")
Back to Journals » Journal of Pain Research » Volume 15
Neurometabolite Levels and Relevance to Central Sensitization in Chronic Orofacial Pain Patients: A Magnetic Resonance Spectroscopy Study
Authors Terumitsu M , Takado Y, Fukuda KI, Kato E, Tanaka S
Received 21 February 2022
Accepted for publication 6 May 2022
Published 16 May 2022 Volume 2022:15 Pages 1421—1432
DOI https://doi.org/10.2147/JPR.S362793
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr David Keith
Makoto Terumitsu,1,2 Yuhei Takado,3 Ken-Ichi Fukuda,2 Eisuke Kato,2 Sei Tanaka4
1Division of Dental Anesthesiology, Department of Human Biology and Pathophysiology, Health Sciences University of Hokkaido, Hokkaido, Japan; 2Division of Special Needs Dentistry and Orofacial Pain, Department of Oral Health and Clinical Science, Tokyo Dental College, Tokyo, Japan; 3Department of Functional Brain Imaging, Institute of Quantum Medical Sciences, National Institutes for Quantum Science and Technology, Chiba, Japan; 4Department of Oral and Maxillofacial Surgery, Tokyo Dental College, Tokyo, Japan
Correspondence: Makoto Terumitsu, Division of Dental Anesthesiology, Department of Human Biology and Pathophysiology, Health Sciences University of Hokkaido, 1757 Kanazawa, Tobetsu-cho, Ishikari-gun, Hokkaido, 061-0293, Japan, Tel/Fax +81 133 23 1445, Email [email protected]
Background: Refractory chronic pain in the orofacial region involves central sensitization (CS). However, not all chronic pain patients exhibit CS. An objective assessment of CS may be useful for pain management. Changes in the balance of excitatory and inhibitory neural activity or excessive activity of nerves and glial cells may cause CS and contribute to pain chronification.
Patients and Methods: 1H-magnetic resonance spectra were acquired from the anterior cingulate cortex (ACC) and thalamus in 20 patients with chronic orofacial pain and suspected CS, and 21 healthy volunteers, using a single-voxel point-resolved spectroscopy sequence. The patients were assessed using the Central Sensitization Inventory.
Results: Aspartate/total creatine (tCr) and glutathione in the ACC were significantly higher in the patient group. However, no significant difference was observed between groups in the neurometabolites measured in the thalamus. Patients also exhibited a tendency for increased gamma-aminobutyric acid (GABA)/tCr in the ACC. There were positive relationships between Central Sensitization Inventory scores and glutamate + glutamine (Glx) in the thalamus, a positive trend for Glx in the ACC and a negative relationship for GABA/tCr in the ACC.
Conclusion: The high levels of aspartate/tCr and glutathione in the patient group suggest excitatory neuronal activity and hyperactivity of neurons and glial cells. The correlation analysis results suggest that excitatory and inhibitory neurometabolites are involved in the chronification of orofacial pain, including CS.
Keywords: orofacial pain, central sensitization, magnetic resonance spectroscopy, Central Sensitization Inventory, anterior cingulate cortex, thalamus
Introduction
Chronic pain in the orofacial region becomes refractory and degrades patients’ quality of life in many cases. Central sensitization (CS) is a possible cause of the transition from acute to chronic pain.1 CS is clinically characterized by disproportionate pain, in which the severity of pain and functional disability are disproportionate to detectable tissue damage and pathophysiology, or which occurs regardless of peripheral lesions.2 CS has been defined as “an amplification of neural signaling within the central nervous system that elicits pain hypersensitivity.”3 Altered brain and brainstem function in the pain processing system corresponds to CS, including functional connectivity changes among brain regions,4 neural plasticity, such as new synapse formation5 and long-term potentiation,6 imbalance of the activity between excitatory and inhibitory neurons,7 and excess glial cell activation.8 To evaluate CS, neurophysiological examinations (quantitative sensory testing, and various stimuli),9 questionnaires,10 functional magnetic resonance imaging (fMRI), and cerebral metabolite measurement (proton magnetic resonance spectroscopy: 1H-MRS) have been used in medical practice and research.
Hyperactivity or hypoactivity of excitatory and inhibitory neurons are important factors in CS. Glutamate (Glu) is a prominent excitatory neurometabolite in the central nervous system and activates several receptors, such as the N-methyl-D-Aspartate (NMDA) receptor, in which excessive activation causes high permeability to calcium and sodium ions, and acts as both a trigger and effector of CS.11 Phosphorylation of these receptors increases their hypersensitivity and decreases inhibitory transmission by reducing gamma-aminobutyric acid (GABA) and glycine tonic inhibition.12 Positive allosteric modulation of the GABA-A receptor reduces CS.13 1H-MRS can be used for noninvasive quantitative measurement of neurometabolites related to excitatory and inhibitory neural activity in the human brain. Previous studies have detected changes in the levels of several neurometabolites corresponding to specific cerebral regions, and these changes have been extensively examined in the anterior cingulate cortex (ACC) and thalamus in chronic pain patients. The ACC and thalamus are involved in the affective components of pain and the essential relay of nociceptive inputs to transmit signals from the brainstem to the cortex, respectively. Moreover, the mediodorsal and intralaminar thalamic nuclei send nociceptive signals to the ACC, suggesting abnormal processing of the affective component.14,15 Various increases or decreases in the patterns of excitatory and inhibitory neurometabolites have been observed in these brain regions in previous 1H-MRS studies of chronic pain.16
1H-MRS studies have been conducted to measure Glu, glutamine (Gln), and GABA in a range of pain conditions and various brain regions.16 However, neurometabolite findings of studies examining chronic pain remain controversial. Trends toward increases in both Glu/total creatine (tCr) and Glu + Gln (Glx)/tCr have been reported in the ACC in patients with various chronic pain conditions in comparison with healthy controls,17 whereas significantly lower Glx or Glu in ACC were reported in patients with severe neuropathic pain caused by spinal cord injury18 and low back pain.19 Other studies reported no significant changes in excitatory or inhibitory neurometabolites in chronic pain patients with low back pain,20,21 knee osteoarthritis,22 musculoskeletal pain23 and migraine.24 The level of GABA in ACC has been reported to be reduced in fibromyalgia25 and pelvic pain patients.26 Migraine patients were found to exhibit significantly increased levels of Glx in the thalamus,27 whereas lower GABA levels were observed in patients with neuropathic pain in the trigeminal regions28 and spinal cord.29 Excitatory neurometabolites in the thalamus were reported to exhibit no significant differences in patients with fibromyalgia30,31 and migraine.24
The nociceptive pathway via the trigeminal nerve differs from that in the extremities of the limbs and body. Few 1H-MRS studies have examined the orofacial region. In addition, although numerous studies have investigated chronic pain, not all chronic pain patients exhibit CS. Therefore, in the current study we aimed to elucidate changes in neurometabolites associated with excitatory and inhibitory neural and glial activity in the ACC and thalamus in CS patients with chronic orofacial pain, and to examine the correlation between CS questionnaire responses and neurometabolite levels.
