")
Back to Journals » Infection and Drug Resistance » Volume 15
NDM-1 and OXA-48-Like Carbapenemases (OXA-48, OXA-181 and OXA-252) Co-Producing Shewanella xiamenensis from Hospital Wastewater, China
Authors Wen Y , Xie X, Xu P, Yang C, Zhu Z, Zhu J, Lv J, Zhang H, Chen L, Du H
Received 16 August 2022
Accepted for publication 9 November 2022
Published 29 November 2022 Volume 2022:15 Pages 6927—6938
DOI https://doi.org/10.2147/IDR.S386345
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Yicheng Wen,1,* Xiaofang Xie,1,* Ping Xu,2,* Chengcheng Yang,1 Zhichen Zhu,1 Jie Zhu,1 Jingnan Lv,1 Haifang Zhang,1 Liang Chen,3,4 Hong Du1
1Department of Clinical Laboratory, The Second Affiliated Hospital of Soochow University, Suzhou, Jiangsu, People’s Republic of China; 2Department of Clinical Laboratory, The Fifth People’s Hospital of Suzhou, Suzhou, Jiangsu, People’s Republic of China; 3Hackensack Meridian Health Center for Discovery and Innovation, Nutley, NJ, USA; 4Department of Medical Sciences, Hackensack Meridian School of Medicine, Nutley, NJ, USA
*These authors contributed equally to this work
Correspondence: Hong Du, Department of Clinical Laboratory, The Second Affiliated Hospital of Soochow University, 1055 Sanxiang Road, Suzhou, Jiangsu, People’s Republic of China, Email [email protected]
Background: Shewanella genus, as an important carrier of resistance genes, has the potential to transmit resistance to many antimicrobials in many circumstances, especially in aquatic environment. The aim of the study was to describe the risk of Shewanella xiamenensis in hospital environment through analysis of genomic comparison and resistance status.
Methods: Seven S. xiamenensis strains were isolated from hospital wastewater. PCR and Sanger sequencing were carried out for detection of common carbapenemase genes. Antimicrobial susceptibility testing was performed to determine the antimicrobial profile. Whole genome sequencing was applied, and sequences were further used for genomic analysis.
Results: Seven Shewanella xiamenensis were all positive for blaNDM and blaOXA-48. Antimicrobial susceptibility testing showed all Shewanella xiamenensis were resistant to cefotaxime, ceftazidime, imipenem, meropenem, gentamycin and trimethoprim-sulfamethoxazole. Whole genome sequencing and phylogenetic analysis demonstrated the diversity of Shewanella xiamenensis despite isolating from one wastewater pool.
Conclusion: To the best of our knowledge, this is the first report of detection of three types blaOXA-48-like genes in one hospital in China. And we have detected multi-drug resistant S. xiamenensis from hospital wastewater. This emphasizes that the presence of naturally existing carbapenemases in the environment may be significantly overlooked and that the blaOXA-48-like genes in China may originate through the horizontal gene transfer from S. xiamenensis to Enterobacterales rather than import from other countries.
Keywords: OXA-48, NDM, Shewanella xiamenensis, hospital wastewater, reservoir, horizontal transmission, surveillance, New Delhi metallo-β-Lactamase
Introduction
Antimicrobial resistance has become a threat for decades, and one representative example is carbapenem-resistant Enterobacterales (CRE), which have spread all over the world. CRE has received great attention in recent years due to its high morbidity, mortality, and lack of effective treatment solutions.1 And the main mechanism of carbapenem resistance is the production of different carbapenemases, and Klebsiella pneumoniae carbapenemase (KPC), New Delhi metallo-β-Lactamase (NDM), Verona integron-mediated metallo-β-lactamase (VIM), imipenemase (IMP), and oxacillinase-48 (OXA-48) are the most common carbapenemases over the world.2 Specifically, KPC and NDM are widely spread in America and China, while OXA-48-like is prevalent in Europe, North Africa, Middle East, and Indian subcontinent. OXA-48-like enzymes remain uncommon in American and most Asian countries, including China.3 In China, OXA-48-like producers have been reported in Shanghai, Jiangsu, Sichuan, Zhejiang, Guangdong. But few cases were related to foreign stay, and whether these blaOXA-48-like genes were originated from abroad was unclear.4–6
Surveillance has been widely regarded as the key point of administration of antibiotic resistance, which is informative for guidance on empirical treatment before antimicrobial susceptibility profile is determined.7,8 To make surveillance data more informative, samples from both clinical patients and healthy individuals should be included. However, this is demanding and difficult to achieve, thus hospital wastewater has been increasingly recognized as an alternative of classical surveillance due to the abundance of antibiotic residual and pathogenic bacteria released from large quantity of hospitalized population.9 Additionally, antimicrobial resistant genes (ARG) in wastewater have also been suggested as a suitable marker to predict clinical resistance prevalence in the hospitals.10 This provides the possibility to evaluate the antimicrobial resistant bacteria (ARB) communities and ARG prevalence in less developing regions with low input of expenditure and labor. On the other hand, detection of uncommon ARG from wastewater in a non-epidemic region indicates a risk of further transmission, with probably much more severe consequences than those have already been circulating in pathogens.8
The genus Shewanella, facultative anaerobic, motile gram-negative bacteria, is widely distributed in various environments, such as spoilt food and aquatic environments.11,12 In recent years, Shewanella infection cases have been rapidly increased and have been reported over the world. Most of cases are reported in tropical regions, and temperate countries and coastal cities like Australia, Spain, Martinique, Canary Islands, also have sporadic cases.13,14 Shewanella-related infections can involve multiple parts of human body, and could be divided into several categories, including ear, nose, and throat (ENT) disorders, central nervous system (CNS) disorders, chest infections, cardiovascular diseases, bloodstream infections, intra-abdominal infections, bone arthropathy, skin and soft-tissue infections (SSTIs).13 Marine exposure, compromised immune system, invasive procedures and poor hygiene in hospital are risk factors of Shewanella infections.11,15–17 Although no antimicrobial treatment guideline for Shewanella infections has been published yet, the empirical treatment could efficiently control infections.13 With the increase of antimicrobial resistance, however, clinicians and laboratory researchers should raise awareness to occurrence and dissemination of multi-drug resistant strains.
