")
Back to Journals » International Journal of Nanomedicine » Volume 11
Nanobody-derived nanobiotechnology tool kits for diverse biomedical and biotechnology applications
Authors Wang Y, Fan Z, Shao L, Kong X, Hou X, Tian D, Sun Y, Xiao Y, Yu L
Received 25 February 2016
Accepted for publication 15 June 2016
Published 21 July 2016 Volume 2016:11 Pages 3287—3303
DOI https://doi.org/10.2147/IJN.S107194
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Linlin Sun
Yongzhong Wang,1 Zhen Fan,2 Lei Shao,3 Xiaowei Kong,1 Xianjuan Hou,1 Dongrui Tian,1 Ying Sun,1 Yazhong Xiao,1 Li Yu4
1School of Life Sciences, Collaborative Innovation Center of Modern Bio-manufacture, Anhui University, Hefei, People’s Republic of China; 2Department of Biomedical Engineering, The Ohio State University, Columbus, OH, USA; 3State Key Laboratory of New Drugs and Pharmaceutical Process, Shanghai Institute of Pharmaceutical Industry, Shanghai, 4Department of Microbiology and Parasitology, Anhui Provincial Laboratory of Microbiology and Parasitology, Anhui Key Laboratory of Zoonoses, Anhui Medical University, Hefei, People’s Republic of China
Abstract: Owing to peculiar properties of nanobody, including nanoscale size, robust structure, stable and soluble behaviors in aqueous solution, reversible refolding, high affinity and specificity for only one cognate target, superior cryptic cleft accessibility, and deep tissue penetration, as well as a sustainable source, it has been an ideal research tool for the development of sophisticated nanobiotechnologies. Currently, the nanobody has been evolved into versatile research and application tool kits for diverse biomedical and biotechnology applications. Various nanobody-derived formats, including the nanobody itself, the radionuclide or fluorescent-labeled nanobodies, nanobody homo- or heteromultimers, nanobody-coated nanoparticles, and nanobody-displayed bacteriophages, have been successfully demonstrated as powerful nanobiotechnological tool kits for basic biomedical research, targeting drug delivery and therapy, disease diagnosis, bioimaging, and agricultural and plant protection. These applications indicate a special advantage of these nanobody-derived technologies, already surpassing the “me-too” products of other equivalent binders, such as the full-length antibodies, single-chain variable fragments, antigen-binding fragments, targeting peptides, and DNA-based aptamers. In this review, we summarize the current state of the art in nanobody research, focusing on the nanobody structural features, nanobody production approach, nanobody-derived nanobiotechnology tool kits, and the potentially diverse applications in biomedicine and biotechnology. The future trends, challenges, and limitations of the nanobody-derived nanobiotechnology tool kits are also discussed.
Keywords: nanobody, VHH, V-NAR, HCAbs, IgNAR, nanobiotechnology
Introduction
Over the decades, single-domain antibodies (sdAbs) have received a progressive interest from pharmaceutical and biotechnology industries owing to their peculiar properties, including small size, robust structure, high affinity and specificity, superior cryptic cleft accessibility, and deep tissue penetration.1–3 The sdAbs could be categorized into man-made sdAbs and naturally occurring counterparts,4 the latter including variable domain of heavy-chain-only antibodies in camelids (VHH), a naturally occurring antigen-binding variable domain of heavy chain from heavy-chain-only antibodies (HCAbs),1 and variable domain of immunoglobulin new antigen receptors in sharks (V-NAR), a variable domain from immunoglobulin new antigen receptors (IgNARs).5 Totally different from the man-made sdAbs, which have been pursued for the ideal ones by scientists for half a century, several naturally occurring sdAbs, including VHHs and V-NARs, already exist in nature in camelids or cartilaginous fish, sharing a surprising structural convergent evolution and performing similar biological functions.1–5
In addition to the classical antibodies containing two heavy and two light chains, all camelidae including camels (Camelus dromedarius and Camelus bactrianus), llama (Lama glama and Lama guanicoe), and vicugna (Vicugna vicugna and Vicugna pacos) have HCAbs in their sera, lacking L chains and devoid of a canonical constant heavy-chain (CH) 1 domain.1 Some cartilaginous fish, including nurse shark (Ginglymostoma cirratum), wobbegong (Orectolobus maculates), and dogfish (Squalus acanthias and Mustelus canis) sharks, also remarkably produce functional heavy-chain-only immunoglobulins (Igs), named IgNARs.1,5–8 Interestingly, some pathological and nonfunctional HCAbs were also discovered in sera of humans or in mouse hybridoma due to a partly genetic deletion of the variable heavy-chain (VH) and CH1 regions.1 In camels or sharks, these HCAbs (or IgNARs) recognize the antigens via single variable domains, referred to as VHHs or V-NARs, respectively. Thus, the VHHs or V-NARs are the smallest intact antigen-binding domain derived from the HCAbs or IgNARs naturally occurring in camelids or cartilaginous fish.
Nanobody is referred to the VHHs, the sdAb fragments derived from naturally occurring camelid HCAbs or the counterpart domain V-NAR of IgNARs, the homodimeric antibodies devoid of light chains in sharks. Owing to the small dimensional size of 2.5 nm in diameter and 4 nm in height (~12–15 kDa), Ablynx in 2003 dubbed these sdAbs “nanobody” to emphasize their smaller dimensional sizes, compared to larger molecular sizes of single-chain variable fragments (scFvs; 27 kDa), antigen-binding fragments (Fabs; ~57 kDa), and the intact conventional immunoglobulin-γ (IgG) antibody (~150 kDa).3,5,8 In this review, we summarize the current state of the art in nanobody research, focusing on the nanobody structural features, nanobody production approach, nanobody-derived nanobiotechnology tool kits, and the potential applications in biomedicine and biotechnology.
Structure of nanobodies
In comparison with the conventional IgG antibody assembled from two identical heavy-chain and two identical light-chain (heterotetrameric structure, Figure 1A–C), camelid HCAbs are HCAbs (homodimeric structure, Figure 1D–F), devoid of the light-chain polypeptide and the first constant domain (CH1) of heavy-chain polypeptide.1–3 The absence of light chains in the whole antibody and lack of the CH1 domain in the heavy chain are two significant features of camelid HCAbs, which provide them with a more compact architecture and smaller dimensional size with a molecular weight of ~90 kDa rather than ~150 kDa for the canonical antibody IgG.1,3 Similarly, the Ig isotype IgNAR (novel antigen receptor) discovered in the shark bloodstream is also a homodimeric structure of two heavy-chain polypeptides, each comprising a single variable domain and five constant domains (homodimeric structure, Figure 1G–I).7
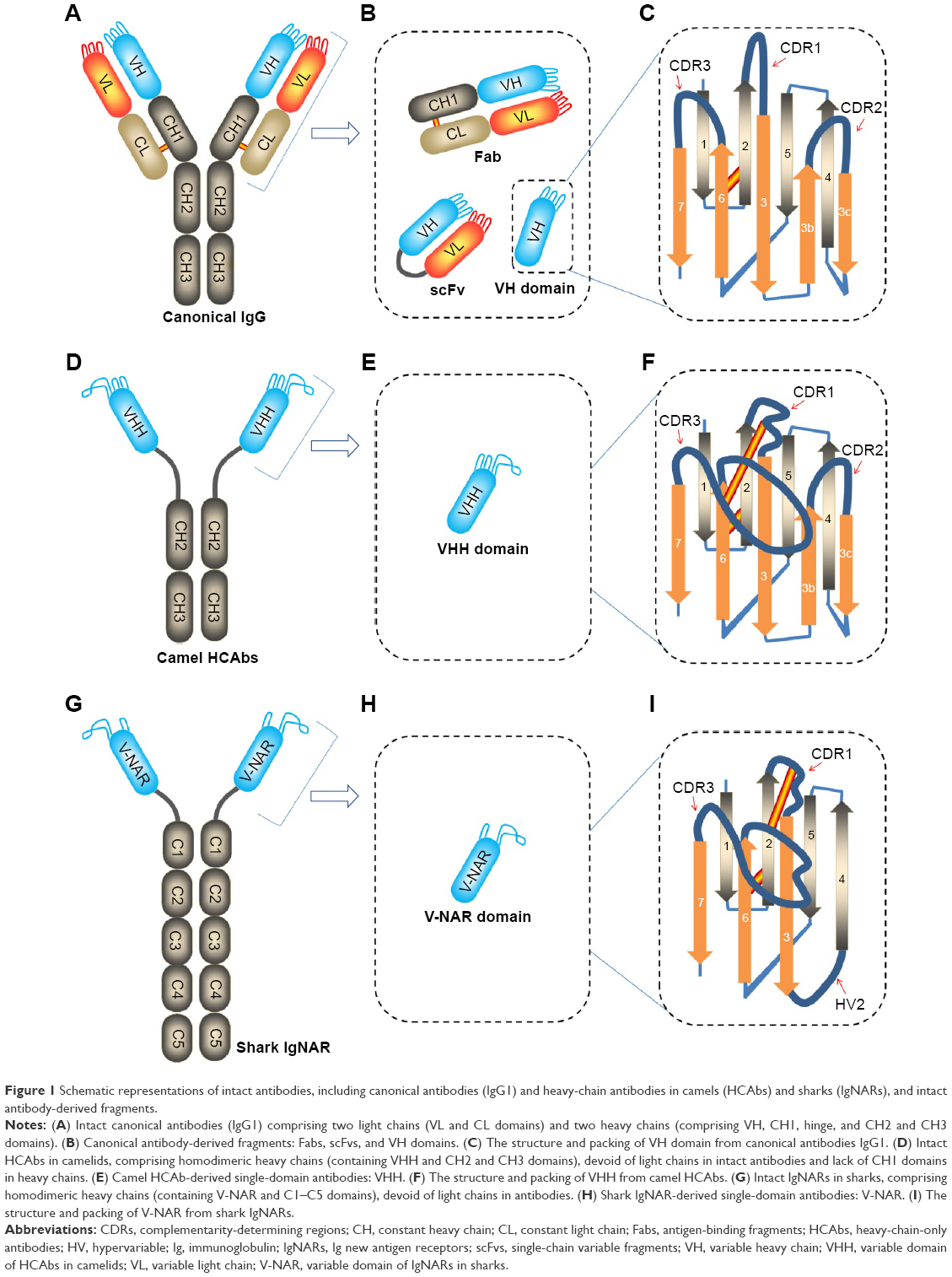 | Figure 1 Schematic representations of intact antibodies, including canonical antibodies (IgG1) and heavy-chain antibodies in camels (HCAbs) and sharks (IgNARs), and intact antibody-derived fragments. |
The percentage of HCAbs in the bloodstream of camelids varies greatly among species because of a variation in mutation rates.3 It might reach relatively high level in camels, ranging from ~50% to ~80%, whereas it totals up to 10%–25% in South American camelid species.1 Relative to the HCAbs in camelidae, the shark IgNARs are slightly at lower level, ranging from ~0.1 mg/mL to 1 mg/mL, counting for ~5% of the total Igs in the bloodstream.5,9 The HCAbs and IgNARs have shown very high hypermutation, apparently in response to antigens in the immune protection of the camelid and the shark.1,5,9 Therefore, to some extent, the nanobodies (VHHs or V-NARs) with a molecular weight of ~12–15 kDa in the HCAbs or IgNARs are the structural and functional counterparts of the Fabs in the conventional IgGs (Figure 1B, E, and H).1,3,5,8