Materials and Methods
Participants
Twenty patients (18 females and two males) with chronic pain in the orofacial region were recruited from the pain clinic of Tokyo Dental College at Suidobashi Hospital. The patients had spontaneous and persistently continuing pain with widespread dysesthesia and/or static allodynia. One of the criteria for CS included disproportionate pain regardless of peripheral lesions2 as determined by imaging examinations, such as computed tomography, conventional MRI, MR neurography (three-dimensional volume rendering magnetic resonance neurography: 3DVR-MRN),32 and 3D anisotropy contrast (3DAC) on the basis of diffusion-weighted imaging.33 In addition, pain control was difficult, and patients were unresponsive or only temporarily responsive to conventional pain therapies such as various medications, local or nerve block anesthesia and local anesthetic injection into myofascial trigger points. Patients’ mean age was 52.4 ± 12.8 years (mean ± standard deviation: SD). Pain duration ranged from 0.5 to 25 years, and the mean duration was 7.6 ± 7.3 years (mean ± SD). Table 1 shows patients’ site and type of pain, occasion of pain, International Classification of Orofacial Pain (ICOP)34 and medications. Already prescribed medication was continued during the study. ICOP classified the patients into three groups: 1) probable persistent idiopathic dentoalveolar pain (n = 9), 2) other trigeminal neuropathic pain (n = 5) and 3) chronic primary myofascial orofacial pain (n = 6) (as shown in Table 1). The type of pain was used to categorize patients into three groups: 1) chronic postsurgical pain (n = 12), 2) myofascial pain (n = 6) and 3) others (n = 2) (as shown in Table 1).
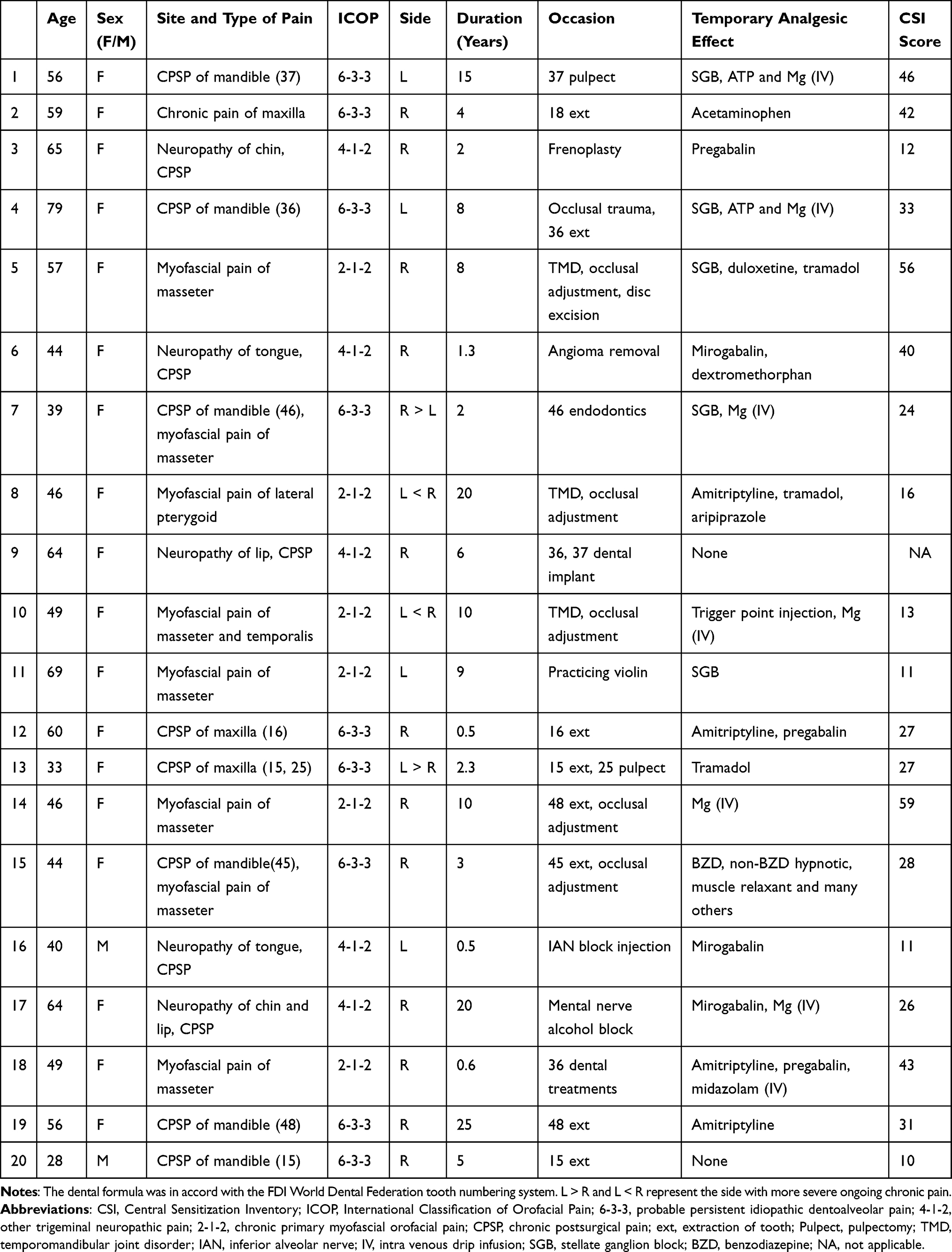 |
Table 1 Orofacial Chronic Pain Patients’ Characteristics |
Twenty-one sex- and age-matched healthy volunteers (19 females and two males) without any pain, medication for central nervous system conditions, or cerebral nerve disease participated. The mean age of the control group was 53.2 ± 12.9 years (mean ± SD), and there was no significant difference in age between the control group and the patient group (p = 0.61, Student’s t-test).
All participants provided written informed consent. The study protocol was approved by the Ethics Committee of Tokyo Dental College (#923).
Questionnaire Evaluation
The Central Sensitization Inventory (CSI) was used as a self-report questionnaire tool.10 The patients completed the Japanese version of the CSI.35 Patients were given the questionnaire to fill out at home and return to us on a day close to the MRI scanning. Scores were calculated using a five-point evaluation for 25 question items in part A of the CSI, and total scores ranged from 0 to 100.
Magnetic Resonance Spectroscopy
All experiments were performed using a 3.0 T Ingenia Philips MRI scanner (Philips Healthcare, Best, Netherlands) with a 32-channel receive head coil and a multitransmit body coil. The MRS protocol included a single-voxel point-resolved spectroscopy (PRESS) sequence with chemical-shift selective water suppression. The MRS acquisition parameters were as follows: repetition time/echo time (TE) = 2000/35 ms, complex points for the spectral data = 1024, and total number of acquisitions = 128. The linewidth of the water spectrum in magnitude mode became smaller than 18 Hz after shimming.
Whole brain images were acquired using sagittal 3D T1-weighted gradient echo imaging with the following parameters: field of view (FOV) = 25.6×25.6 cm, voxel size = 0.5×0.5×0.5 mm, repetition time/TE = 6.3/3.5 ms and flip angle = 8°.