Here, we identified seven Shewanella xiamenensis strains that coproduce NDM-1 and OXA-48-like (OXA-48, OXA-181 and OXA-252) carbapenemases from hospital wastewater in Suzhou, China, where OXA-48-like-producing bacteria have not been detected before. We aimed to describe the risk of Shewanella xiamenensis in hospital environment through analysis of genomic comparison and resistance analysis.
Materials and Methods
Sample Collection, Culture Condition and Species Determination
To screen carbapenem-resistant bacteria in hospital wastewater, we performed the assay as described previously with minor modification.18 Briefly, water samples were collected from main sewer outlet at different depth of 0.5 to 1 m below the surface using 50mL sterile bottles. (Samples were chilled on ice during the sampling procedure and after being collected.) The samples were concentrated by 0.45 μm membrane filters, and ten-fold serial dilutions (10–1000 times) of each water sample were made in sterile saline solution. A total of 100 µL of each dilution was then plated onto LB agar plates supplemented with 2 mg/L meropenem. After incubated for 18 to 24 hours at 37°C, bacterial colonies with distinct coloration and morphologies were randomly picked and subcultured onto meropenem plates for further purification. Then each isolate was stored at −80 °C in 30% (vol/vol) glycerol for further investigation. And the species preliminary identification was determined using matrix-assisted laser desorption/ionization time of flight mass spectrometry (MALDI-TOF-MS, Bruker).
Carbapenemase Gene Assay and Antimicrobial Susceptibility Testing
To determine resistance mechanism to carbapenem of these strains, common carbapenemase genes were detected (blaKPC, blaNDM, blaIMP, blaVIM, blaOXA-48) using specific primer pairs followed by Sanger sequencing for their subtypes (Sangon, Shanghai). Primers were synthesized by Sangon and were listed in Table 1. Reaction buffer 2× Taq Mater Mix was provided by Vazyme (Nanjing).
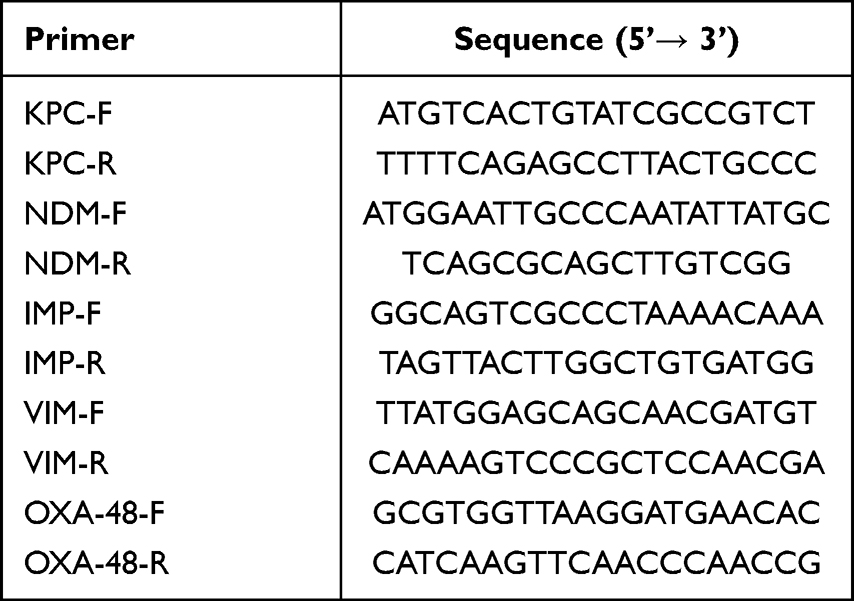 |
Table 1 Primers Used in This Study |
To characterize the antimicrobial susceptibility profile, we performed antimicrobial susceptibility testing. Isolates were suspended in 0.9% saline to 0.5 McFarland. Panel Phoenix NMIC/ID (BD) was used to determine MIC values on Phoenix M50 Automated System (BD, USA) and a total of 21 antibiotics were tested, including ampicillin, piperacillin, amoxicillin-clavulanate, ampicillin-sulbactam, piperacillin-tazobactam, imipenem, meropenem, cefazolin, cefepime, cefotaxime, ceftazidime, aztreonam, chloramphenicol, colistin, tetracycline, trimethoprim-sulfamethoxazole, amikacin, gentamycin, ciprofloxacin, levofloxacin and moxifloxacin. Results were interpreted according to “Other Non-Enterobacterales” in Clinical and Laboratory Standards Institute criteria (CLSI, 2020).
Whole Genome Sequencing
To further characterize the strains, we have performed genome sequencing. Bacterial genomic DNA were extracted by The Omega Bio-Tek Bacterial DNA Kit (Doraville, GA, USA) following instructions provided. Whole genome sequencing used the Illumina NovaSeq 6000 platform (Illumina, CA, USA) and then the corrected reads were assembled de novo utilizing SPAdes v3.11. In addition, sequencing of the complete genome sequence of isolate HD6416 and HD6452 were respectively performed with a sheared DNA library with average size of 10 kb on a Nanopore PromethION sequencer (Oxford Nanopore Technologies, OX, UK). The paired-end short Illumina reads were used to correct the long Nanopore reads, and then the corrected Nanopore reads were assembled de novo utilizing Unicycler v0.4.9 (https://github.com/rrwick/Unicycler).