Similar to the VH domain in conventional IgGs, the folded VHH in camel HCAbs comprises nine β-strands. These β-strands, organized in a four-stranded β-sheet and a five-stranded β-sheet, were connected by three hypervariable (HV) loops, also called complementarity-determining regions (CDRs), and by a conserved disulfide bond between Cys23 within framework region (FR) 1 and Cys94 within FR3.1 The three CDRs located at the N-terminal end of the domain forms a continuous surface (paratope) in response to recognizing epitopes of antigens. This means that VHHs of the camel HCAbs and VH domains of the conventional IgGs share the similar structural architecture of the FRs and loops (Figure 1C and F).1,2,10,11 The alignment of the VHH amino acid sequences indicates that two major significant differences between both the VHH and VH domains exist within the FR2 and in the CDRs, especially in the CDR3.
The first notable difference between VHH and VH is within FR2.1,3 The highly conserved hydrophobic amino acids found within FR2 of the conventional VH region at positions (Val37, Gly44, Leu45, and Trp47) are replaced by hydrophilic and/or smaller amino acids, mostly Phe42, Glu49, Arg50, and Gly52, respectively.1,12–15 These hydrophobic amino acids normally participate in the interaction with chaperone proteins and the variable light chain (VL) domain during the assembly of heavy chains and light chains of the conventional antibodies.1 Thus, reshaping this VL side of the domain deprives the heavy chains of their binding sites for any VL interaction, enhancing solubility and reducing aggregation of VHH in the absence of a VL domain.1,3
The second difference between VHH and VH is found in the HV loops. There is a broader structural repertoire of loops in VHH, especially an enlarged HV region in the H1 loop (CDR1) and an extended H3 loop (CDR3) (Figure 1C and F). These uniquely extended loops provide a sufficient antigen-interacting surface, large enough to 600–800 Å2 as offered by six loops from the VH–VL domain pair in the traditional IgGs.1,3 It has been established that the elongated CDR3 is capable of extending into cavities on antigens, such as the active site crevice of enzymes and ligand-binding cavity of receptors.1,3 However, an enlarged loop suggests broader flexibility, expectedly leading to be entropically counterproductive for binding. This problem is solved in camel VHHs by fastening the extended loop of CDR3 with an extra disulfide bond toward either the CDR1 loop (Figure 1F) or the CDR2 loop or even the FR2.1,3,16 These disulfide bonds might maximally optimize the binding surface topology and facilitate the orientation of the CDR3 toward the antigens.3
As for the V-NARs, it shows homology to Vα domain of the T-cell receptor.5,17 Contrary to VHH, V-NAR only consists of seven β-strands instead of nine β-strands due to the truncation in FR2–CDR2 (Figure 1F and I). This means that V-NARs have only two CDRs, ie, CDR1 and CDR3 (Figure 1I). Similar to the camelid VHHs, the V-NAR has abnormally long CDR3 loops that considerably reflect the greatest diversity of the V-NAR in both sequence and length.7 Furthermore, highly frequent somatic mutation is also found in CDR1, at the equivalent site of the deleted CDR2, where the shorten loop forms a belt-like structure at the opposite side of the CDR1 and CDR3, and in a loop that resembles HV4 in T-cell receptors.5,18 Similar to the camel VHHs, the shark V-NARs also evolved to generate equivalent disulfide bridges tethering the extended antigen-binding loops.1,19,20
Although the sequences of VHHs and V-NARs are very diverse,1 the camel VHHs shared the similar architecture with V-NARs. Overall, both show a rugby ball-shaped structure and a convex or protruding surface at the N-terminal end of the domains.19,21 This protruding surface increases the actual interaction surface of the paratopes, extremely facilitating insertion of nanobodies in cavities on the surface of the antigens or ligand-binding sites of receptors.22 In limited cases, a flat paratope surface23 and occasionally a cavity for the antihapten binders24,25 were also observed owing to the long loops in most nanobody structures folding over the FR2 region in VHH.1 These different shapes of the paratope surfaces demonstrated the extreme flexibility and great diversity of both the sdAbs.
Nanobody libraries
To fish out a desired nanobody with high stability and subnanomolar or picomolar affinity, preparation of nanobody libraries using different approaches was reasonably proposed.1,4,7,26,27 The technologies used for the preparation of nanobody library do not significantly differ from the ones otherwise used for recovering the Fab and scFv libraries. First of all, retrieval of nanobodies from an immune library is a priority consideration because somatic maturation in lymphocytes of immunized Camelidae will give antibody libraries more specific and higher affinity to antigens of interest.4 However, for each new antigen, we have to prepare a new immune library, which might unnecessarily spend more time and costs than other strategies proposed where else, such as large one-pot libraries without immunization of animals.4 Thus, a suitable naive library using blood samples from nonimmunized animals or the semisynthetic and synthetic libraries are practically alternative choices. Despite the lack of somatic maturation, it is possible that the selection based on phage display using such one-pot large libraries (>109 clones per library) allowed to isolate such VHHs with high affinity in the subnanomolar or picomolar range that are suitable for the diverse biomedical applications.
Immune library
Preparation of the immune nanobody library first needs an immunization of camelidae, through which antigen-specific HCAbs are affinity matured. The immunization procedures are mostly involved in prime–boost strategy using various antigens of interest as immunogens. After a brief immunization, the nanobodies are generally readily obtained by cloning the V gene repertoire from peripheral blood lymphocytes and by screening through phage display or other biological carriers.1,26 The entire Fab of the HCAb comprises only one VHH, and it contains ~120 amino acid residues, encoded by a gene fragment of only ~360 bp. Thus, the VHH gene is easily cloned by polymerase chain reaction (PCR) in one single amplicon. As a result, small libraries (just ~106 individual clones per library) created by using a ~50 mL of blood sample already represent the immune VHH repertoire of lymphocytes present in bloodstream of the immunized animals.1
As for the amplification and cloning of an scFv, the VH and VL exons needed to be first individually PCR amplified, which probably results in scrambled pairs of the VH and VL domains due to their random assembly. In contrast to the scFv cloning, the PCR amplification of the VHH present in only one exon generally facilitated generation of intact and affinity-matured VHHs from peripheral B-lymphocytes. Thus, the unique specificity and high affinity of nanobodies from immune libraries are guaranteed. Kinetic kon and koff rate constants of the nanobodies retrieved from the immune libraries are routinely reported to be low nanomolar or even picomolar levels, ranging from 105 to 106 M−1 s−1 and 10−2–10−4 s−1, respectively.1 Such affinity parameters are excellent properties for the most biomedical applications, including disease diagnosis, bioimaging, drug screening, and targeting therapy.
