")
Back to Archived Journals » Open Access Animal Physiology » Volume 8
Negative correlation between gestation and sleep durations in mammals
Authors Gonfalone A
Received 8 April 2016
Accepted for publication 29 April 2016
Published 1 September 2016 Volume 2016:8 Pages 1—7
DOI https://doi.org/10.2147/OAAP.S110108
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Peter Koulen
Alain A Gonfalone
Microgravity Department, European Space Agency, France
Abstract: Many studies of sleep have tempted to establish a relation between environmental, ecological, and physiological variables and sleep duration. This article establishes unambiguously that there is a negative correlation between gestation and sleep durations in mammals. Gestation offers the advantage that one species is characterized by a precise value of its gestation time, with relative small variations, and it is a quantity that can easily be measured. The duration of gestation is a parameter which differs from other parameters that have been used for comparison, such as weight, brain size, and predatory danger. The review of all relevant research articles clearly demonstrates that those mammals, which have longer gestation period, experience less sleep. Within the class of mammals, precocial mammals, capable of moving at birth, show a relatively short amount of rapid eye movement (REM) sleep (a unique phase of mammalian sleep) that does not vary much after birth, while altricial species with a short gestation that are relatively more dependent at birth adjust their amount of REM sleep after birth and show longer sleep duration in-line with the present findings. The essential data on which this result is obtained has been deduced from an extensive and attentive review of previously published articles on mammalian sleep and mammalian gestation, among others. In answering the question “Why does species with a shorter gestation period need more sleep?”, many similarities in behavior between the fetal state and REM sleep state have been identified. Even if at birth all sensory organs are fully developed, their functions have not been exercised, and it is proposed that the amount of REM sleep may be the time needed to strengthen adaptation of the senses to the external world and its components, such as light, sound, contact, taste, odors, and particularly cold and gravity. The assumption proposed is that REM sleep is needed to compensate for the short gestation. It is a period for the brain to reorder and rearrange all sensations accumulated during wake and prepare for the following wake period, and that REM sleep is a carry over of the fetal state.
Keywords: gravity, REM sleep, mammal, brain, fetus, neuron
Introduction
Sleep is the object of sustained scientific research; some of its functions are unclear, and although many features have been discovered and explained, numerous questions remain. Currently, it is thought that all species throughout the animal kingdom, have periods of reduced activity or quiescence that can be equated to a state of sleep.1 The sleep of mammals, including the human species, has been the subject of numerous analyses. Mammalian sleep involves complex mechanisms, some of which have been identified and are well understood. Siegel2 quotes that “Across mammals, the amount and nature of sleep are correlated with age, body size and ecological variables, such as whether the animals live in a terrestrial or an aquatic environment, their diet and the safety of their sleeping site.” This encapsulates in one sentence that all of the parameters are thought to influence sleep in mammals.
A comparison of the durations of mammalian sleep periods and their weights led to the conclusion that there are differences between carnivores, herbivores, and omnivores with regard to correlations between weight and sleep duration.2
Predation risk can also influence sleep duration;3 however, the degree of risk was determined in the cited study using a number of poorly defined, nonquantifiable factors that are not established in the literature. Lighter animals are at greater risk of predation, and this is demonstrated in the article; however, this could indicate that the observed correlation between predation risk and sleep could be attributed to weight differences between species.
Other parameters studied in relation to sleep duration include life span, heart rate, and brain weight. All these constitutive variables are interrelated and may be a consequence of the same underlying relationship, as noted by Lesku et al.4 Sleep duration can also be understood in terms of energy expenditure. For example, it has been stated that “A platypus may sleep more (fourteen hours), perhaps because a frugal meal of crustaceans is enough to bring him a sufficient dose of calories.”5 The time required to forage for food may also have an effect, since herbivores generally require more time to collect sufficient vegetation to meet their energy requirements.
Nonetheless, close examination of the results reveals a trend for shorter sleep duration in heavier species, and the influence of weight, or gravity, on rapid eye movement (REM) sleep was confirmed by Gonfalone and Jha,6 who show in their article that REM sleep can only occur if the sensation of weight has been reduced.