Phylogenetic Analysis
To investigate the relationships of the blaOXA-48-like genes, the maximum-likelihood phylogenetic tree of blaOXA-48-like genes on Shewanella species available on Beta-Lactamase DataBase (BLDB, http://bldb.eu/) was constructed using MEGA 7.0.19
To describe the phylogenetic relationships of Shewanella xiamenensis strains, we collected all accessible genomes of Shewanella xiamenensis on NCBI to perform phylogenetic analysis. Based on recombination-free core genome, single-nucleotide polymorphisms (SNPs) was conducted with MUMmer 3.0, ClonalFrameML and MEGA7.0 as described previously.20 The sequence of Shewanella xiamenensis HD6416 (GenBank accession no. CP079717) was used as the reference, and the sequence of Shewanella putrefaciens CN-32 (GenBank accession no. NC_021505) was used as the outgroup. And the phylogenetic tree was further improved on iTOL and Inkscape v1.0 (https://inkscape.org/).21
Bioinformatic Analysis
Species identification was performed by calculating the pairwise average nucleotide identity (ANI) value between the genome sequence of the query strain and type strain of Shewanella xiamenensis (Accession number: BMOP01000000) by using OAT 0.93.1 (cutoff value of 96%).22
Open reading frames (ORFs) and pseudogenes were predicted using RAST 2.023 combined with BLASTP/BLASTN searches against the UniProtKB/Swiss-Prot database and the RefSeq database.23 Annotation of resistance genes, mobile elements, and other features were carried out using the online databases including CARD,24 ResFinder 4.0,25 ISfinder,26 and Tn Number Registry. Gene organization diagram was drawn in Inkscape v1.0 (https://inkscape.org/). BLAST Ring Image Generator (BRIG)27 was used to compare the differences between the plasmids.
Results
Isolate Characterization
Seven Shewanella putrefaciens were identified by MALDI-TOF-MS, but they were further determined as Shewanella xiamenensis in WGS analysis. The Shewanella xiamenensis were isolated from three different spots of the sewage outlets (Depth A, B and C) (Table 2). PCR testing and Sanger sequencing showed that all isolates co-harbored blaNDM-1 and blaOXA-48-like carbapenemase genes (including blaOXA-48, blaOXA-181 and blaOXA-252, Table 2). Antimicrobial susceptibility testing (Table 3) confirmed that all seven Shewanella strains were resistant to cefotaxime, ceftazidime, imipenem, meropenem, gentamycin and trimethoprim-sulfamethoxazole. And all of them were determined as multidrug-resistant. Only amikacin (100%), aztreonam (85.7%), tetracycline (71.4%) and levofloxacin (85.7%) retained high antibiotic activity to them. Additionally, just part of the strains were susceptible to piperacillin (14.3%), cefepime (14.3%), piperacillin-tazobactam (42.9%) and ciprofloxacin (28.6%), respectively.
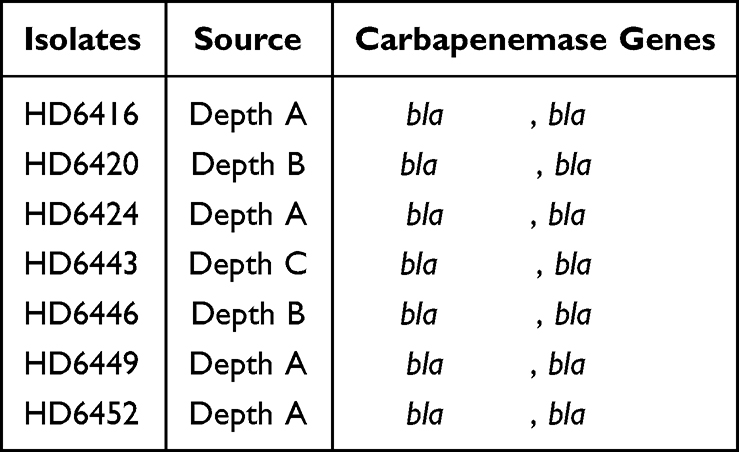 |
Table 2 Isolation Spot and Resistance Genes of Seven Shewanella xiamenensis |
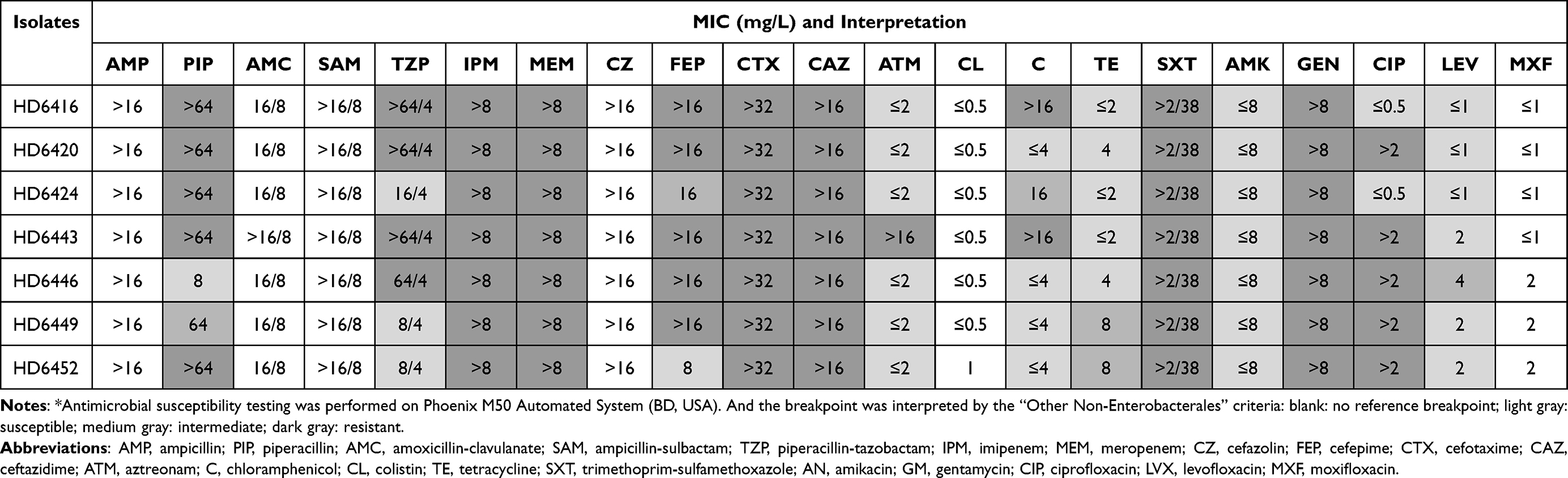 |
Table 3 Antimicrobial Drug Susceptibility of Seven Shewanella xiamenensis* |
Genome Analysis
Genetic Characterization
Whole genome sequencing (Illumina Hiseq) (Bioproject PRJNA755833) and analysis showed that blaOXA-48-like genes were on chromosomes while blaNDM-1 were likely on plasmids. Sequence analysis showed that HD6416, HD6424, HD6449 and HD6452 carry blaOXA-48, and HD6443 harbors blaOXA-252, while HD6420 and HD6446 contain blaOXA-181. The phylogenetic analysis of blaOXA-48-like of Shewanella spp. from NCBI showed the blaOXA-48 genes have 3 genotypes (different nucleotide sequences) despite being the same amino acid sequences while blaOXA-181 gene only has one genotype. In addition, plasmid-borne and chromosome-borne blaOXA-48 displayed some nucleotide variations but blaOXA-181 were identical (Figure 1A).