Naive library
When toxic or nonimmunogenic antigens are potentially used as immunogens, or immunization is not available for some other reasons, naive V repertoires could be employed to replace the immune VHH libraries. This approach allows us saving the time and costs related to preparing a new library for any new antigens of interest. Due to the lack of somatic maturation stimulated in vivo by immunization process, such libraries theoretically need to include ~109 individual clones to allow the retrieval of high-affinity binders to a given antigen in general.1 Practically, the theoretical diversity of a naive library increases with increasing of the number of lymphocytes initially collected. Thus, large volume of blood samples (>1 L) collected from different animals is a prerequisite for preserving the greatest genetic diversity. In addition, to avoid unnecessary diversity reduction during cloning of this type of library, all of the necessary steps should be performed with the highest care to reduce the material loss. As a practical alternative, the final library size could also be beneficially increased by mixing independently prepared collections to guarantee the diversity of the library.4
Semisynthetic/synthetic library
Limited size and diversity is a key problem that affects even large size naive-based nanobody libraries.27 Without an enormous germline diversity and the recombinatorial diversity from VH/VL pairing in traditional IgGs, affinity maturation in camelids or sharks relies to a larger extent on somatic hypermutation that precisely tunes the CDRs to recognize any given antigens.27 Thus, in an effort to mimic such in vivo diversification to yield diverse enough libraries capable of generating nanobodies to any given antigen, another strategy, ie, semisynthetic/synthetic library, was proposed. The high-affinity nanobody could also be fished out using this type of library without immunization of animals. The strategy is trying to conserve the FRs surrounding the CDRs, which may be crucial in conserving the structural integrity of nanobodies, and to randomly diversify the sequences of the CDRs, especially of CDR3.27
Based on naturally occurring VHH or V-NAR sequences, the semisynthetic or synthetic nanobody libraries could be created by introducing length and sequence variations in CDR3 using randomized CDR3 primers,18 or error-prone PCR combined with splice-overlap extension PCR method.7,27 Using small blood samples (<10 mL), the complexity of diversity-enhanced semisynthetic or synthetic nanobody library is close to 109, whereas the complexity of 106 of naive library could only be reached by using the same volume of blood samples. At least >1 L of blood samples might be consumed in order to obtain ~109 individual clones per library for the naive library.4 This means that the semisynthetic or synthetic library had better CDR3 diversity and better utility than the naturally occurring naive VHH or V-NAR libraries without immunization of animal or collection of large volume of blood samples. Thus, this may be a promising path toward obtaining a limitless source of nanobodies against a variety of antigens without immunization of animals.
Independent of the strategies to construct libraries, the established libraries further need to be displayed on different biological carries, including phages, bacteria, yeasts, and ribosomes, to facilitate screening and panning for a given antigen-specific nanobody. Retrieval of nanobodies from the libraries by phage display or any other selection protocols described earlier, including bacterial display, yeast display, intracellular two-hybrid selection, and ribosome display, has been well documented.1 By a variety of standard biopanning strategies, these libraries are preciously hidden treasure with great molecular diversity and could be utilized to fish out the nanobody binders with desired properties to various antigens of interest.
Nanobody-derived nanobiotechnology tool kits
Within the new vista of nanobiotechnological applications, different nanosized biotools, nanoscaled biomacromolecules, and engineered bacteriophages have been employed as promising approaches to meet the unmet needs of biomedicine and biotechnology development for human health. Owing to the desired properties of nanobody, including nanoscaled size, stable and soluble behavior in aqueous solution, reversible refolding, humanizable sequences, and specific and high affinity for only one cognate target, as well as a sustainable source, nanobody has been an ideal research tool for the development of sophisticated nanobiotechnologies. Currently, the nanobody has been evolved into versatile research and application tool kits for diverse nanobiotechnology applications.1–3,10,11,28 A variety of nanobody-derived formats, including the nanobody itself, the radionuclide or fluorescent dye-labeled nanobodies, fluorescent protein or chromogenic enzyme fusion nanobodies, bivalent nanobodies, self-assembly motif-mediated nanobody homo- or heteromultimers, nanobody-coated nanoparticles, and nanobody-displayed bacteriophages (Figure 2), have been successfully demonstrated as powerful nanobiotechnological tool kits for diverse biomedical applications, including targeting drug delivery and therapy,1,3 disease diagnosis,2,3 bioimaging,11,29–31 and agricultural and plant protection.32,33 These applications indicate a special advantage of these nanobody-derived technologies, already surpassing the “me-too” products of other equivalent binders, such as the full-length antibodies, scFvs, Fabs, targeting peptides, and DNA-based aptamers.
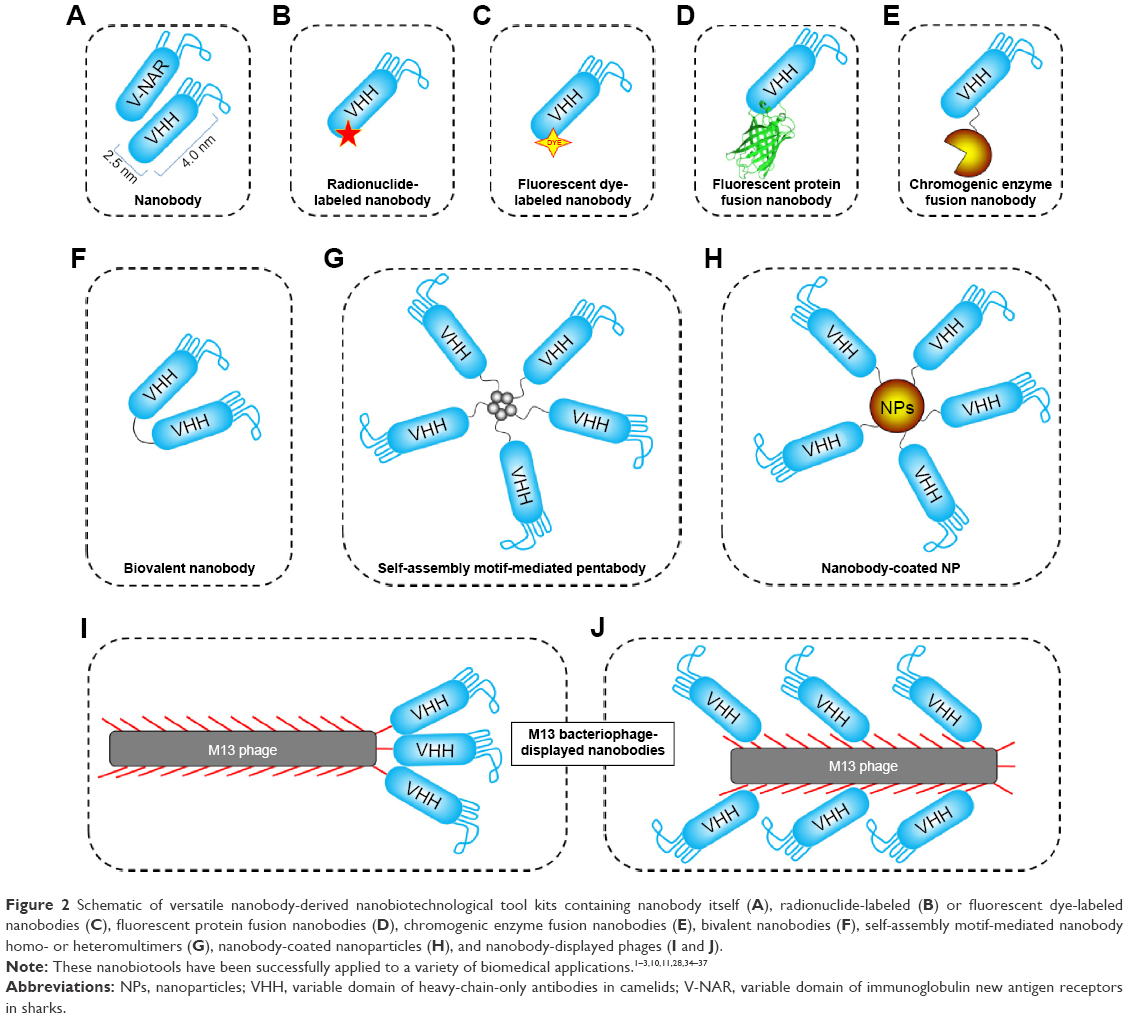 | Figure 2 Schematic of versatile nanobody-derived nanobiotechnological tool kits containing nanobody itself (A), radionuclide-labeled (B) or fluorescent dye-labeled nanobodies (C), fluorescent protein fusion nanobodies (D), chromogenic enzyme fusion nanobodies (E), bivalent nanobodies (F), self-assembly motif-mediated nanobody homo- or heteromultimers (G), nanobody-coated nanoparticles (H), and nanobody-displayed phages (I and J). |
Versatile applications of nanobody-derived nanobiotechnologies
There are various biomedical applications using the nanobody-derived nanobiotechnologies, which has been extensively covered recently elsewhere.1–3,10,11,28,34–37 Here, we focus on a number of examples, wherein nanobodies provide special advantages over other equivalent binders. These applications demonstrated a promising future of the use of nanobodies in versatile environments, including basic research, bioimaging, clinical diagnosis, therapeutics, and agricultural and plant protection. Compared to the conventional sdAbs, these diverse applications indicate the versatile and novel properties of nanobodies as promising sdAbs.
Nanobodies as versatile research tools in biotechnology
With the versatile properties of the nanobodies, they have been developed into various research tools used in basic research, including affinity purification, immunoprecipitation, chaperone-assisted crystallization, protein degradation, gene activation or inactivation, protein–protein interaction, and many others (Table 1).