Voxel Location
1H-MRS voxels of interest (VOIs) were set in the ACC (20×30×20 mm) and thalamus (20×20×20 mm), as shown in Figure 1. VOIs in the ACC were positioned parallel to and above the corpus callosum, starting from the genu of the corpus callosum and extending 3 cm posteriorly and covered left and right ACC17 in patients. VOIs in the thalamus were set in the side that was contralateral to that of the ongoing chronic pain. Similar VOIs were located in a healthy volunteer who matched the patients in terms of sex and approximate age.
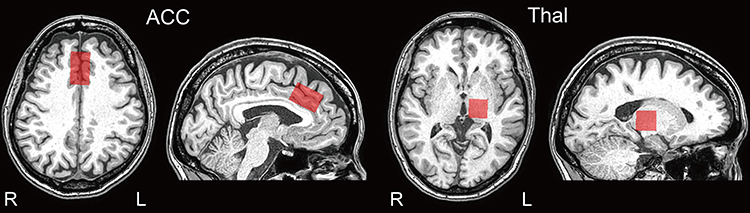 |
Figure 1 Positions of voxels of interests. Abbreviations: ACC, anterior cingulate cortex; Thal, thalamus. Notes: Each 1H-MRS voxel of interest (VOI) was set in the ACC (20 × 30×20 mm) and Thal (20 × 20×20 mm). |
MRS Data Processing and Measurement
The MRS data were processed using a linear combination model (LCModel Version 6.3).36 Tissue composition inside the VOI was calculated on the basis of the segmentation of 3D T1-weighted images using Gannet3.0.37 Water concentrations used in the LCModel analysis were calculated on the basis of the volume fractions of white matter (WM), gray matter (GM), and cerebrospinal fluid (CSF). Metabolite concentrations were then divided by the fraction of WM and GM to correct for CSF inside the VOI, because metabolites are mainly present in the WM and GM.38 Water scaling and eddy-current correction were performed and metabolites were fitted in the chemical shift range (0.2–4.2 ppm).
We measured Glu, Glx, aspartate (Asp)/tCr, GABA/tCr, and glutathione (GSH). The tCr (Cr + PCr) was used for normalization, because tCr is widely used as an internal reference in human studies.39 Glu was analyzed independently because it was reliably determined in high quality control of spectra at 3T with a short TE.40 Glx and GABA/tCr were evaluated for correlation analysis between the neurometabolites and CSI scores in the patient group.
Metabolites processed by the LCModel with Cramer-Rao Lower Bounds < 25% for GSH and 15% for the others were analyzed. A representative 1H MR spectrum obtained from the ACC and thalamus, LCModel spectral fit, and neurometabolite fits in a patient are shown in Figure 2. Mean signal-to-noise ratio (SNR) ± SD values for patients and controls were 27.35 ± 6.67, 31.29 ± 2.97 in the ACC and 19.55 ± 2.63, 20.38 ± 2.58 in the thalamus, respectively. Mean full-width of half maximum (FWHM) ± SD values for the patients and controls were 0.033 ± 0.010 ppm and 0.037 ± 0.006 ppm in the ACC and 0.051 ± 0.005 ppm and 0.049 ± 0.006 ppm in the thalamus, respectively. No significant differences were observed in FWHM in both regions between the patients and the controls (F[1, 78] = 2.9929, p = 0.0876, two-way analysis of variance [ANOVA]). Patients’ data showed lower SNR compared with that of the controls in the ACC, but there were no significant differences in the thalamus (F[1, 78] = 3.9945, p = 0.04913, Tukey’s honestly significant difference test for post hoc test: p = 0.0141 [ACC], p = 0.914 [thalamus]).
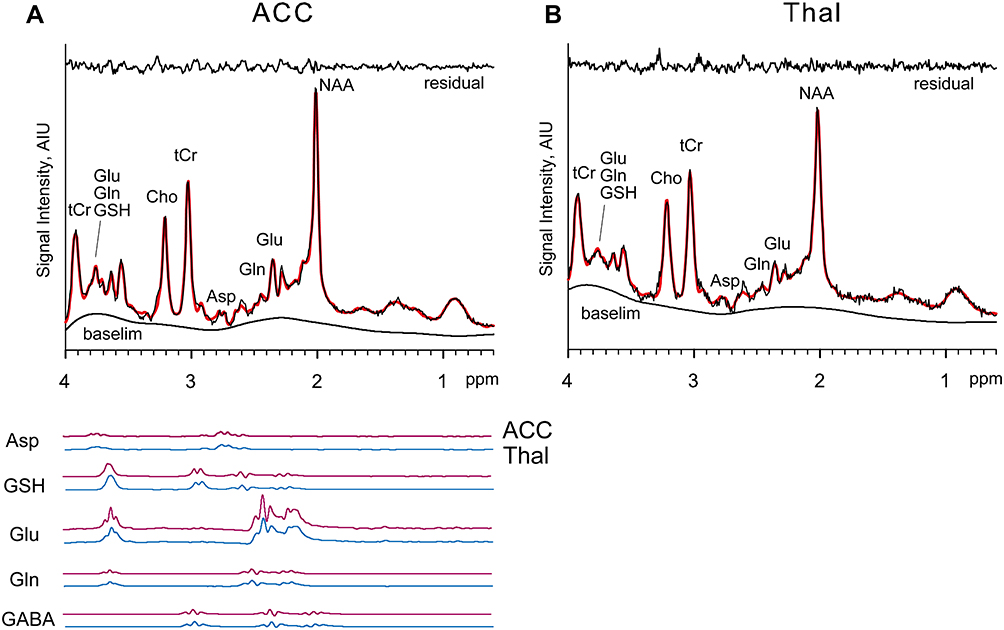 |
Figure 2 Spectra and LCModel fitting. Abbreviations: Asp, aspartate; tCr, creatine (Cr) + PCr; GABA, gamma-aminobutyric acid; GSH, glutathione; Glu, glutamate; Gln, glutamine; mI, Cho, choline; NAA, N-acetylaspartate; AIU, arbitrary signal intensity institutional units ACC, anterior cingulate cortex; Thal, thalamus. Notes: Example spectra and LCModel fits from ACC (A) and Thal (B) in one participant are shown. The spectra and fitted line are indicated in black and red line, respectively. The residual signals following fitting are shown at the top of each panel. LCModel estimated baselines are shown at the bottom of each panel. |
Statistical Analysis
Group differences were tested using Student’s t-test with Bonferroni-adjusted p-value < 0.005 on each 1H-MRS measure (Glu, Glx, Asp/tCr, GABA/tCr, and GSH) separately for each of the two voxel regions in ACC and thalamus between the patients and the controls. Correlation coefficients were calculated between the neurometabolites (Glx and GABA/tCr) of the two regions and CSI scores in the patient group. Pearson’s correlation coefficient, r, and Spearman correlation coefficient, ρ, were calculated for normal and non-normal distribution data, respectively. Normality was tested using the Shapiro–Wilk test. Correlation coefficients were interpreted as follows: >0.75, good to excellent relationship; 0.50 to 0.75, moderate to good relationship; 0.25 to 0.50, fair relationship; and 0.00 to 0.25, little or no relationship.41 The p-values for each correlation coefficient were calculated.