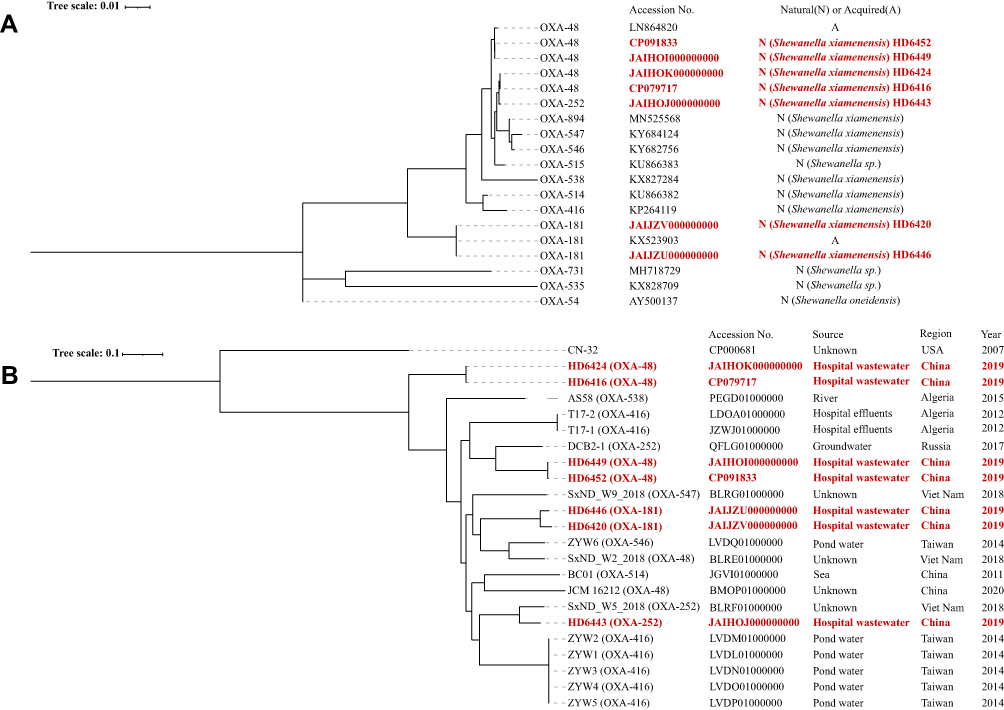 |
Figure 1 Phylogenetic trees of OXA-48-like genes and Shewanella xiamenensis strains. (A) Phylogenetic trees of OXA-48-like genes. blaOXA-48 genes have three genotypes despite being the same amino acid sequences while blaOXA-181 gene only has one genotype. Plasmid-borne and chromosome-borne blaOXA-48 displayed some nucleotide variations but blaOXA-181 were identical; (B) Phylogenetic trees of Shewanella xiamenensis strains. Seven S. xiamenensis genomes were distributed into different clades of the tree, and the phylogenetic clades in general correlated with the blaOXA-48-like gene sequences. Bootstrap values for each cluster of associated taxa are shown next to each branch. Bar corresponds to the scale of sequence divergence. Shewanella xiamenensis from our study were characterized in boldly red. |
SNP Analysis
Core genome single-nucleotide polymorphisms (SNPs) maximum-likelihood phylogenetic tree result (Figure 1B) showed that S. xiamenensis were classified into multiple clades with different subtree lengths. For the strains from our study, HD6416 and HD6424, HD6420 and HD6446, HD6449 and HD6452 were respectively grouped together, while HD6443 was located on a different branch. In addition, the seven genomes were distributed into different clades of the tree, indicating genomic diversity of the seven S. xiamenensis isolates, despite isolating from the same hospital. The S. xiamenensis phylogenetic clades in general correlated with the blaOXA-48-like gene sequences, but interestingly the OXA-48 producing HD6416/HD6424 and HD6420/HD6446 were located on different clusters. And a pairwise comparison of SNPs and similarities for all strains have been shown in Table S1.
Plasmid Analysis
Nanopore sequencing was further conducted in strain HD6416 and HD6452 (Bioproject PRJNA755833) to obtain completely closed chromosome and plasmid sequences. HD6416 and HD6452 both contain one circular chromosome and a plasmid. The PlasmidFinder analysis showed no known plasmid type were matched to pHD6416 and pHD6452, indicating that both plasmids belong to novel plasmid type.28,29 And plasmid comparison was further conducted using BRIG and result (Figure 2) indicated some variation between the backbone region of blaNDM-1-harboring plasmids. The backbone region of pHD6420, pHD6424 and pHD6443 share 99% identity with pHD6416, and pHD6446 has about 90% similarity with pHD6416 backbone, while pHD6449 and pHD6452 contain a different backbone. Notably, the multidrug-resistant regions of the seven plasmids were highly comparable with just a few mobile elements different. We then depicted gene diagrams of MDR region of two closed plasmids from our study (pHD6416 and pHD6452) for further comparison (Figure 3). The MDR region of both plasmids replaced In834 on Tn6358a, but with slight difference in resistance gene organization, so we designated them as Tn6358b (on pHD6416) and Tn6358c (on pHD6452), respectively. On Tn6358c, An ISCR-bleMBL-blaNDM-1 unit and an ISCR-qnrV6-blaCMY-10 unit were inserted in the In469 on Tn6358b. And the ΔIn1357 on Tn6358b was replaced by Tn6309 and part of ΔIn2145. The MDR region of Tn6358b and Tn6358c encode resistance to various antibiotics, such as aminoglycoside (dfrA27, aadA16), disinfectant (qacE), macrolides (msr(E)), rifampin (arr-3), sulfonamide (sul1), β-lactam, carbapenem (blaNDM-1), as well as efflux pump, such as acrABC-tolC. This suggests there were two backbone types between seven blaNDM-1 plasmids. The MDR regions of seven blaNDM-1 plasmids were highly similar with minor differences on resistance elements and arrangement, and they likely evolve from the same Tn element.