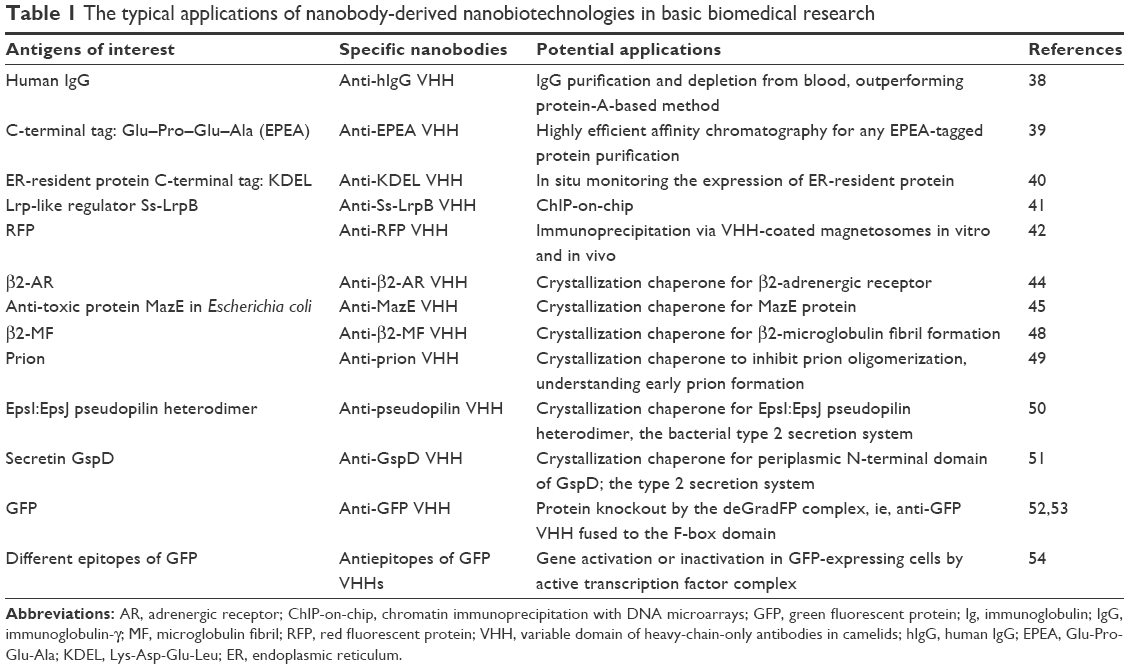 | Table 1 The typical applications of nanobody-derived nanobiotechnologies in basic biomedical research |
Owing to the intrinsic stability, monomeric nature, and easy directional immobilization to solid substrates, the nanobody is considered an ideal ligand for biomolecule purifications. Compared to full-length antibodies, nanobodies could yield a high-column regeneration capacity, produce an increased amount of paratopes per gram of support materials, and only need milder elution conditions.1–3 For example, antihuman IgG VHH has been developed for IgG purification and depletion from blood, outperforming canonical protein A-based method.38 Apart from concave epitopes on properly folded proteins, nanobodies could recognize small linear peptide sequences, which has been confirmed by isolating anti-EPEA VHHs. EPEA is a C-terminal tetra-amino-acid Glu–Pro–Glu–Ala sequence that can be cloned as a tag behind any protein,39 facilitating a rapid and robust affinity purification of proteins. This linear peptide sequence recognition by nanobodies has also been demonstrated by isolating VHHs against another tetra-amino-acid sequence KDEL, a C-terminal signature tag of endoplasmic reticulum-resident protein.40 This anti-KDEL VHH nanobody was shown to be an excellent tool to study differences in ER-resident protein expression by recognizing the KDEL sequence at the C-terminus of proteins, irrespective of the protein context.40
The high affinity and unique specificity of nanobodies also make themselves excellent candidates for immunoprecipitation applications and for chromatin immunoprecipitation with DNA microarray (ChIP-on-chip) technology, facilitating uncovering new transcription factor-binding sites.2,41 Furthermore, by the combination of nanobody and magnetosome, a VHH-coated magnetosome approach was proposed for in vitro and in vivo immunoprecipitation by magnetical recruitment of antigen partners from cell extracts or within living bacteria.42 As membranous organelles present in magnetotactic bacteria, magnetosomes contain magnetite particles enabling orientation of bacteria in a magnetic field.2 By expressing red fluorescent protein (RFP)-binding nanobody with a magnetosome membrane protein MamC, VHH-coated magnetosomes were generated to efficiently recognize and bind their antigens in vitro by magnetically separating the VHH particle–antigen complexes from unbound protein contaminants. Such VHH particles could potentially also be used for intracellular recognition and magnetosome recruitment of RFP-tagged proteins and their interaction partners within living bacteria.
Nanobodies were also used as effective chaperones to assist crystallization process and structural determination of membrane proteins and large protein complexes for which it is difficult to determine these protein structures by X-ray crystallography. Since the convex or protruding surface at the N-terminal end of nanobodies could be targeting and recognizing clefts on the surface of antigens and these clefts usually coincide with active enzymatic sites or ligand-/receptor-binding cavities, the nanobodies are very suitable chaperones to assist crystallization and structural determination of these challenging targets.43,44 It has been demonstrated that, compared to the conventionally full antibodies, such chaperones could facilitate crystal formation by maintaining highly dynamic proteins in one of the particular protein conformations and stabilize intrinsic flexible regions or detergent-solubilized membrane proteins through preventing hydrophobic surfaces from contact with solvent to facilitate the effective crystal formation (Figure 3).45–47 For example, β2-adrenergic receptor crystal has been elucidated in the presence of active state-specific VHH, which could stabilize the instable active state of the receptor,2,44 and a VHH also plays its unique roles as a chaperone for the formation of β2-microglobulin fibril by stabilizing early fibril intermediates and preventing their self-oligomerization.2,48 The structural information on the disordered N-terminal prion protein region has also been elucidated by a VHH-inhibiting prion oligomerization, eventually contributing to the understanding of early prion formation.2,49 Similarly, crystallization process of components of bacterial type 2 secretion system demonstrated that the VHHs could substantially facilitate well-diffracting crystal formation by merely providing additional contact surface to the target proteins.2,50,51
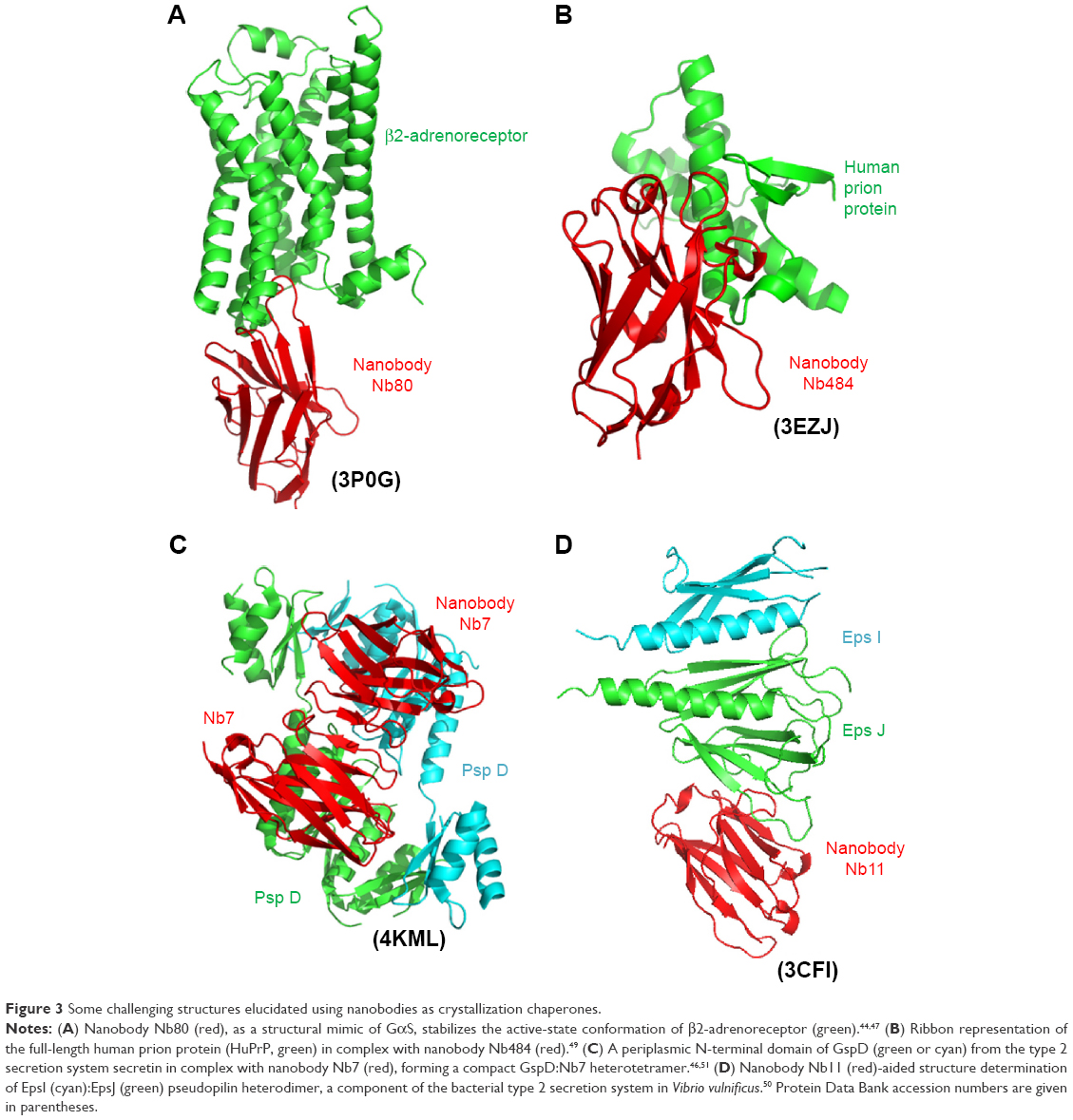 | Figure 3 Some challenging structures elucidated using nanobodies as crystallization chaperones. |
By an elegantly experimental design, another example of the use of nanobody is to trigger the depletion of antigen via the ubiquitin pathway. Caussinus et al52,53 designed a deGradFP nanocomplex to induce the degradation of protein in vivo. A proof of concept study was conducted in fruitfly (Drosophila melanogaster) and zebrafish (Danio rerio). The anti-green fluorescent protein (GFP) nanobody was fused to the F-box domain that recruits the polyubiquitination machinery. Once a target was captured in the ubiquitinated nanobody-based nanocomplex, the proteasome-mediated degradation could be initiated.1,52 This is a new protein knockout tool potentially for more sophisticated applications in biomedicine and biotechnology. The resulting ubiquitinated nanobody-based nanocomplex could be effectively restricted in certain tissues, and the extent of protein degradation could be real-time monitored by just measuring GFP fluorescence.2
Recently, some other elegant works also demonstrated a nanobody-based system using fluorescent proteins as scaffolds for cell-specific gene manipulation.54 Two different nanobodies that could bind different regions of GFP were fused to a transcriptional activation domain or a DNA-binding domain, respectively. As a result, the GFP expression in GFP-expressing cells could specifically lead to the formation of a nanoscaled active transcription factor.54 This approach could be utilized to conveniently induce cell-specific transgene expression or gene knockdown by RNAi in a vast collection of transgenic GFP cell lines.2
Nanobodies as powerful bioimaging reagents
Owing to small dimensional size and high affinity of the nanobodies against various targets of interest, eg, intracellular signaling molecules and cancer biomarkers, the nanobodies and their derivative formats used as versatile nanotracers have been successfully employed in this postgenomic era (Table 2).1 Using fluorescent protein fusion nanobodies or anti-GFP nanobodies, some elegantly fabricated nanobody-based tracers have been developed for bioimaging in living cells. As a proof of concept, intracellular expression of the genetic fusion of a fluorescent protein with a nanobody produces useful chromobodies or fluobodies to trace in vivo intracellular target in various cellular compartments in living cells,55,56 avoiding the need of genetic modification of target proteins with fluorescent tags. For example, an anti-GFP nanobody, termed GFP-binding protein (GBP), was fused to a monomeric RFP to generate a chromobody. The resulting GFP-based chromobody could specifically label the intracellular GFP fusion proteins localized in cytoplasm or nuclear.55 Several of these nanobody-based tracers were also developed for high-content analysis, eg, the direct visualization of endogenous native proteins or infectious HIV virions in living cells.57,58 Furthermore, the GBP is also applied in super-resolution microscopy for the visualization of GFP fusion proteins. By coupling organic dyes to GFP-binding nanobodies, the nanobody-based tracker could recognize any GFP-tagged construct, enabling single-molecule localization with super-resolution imaging techniques.1,59 Similarly, a GBP-coated gold nanoparticle was also employed as a single-molecule tracer to monitor GFP-tagged membrane proteins and is even capable of tracking intracellular proteins in living cells by internalization.2,60