The CSI score and five metabolites in the two regions were tested at the 5% significance level among three patient groups classified by ICOP and the type of pain, using one-way ANOVA and two-way ANOVA, respectively. Statistical analyses were performed using Statistica version 13 (TIBCO Software Inc. CA, USA).
Results
The levels of neurometabolites and p-values between the patient group and healthy controls are shown in Table 2. Asp/tCr and GSH were significantly higher in the patient group in the ACC (Student’s t-test, Bonferroni-adjusted p-value < 0.005). Patients showed a tendency toward increased GABA/tCr in the ACC, although this effect did not reach significance (p = 0.0388). There was no significant difference between the two groups in any neurometabolite in the thalamus.
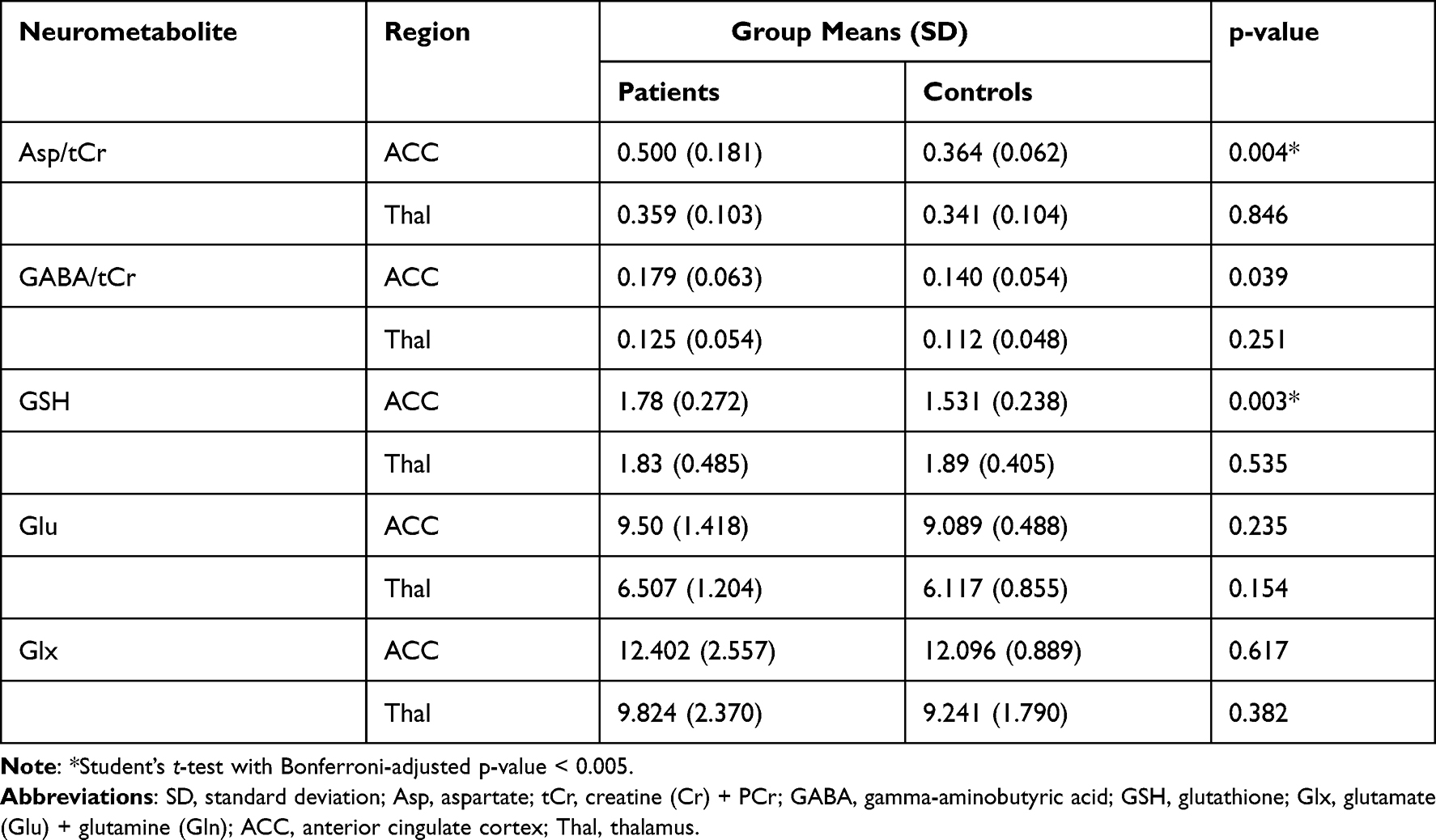 |
Table 2 Neurometabolite Levels |
A moderate to good relationship between Glx and CSI scores was observed in the thalamus (r = 0.556, p = 0.013) (Figure 3A). Fair relationships were found between GABA/tCr (r = −0.467, p = 0.044) and Glx (ρ = 0.420, p = 0.074) in the ACC (Figure 3B and C). GABA/tCr showed little or no relationship in the thalamus (r = −0.248, p = 0.307) (Figure 3D).
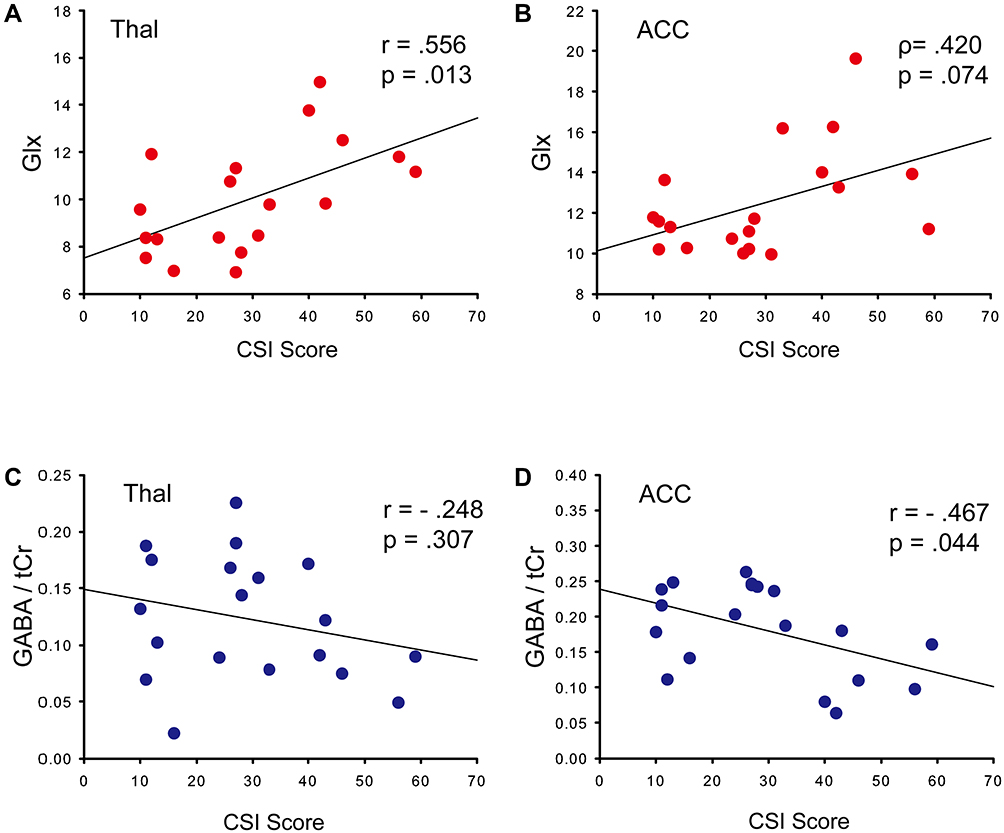 |
Figure 3 Correlation between CSI scores and neurometabolites. Abbreviations: p, Pearson’s correlation coefficient; ρ, Spearman correlation coefficient; Glx, glutamate (Glu) + glutamine (Gln); tCr, creatine (Cr) + PCr; GABA, gamma-aminobutyric acid; ACC, anterior cingulate cortex; Thal, thalamus. Notes: Scatter plot with regression line and correlation between CSI scores and neurometabolite levels. (A) Glx in Thal, (B) Glx in ACC, (C) GABA/tCr in Thal, (D) GABA/tCr in ACC. |
There were no significant differences among the three patient groups classified by ICOP for CSI score (F[2, 16] = 1.508, p = 0.861) and each neurometabolite in the two regions (F[10, 60] = 1.332, p = 0.235), and by the type of pain for CSI score (F[2, 16] = 0.398, p = 0.678) and each neurometabolite in the two regions (F[10, 60] = 0.788, p = 0.641).