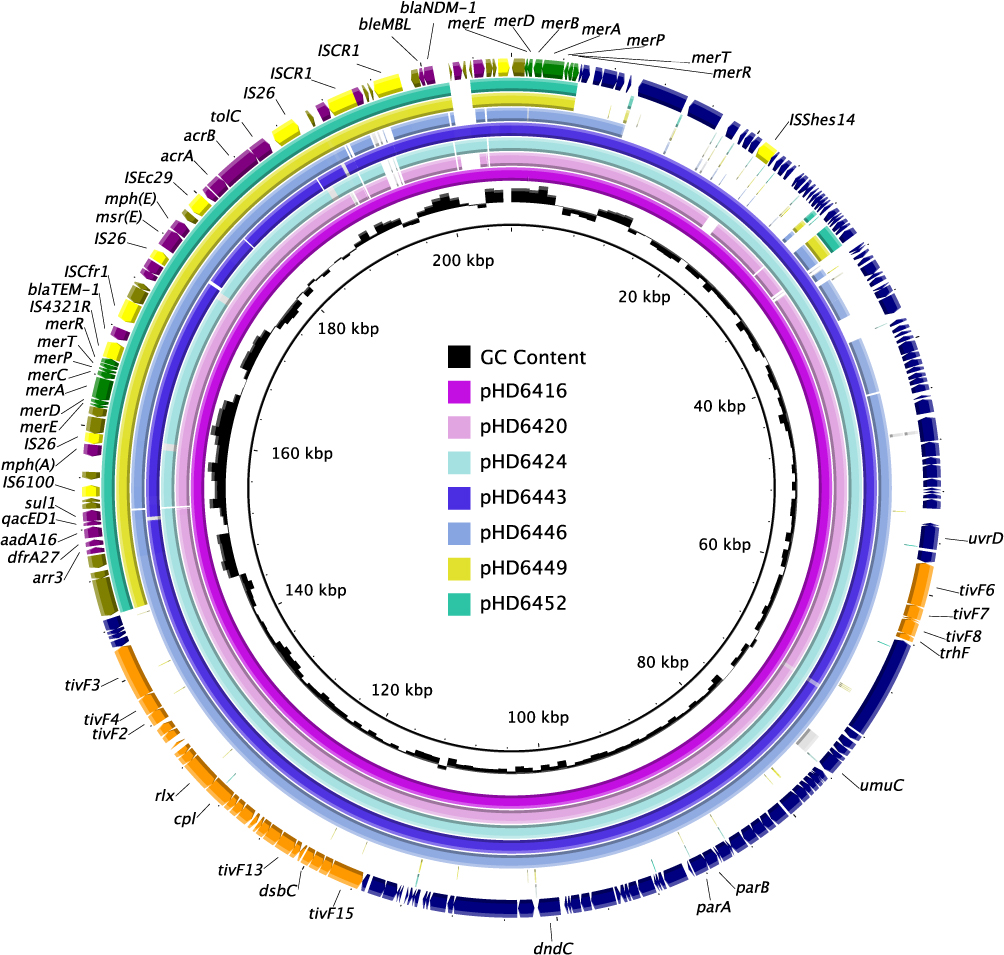 |
Figure 2 Plasmid structure comparison between blaNDM-1-harbouring pHD6416 and the other strains. The backbone region of pHD6420, pHD6424, pHD6443, pHD6446 was highly comparable with pHD6416 backbone, respectively, while pHD6449 and pHD6452 contain a different backbone. The multidrug-resistant regions of the seven plasmids were also highly comparable with just a few mobile elements different. Colored arrows indicate ORFs, with navy, yellow, Orange, olive, purple and green arrows representing plasmid backbone genes, mobile elements, plasmid transfer genes, Tn6358b/c backbone genes, the antimicrobial and heavy metal resistance genes, respectively. |
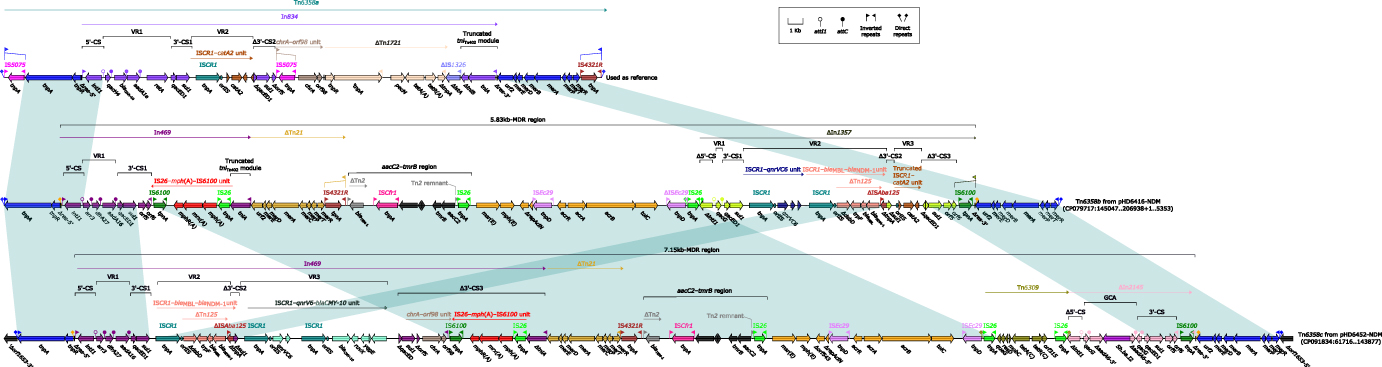 |
Figure 3 Linear comparison of Tn6358a, Tn6358b, Tn6358c. The MDR regions from pHD6416 and pHD6452 were highly similar and could evolve from the same Tn element. Genes are denoted by arrows. Gene, mobile elements, and other features are colored based on their functional classification. Shading denotes regions of homology (nucleotide identity ≥95%). Numbers in brackets indicate nucleotide positions within the plasmid pHD6416-NDM and pHD6452-NDM. Tn6358a was used as reference and its accession number was JX141473. |
Besides, oriTfinder analysis (https://tool-mml.sjtu.edu.cn/oriTfinder) failed to identify transfer origin regions (oriT) and relaxases sequences in all seven S. xiamenensis, suggesting the plasmid is non-transferable. Further conjugation assay also failed to transfer blaNDM-1-bearing plasmids to recipients of E. coli EC600 despite multiple attempts, which is consistent with the sequence analysis results. And blaOXA-48 genes were also not detected in recipients.