The duration of mammalian sleep varies widely, and there remains no clear explanation of the fact that elephants sleep for ~3 hours and opossums sleep for 18 hours, each day.7 Mammals exhibit a number of sleep phases, including paradoxical sleep, or its equivalent, REM sleep. REM sleep represents ~20%–25% of total sleep time in most species and is a characteristic of the sleep of adult mammals. REM sleep usually occurs after a period of slow wave sleep and is characterized by more rapid waves indicating brain activity, while the sleeper is profoundly asleep and disconnected from the external world.8,9
This article demonstrates an unambiguous relationship between gestation and REM sleep durations for a large number of terrestrial mammal species. Furthermore, similarities between the fetal state and the REM sleep state are highlighted, and a possible explanation for the relationship is provided, associating REM sleep with the need for adaptation to ecological variables, such as cold and gravity. In addition to light, sound, contact, odor, and taste, which can be stimulated during gestation, these two variables, such as cold and gravity, do not affect the fetus, which develops in a constant thermal environment and in a state of weightlessness.
Zepelin et al1 in their article noted that “There are strikingly strong correlations of REM sleep quotas with degree of maturity at birth. REM sleep may be a carryover from fetal life.” The present study demonstrates the validity of this statement and pursues it further to confirm the indisputable similarity between gestation and the REM sleep state.
Methods
Numerous scientific reports related to sleep and REM sleep duration in mammals have been published in recent years.1–11 In this article, all these published data are carefully reviewed and reinterpreted, and a new hypothesis is proposed. Gestation and corresponding REM sleep durations were collated for 79 mammalian species (Table 1). It seems that gestation has not often been compared with sleep amount before and when it has and has never been appreciated as the important parameter,1–4 rather other parameters such as weight and lifespan for instance, have been considered in an analysis of REM sleep duration. Data are shown in Table 1, and gestation duration and REM sleep duration are compared subsequently using a correlation method. The sleep data are taken from Gonfalone and Jha,6 and the gestation data are taken from the very complete study of Lesku et al.4
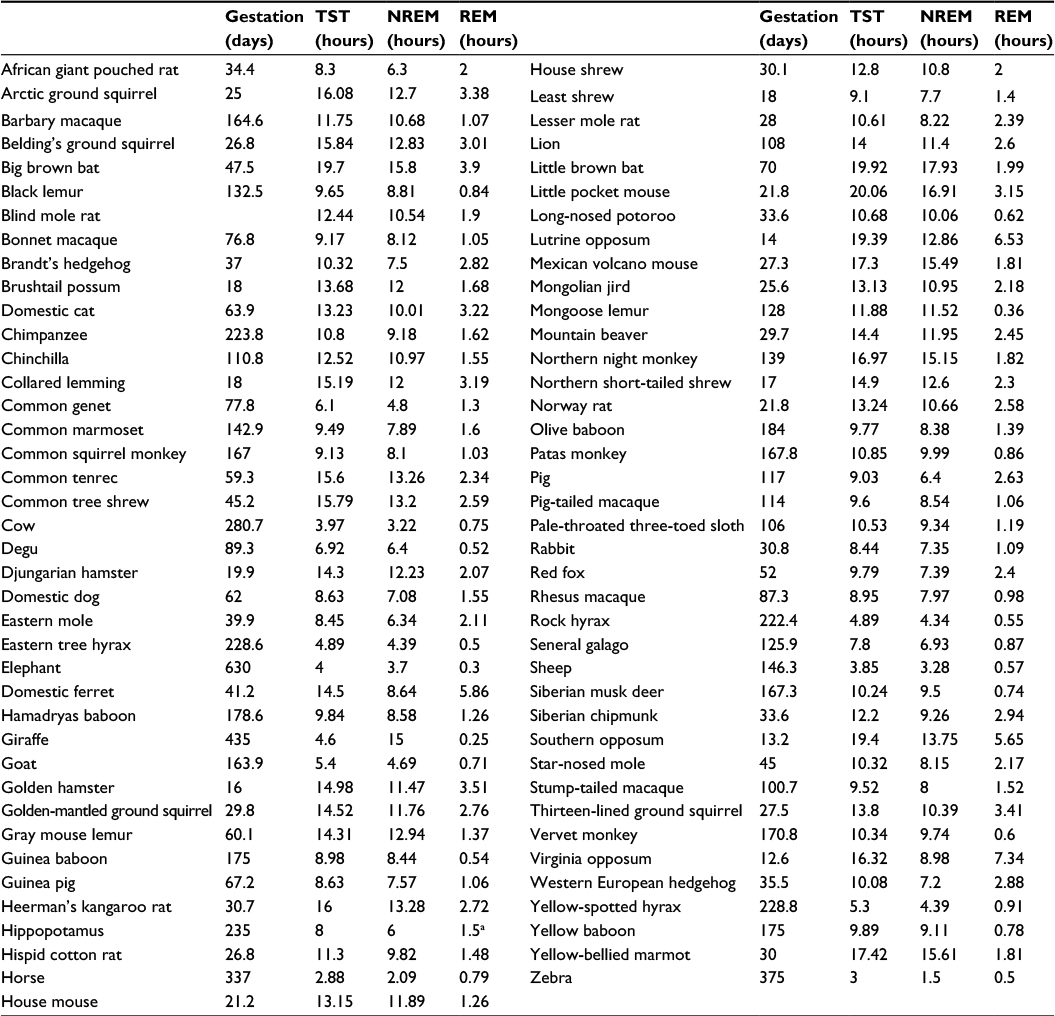 | Table 1 REM sleep and gestation data from 79 species of mammals Notes: Data are derived from the reports of Madan and Jha39 and Gonfalone and Jha,6 with some additional data from the lion, the giraffe,25 and the platypus. Gestation times are reported in the article by Lesku5 and are widely available on the Internet. aREM sleep duration was estimated as 25% of NREM sleep duration. Adapted from Neurosci Bull. Sleep alterations in mammals: did aquatic conditions inhibit rapid eye movement sleep? 2012;28:746–758. Madan V, Jha SK. © Shanghai Institutes for Biological Sciences, CAS and Springer-Verlag Berlin Heiedelberg 2012. With permission of Springer and SK Jha.39 Copyright ©2015. Dove Medical Press. Adapted from Gonfalone A, Jha SK. The influence of gravity on REM sleep. Open Access Anim Physiol. 2015;7:65–72.6 Abbreviations: NREM, nonrapid eye movement; REM, rapid eye movement; TST, total sleep time. |