Discussion
1H-MRS revealed higher levels of Asp/tCr and GSH in the ACC in chronic orofacial pain patients compared with the levels in healthy controls, but no differences in metabolite levels in the thalamus. Glx levels in the ACC and thalamus indicated a positively moderate to good or fair relationship with CSI scores, whereas GABA/tCr levels in the ACC showed a negatively fair relationship with CSI scores.
Asp is an excitatory amino acid neurotransmitter and a highly selective agonist for NMDA receptor-type glutamate receptors,42 and is also part of a metabolic pathway involved in the production of NAA.43 Although Asp has not been frequently evaluated in 1H-MRS studies of pain, a complex combination of alterations of Asp/Cr, N-acetyl aspartate/Cr, and Gln/Cre in ACC has been reported to differentiate migraine patients from controls.44 These three metabolites act as eventual precursors of Glu.45 Glu is transported into mitochondria via the Asp/Glu carrier, then decarboxylated and converted to Asp.46 This process is an essential component of the malate/Asp NDH shuttle, which links glycolysis and the tricarboxylic acid cycle in brain cells, as well as neurotransmitter labeling from glucose and the formation of the neurotransmitter Glu from Gln Asp aminotransferase in astrocytes and neurons. This shuttle is also essential for transferring the reducing equivalents from glycolysis to the mitochondrial electron transport chain and subsequent oxidative phosphorylation.47 Consequently, increased levels of Asp suggest both neural and glial hyperactivity in ACC. Regarding increases in the levels of excitatory neurotransmitters in ACC, our results are consistent with those of a previous MRS study of chronic pain in miscellaneous pain patients.17
Long-term potentiation of neuronal synapses in ACC is thought to play an important role in the development of CS.48 Short-term plasticity in ACC is also related to CS, and nociceptive signals are transmitted through the thalamic-anterior cingulate pathway.49 The absence of group differences in neurometabolites in the thalamus in the current study suggests the CS may involve reduced activity of this pathway, or that CS was already established in the ACC.
GSH is a major intracellular antioxidant, and its prevalence in glial cells is approximately twice that in neurons.50 The fundamental role of GSH is detoxification of reactive oxygen species, which is critical for maintaining normal functioning of the human brain.51 Excessive stimulation of the NMDA receptor and an influx of calcium ions causes the production of excess free radicals and oxidative stress, which leads to CS and cellular apoptosis.52 GSH provides neuroprotection and modulates the activity of several membrane receptors, including NMDA receptors.53 Previous studies reported that GSH decreased the expression of NMDA receptor subunit GluN1, and inhibited mechanical allodynia and CS in chronic post-ischemia pain model rats.54 A GABA agonist and endomorphin-1 increased the pain threshold and GSH in spinal cord injury model rats.55 Glu-induced GABA release was prolonged by GSH and oxidized GSH in rat hippocampal slices.56 In addition, Asp induced GABA release via the selective activation of NMDA receptors in retina cells.57 In the current study, significantly higher Asp/tCr and a trend toward higher GABA/tCr compared with the levels in controls suggests that excessive activation of NMDA receptors induced by pain causes the production of GSH in astrocytes, maintains oxidative stress, and modulates the activation of NMDA receptors and release of GABA.
The CSI can be used to assess both somatic and emotional complaints that are commonly associated with CS. Few studies have investigated the associations between neurometabolite levels and CSI scores. Aguila et al reported a positive relationship between GABA levels and CSI scores in posterior cingulate cortex in migraine patients,58 whereas a negative correlation between GABA/tCr in the ACC and CSI scores was observed in the current study. In addition, Aguila et al’s research group recently reported that migraine and other chronic pain patients exhibited significantly higher CSI scores than controls, whereas GABA levels in ACC were not significantly different in any of the examined pain groups compared with levels in healthy controls.59 In contrast, the level of GABA in ACC was negatively correlated with other indices associated with chronic pain, such as the intensity of ongoing osteoarthritis pain22 and fibromyalgia,60 and the Crohn’s Disease Activity Index for patients with abdominal pain.40 The current results revealed a positive correlation between Glx and Gln levels in ACC and CSI scores. These findings suggest that an imbalance between GABA and glutamate neural activity may promote the development of CS. Our findings are consistent with those of several previous investigations. In fibromyalgia patients with low levels of GABA, the level of Glx was found to be negatively correlated with clinical pain scores after transcranial direct current stimulation treatment.60 Furthermore, the Glu/GABA ratio was positively associated with anxiety and depressive scores in veterans with chronic pain23 and in chronic back pain patients.21 Inhibitory-excitatory imbalances in the ACC may be related to both the somatosensory and emotional aspects of CS.
The thalamus is an essential relay for trigeminal-nociceptive inputs that are transmitted from the medulla oblongata to the cortices. In the current study, Glx in the thalamus was positively correlated with CSI scores. Conflicting findings have been reported regarding the correlation between Glu and Gln levels and pain intensity. Although the level of Glx in the thalamus of migraine patients was reported to be higher than that in healthy controls, correlation analysis revealed no significant correlation between this neurometabolite and pain intensity during the last pain attack.27 However, a significant positive correlation was found between Glu/Cr of the left thalamus and visual analog scale pain scores in fibromyalgia patients.31 The Glx level was correlated with pain sensitivity in experimental pain stimulation in healthy individuals.61 A previous study reported a strong correlation between nociceptive-related neuronal activity in the thalamus and local field potentials in the ACC in mice.62 Nociceptive thalamocingulate connections are considered to be crucial to the transition from acute nociceptive stimulation to chronic pain conditions. Because the pain duration of our patient group ranged from 0.5 to 25 years, our findings were likely to have reflected the transition to CS.