Discussion
Shewanella is an opportunistic pathogen and generally invades the immunocompromised and those who are exposed to marine environment. And the Shewanella genus has been frequently isolated from clinical specimens in recent years. According to a latest review, bloodstream infections, SSTIs and ENT disorders are most dominant Shewanella-related diseases, accounting for 37.0%, 35.9% and 27.5% of all cases, respectively. Unlike the former three, less Shewanella strains were isolated from patients of intra-abdominal infection (9.2%), bone arthropathy (6.6%), chest infection (4.4%), CNS disease (2.2%), cardiovascular infection (1.8%). And patients who suffer the infections are distributed in all generations, ranging from newborn to 92-years-old.13 Besides being pathogenic to human, Shewanella have been considered as a reservoir and vehicle of multiple antimicrobial resistance since the description of OXA-54 on S. oneidensis as the progenitor of OXA-48 in K. pneumoniae.30 Studies have found a variety of resistance genes from Shewanella genus, such as qnrA variants, dfrA1, aadA1, sul1, sul2, and blaOXA-48-like variants, conferring them resistance to many antibiotics, including β-lactams, quinolones, trimethoprim, aminoglycoside, sulfonamides, and carbapenems.10,31–33 And the occurrence of ARBs and ARGs in environment raises worries due to the potential hazard to public health. In our study, we reported three types of OXA-48-like carbapenemases (OXA-48, OXA-181 and OXA-252) from hospital wastewater, where blaOXA-48-like has not been detected before. Although the occurrence of waterborne species harboring NDM-1-encoding plasmids in China has been frequently reported,34,35 we describe a novel type blaNDM-1 plasmid of complicated structure in our study. We then further searched for similar blaNDM-1 plasmids in our isolate’s storage, but no similar plasmid was found. This suggested that the blaNDM-1 plasmids were assembled in wastewater, which could reflect the resistance gene diversity and complex genetic interaction in hospital wastewater.
The contributing factor to the ARG horizontally transmission is usually mobile element, such as insertion sequence. Insertion sequence has been also suggested accountable for the mobilization of blaOXA-48-like genes from chromosome to plasmid in this situation. We compared the genetic environment of blaOXA-48, blaOXA-181 and blaOXA-252 from our study with those on plasmids such as Tn1999 and Tn2013, respectively (Figure 4). blaOXA-48 and lysR on plasmid borne Tn1999 shared 2.237 kb of and >95% similarity with those on chromosomes of S. xiamenensis (cHD6416, cHD6420, cHD6449 and cHD6452). The blaOXA-48-lysR from Tn1999 in Enterobacterales plasmids were flanked by two copies of IS1999, indicating Tn1999 was likely responsible for the capture and mobilization of blaOXA-48-lysR region from chromosomes of S. xiamenensis to plasmid. Similarly, blaOXA-181-ΔlysR region on plasmid carried by Tn2013 shared 1.299 kb and >95% similarity with chromosome of S. xiamenensis HD6424. And blaOXA-181-ΔlysR region from Tn2013 in Enterobacterales plasmids were adjacent to ISEcp1, thus Tn2013 was more than likely to catch blaOXA-181-ΔlysR from chromosomes to plasmids.36,37 This is consistent with previous study that chromosome-encoded β-lactamases of Shewanella spp., eg S. xiamenensis, are likely the progenitors of blaOXA-48-like genes in Enterobacterale members.38 Different insertion sequences have been found to be linked to blaOXA-48-like genes on plasmids in Enterobacterales. The plasmid borne blaOXA-48 and blaOXA-163 are associated with IS1999 and ISEcl4, respectively, while blaOXA-181, blaOXA-204, and blaOXA-232 genes are associated with ISEcp1 elements but each in slightly different arrangements.38 These results suggested that blaOXA-48-like genes have been independently mobilized from their original location in Shewanella species by different insertion elements on multiple occasions. Under specific occasions, these blaOXA-48-like genes from the environmental S. xiamenensis may be recruited by additional mobile elements, followed by horizontal transfer and dissemination. In China, blaOXA-48-like producers are not as common as blaKPC producers, and so far they have been found in several provinces, including Shanghai, Jiangsu, Sichuan, Zhejiang, Guangdong, but few patients had a foreign stay before the blaOXA-48-like producers were isolated.4,39–41 For example, the first blaOXA-181 gene in China was found in a clinical ST410 E. coli strain WCHEC14828 from a Chinese patient without recent travel history.5 In this case, the possibility that blaOXA-48-like genes in China may originate through the horizontal gene transfer from S. xiamenensis to Enterobacterales rather than import from other countries, cannot be ruled out. Therefore, detection of OXA-48-like-producing S. xiamenensis from hospital wastewater in non-epidemic region, such as China, is rather indicative of the possibility of further transmission, which could lead to local epidemic of blaOXA-48-like genes. According to the surveillance of CRE in our hospital, OXA-48-like-producing Enterobacterales have not been detected yet, however, blaOXA-48-like transmission in our hospital is still possible due to the emergence of S. xiamenensis harboring blaOXA-48-like in hospital environment. This observation raised the concern that the S. xiamenensis from the hospital wastewater may serve as a reservoir for the transmission of ARGs, suggesting the presence of naturally existing carbapenemases in the environment may be significantly overlooked. Hence, the hospital surveillance towards both clinical isolates and wastewater should afterward continue to detect blaOXA-48-like gene prevalence in real time.