 | Table 2 The representative applications of chromobodies-based nanobiotechnologies for intracellular bioimaging |
Recently, the nanobody-based tracer was applied to study in vivo protein–protein interactions.2 By a sophisticated experimental design, the GBP was first fused to an anchoring protein that will localize the GBP at a particular subcellular compartment. The fluorescent protein GFP or RFP was then fused to the two proteins of interest. Once interaction of the two proteins occurs, both the proteins tethered together will lead to a strong GFP–RFP colocalization signal at the subcellular compartment defined by the GBP. In the nucleus and cytosol of human cells, it has been successfully demonstrated the effectiveness of this approach. By this way, peptide inhibitors of protein–protein interactions in these intracellular environments were analyzed and screened.2,61 More importantly, intracellularly expressed nanobodies themselves remaining soluble and specific antigen recognition activities could also be used to interfere with particular protein functions by competing with normal in situ protein–protein interactions. Thus, the identification of such bioactive nanobodies could offer an opportunity for target validation and lead molecule optimization to investigate difficult interactions or interactions considered undruggable.1,62,63
More interestingly, versatile nanotraps have been developed, by which nanobodies were exploited to report particular conformational variants of a target and even to alter target translocation and localization in different organelles of living cells.64–67 For example, nanobodies could modulate the conformation and spectral properties of GFP. The tamoxifen-induced translocation of human estrogen receptor from the cytoplasm to the nucleus could be monitored in a sensitive, high-throughput manner by nuclear expression of a GFP-enhancing GBP and then by monitoring the ratio of imaging.2,64 Promyelocytic leukemia protein originally dispersed throughout the nucleus, whereas GFP fusion promyelocytic leukemia protein is specifically redirected to the nuclear lamina by coexpressing GBP-lamin1 as a nuclear lamina anchor.2,67 Contrary to trapping target proteins in nuclear lamina, GBP was also demonstrated to block the action of nuclear GFP fusion proteins by trapping them in the cytoplasm of plant cells.2,68
Nanobodies for disease diagnosis
Apart from the use of nanobodies as versatile bioimaging tools in living cells, nanobodies have been used as valuable in vivo detection probes for cancer, infectious disease, atherosclerotic lesions, inflammatory response, and many other diseases in preclinical and clinical environments (Table 3).
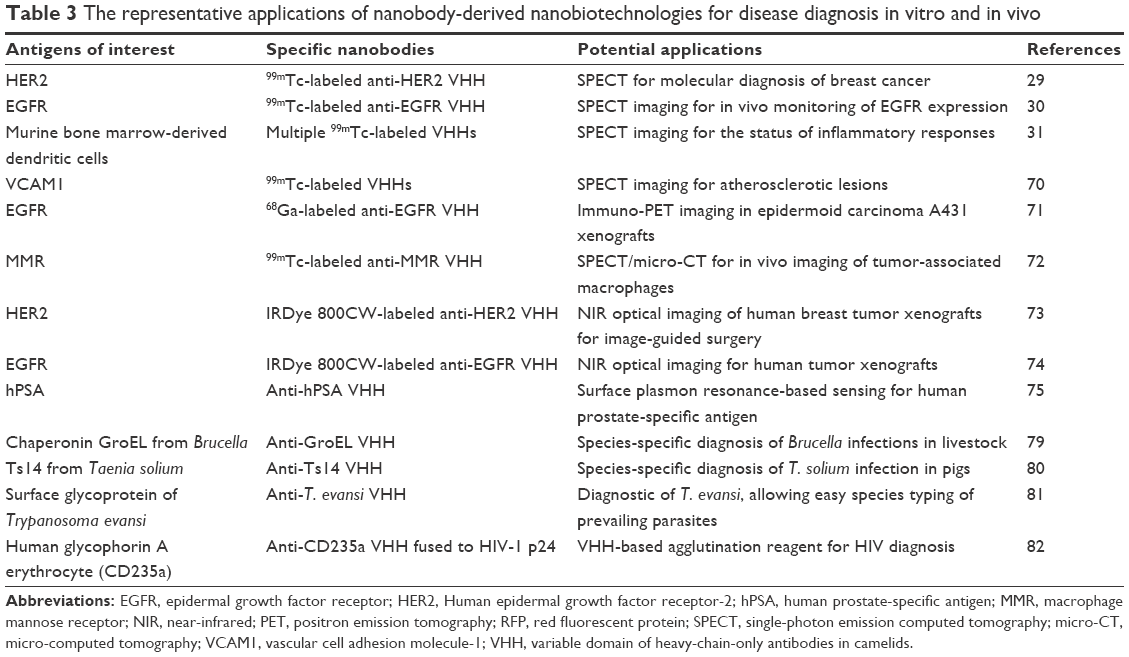 | Table 3 The representative applications of nanobody-derived nanobiotechnologies for disease diagnosis in vitro and in vivo |
The nanobodies could be quickly eliminated from bloodstream due to their small sizes with a molecular weight of ~12–15 kDa that is well below the renal clearance cutoff of molecular weight of ~50 kDa.1,69 Thus, the small-size property of nanobodies facilitates their fast extravasation, good tumor penetration, and rapid renal clearance, which eventually lead to rapid and sensitive imaging of target tissue just within a few hours postinjection.2,29,30 This is requirement exactly for a good in vivo imaging agent.1 Radionuclide-labeled nanobodies have been successfully applied to noninvasively image in vivo tumors via positron emission tomography and single-photon emission computed tomography. These nanobody-derived nanoprobes demonstrated a high target specificity, high stability, good solubility, high tumor-to-background signal, fast clearance of excess tracer, and low immunogenicity.2 For example, 99mTc-labeled VHHs could be used to recognize HER2, a cancer antigen for breast cancer diagnosis in a preclinical setting,29 to distinguish moderate and high expression of epidermal growth factor receptor for improved prognosis of cancer therapy,30 to detect the status of inflammatory responses by imaging dendritic cells,31 and to target VCAM1, an antigen used to diagnose vulnerable atherosclerotic plaques.70
Besides the promising results obtained by using 99mTc-VHH with half-life time (t1/2) of ~6 hours, some short-lived nuclides were also tested for more patient-friendly diagnoses. Rapid targeting to diseased tissue and fast blood clearance of unbound nanobodies make it possible to use short-lived nuclide-labeled formats for in vivo diagnosis, such as 68Ga and 18F with the t1/2 of 68 minutes and 110 minutes, respectively. This approach makes it possible to measure picomolar concentrations of nanoprobe within 1–3 hours postinjection by using positron emission tomography and single-photon emission computed tomography imaging techniques, thus resulting in a very low radiation burden for patients.1 Using a mouse epidermoid carcinoma A431 xenograft model, the 68Ga-labeled nanobodies against an epidermal growth factor receptor were recently tested for their performances, yielding a relatively high ratio of tumor to blood after 3 hours postinjection.1,71
Owing to rapid renal clearance of nanobodies, the disadvantage of using nanobodies as in vivo probes is high accumulation of unbound nanobodies in kidneys soon after the probe injection, finally leading to a high radiation dose in kidneys and a complicated analysis of nearby tissues.2,29,31,70 A method has been further proposed to reduce the background level encountered with radiolabeled VHHs by preinjection of unlabeled bivalent VHHs to first occupy all extratumoral sites.72 Alternatively, nonradioactive VHH-based probes with near-infrared fluorophores have also been fabricated to image in vivo tumors. These near-infrared fluorophore-labeled VHHs also showed faster imaging compared to approved monoclonal antibodies (McAbs) targeting the same antigens.2,73,74
Like a coin has two sides, the small size of nanobodies, however, might affect antigen–probe interaction when they are coated in adsorptive substrates in an in vitro enzyme linked immunosorbent assay (ELISA) for disease biomarkers in blood or other biopsies. Compared to traditional IgGs, when these small-size nanobodies are coated on adsorptive substrates, the paratopes of nanobodies are very close to the vicinity of adsorbing surface, which might hinder antigen–probe interaction.75 C-terminal peptide extension could improve the accessibility of coated VHHs in ELISA, eg, by fusion to peptide fragments, including a myc-His-tag, a llama long hinge region-His-tag,76 and an Fc chain.2,75 Through this way, the engineered camel sdAbs were immobilized for sensing human prostate-specific antigen, which demonstrated that a higher probe density mediates enhanced detection sensitivity in a surface plasmon resonance-based detection system.75,77 This means that the use of nanobodies to generate sensitive and selective biosensors for in vitro disease diagnosis is highly feasible.2,75