The same metabolites were not detected in the correlation analysis within patient groups and the comparison between patients and the controls. Asp/tCr and GSH indicated little or no relationship (r < 0.2) with CSI score in the ACC and thalamus. Although Asp/tCr and GSH were elevated in the patient group, they did not fluctuate in conjunction with the severity of central sensitization. Conversely, Glx and GABA/tCr, which were correlated with CSI scores, varied in conjunction with severity, although the patient group did not necessarily show higher values than controls. The main reason the same metabolites were not detected might be related to differences in the method used to search for the relationship between CSI and metabolites, and the method used to test for high and low metabolite levels between the groups.
There were no significant differences among the three groups classified by ICOP and the type of pain for both CSI score and each metabolite. Although the absence of differences might have been caused by the small sample size in the current study, the findings suggested that CS and the levels of associated neurometabolites can fluctuate regardless of classification by diagnosis and the characteristics of pain.
The current study involved several limitations. We did not use spectral editing techniques such as J-difference editing MEGA-PRESS (MEscher-GArwood-Point RESolved Spectroscopy) or multiple quantum coherence editing. Thus, neurometabolites with similar spectral peaks were possibly segregated into imprecise concentrations, including Asp, GSH, GABA, Glu, and Gln. The Asp concentration in the ACC of healthy controls in the current study (2.01 ± 0.327 mM, mean ± SD) was lower than that reported in a previous study using MEGA-PRESS (2.80 ± 0.31 mM).63 Asp could be detected in PRESS, but exhibited weak to moderate correlations with other measures.64 Measurements that separate Asp from other neurometabolites could reveal the role of Asp and imbalance of excitatory and inhibitory neural activity to develop CS in ACC. Regarding GSH, one previous study reported that estimated GSH in ACC was not significantly different between PRESS and MEGA-PRESS using a 3T system.65 In addition, the voxel size in the thalamus was larger than the region of the trigeminal nucleus in size and lower SNR in patients compared with the levels in controls in the ACC, which is likely to have affected the results. Regarding GABA, the results of the current study should be considered as preliminary findings because the measurements made by PRESS have limited accuracy.64 Moreover, medication was not controlled in our patients, and anticonvulsants and antidepressants were frequently used (Table 1). A previous study reported that a selective serotonin reuptake inhibitor caused changes in the levels of GABA in the ACC but not the levels of Glu or Gln.66 Therefore, the possibility that medication influenced the current results cannot be excluded. Moreover, the correlation coefficients were not high. Therefore, the correlation analysis in this study should be considered as exploratory. Finally, in the Japanese version of the CSI, scores represent subclinical (0–29), mild (30–39), moderate (40–49), severe (50–59), and extreme (> 60) states.35 Thus, the current study sample included subclinical patients (Table 1). However, it is possible that the accompanying symptoms associated with chronic pain listed in the questionnaire were not matched to patients with orofacial pain, because the CSI was originally developed for a population of patients with chronic pain in the whole musculoskeletal system. This issue should be examined in more depth in future studies.
Conclusion
Chronic orofacial pain patients exhibited higher Asp/tCr, and GSH in the ACC, suggesting that the activity of excitatory neurons and glial cells is related to the chronicity of emotional-affective aspects of pain. In addition, the results revealed positive relationships between CSI scores and Glx in the thalamus, as well as trends toward a positive relationship with Glx in ACC and a negative relationship with GABA/tCr in the ACC. These findings suggest that neuronal and glial activities and excitatory and inhibitory neurometabolites are related to the chronification of orofacial pain, including CS. The current findings indicate that the combined use of questionnaires and 1H-MRS may increase the accuracy of CS diagnosis.
Acknowledgments
The authors would like to thank the participants in this study. The authors are also grateful to Takashi Namiki for technical assistance in MR data collection, Mark Mikkelsen for usage of Gannet software, MR staff at Ichikawa general hospital of the Tokyo Dental College and the AIC Yaesu Clinic. We thank Benjamin Knight, MSc., from Edanz (https://jp.edanz.com/ac) for editing a draft of this manuscript.
This work was supported by Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Number JP 18K09821.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Baliki MN, Petre B, Torbey S, et al. Corticostriatal functional connectivity predicts transition to chronic back pain. Nat Neurosci. 2012;15(8):1117–1119. doi:10.1038/nn.3153
2. Nijs J, Torres-Cueco R, van Wilgen CP, et al. Applying modern pain neuroscience in clinical practice: criteria for the classification of central sensitization pain. Pain Physician. 2014;17(5):447–457. doi:10.36076/ppj.2014/17/447
3. Woolf CJ. Central sensitization: implications for the diagnosis and treatment of pain. Pain. 2011;152(3 Suppl):S2–S15. doi:10.1016/j.pain.2010.09.030
4. Seifert F, Maihofner C. Functional and structural imaging of pain-induced neuroplasticity. Curr Opin Anaesthesiol. 2011;24(5):515–523. doi:10.1097/ACO.0b013e32834a1079
5. Larsson M, Broman J. Synaptic plasticity and pain: role of ionotropic glutamate receptors. Neuroscientist. 2011;17(3):256–273. doi:10.1177/1073858409349913
6. Zhuo M. Cortical plasticity as synaptic mechanism for chronic pain. J Neural Transm. 2020;127(4):567–573. doi:10.1007/s00702-019-02071-3
7. Li C, Lei Y, Tian Y, et al. The etiological contribution of GABAergic plasticity to the pathogenesis of neuropathic pain. Mol Pain. 2019;15:1744806919847366. doi:10.1177/1744806919847366
8. Hossain MZ, Unno S, Ando H, Masuda Y, Kitagawa J. Neuron-glia crosstalk and neuropathic pain: involvement in the modulation of motor activity in the orofacial region. Int J Mol Sci. 2017;18(10):2051. doi:10.3390/ijms18102051
9. Treede RD. The role of quantitative sensory testing in the prediction of chronic pain. Pain. 2019;160 Suppl 1:S66–S69. doi:10.1097/j.pain.0000000000001544
10. Mayer TG, Neblett R, Cohen H, et al. The development and psychometric validation of the central sensitization inventory. Pain Pract. 2012;12(4):276–285. doi:10.1111/j.1533-2500.2011.00493.x