In that study, Lesku et al considered the weight of the brain at birth, compared with its weight in the adult; a trend was observed, but the data are not conclusive, are interpreted with an emphasis on phylogenetic links between species, and appear to invalidate the majority of the conclusions of previous research articles. However, this report contains an extensive data set related to both gestation and REM sleep.
Brain maturity and subsequent adaptation of a newborn animal to the external world depend on the length of gestation. In addition, adaptation to the external world and its ecological variables are the keys to sleep duration.
Gestation period is a relatively invariable, species-specific parameter,12,13 as opposed to, for instance, weight. Moreover, gestation duration can be measured accurately and is a permanent characteristic of a given species. Where there are variations, these are in the order of only a few percent. This is much less than intraspecies variations in weight, which in dogs, eg, can vary by a factor of 20 (between 1.5 kg and 30 kg).
Excluded species and justification
All aquatic mammals were excluded from the analysis because their sleep is unihemispheric,14 eg, dolphins shut down only half of their brains,15 along with the opposite eye, when sleeping. These species deserve additional research and, potentially, a different interpretation of their REM sleep.
Two other groups of animal species have sleep durations that differ considerably from the majority of mammals: the armadillo family (eg, nine-banded, large hairy, and giant), which are terrestrial mammals, and the seals (eg, cape fur, northern fur, grey, caspian seals, and southern sea lion), which are semiaquatic.16,17 A very particular and unusual trait in common between these two groups is that their gestation is delayed;18,19 embryo development does not proceed immediately after fecundation but is delayed until a few months before the time when the birth conditions are favorable, eg, until spring or summer, when food availability and temperature are adequate. In addition, eared seals have been noted for an absence of REM sleep when in water, while they do experience REM sleep when on shore. A possible explanation for this phenomenon is that seals are subject to their own weight on shore, where they do have REM sleep and behave more like fish, which have no REM sleep when in water.3,20 Furthermore, these animals move by wriggling on their bellies and not by using their front limbs. Although this is not their main mode of locomotion, the nine-banded armadillo is often observed moving on its hind limbs,21 which may require additional exertion of effort to maintain balance and an effort to oppose gravity, leading to a need for a longer rest period (ie, sleep). However, the definitive explanation for these particular behaviors has yet to be identified.
The other species excluded from the analysis is humans. The sleep duration of humans is much longer than that would be expected for a species with a 9-month gestation. A possible explanation for this may be that humans are the only true, permanently two-legged species; therefore, the waking behavior of humans differs from four-legged mammal species. The anomalous sleep patterns observed in humans may, therefore, be due to the permanent effort required to support their weight and maintain their balance and posture, opposing the effects of gravity. Hence, locomotion on two or four limbs may influence the need for sleep.