More particularly, the nanobodies have also been developed to detect disease biomarkers in human biopsies by antibody-based slide and bead arrays. For an instance, a biotinylated VHH has been applied to streptavidin beads for sensitive biomarker detection in patient sera.2,78 As for pathogen diagnosis, traditionally McAb-derived antibodies have been used for decades. Although these McAb-derived formats recognize pathogen antigens with high sensitivity, they often lack the required specificity, leading to unsatisfactory performances. It has been demonstrated that the performances of nanobodies far exceed traditional McAbs. For example, Brucella and Yersinia infections in livestock have been successfully distinguished by a VHH rather than conventional antibodies.79 Similarly, Taenia solium infection in pigs could be successfully detected by species-specific VHHs, whereas the existing genus-specific McAbs have failed to discriminate between T. solium and Taenia hydatigena infections.80 Both species-specific and genus-specific VHHs have been developed for the diagnosis of Trypanosoma parasites without purification of antigens, leading to easy species typing of the prevailing parasites.2,81 Recently, a general nanobody-based agglutination reagent, consisting of a fusion protein between a red blood cell-specific VHH and a disease antigen of interest, has been developed for diagnosing a variety of diseases when different disease-specific antigens are available. This elegantly designed system could be used for HIV diagnosis when an HIV-1 p24 antigen was fused with red blood cell-specific VHH.82 Thus, agglutination mediated by anti-p24 antibodies in patient serum and the added p24-VHH fusion protein could be observed if the anti-p24 antibodies are present in HIV-positive patient sera.2
Nanobodies as targeting therapeutics
The development of nanobodies as targeting therapeutics is still in a very early stage. Some elegant works have demonstrated that the use of the bioactive nanobodies for antitoxin, anti-infection, anti-inflammation, or enzyme inhibition is a potentially feasible way for novel therapeutic development (Table 4). For example, nanobodies have been evaluated for passive immunization to treat envenomed victims, demonstrating extreme high-neutralization potency. By far, nanobodies for antiscorpion toxins, antibacterial toxins, and anti-snake venom are actively being investigated.83–87 Owing to their small size and extended CDR3, nanobodies also showed special advantages as therapeutics for infectious disease, including the infection of viruses, bacteria, and parasites, over conventional antibodies that usually obstruct the access of hidden and essential epitopes on pathogens.88–96 The added value of the nanobodies as targeting therapeutics stems from their capacity to distinguish the cognate target from closely related variants. Most of the small organic antagonists or even the conventional antibodies to a larger extent cannot reach such high specificity. Therefore, nanobodies could be used to specifically inhibit unwanted enzymatic activities related to different cellular signaling pathways. For example, ecto-ADP-ribosyl transferase ART2.2-specific VHH effectively blocked the enzymatic and cytotoxic activities of ART2.2 in lymphatic organs following intravenous injection. This blockade was highly specific, ie, only targeting blockade of enzyme ART2.2 but not the related enzymes ART1 or ART2.1, which eventually leads to develop new antidotes against ADP-ribosylating toxins.97 In addition, several other nanobody-derived therapeutics are already in the pipeline. For instances, anti-IL6R and anti-tumor necrosis factor α nanobodies were developed to treat rheumatoid arthritis,98–100 and an anti-RANKL nanobody was generated for bone loss disorder therapy.1 Some therapeutic nanobodies, such as an anti-von Willebrand factor nanobody for the treatment of acute thrombotic thrombocytopenic purpura, have passed Phase I and Phase II clinical trials.101 However, there is expectedly high competition from other therapeutics, including conventional McAbs and other antibody-derived small-size formats, such as scFvs and Fabs.1
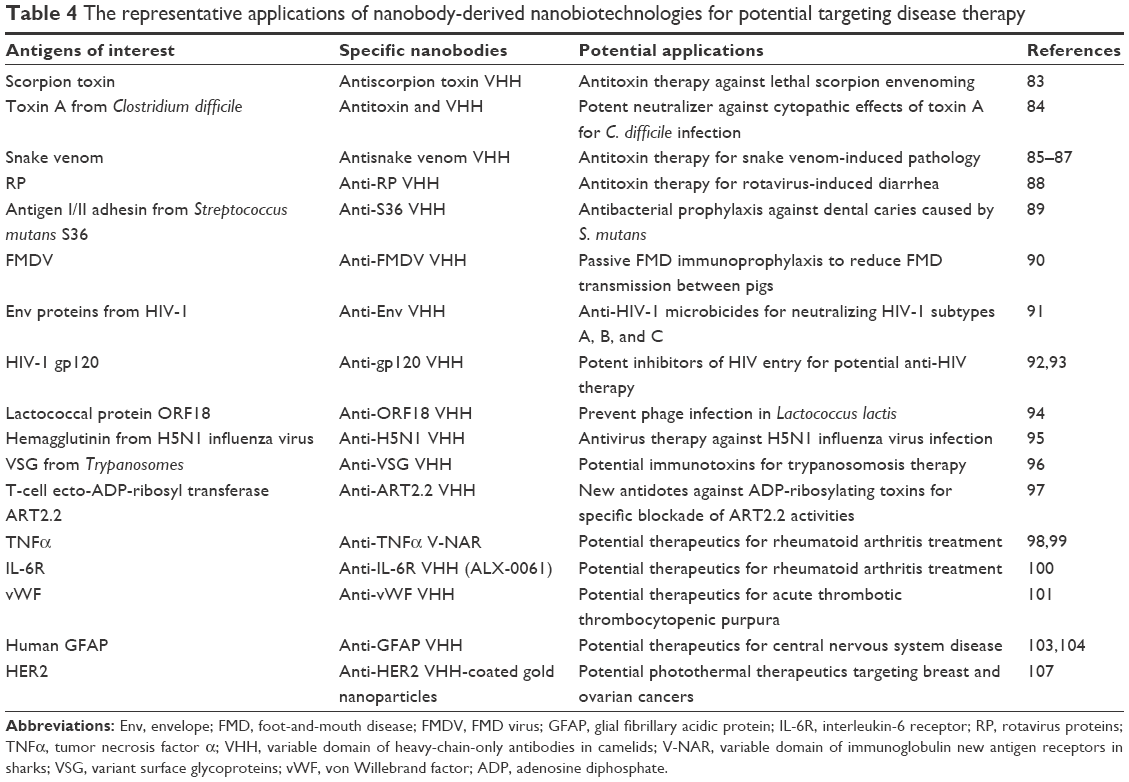 | Table 4 The representative applications of nanobody-derived nanobiotechnologies for potential targeting disease therapy |
On the other hand, the success of nanobodies as targeting therapeutics in clinic environments will also come from a more patient friendly administration, such as topical, oral, and inhalation or targeting delivery of cargoes to tissues that are difficult to access, and from obtaining improved biopharmaceutical parameters, including blood concentrations and prolonged residence time.1,102 To this end, nanobodies have been extensively used for targeting delivery of cargoes to different targeting antigens in in vivo organs to improve their biopharmaceutical parameters. For example, nanobodies have been assessed in their capacity of crossing blood–brain barrier and transcytosis across epithelia, which will potentially lead to develop nanobody-based brain-targeting therapeutics.103–106 Moreover, chemical conjugation of nanobodies to branched gold nanoparticles also effectively produced antigen-targeting photothermal therapeutics upon light irradiation in a near biological window.107
Nanobodies for agricultural protection and food analysis
Besides the promising results from enzyme inhibition exerted by nanobodies in mammals, nanobodies could also be used as enzyme inhibitors to modulate plant physiology and increase starch content in plant. For example, an inhibitory VHH against starch-branching enzyme A has been generated to interfere with the function of enzymes in potato.108 Because of specific enzyme inhibition by an inhibitory VHH, the content of amylose in potato tuber increases upon targeting the VHH to plastids. An increase in amylose is even higher than that of the antisense controls, demonstrating the unique performance of the nanobodies as plant enzyme inhibitors. Furthermore, nanobodies could be used as nanocarriers for more efficient and specific delivery of existing chemicals to crops and weeds. To this respect, plant-specific VHHs were covalently coupled to engineered agrocapsules to improve the chemical retention time at target sites and reduce the chemical amounts needed.109 The plant-specific VHHs have also been generated that could bind stomata, trichomes, grass leaves, and the surface area of potato.32 Importantly, these plant-specific VHHs also showed good tolerance for harsh field conditions, such as temperature, variable pH, and salt concentrations. Besides the crops and weeds, nanobodies have also been applied for specific and targeting delivery of insecticides to insects.33,109 To pursue a high-specific VHH, the VHH phages are selected directly against living aphids and even whole-insect ELISA has been developed to characterize these insect-specific VHHs. Interestingly, nanobodies also been applied to food analysis because of highly extreme themostability. Several anticaffeine VHHs have been already generated for the quantification of caffeine in hot beverages. At 70°C, one of these VHHs could recognize caffeine and amazingly recover its binding functionality after an incubation step at 90°C,110 demonstrating the excellent thermostability of these nanobodies.