11. Latremoliere A, Woolf CJ. Central sensitization: a generator of pain hypersensitivity by central neural plasticity. J Pain. 2009;10(9):895–926. doi:10.1016/j.jpain.2009.06.012
12. Lin Q, Peng YB, Willis WD. Inhibition of primate spinothalamic tract neurons by spinal glycine and GABA is reduced during central sensitization. J Neurophysiol. 1996;76(2):1005–1014. doi:10.1152/jn.1996.76.2.1005
13. Besson M, Matthey A, Daali Y, et al. GABAergic modulation in central sensitization in humans: a randomized placebo-controlled pharmacokinetic-pharmacodynamic study comparing clobazam with clonazepam in healthy volunteers. Pain. 2015;156(3):397–404. doi:10.1097/01.j.pain.0000460331.33385.e8
14. Vogt BA, Pandya DN, Rosene DL. Cingulate cortex of the rhesus monkey: i. Cytoarchitecture and thalamic afferents. J Comp Neurol. 1987;262(2):256–270. doi:10.1002/cne.902620207
15. Magon S, May A, Stankewitz A, et al. Morphological abnormalities of thalamic subnuclei in migraine: a multicenter MRI Study at 3 Tesla. J Neurosci. 2015;35(40):13800–13806. doi:10.1523/JNEUROSCI.2154-15.2015
16. Peek AL, Rebbeck T, Puts NA, Watson J, Aguila MR, Leaver AM. Brain GABA and glutamate levels across pain conditions: a systematic literature review and meta-analysis of 1H-MRS studies using the MRS-Q quality assessment tool. Neuroimage. 2020;210:116532. doi:10.1016/j.neuroimage.2020.116532
17. Ito T, Tanaka-Mizuno S, Iwashita N, et al. Proton magnetic resonance spectroscopy assessment of metabolite status of the anterior cingulate cortex in chronic pain patients and healthy controls. J Pain Res. 2017;10:287–293. doi:10.2147/JPR.S123403
18. Widerstrom-Noga E, Pattany PM, Cruz-Almeida Y, et al. Metabolite concentrations in the anterior cingulate cortex predict high neuropathic pain impact after spinal cord injury. Pain. 2013;154(2):204–212. doi:10.1016/j.pain.2012.07.022
19. Gussew A, Rzanny R, Gullmar D, Scholle HC, Reichenbach JR. 1H-MR spectroscopic detection of metabolic changes in pain processing brain regions in the presence of non-specific chronic low back pain. Neuroimage. 2011;54(2):1315–1323. doi:10.1016/j.neuroimage.2010.09.039
20. Grachev ID, Fredrickson BE, Apkarian VA. Abnormal brain chemistry in chronic back pain: an in vivo proton magnetic resonance spectroscopy study. Pain. 2000;89(1):7–18. doi:10.1016/S0304-3959(00)00340-7
21. Janetzki L, Gussew A, Malessa R, et al. [Cerebral metabolic changes and chronic back pain: study taking into consideration clinical and psychological parameters]. Schmerz. 2016;30(2):134–140. German. doi:10.1007/s00482-015-0082-5
22. Reckziegel D, Raschke F, Cottam WJ, Auer DP. Cingulate GABA levels inversely correlate with the intensity of ongoing chronic knee osteoarthritis pain. Mol Pain. 2016;12:174480691665069. doi:10.1177/1744806916650690
23. Legarreta MD, Sheth C, Prescot AP, Renshaw PF, McGlade EC, Yurgelun-Todd DA. An exploratory proton MRS examination of gamma-aminobutyric acid, glutamate, and glutamine and their relationship to affective aspects of chronic pain. Neurosci Res. 2021;163:10–17. doi:10.1016/j.neures.2020.03.002
24. Niddam DM, Lai KL, Tsai SY, et al. Neurochemical changes in the medial wall of the brain in chronic migraine. Brain. 2018;141(2):377–390. doi:10.1093/brain/awx331
25. Foerster BR, Petrou M, Edden RA, et al. Reduced insular gamma-aminobutyric acid in fibromyalgia. Arthritis Rheum. 2012;64(2):579–583. doi:10.1002/art.33339
26. Harper DE, Ichesco E, Schrepf A, et al. Relationships between brain metabolite levels, functional connectivity, and negative mood in urologic chronic pelvic pain syndrome patients compared to controls: a MAPP research network study. Neuroimage Clin. 2018;17:570–578. doi:10.1016/j.nicl.2017.11.014
27. Bathel A, Schweizer L, Stude P, et al. Increased thalamic glutamate/glutamine levels in migraineurs. J Headache Pain. 2018;19(1):55. doi:10.1186/s10194-018-0885-8
28. Henderson LA, Peck CC, Petersen ET, et al. Chronic pain: lost inhibition? J Neurosci. 2013;33(17):7574–7582. doi:10.1523/JNEUROSCI.0174-13.2013
29. Gustin SM, Wrigley PJ, Youssef AM, et al. Thalamic activity and biochemical changes in individuals with neuropathic pain after spinal cord injury. Pain. 2014;155(5):1027–1036. doi:10.1016/j.pain.2014.02.008
30. Valdes M, Collado A, Bargallo N, et al. Increased glutamate/glutamine compounds in the brains of patients with fibromyalgia: a magnetic resonance spectroscopy study. Arthritis Rheum. 2010;62(6):1829–1836. doi:10.1002/art.27430
31. Feraco P, Bacci A, Pedrabissi F, et al. Metabolic abnormalities in pain-processing regions of patients with fibromyalgia: a 3T MR spectroscopy study. AJNR Am J Neuroradiol. 2011;32(9):1585–1590. doi:10.3174/ajnr.A2550
32. Terumitsu M, Seo K, Matsuzawa H, Yamazaki M, Kwee IL, Nakada T. Morphologic evaluation of the inferior alveolar nerve in patients with sensory disorders by high-resolution 3D volume rendering magnetic resonance neurography on a 3.0-T system. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2011;111(1):95–102. doi:10.1016/j.tripleo.2010.09.002
33. Terumitsu M, Matsuzawa H, Seo K, et al. High-contrast high-resolution imaging of posttraumatic mandibular nerve by 3DAC-PROPELLER magnetic resonance imaging: correlation with the severity of sensory disturbance. Oral Surg Oral Med Oral Pathol Oral Radiol. 2017;124(1):85–94. doi:10.1016/j.oooo.2017.02.017
34. Orofacial Pain Classification Committee. International classification of orofacial pain, 1st edition (ICOP). Cephalalgia. 2020;40(2):129–221. doi:10.1177/0333102419893823
35. Tanaka K, Nishigami T, Mibu A, et al. Validation of the Japanese version of the Central Sensitization Inventory in patients with musculoskeletal disorders. PLoS One. 2017;12(12):e0188719. doi:10.1371/journal.pone.0188719
36. Provencher SW. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn Reson Med. 1993;30(6):672–679. doi:10.1002/mrm.1910300604
37. Harris AD, Puts NA, Edden RA. Tissue correction for GABA-edited MRS: considerations of voxel composition, tissue segmentation, and tissue relaxations. J Magn Reson Imaging. 2015;42(5):1431–1440. doi:10.1002/jmri.24903
38. Xin L, Mekle R, Fournier M, et al. Genetic polymorphism associated prefrontal glutathione and its coupling with brain glutamate and peripheral redox status in early psychosis. Schizophr Bull. 2016;42(5):1185–1196. doi:10.1093/schbul/sbw038
39. Wilson M, Andronesi O, Barker PB, et al. Methodological consensus on clinical proton MRS of the brain: review and recommendations. Magn Reson Med. 2019;82(2):527–550. doi:10.1002/mrm.27742
40. Kun L, Song W, Tang R, et al. Neurotransmitter alterations in the anterior cingulate cortex in Crohn’s disease patients with abdominal pain: a preliminary MR spectroscopy study. Neuroimage Clin. 2018;20:793–799. doi:10.1016/j.nicl.2018.09.008