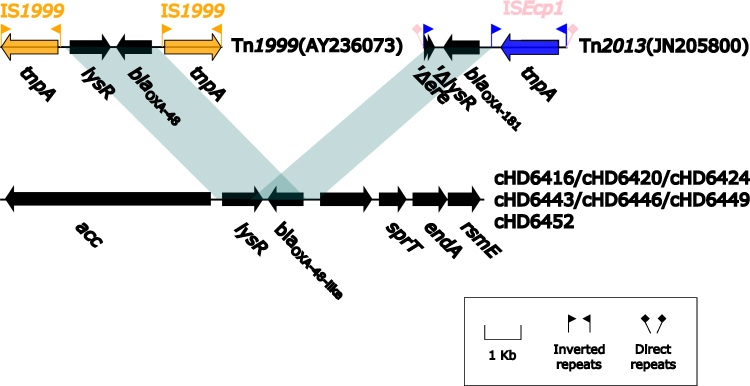 |
Figure 4 Linear Comparison of blaOXA-48-like-related region and related regions. Genes are denoted by arrows. Shading denotes regions of homology (nucleotide identity ≥90%). Sequences of Tn1999 and Tn2013 are acquired from pOXA-48a plasmid and pKP3-A plasmid on NCBI, respectively, and accession numbers are noted in bracket. |
Wastewater-based surveillance has been recommended as a low-cost methodology to monitor antimicrobial resistance prevalence, and could be useful in monitoring environmental resistance gene and helping decrease the health risks coming from the wastewater.42 However, research about hospital wastewater remains rare in China because of a lack of understanding and underestimation of surveillance. Future work should be considered to establish wastewater surveillance system in China hospitals to assess antimicrobial resistance abundances and monitor the transmission of resistance between human and the environment.
Conclusion
In our study, we reported seven S. xiamenensis co-producing NDM-1 and OXA-48-like carbapenemases (OXA-48, OXA-181 and OXA-252) from hospital wastewater in China and characterized them in some profiles. And to the best of our knowledge, this is the first report of detection of three types blaOXA-48-like genes in one hospital in China. This study proposes the possible origin of the blaOXA-48-like cases in China and warns the risk of blaOXA-48-like transmission in our hospital. Besides, an active antimicrobial resistance surveillance system in both clinical isolates and the hospital environment should be considered. And more attention is urgently needed on multi-drug resistant Shewanella genus.
Institutional Review Board Statement
Ethical approval is not applicable in this study as the samples were derived from environment. And the study was conducted in accordance with the principles of the Declaration of Helsinki.
Data Sharing Statement
The whole genome sequences of strains in this study were deposited in GenBank Bioproject PRJNA755833.
Acknowledgment
We are grateful for professor Dongsheng Zhou from State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology for the assistance in the drawing of figures. Yicheng Wen, Xiaofang Xie, and Ping Xu are co-first authors for this study.
Funding
This study was supported by the Science Foundation of Jiangsu Province Health Department [ZDB2020014]; Science Foundation of Suzhou Health Department [LCZX202106]; the Discipline Construction Program of the Second Affiliated Hospital of Soochow University [XKTJ-TD20200]; the Science and Technology Program of Suzhou [SZS201715].
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Nordmann P, Naas T, Poirel L. Global spread of Carbapenemase-producing Enterobacteriaceae. Emerg Infect Dis. 2011;17(10):1791–1798. doi:10.3201/eid1710.110655
2. Cui X, Zhang H, Du H. Carbapenemases in Enterobacteriaceae: detection and antimicrobial therapy. Front Microbiol. 2019;10:1823. doi:10.3389/fmicb.2019.01823
3. Poirel L, Potron A, Nordmann P. OXA-48-like carbapenemases: the phantom menace. J Antimicrob Chemother. 2012;67(7):1597–1606.
4. Li X, Ma W, Qin Q, et al. Nosocomial spread of OXA-232-producing Klebsiella pneumoniae ST15 in a teaching hospital, Shanghai, China. BMC Microbiol. 2019;19(1):235.
5. Liu Y, Feng Y, Wu W, et al. First report of OXA-181-producing Escherichia coli in China and characterization of the isolate using whole-genome sequencing. Antimicrob Agents Chemother. 2015;59(8):5022–5025.
6. Guo L, An J, Ma Y, et al. Nosocomial outbreak of OXA-48-producing Klebsiella pneumoniae in a Chinese hospital: clonal transmission of ST147 and ST383. PLoS One. 2016;11(8):e0160754.
7. Aarestrup FM, Woolhouse MEJ. Using sewage for surveillance of antimicrobial resistance. Science. 2020;367(6478):630–632.
8. Huijbers PMC, Flach CF, Larsson DGJ. A conceptual framework for the environmental surveillance of antibiotics and antibiotic resistance. Environ Int. 2019;130:104880.
9. Flach CF, Hutinel M, Razavi M, Ahren C, Larsson DGJ. Monitoring of hospital sewage shows both promise and limitations as an early-warning system for carbapenemase-producing Enterobacterales in a low-prevalence setting. Water Res. 2021;200:117261.