Conclusion and perspectives
In this postgenomic era, searching for picomolar affinity and high specific binders against different targets of interest, including proteins, peptides, DNAs, RNAs, polysaccharides, and small molecules, are overwhelming challenges. Compared to conventional antibodies, nanobodies have following excellent properties: nanoscale size, robust structure, stable and soluble behavior in aqueous solution, reversible refolding, high affinity and specificity for only one cognate target, superior cryptic cleft accessibility, and deep tissue penetration, as well as a sustainable source. It is the amazingly excellent properties of nanobodies that could meet the unmet demand for screening these novel types of binders from renewable nanobody resources, making them attractive alternatives to conventional antibodies and their single-domain formats, such as Fab and scFv. The nanobody-derived binders have already been demonstrated to not only recognize only one cognate target of interest with high affinity but also differentiate different conformations of these targets with superior specificity. Therefore, these unique properties of nanobodies offer us opportunities to develop specific nanobiotechnological tool kits for various biomedical and biotechnological applications.
Current research on applying the nanobody-derived nanobiotechnological tool kits in affinity purification, immunoprecipitation, chaperone-assisted crystallization, protein degradation, gene activation or inactivation, protein–protein interactions, cellular bioimaging, in vivo and in vitro disease diagnosis, targeting therapeutics, agricultural and plant protection, and food analysis indicate a promising future of the use of these nanobodies and their derivative formats in biomedical and biotechnological fields. In the future, application of nanobodies as versatile molecules in different fields, including targeting therapy, targeting delivery, immunosensors, and in vivo imaging, nano-based research tools, is highly anticipated. Among these future trends, commercialization of nanobodies as a next-generation targeting therapeutics is high priority and technical challenge.111,112 Although some nanobody-derived products are in the pipeline, some have passed Phase I and Phase II trials,1,101 the competition from conventional McAbs or other antibody-derived small-size formats is huge. As compared to conventional McAbs, the small size of nanobodies not only confers their good tumor penetration but also enables them for a rapid renal clearance, indicating the challenge to balance improved systemic distribution with decreased plasma half-life in designing nanobody-based targeting therapeutics.111 Therefore, for some intended uses, properties and pharmacokinetics should be tailored by linking nanobodies to albumin-binding moieties or by changing their hydrodynamic volume in various ways to reach a high blood concentration over a prolonged blood residence time,1,102 ie, obtaining long-circulating nanobody-based therapeutics. Furthermore, several other obstacles, such as immunogenicity and functionalization,3 still have to be clinically addressed before the use of nanobodies in the clinic as targeting therapeutics become feasible.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (YW, grant no 31470218; LY, grant no 81572022). The authors also thank the anonymous reviewers for their insightful suggestions on improving the original article.
Disclosure
The authors report no conflicts of interest in this work.
References
Muyldermans S. Nanobodies: natural single-domain antibodies. Annu Rev Biochem. 2013;82:775–797. | ||
De Meyer T, Muyldermans S, Depicker A. Nanobody-based products as research and diagnostic tools. Trends Biotechnol. 2014;32(5):263–270. | ||
Siontorou CG. Nanobodies as novel agents for disease diagnosis and therapy. Int J Nanomedicine. 2013;8:4215–4227. | ||
Saerens D, Muyldermans S. Single Domain Antibodies: Methods and Protocols. Totowa, NJ: Humana Press; 2012. | ||
Zielonka S, Empting M, Grzeschik J, Konning D, Barelle CJ, Kolmar H. Structural insights and biomedical potential of IgNAR scaffolds from sharks. MAbs. 2015;7(1):15–25. | ||
Flajnik MF, Kasahara M. Origin and evolution of the adaptive immune system: genetic events and selective pressures. Nat Rev Genet. 2010;11(1):47–59. | ||
Shao CY, Secombes CJ, Porter AJ. Rapid isolation of IgNAR variable single-domain antibody fragments from a shark synthetic library. Mol Immunol. 2007;44(4):656–665. | ||
Liu JL, Anderson GP, Delehanty JB, Baumann R, Hayhurst A, Goldman ER. Selection of cholera toxin specific IgNAR single-domain antibodies from a naive shark library. Mol Immunol. 2007;44(7):1775–1783. | ||
Dooley H, Flajnik MF. Antibody repertoire development in cartilaginous fish. Dev Comp Immunol. 2006;30(1–2):43–56. | ||
Muyldermans S, Baral TN, Retamozzo VC, et al. Camelid immunoglobulins and nanobody technology. Vet Immunol Immunopathol. 2009;128(1–3):178–183. | ||
Helma J, Cardoso MC, Muyldermans S, Leonhardt H. Nanobodies and recombinant binders in cell biology. J Cell Biol. 2015;209(5):633–644. | ||
Muyldermans S, Atarhouch T, Saldanha J, Barbosa JA, Hamers R. Sequence and structure of VH domain from naturally occurring camel heavy chain immunoglobulins lacking light chains. Protein Eng. 1994;7(9):1129–1135. | ||
Vu KB, Ghahroudi MA, Wyns L, Muyldermans S. Comparison of llama VH sequences from conventional and heavy chain antibodies. Mol Immunol. 1997;34(16–17):1121–1131. | ||
Harmsen MM, Ruuls RC, Nijman IJ, Niewold TA, Frenken LGJ, de Geus B. Llama heavy-chain V regions consist of at least four distinct subfamilies revealing novel sequence features. Mol Immunol. 2000;37(10):579–590. | ||
Maass DR, Sepulveda J, Pernthaner A, Shoemaker CB. Alpaca (Lama pacos) as a convenient source of recombinant camelid heavy chain antibodies (VHHs). J Immunol Methods. 2007;324(1–2):13–25. | ||
Govaert J, Pellis M, Deschacht N, et al. Dual beneficial effect of interloop disulfide bond for single domain antibody fragments. J Biol Chem. 2012;287(3):1970–1979. | ||
Criscitiello MF, Saltis M, Flajnik MF. An evolutionarily mobile antigen receptor variable region gene: doubly rearranging NAR-TcR genes in sharks. Proc Natl Acad Sci U S A. 2006;103(13):5036–5041. | ||
Liu JL, Anderson GP, Goldman ER. Isolation of anti-toxin single domain antibodies from a semi-synthetic spiny dogfish shark display library. BMC Biotechnol. 2007;7:78–87. | ||
Stanfield RL, Dooley H, Verdino P, Flajnik MF, Wilson IA. Maturation of shark single-domain (IgNAR) antibodies: evidence for induced-fit binding. J Mol Biol. 2007;367(2):358–372. | ||
Stanfield RL, Dooley H, Flajnik MF, Wilson IA. Crystal structure of a shark single-domain antibody V region in complex with lysozyme. Science. 2004;305(5691):1770–1773. | ||
Desmyter A, Transue TR, Ghahroudi MA, et al. Crystal structure of a camel single-domain VH antibody fragment in complex with lysozyme. Nat Struct Biol. 1996;3(9):803–811. | ||
De Genst E, Silence K, Decanniere K, et al. Molecular basis for the preferential cleft recognition by dromedary heavy-chain antibodies. Proc Natl Acad Sci U S A. 2006;103(12):4586–4591. | ||
Desmyter A, Decanniere K, Muyldermans S, Wyns L. Antigen specificity and high affinity binding provided by one single loop of a camel single-domain antibody. J Biol Chem. 2001;276(28):26285–26290. | ||
Spinelli S, Tegoni M, Frenken L, van Vliet C, Cambillau C. Lateral recognition of a dye hapten by a llama VHH domain. J Mol Biol. 2001;311(1):123–129. | ||
