating a Wnt/β-catenin positive feedback loop. J Exp Clin Cancer Res. 2018;37(1):238. doi:10.1186/s13046-018-0879-z
47. Wang W, Xiao X, Chen X, et al. Tumor-suppressive miR-145 co-repressed by TCF4-β-catenin and PRC2 complexes forms double-negative regulation loops with its negative regulators in colorectal cancer. Int J Cancer. 2018;142(2):308–321. doi:10.1002/ijc.31056
48. Atlasi Y, Looijenga L, Fodde R. Cancer stem cells, pluripotency, and cellular heterogeneity: a WNTer perspective. Curr Top Dev Biol. 2014;107:373–404. doi:10.1016/B978-0-12-416022-4.00013-5
49. de Sousa e Melo F, Kurtova AV, Harnoss JM, et al. A distinct role for Lgr5 stem cells in primary and metastatic colon cancer. Nature. 2017;543(7647):676–680. doi:10.1038/nature21713
50. McKenzie MD, Ghisi M, Oxley EP, et al. Interconversion between tumorigenic and differentiated states in acute myeloid leukemia. Cell Stem Cell. 2019;25(2):258–272. doi:10.1016/j.stem.2019.07.001
51. Huels DJ, Sansom OJ. Stem vs non-stem cell origin of colorectal cancer. Br J Cancer. 2015;113(1):1–5. doi:10.1038/bjc.2015.214
52. Shin S, Wangensteen KJ, Teta-Bissett M, et al. Genetic lineage tracing analysis of the cell of origin of hepatotoxin-induced liver tumors in mice. Hepatology. 2016;64(4):1163–1177. doi:10.1002/hep.28602
53. Reilein A, Melamed D, Park KS, et al. Alternative direct stem cell derivatives defined by stem cell location and graded Wnt signalling. Nat Cell Biol. 2017;19(5):433–444. doi:10.1038/ncb3505
54. Liao W-Y, Hsu -C-C, Chan T-S, et al. Dishevelled 1-regulated superpotent cancer stem cells mediate Wnt heterogeneity and tumor progression in hepatocellular carcinoma. Stem Cell Rep. 2020;14(3):462–477. doi:10.1016/j.stemcr.2020.02.003
55. Chang YW, Su YJ, Hsiao M, et al. Diverse targets of β-catenin during the epithelial-mesenchymal transition define cancer stem cells and predict disease relapse. Cancer Res. 2015;75(16):3398–3410. doi:10.1158/0008-5472.CAN-14-3265
56. Malladi S, Macalinao DG, Jin X, et al. Metastatic latency and immune evasion through autocrine inhibition of WNT. Cell. 2016;165(1):45–60. doi:10.1016/j.cell.2016.02.025
57. Ordonez-Moran P, Dafflon C, Imajo M, Nishida E, Huelsken J. HOXA5 counteracts stem cell traits by inhibiting Wnt signaling in colorectal cancer. Cancer Cell. 2015;28(6):815–829. doi:10.1016/j.ccell.2015.11.001
58. Tan SH, Barker N. Stemming colorectal cancer growth and metastasis: HOXA5 forces cancer stem cells to differentiate. Cancer Cell. 2015;28(6):683–685. doi:10.1016/j.ccell.2015.11.004
59. Funck-Brentano T, Nilsson KH, Brommage R, et al. Porcupine inhibitors impair trabecular and cortical bone mass and strength in mice. J Endocrinol. 2018;238(1):13–23. doi:10.1530/JOE-18-0153
60. Li C, Liang Y, Cao J, et al. The delivery of a Wnt pathway inhibitor toward CSCs requires stable liposome encapsulation and delayed drug release in tumor tissues. Mol Ther. 2019;27(9):1558–1567. doi:10.1016/j.ymthe.2019.06.013
61. You L, Zhang C, Yarravarapu N, et al. Development of a triazole class of highly potent Porcn inhibitors. Bioorg Med Chem Lett. 2016;26(24):5891–5895. doi:10.1016/j.bmcl.2016.11.012
62. Lee H, Bao J, Miller A, et al. Structure-based discovery of novel small molecule Wnt signaling inhibitors by targeting the cysteine rich domain of Frizzled. J Biol Chem. 2015;290(51):30596–30606. doi:10.1074/jbc.M115.673202