Results
Gonfalone and Jha6 in their article compared the sleep duration of ~80 mammals and revealed a clear trend for the association of short sleep durations with increased weight and longer gestation periods. In the present study, data from a number of additional species, including the giraffe (a short sleeper),22 the lion,23 and the platypus (a long sleeper),5 have been added, while some were excluded (as explained in the “Methods” section). The data concerning the REM sleep of these mammals were compared with their gestation time.
The logarithm of gestation length (days) was plotted against the logarithm of REM sleep duration (minutes) (Figure 1). This mode of presentation amplifies changes in values close to zero and moderate changes to large numbers and highlights relative variations. A trend line was added to illustrate the negative correlation between the durations of gestation and REM sleep. The spreadsheet software was used to calculate the correlation from the data set. The straight line indicates the negative correlation between coordinates of the data point.
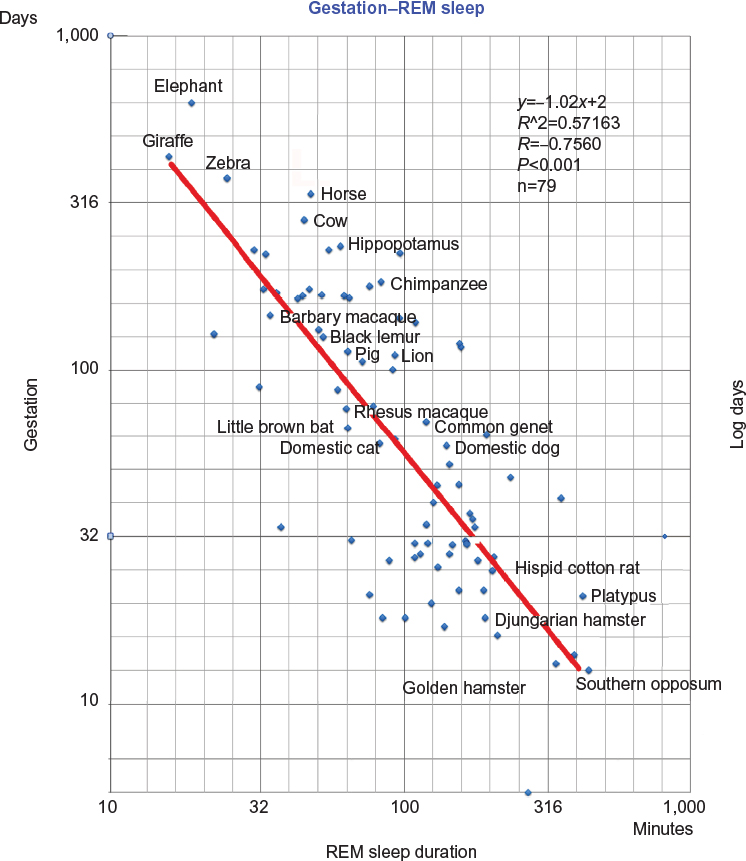 | Figure 1 Negative correlation between gestation and REM sleep durations from 79 mammalian species. Notes: Common logarithmic transformations were used for gestation period and paradoxical sleep values. Correlation coefficient is −0.7560. The strength of the correlation found is higher than the value reported in Zepelin et al1 and Capellini et al.7 Abbreviation: REM, rapid eye movement. |
Although Figure 1 contains data from ~80 mammalian species, it was obtained by excluding a small number of species for which the data did not fit the general trend (as explained in the “Methods” section); however, these exceptions do not invalidate the general trend.
In Figure 1, the platypus, an egg-laying mammal with a short gestation period, demonstrates a sleep duration consistent with that of other mammals. The sleep of the platypus was the subject of specific study, which concluded that “Our findings indicate that REM sleep may have been present in large amounts in the first mammals and suggest that it may have evolved in pre-mammalian reptiles.”5 However, taking into account gestation length places the sleep duration of the platypus in-line with that of other mammals indicating the validity of the results presented in this study.
Discussion
The plot in Figure 1 shows unambiguously that there is a negative correlation between the durations of REM sleep and gestation in mammals. This raises the question “Why does a species with a short gestation period need a longer period of sleep?”. Here, a new hypothesis is proposed that the development of the functions of the sensory organs begins at birth but is consolidated during REM sleep.