10. Huijbers PMC, Larsson DGJ, Flach CF. Surveillance of antibiotic resistant Escherichia coli in human populations through urban wastewater in ten European countries. Environ Pollut. 2020;261:114200. doi:10.1016/j.envpol.2020.114200
11. Janda JM, Abbott SL. The genus Shewanella: from the briny depths below to human pathogen. Crit Rev Microbiol. 2014;40(4):293–312. doi:10.3109/1040841X.2012.726209
12. Holt HM, Gahrn-Hansen B, Bruun B. Shewanella algae and Shewanella putrefaciens: clinical and microbiological characteristics. Clin Microbiol Infect. 2005;11(5):347–352. doi:10.1111/j.1469-0691.2005.01108.x
13. Yu K, Huang Z, Xiao Y, Wang D. Shewanella infection in humans: epidemiology, clinical features and pathogenicity. Virulence. 2022;13(1):1515–1532. doi:10.1080/21505594.2022.2117831
14. Gram L, Bundvad A, Melchiorsen J, Johansen C, Fonnesbech Vogel B. Occurrence of Shewanella algae in Danish coastal water and effects of water temperature and culture conditions on its survival. Appl Environ Microbiol. 1999;65(9):3896–3900. doi:10.1128/AEM.65.9.3896-3900.1999
15. Tsai MS, You HL, Tang YF, Liu JW. Shewanella soft tissue infection: case report and literature review. Int J Infect Dis. 2008;12(6):e119–24. doi:10.1016/j.ijid.2008.03.020
16. Oh HS, Kum KA, Kim EC, Lee HJ, Choe KW, Oh MD. Outbreak of Shewanella algae and Shewanella putrefaciens infections caused by a shared measuring cup in a general surgery unit in Korea. Infect Control Hosp Epidemiol. 2008;29(8):742–748. doi:10.1086/589903
17. Brink AJ, van Straten A, van Rensburg AJ. Shewanella (Pseudomonas) putrefaciens bacteremia. Clin Infect Dis. 1995;20(5):1327–1332. doi:10.1093/clinids/20.5.1327
18. Zhang L, Ma X, Luo L, Hu N, Duan J, Tang Z, Zhong R and Li Y. (2020). The Prevalence and Characterization of Extended-Spectrum β-Lactamase- and Carbapenemase-Producing Bacteria from Hospital Sewage, Treated Effluents and Receiving Rivers. IJERPH, 17(4), 1183 10.3390/ijerph17041183
19. Naas T, Oueslati S, Bonnin RA, et al. Beta-lactamase database (BLDB) - structure and function. J Enzyme Inhib Med Chem. 2017;32(1):917–919. doi:10.1080/14756366.2017.1344235
20. Liang Q, Jiang X, Hu L, et al. Sequencing and genomic diversity analysis of IncHI5 plasmids. Front Microbiol. 2018;9:3318. doi:10.3389/fmicb.2018.03318
21. Letunic I, Bork P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021;49(W1):W293–W6. doi:10.1093/nar/gkab301
22. Lee I, Ouk Kim Y, Park SC, Chun J. OrthoANI: an improved algorithm and software for calculating average nucleotide identity. Int J Syst Evol Microbiol. 2016;66(2):1100–1103. doi:10.1099/ijsem.0.000760
23. Aziz RK, Bartels D, Best AA, et al. The RAST server: rapid annotations using subsystems technology. BMC Genomics. 2008;9:75. doi:10.1186/1471-2164-9-75
24. Alcock BP, Raphenya AR, Lau TTY, et al. CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020;48(D1):D517–D25. doi:10.1093/nar/gkz935
25. Bortolaia V, Kaas RS, Ruppe E, et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J Antimicrob Chemother. 2020;75(12):3491–3500. doi:10.1093/jac/dkaa345
26. Siguier P, Perochon J, Lestrade L, Mahillon J, Chandler M. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006;34(Database issue):D32–6.
27. Alikhan NF, Petty NK, Ben Zakour NL, Beatson SA. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics. 2011;12:402.
28. Carattoli A, Zankari E, Garcia-Fernandez A, et al. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58(7):3895–3903.
29. Camacho C, Coulouris G, Avagyan V, et al. BLAST+: architecture and applications. BMC Bioinform. 2009;10:421.
30. Poirel L, Heritier C, Nordmann P. Chromosome-encoded ambler class D beta-lactamase of Shewanella oneidensis as a progenitor of carbapenem-hydrolyzing oxacillinase. Antimicrob Agents Chemother. 2004;48(1):348–351.
31. Kim HB, Park CH, Gavin M, Jacoby GA, Hooper DC. Cold shock induces qnrA expression in Shewanella algae. Antimicrob Agents Chemother. 2011;55(1):414–416.
32. Poirel L, Rodriguez-Martinez JM, Mammeri H, Liard A, Nordmann P. Origin of plasmid-mediated quinolone resistance determinant QnrA. Antimicrob Agents Chemother. 2005;49(8):3523–3525.
33. Soledad Ramirez M, Merkier AK, Almuzara M, Vay C, Centron D. Reservoir of antimicrobial resistance determinants associated with horizontal gene transfer in clinical isolates of the genus Shewanella. Antimicrob Agents Chemother. 2010;54(10):4516–4517.
34. Jiang N, Zhang X, Zhou Y, Zhang Z, Zheng X. Whole-genome sequencing of an NDM-1- and OXA-58-producing Acinetobacter towneri isolate from hospital sewage in Sichuan Province, China. J Glob Antimicrob Resist. 2019;16:4–5.
35. Hu Y, Feng Y, Qin J, Zhang X, Zong Z. Acinetobacter chinensis, a novel Acinetobacter species, carrying blaNDM-1, recovered from hospital sewage. J Microbiol. 2019;57(5):350–355.
36. Zong Z. Discovery of bla(OXA-199), a chromosome-based bla(OXA-48)-like variant, in Shewanella xiamenensis. PLoS One. 2012;7(10):e48280.
37. Potron A, Poirel L, Nordmann P. Origin of OXA-181, an emerging carbapenem-hydrolyzing oxacillinase, as a chromosomal gene in Shewanella xiamenensis. Antimicrob Agents Chemother. 2011;55(9):4405–4407.
38. Evans BA, Amyes SG. OXA beta-lactamases. Clin Microbiol Rev. 2014;27(2):241–263.
39. Zhu Z, Huang H, Xu Y, et al. Emergence and genomics of OXA-232-producing Klebsiella pneumoniae in a hospital in Yancheng, China. J Glob Antimicrob Resist. 2021;26:194–198.
40. Patil S, Chen H, Zhang X, Lian M, Ren PG, Wen F. Antimicrobial resistance and resistance determinant insights into multi-drug resistant gram-negative bacteria isolates from paediatric patients in China. Infect Drug Resist. 2019;12:3625–3634.
41. Shu L, Dong N, Lu J, et al. Emergence of OXA-232 carbapenemase-producing Klebsiella pneumoniae that carries a pLVPK-like virulence plasmid among elderly patients in China. Antimicrob Agents Chemother. 2019;63(3):Mar.
42. Samreen AI, Malak HA, Abulreesh HH. Environmental antimicrobial resistance and its drivers: a potential threat to public health. J Glob Antimicrob Resist. 2021;27:101–111.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.