63. Qu Y, Olsen JR, Yuan X, et al. Small molecule promotes β-catenin citrullination and inhibits Wnt signaling in cancer. Nat Chem Biol. 2018;14(1):94–101. doi:10.1038/nchembio.2510
64. Hwang SY, Deng X, Byun S, et al. Direct targeting of β-catenin by a small molecule stimulates proteasomal degradation and suppresses oncogenic Wnt/β-catenin signaling. Cell Rep. 2016;16(1):28–36. doi:10.1016/j.celrep.2016.05.071
65. Ferri M, Liscio P, Carotti A, et al. Targeting Wnt-driven cancers: discovery of novel tankyrase inhibitors. Eur J Med Chem. 2017;142:506–522. doi:10.1016/j.ejmech.2017.09.030
66. Arques O, Chicote I, Puig I, et al. Tankyrase inhibition blocks Wnt/β-catenin pathway and reverts resistance to PI3K and AKT inhibitors in the treatment of colorectal cancer. Clin Cancer Res. 2016;22(3):644–656. doi:10.1158/1078-0432.CCR-14-3081
67. Menon M, Elliott R, Bowers L, et al. A novel tankyrase inhibitor, MSC2504877, enhances the effects of clinical CDK4/6 inhibitors. Sci Rep. 2019;9(1):201. doi:10.1038/s41598-018-36447-4
68. Scarborough H, Helfrich B, Casasselves M, et al. AZ1366: an inhibitor of tankyrase and the canonical Wnt pathway that limits the persistence of non–small cell lung cancer cells following EGFR inhibition. Clin Cancer Res. 2017;23(6):1531–1541. doi:10.1158/1078-0432.CCR-16-1179
69. Zhong Y, Katavolos P, Nguyen T, et al. Tankyrase inhibition causes reversible intestinal toxicity in mice with a therapeutic index < 1. Toxicol Pathol. 2016;44(2):267–278. doi:10.1177/0192623315621192
70. An T, Gong Y, Li X, et al. USP7 inhibitor P5091 inhibits Wnt signaling and colorectal tumor growth. Biochem Pharmacol. 2017;131(1):29–39. doi:10.1016/j.bcp.2017.02.011
71. Xiong Y, Zhou L, Su Z, et al. Longdaysin inhibits Wnt/β-catenin signaling and exhibits antitumor activity against breast cancer. Onco Targets Ther. 2019;12:993–1005. doi:10.2147/OTT.S193024
72. Jang G, Hong I, Kim R, et al. Wnt/β-catenin small-molecule inhibitor CWP232228 preferentially inhibits the growth of breast cancer stem-like cells. Cancer Res. 2015;75(8):1691–1702. doi:10.1158/0008-5472.CAN-14-2041
73. Masuda M, Sawa M, Yamada T. Therapeutic targets in the Wnt signaling pathway: feasibility of targeting TNIK in colorectal cancer. Pharmacol Ther. 2015;156:1–9. doi:10.1016/j.pharmthera.2015.10.009
74. Neri F, Dettori D, Incarnato D, et al. TET1 is a tumour suppressor that inhibits colon cancer growth by derepressing inhibitors of the WNT pathway. Oncogene. 2015;34(32):4168–4176. doi:10.1038/onc.2014.356
75. Duan H, Yan Z, Chen W, et al. TET1 inhibits EMT of ovarian cancer cells through activating Wnt/β-catenin signaling inhibitors DKK1 and SFRP2. Gynecol Oncol. 2017;147(2):408–417. doi:10.1016/j.ygyno.2017.08.010
76. Ye G, Sun GB, Jiao P, et al. OVOL2, an inhibitor of WNT signaling, reduces invasive activities of human and mouse cancer cells and is downregulated in human colorectal tumors. Gastroenterology. 2016;150(3):659–671.e616. doi:10.1053/j.gastro.2015.11.041
77. Zhang K, Wang J, Yang L, et al. Targeting histone methyltransferase G9a inhibits growth and Wnt signaling pathway by epigenetically regulating HP1α and APC2 gene expression in non-small cell lung cancer. Mol Cancer. 2018;17(1):1–15. doi:10.1186/s12943-018-0896-8
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.