Sense organs become morphologically mature before birth.23 Although a neonate possesses all the necessary sense organs that have developed during gestation, no stimulation has been consistently applied to these organs, and their functions essentially begin at birth, with exposure to light, sound, air, cold, and gravity.
During gestation, under special circumstances and with sufficiently strong stimuli, sense organs detecting light, sound, tastes, and touch are active;24 however, stimulation by cold or gravity is not possible. During gestation, the embryo exists under conditions of low gravity and constant temperature within the womb, a state that persists for a period of months in some species. It is generally assumed that the fetus floats almost weightlessly in its environment, hence gravitational stimuli are not present in utero.
At birth, the brain is, in principle, prepared to react with these new stimuli. However, gestation does not lead to equal levels of brain maturity at birth for all mammalian species, with some producing precocial, which can move at birth, and other altricial neonates. In the minutes following the birth of a baby giraffe, it is striking to see that the first activity is to rise on to its four limbs, before even looking for milk.25 If the brain has developed sufficiently, adaptation to stimuli in the external world, particularly cold and gravity, will occur rapidly, particularly in species with long gestation periods, despite the fact that the fetus has not been exposed to cold and gravity before birth. In contrast, a short gestation period does not prepare the newborn to face cold and gravity at birth, and most probably, adaptation to the external world takes place during REM sleep episodes.
In this case, REM sleep may be conceived as an additional period for the development of brain capabilities required for adaptation to certain ecological variables in the external world.26 Surprisingly, this adaptation appears to continue throughout life. This may be because adaptations not acquiring before birth need to be strengthened or revised periodically. As mentioned by Siegel,27 sleep is a period of inactivity but adaptive inactivity. In mammals, neurogenesis ceases within a month of birth, “So why not conceive that certain genetic programs may be reinforced periodically.”28
This hypothesis certainly does not provide a full explanation of the need for REM sleep, but similarities between the behavior of the fetus and some reactions observed during REM sleep suggest a relationship. The fetus exhibits REMs and jerky motions and also has penile erections,29,30 all things which also occur during REM sleep.31,32 Body temperature is not well regulated during REM sleep, and the sensation of weight is strongly reduced, as in the fetal state. Anecdotally, the sleeping position most frequent among humans is the “fetal position”. In addition, astronauts experiencing weightlessness sleep in a position very similar to the fetal position.33 Moreover, it has been stated that “Suppression of REM sleep during early development can produce long-lasting behavioral as well as brain morphological and biochemical abnormalities in later life.”34
As mentioned earlier, there is a correlation between gestation duration and animal weight. In a study that scaled the gestation periods of a sample of 429 mammals against their weight, the following two slopes were observed: one for those species that give birth to altricial neonates and a second for those species with precocial neonates.11 Precocial species tend to be heavier, in agreement with the plot in Figure 1, which shows the short REM sleep durations for heavier species, such as the elephant and the cow. The altricial neonate shows a large amount of REM sleep at birth, but the amount of REM sleep diminishes to reach the adult value within weeks.1 It is supposed that given all of the sensory information a neonate can take in, it must begin to develop its processing skills and learn how to use the incoming sensory information to effectively act on its environments, such as light, cold, and gravity.
In a recent publication, Herculano-Houzel35 related the duration of sleep with the amount of specific neurons in the brain. The results corroborate those presented in the present article and indicate that brain size and brain maturation play a role in the duration of sleep.
Another idea that should be explored is that the amount of waking time may be related to the quantity of stimulations and sensations that the brain can acquire before reaching saturation. During the course of a typical day, the brain is constantly submitted to external and internal sensations, and each of them leaves a trace; at night, the brain needs to reorder and clear all of these traces.36 The present findings have their limitations: as described in the “Excluded species and justification” section, many species have REM sleep durations that do not fit with the correlation established in Figure 1, particularly the human species. Furthermore, this article concerns the gestation of mammals and their REM sleep. It cannot be applied easily to other classes of vertebrates, such as fishes, reptiles, amphibians, and birds, where the embryo usually develops in an egg.
Another questionable conclusion of this study is that REM sleep is “needed” all life long, even if its amount changes, and it poses the question of a permanent need for adaptation to the external world. At present, there is no simple answer.