41. Portney LG. Foundations of Clinical Research: Applications to Practice.
42. Patneau DK, Mayer ML. Structure-activity relationships for amino acid transmitter candidates acting at N-methyl-D-aspartate and quisqualate receptors. J Neurosci. 1990;10(7):2385–2399. doi:10.1523/JNEUROSCI.10-07-02385.1990
43. Bates TE, Strangward M, Keelan J, Davey GP, Munro PM, Clark JB. Inhibition of N-acetylaspartate production: implications for 1H MRS studies in vivo. Neuroreport. 1996;7(8):1397–1400. doi:10.1097/00001756-199605310-00014
44. Becerra L, Veggeberg R, Prescot A, et al. A “complex” of brain metabolites distinguish altered chemistry in the cingulate cortex of episodic migraine patients. Neuroimage Clin. 2016;11:588–594. doi:10.1016/j.nicl.2016.03.020
45. Albrecht J, Sidoryk-Wegrzynowicz M, Zielinska M, Aschner M. Roles of glutamine in neurotransmission. Neuron Glia Biol. 2010;6(4):263–276. doi:10.1017/S1740925X11000093
46. Palmieri L, Pardo B, Lasorsa FM, et al. Citrin and aralar1 are Ca(2+)-stimulated aspartate/glutamate transporters in mitochondria. EMBO J. 2001;20(18):5060–5069. doi:10.1093/emboj/20.18.5060
47. McKenna MC, Hopkins IB, Lindauer SL, Bamford P. Aspartate aminotransferase in synaptic and nonsynaptic mitochondria: differential effect of compounds that influence transient hetero-enzyme complex (metabolon) formation. Neurochem Int. 2006;48(6–7):629–636. doi:10.1016/j.neuint.2005.11.018
48. Zhuo M. A synaptic model for pain: long-term potentiation in the anterior cingulate cortex. Mol Cells. 2007;23(3):259–271.
49. Shyu BC, Vogt BA. Short-term synaptic plasticity in the nociceptive thalamic-anterior cingulate pathway. Mol Pain. 2009;5:51. doi:10.1186/1744-8069-5-51
50. Rice ME, Russo-Menna I. Differential compartmentalization of brain ascorbate and glutathione between neurons and glia. Neuroscience. 1998;82(4):1213–1223. doi:10.1016/S0306-4522(97)00347-3
51. Dringen R. Metabolism and functions of glutathione in brain. Prog Neurobiol. 2000;62(6):649–671. doi:10.1016/s0301-0082(99)00060-x
52. Xu B, Xu ZF, Deng Y, Liu W, Yang HB, Wei YG. Protective effects of MK-801 on methylmercury-induced neuronal injury in rat cerebral cortex: involvement of oxidative stress and glutamate metabolism dysfunction. Toxicology. 2012;300(3):112–120. doi:10.1016/j.tox.2012.06.006
53. Sucher NJ, Lipton SA. Redox modulatory site of the NMDA receptor-channel complex: regulation by oxidized glutathione. J Neurosci Res. 1991;30(3):582–591. doi:10.1002/jnr.490300316
54. Yeo J, Jung H, Lee H. Effects of glutathione on mechanical allodynia and central sensitization in chronic postischemic pain rats. Pain Res Manag. 2017;2017:7394626. doi:10.1155/2017/7394626
55. Hosseini M, Karami Z, Yousefifard M, Janzadeh A, Zamani E, Nasirinezhad F. Simultaneous intrathecal injection of muscimol and endomorphin-1 alleviates neuropathic pain in rat model of spinal cord injury. Brain Behav. 2020;10(5):e01576. doi:10.1002/brb3.1576
56. Janaky R, Varga V, Oja SS, Saransaari P. Release of [3H]GABA evoked by glutamate agonists from hippocampal slices: effects of dithiothreitol and glutathione. Neurochem Int. 1994;24(6):575–582. doi:10.1016/0197-0186(94)90010-8
57. Kubrusly RC, de Mello MC, de Mello FG. Aspartate as a selective NMDA receptor agonist in cultured cells from the avian retina. Neurochem Int. 1998;32(1):47–52. doi:10.1016/S0197-0186(97)00051-X
58. Aguila MR, Rebbeck T, Leaver AM, et al. The association between clinical characteristics of migraine and brain GABA levels: an exploratory study. J Pain. 2016;17(10):1058–1067. doi:10.1016/j.jpain.2016.06.008
59. Peek AL, Leaver AM, Foster S, et al. Increased GABA+ in people with migraine, headache, and pain conditions- a potential marker of pain. J Pain. 2021;22:1631–1645.
60. Foerster BR, Nascimento TD, DeBoer M, et al. Excitatory and inhibitory brain metabolites as targets of motor cortex transcranial direct current stimulation therapy and predictors of its efficacy in fibromyalgia. Arthritis Rheumatol. 2015;67(2):576–581. doi:10.1002/art.38945
61. Zunhammer M, Schweizer LM, Witte V, Harris RE, Bingel U, Schmidt-Wilcke T. Combined glutamate and glutamine levels in pain-processing brain regions are associated with individual pain sensitivity. Pain. 2016;157(10):2248–2256. doi:10.1097/j.pain.0000000000000634
62. Lee CM, Chang WC, Chang KB, Shyu BC. Synaptic organization and input-specific short-term plasticity in anterior cingulate cortical neurons with intact thalamic inputs. Eur J Neurosci. 2007;25(9):2847–2861. doi:10.1111/j.1460-9568.2007.05485.x
63. Menshchikov P, Manzhurtsev A, Ublinskiy M, Akhadov T, Semenova N. T2 measurement and quantification of cerebral white and gray matter aspartate concentrations in vivo at 3T: a MEGA-PRESS study. Magn Reson Med. 2019;82(1):11–20. doi:10.1002/mrm.27700
64. Dhamala E, Abdelkefi I, Nguyen M, Hennessy TJ, Nadeau H, Near J. Validation of in vivo MRS measures of metabolite concentrations in the human brain. NMR Biomed. 2019;32(3):e4058. doi:10.1002/nbm.4058
65. Sanaei Nezhad F, Anton A, Parkes LM, Deakin B, Williams SR. Quantification of glutathione in the human brain by MR spectroscopy at 3 Tesla: comparison of PRESS and MEGA-PRESS. Magn Reson Med. 2017;78(4):1257–1266. doi:10.1002/mrm.26532
66. Brennan BP, Admon R, Perriello C, et al. Acute change in anterior cingulate cortex GABA, but not glutamine/glutamate, mediates antidepressant response to citalopram. Psychiatry Res Neuroimaging. 2017;269:9–16. doi:10.1016/j.pscychresns.2017.08.009
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.