Conclusion
A number of previous propositions for factors that may play a role in mammalian sleep duration have been presented in research articles, and clearly, none can be ignored or rejected completely as potential contributors. In addition, it must be noted that this article does not propose a unique explanation for REM sleep duration in mammalian species, rather it identifies a trend in ~80 species of mammals. The most likely explanation for the observation presented in this article is that during REM sleep, cerebral activity takes place to consolidate adaptation to the external world.
This assumption is somehow indirectly supported by previous work demonstrating that REM sleep deprivation in neonate rats has negative effects on their development.
Mirmiran et al34 quote “We conclude, therefore, that chronic clomipramine-induced suppression of AS (REM sleep) during early postnatal life …results in a characteristic ‘syndrome’ consisting of specific behavioral, neurophysiological and structural brain abnormalities.” A more positive confirmation may come from space: astronauts do not feel gravity, and their external world misses this component, while other components are still present, such as light and sound. As a result of this adaptation to a “truncated” environment, their REM sleep is reduced.37,38 Experiments submitting rats to an environment of hypergravity are planned, and it is expected that their REM sleep will be longer.
There are strong arguments to support the assertion that REM sleep is a carryover from the fetal state; however, further investigation is needed. Beyond this assertion, there are clearly many questions requiring further study to fully interpret REM sleep. It is possible that there is no single explanation for the amount of sleep required by different species, as a combination of many parameters may influence this phenomenon.
Acknowledgment
Alain Gonfalone is former staff of the European Space Agency.
Disclosure
The author reports no conflicts of interest in this work.
References
Zepelin H, Siegel JM, Tobler I. Mammalian sleep. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practice of Sleep Medicine. Philadelphia, PA: WB Saunders Company; 2005:91–100. | ||
Siegel JM. Clues to the functions of mammalian sleep. Nature. 2005;437:1264–1271. | ||
Allison T, Cicchetti DV. Sleep in mammals: ecological and constitutional correlates. Science. 1976;194:732–734. | ||
Lesku JA, Roth TC, Rattenborg NC, Amlaner CJ, Lima SL. Phylogenetics and the correlates of mammalian sleep: a reappraisal. Sleep Med Rev. 2008;12:229–244. | ||
Siegel JM, Manger PR, Nienhuis R, Fahringer HM, Shalita T, Pettigrew JD. Sleep in the platypus. Neuroscience. 1999;91(1):391–400. | ||
Gonfalone A, Jha SK. The influence of gravity on REM sleep. Open Access Anim Physiol. 2015;7:65–72. | ||
Capellini I, Barton RA, McNamara P, Preston BT, Nunn CL. Phylogenetic analysis of the ecology and evolution of mammalian sleep. Evolution. 2008;62:1764–1776. | ||
Jouvet M. The Paradox of Sleep, the Story of Dreaming. Le Sommeil et le Rêve. Cambridge, MA: MIT Press; 1999. | ||
Carskadon MA, Dement WC. Monitoring and staging human sleep. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practice of Sleep Medicine. 5th ed. St. Louis, MO: Elsevier Saunders; 2011:16–26. | ||
Campbell SS, Tobler I. Animal sleep: a review of sleep duration across phylogeny. Neurosci Biobehav Rev. 1984;8(3):269–300. | ||
Zepelin H, Rechtschaffen A. Mammalian sleep, longevity, and energy metabolism. Brain Behav Evol. 1974;10(6):425–470. | ||
Martin RD, Genoud M, Hemelrijk CK. Problems of allometric scaling analysis: examples from mammalian reproductive biology. J Exp Biol. 2005;208:1731–1747. | ||
Guerrier J, Journaux L, Chatelin YM, Ledos H [webpage on the Internet]. Gestation Length of French Bovine Breeds. Renc Rech Ruminants. 14 INRA- Institut de l’Elevage, Département génétique; 2007. (in French). Available from: www.journees3r.fr/IMG/pdf/2007_03_Genetique_13_Guerrier.pdf. Accessed May 23, 2016. | ||
Mukhametov LM. Unihemispheric slow-wave sleep in the Amazonian dolphin, Inia geoffrensis. Neurosci Lett. 1987;79(1–2):128–132. | ||
Lyamin OI, Chetyrbok IS. Unilateral EEG activation during sleep in the Cape fur seal, Arctocephalus pusillus. Neurosci Lett. 1992;143:263–266. | ||
Lyamin OI, Mukhametov LM, Chetyrbok IS, Vassiliev AV. Sleep and wakefulness in the southern sea lion. Behav Brain Res. 2002;128:129–138. | ||
Lyamin OI, Lapierre JL, Mukhametov LM. Sleep in aquatic species. In: Kushida C, editor. The Encyclopedia of Sleep. Waltham, MA: Academic Press; 2013;1:57–62. | ||
Pearson OP, Pearson AK. Certain aspects of reproduction in the fur seals. Nature. 2005;437:1264–1271. | ||
Hamlett GWD. Delayed implantation and discontinuous development in the mammals. Q Rev Biol. 1935;10:435–436. | ||
Lyamin OI, Kosenko PO, Lapierre JL, Mukhametov LM, Siegel JM. Fur seals display a strong drive for bilateral slow-wave sleep while on land. J Neurosci. 2008;28(48):12614–12621. | ||
Mengak MT [webpage on the Internet]. 0000 Natural History Series: Nine-Banded Armadillo (C 866-2), 2005. Available from: http://extension.uga.edu/publications/detail.cfm?number=C866-2. Accessed May 23, 2016. | ||
Tobler I, Schwierin BJ. Behavioural sleep in the giraffe (Giraffa camelopardalis) in a zoological garden. Sleep Res. 1996;5:21–32. | ||
TOOLITO BIO [webpage on the Internet]. Combien de temps dorment les animaux? Available from: http://www.toolito.com/bio/combien-dorment-animaux-sommeil/. Accessed May 23, 2016. | ||
Bradley RM, Mistretta CM. Fetal sensory receptors. Physiol Rev. 1975;55(3):352–382. | ||
Giraffe Birth at the Memphis Zoo, 2008 [webpage on the Internet]. Available from: https://www.youtube.com/watch?v=1kZW8e79Bm0. Accessed May 23, 2016. | ||
Lagercrantz H, Changeux JP. The emergence of human consciousness: from fetal to neonatal life. Pediatr Res. 2009;65:255–260. | ||
Siegel JM. Sleep viewed as a state of adaptive inactivity. Nat Rev Neurosci. 2009;10(10):747–753. | ||
Dawes GS, Fox HE, Leduc BM, Liggins GC, Richards RT. Respiratory movements and rapid eye movement sleep in the foetal lamb. Physiology. 1972;220:119–143. | ||
Bekoff A. Embryonic motor output and movement patterns: relationship to postnatal behavior. In: Smotherman WP, Scott R, editors. Behaviour of the Fetus. New Jersey: The Telford Press Caldwell; 1990:191–206. | ||
Shirozu H, Koyanagi T, Takashima T, Horimoto N, Akazawa K, Nakano H. Penile tumescence in the human fetus at term, a preliminary report. Early Hum Dev. 1995;41(3):159–166. | ||
Schmidt MH, Schmidt HS. Sleep-related erections: neural mechanisms and clinical significance. Curr Neurol Neurosci Rep. 2004;4(2):170–178. | ||
Blumberg MS, Coleman CM, Gerth AI, McMurray B. Spatiotemporal structure of REM sleep twitching reveals developmental origins of motor synergies. Curr Biol. 2013;23(21):2100–2109. | ||
Birnholz JC. On observing the human fetus. In: Smotherman WP, Scott R, editors. Behaviour of the Fetus. New Jersey: The Telford Press Caldwell; 1990:47–60. | ||
Mirmiran M, Scholtens J, van de Poll NE, Uylings HB, van der Gugten J, Boer GJ. Effects of experimental suppression of active (REM) sleep during early development upon adult brain and behavior in the rat. Brain Res. 1983;283(2–3):277–286. | ||
Herculano-Houzel S. Decreasing sleep requirement with increasing numbers of neurons as a driver for bigger brains and bodies in mammalian evolution. Proc Biol Sci. 2015;282(1816):20151853. | ||
Tononi G, Cirelli C. New hypothesis explains why we sleep. Sci Am. 2013;309(2):26–31. | ||
Pandi-Perumal SR, Gonfalone AA. Sleep in space as a new medical frontier: the challenge of preserving normal sleep in the abnormal environment of space missions. Sleep Sci. 2016;9(1):1–4. | ||
Hobson JA, Stickgold R. Available from: http://spaceflight.nasa.gov/history/shuttle-mir/science/hls/neuro/sc-hls-sleep.htm. Accessed June 7, 2016. | ||
Madan V, Jha SK. Sleep alterations in mammals: did aquatic conditions inhibit rapid eye movement sleep? Neurosci Bull. 2012;28:746–758. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.