Spinelli S, Frenken LG, Hermans P, et al. Camelid heavy-chain variable domains provide efficient combining sites to haptens. Biochemistry. 2000;39(6):1217–1222. | ||
Holliger P, Hudson PJ. Engineered antibody fragments and the rise of single domains. Nat Biotechnol. 2005;23(9):1126–1136. | ||
Goldman ER, Anderson GP, Liu JL, et al. Facile generation of heat-stable antiviral and antitoxin single domain antibodies from a semisynthetic llama library. Anal Chem. 2006;78(24):8245–8255. | ||
Huang L, Muyldermans S, Saerens D. Nanobodies(R): proficient tools in diagnostics. Expert Rev Mol Diagn. 2010;10(6):777–785. | ||
Vaneycken I, Devoogdt N, Van Gassen N, et al. Preclinical screening of anti-HER2 nanobodies for molecular imaging of breast cancer. FASEB J. 2011;25(7):2433–2446. | ||
Huang L, Gainkam LOT, Caveliers V, et al. SPECT imaging with Tc-99m-labeled EGFR-specific nanobody for in vivo monitoring of EGFR expression. Mol Imaging Biol. 2008;10(3):167–175. | ||
De Groeve K, Deschacht N, De Koninck C, et al. Nanobodies as tools for in vivo imaging of specific immune cell types. J Nucl Med. 2010;51(5):782–789. | ||
Jongedijk E, Verheesen P, inventor; AgroSavfe NV, assignee (Gent, BE). Specific delivery of agrochemicals. US patent 8598081. Dec 3, 2013. | ||
Verheesen P, Jongedijk E, inventors; AgroSavfe NV, assignee (Gent, BE). Insect binding antibodies. US patent 20130224226. Aug 29, 2013. | ||
Harmsen MM, De Haard HJ. Properties, production, and applications of camelid single-domain antibody fragments. Appl Microbiol Biotechnol. 2007;77(1):13–22. | ||
Wesolowski J, Alzogaray V, Reyelt J, et al. Single domain antibodies: promising experimental and therapeutic tools in infection and immunity. Med Microbiol Immunol. 2009;198(3):157–174. | ||
de Marco A. Biotechnological applications of recombinant single-domain antibody fragments. Microb Cell Fact. 2011;10:44. | ||
Dmitriev OY, Lutsenko S, Muyldermans S. Nanobodies as probes for protein dynamics in vitro and in cells. J Biol Chem. 2016;291(8):3767–3775. | ||
Klooster R, Maassen BTH, Stam JC, et al. Improved anti-IgG and HSA affinity ligands: clinical application of VHH antibody technology. J Immunol Methods. 2007;324(1–2):1–12. | ||
De Genst EJ, Guilliams T, Wellens J, et al. Structure and properties of a complex of alpha-synuclein and a single-domain camelid antibody. J Mol Biol. 2010;402(2):326–343. | ||
Klooster R, Eman MR, le Duc Q, et al. Selection and characterization of KDEL-specific VHH antibody fragments and their application in the study of ER resident protein expression. J Immunol Methods. 2009;342(1–2):1–12. | ||
Nguyen-Duc T, Peeters E, Muyldermans S, Charlier D, Hassanzadeh-Ghassabeh G. Nanobody(R)-based chromatin immunoprecipitation/micro-array analysis for genome-wide identification of transcription factor DNA binding sites. Nucleic Acids Res. 2013;41(5):e59. | ||
Pollithy A, Romer T, Lang C, et al. Magnetosome expression of functional camelid antibody fragments (nanobodies) in Magnetospirillum gryphiswaldense. Appl Environ Microbiol. 2011;77(17):6165–6171. | ||
Baranova E, Fronzes R, Garcia-Pino A, et al. SbsB structure and lattice reconstruction unveil Ca2+ triggered S-layer assembly. Nature. 2012;487(7405):119–122. | ||
Rasmussen SGF, Choi H-J, Fung JJ, et al. Structure of a nanobody-stabilized active state of the beta(2) adrenoceptor. Nature. 2011;469(7329):175–180. | ||
Loris R, Marianovsky I, Lah J, et al. Crystal structure of the intrinsically flexible addiction antidote MazE. J Biol Chem. 2003;278(30):28252–28257. | ||
Koide S. Engineering of recombinant crystallization chaperones. Curr Opin Struct Biol. 2009;19(4):449–457. | ||
Pardon E, Laeremans T, Triest S, et al. A general protocol for the generation of nanobodies for structural biology. Nat Protoc. 2014;9(3):674–693. | ||
Domanska K, Vanderhaegen S, Srinivasan V, et al. Atomic structure of a nanobody-trapped domain-swapped dimer of an amyloidogenic beta 2-microglobulin variant. Proc Natl Acad Sci U S A. 2011;108(4):1314–1319. | ||
Abskharon RNN, Giachin G, Wohlkonig A, et al. Probing the N-terminal beta-sheet conversion in the crystal structure of the human prion protein bound to a nanobody. JACS. 2014;136(3):937–944. | ||
Lam AY, Pardon E, Korotkov KV, Hol WGJ, Steyaert J. Nanobody-aided structure determination of the EpsI:EpsJ pseudopilin heterodimer from Vibrio vulnificus. J Struct Biol. 2009;166(1):8–15. | ||
Korotkov KV, Pardon E, Steyaert J, Hol WGJ. Crystal structure of the N-terminal domain of the secretin GspD from ETEC determined with the assistance of a nanobody. Structure. 2009;17(2):255–265. | ||
Caussinus E, Kanca O, Affolter M. Fluorescent fusion protein knockout mediated by anti-GFP nanobody. Nat Struct Mol Biol. 2011;19(1):117–121. | ||
Caussinus E, Kanca O, Affolter M. Protein knockouts in living eukaryotes using deGradFP and green fluorescent protein fusion targets. Curr Protoc Protein Sci. 2013;73(73):Unit30.2. | ||
Tang JCY, Szikra T, Kozorovitskiy Y, et al. A nanobody-based system using fluorescent proteins as scaffolds for cell-specific gene manipulation. Cell. 2013;154(4):928–939. | ||
Rothbauer U, Zolghadr K, Tillib S, et al. Targeting and tracing antigens in live cells with fluorescent nanobodies. Nat Methods. 2006;3(11):887–889. | ||
Olichon A, Surrey T. Selection of genetically encoded fluorescent single domain antibodies engineered for efficient expression in Escherichia coli. J Biol Chem. 2007;282(50):36314–36320. | ||
Zolghadr K, Gregor J, Leonhardt H, Rothbauer U. Case study on live cell apoptosis-assay using lamin-chromobody cell-lines for high-content analysis. Methods Mol Biol. 2012;911:569–575. | ||
Helma J, Schmidthals K, Lux V, et al. Direct and dynamic detection of HIV-1 in living cells. PLoS One. 2012;7(11):E50026. | ||
Ries J, Kaplan C, Platonova E, Eghlidi H, Ewers H. A simple, versatile method for GFP-based super-resolution microscopy via nanobodies. Nat Methods. 2012;9(6):582–584. | ||
Leduc C, Si S, Gautier J, et al. A highly specific gold nanoprobe for live-cell single-molecule imaging. Nano Lett. 2013;13(4):1489–1494. | ||
Herce HD, Deng W, Helma J, Leonhardt H, Cardoso MC. Visualization and targeted disruption of protein interactions in living cells. Nat Commun. 2013;4:2660. | ||
Tanaka T, Sewell H, Waters S, Phillips SEV, Rabbitts TH. Single domain intracellular antibodies from diverse libraries: emphasizing dual functions of LMO2 protein interactions using a single VH domain. J Biol Chem. 2011;286(5):3707–3716. | ||
Perez-Martinez D, Tanaka T, Rabbitts TH. Intracellular antibodies and cancer: new technologies offer therapeutic opportunities. Bioessays. 2010;32(7):589–598. | ||
Kirchhofer A, Helma J, Schmidthals K, et al. Modulation of protein properties in living cells using nanobodies. Nat Struct Mol Biol. 2010;17(1):133–138. | ||
Romer T, Leonhardt H, Rothbauer U. Engineering antibodies and proteins for molecular in vivo imaging. Curr Opin Biotechnol. 2011;22(6):882–887. | ||
Schmidthals K, Helma J, Zolghadr K, Rothbauer U, Leonhardt H. Novel antibody derivatives for proteome and high-content analysis. Anal Bioanal Chem. 2010;397(8):3203–3208. | ||
Rothbauer U, Zolghadr K, Muyldermans S, Schepers A, Cardoso MC, Leonhardt H. A versatile nanotrap for biochemical and functional studies with fluorescent fusion proteins. Mol Cell Proteomics. 2008;7(2):282–289. | ||
Schornack S, Fuchs R, Huitema E, Rothbauer U, Lipka V, Kamoun S. Protein mislocalization in plant cells using a GFP-binding chromobody. Plant J. 2009;60(4):744–754. | ||
Vaneycken I, D’Huyvetter M, Hernot S, et al. Immuno-imaging using nanobodies. Curr Opin Biotechnol. 2011;22(6):877–881. | ||
Broisat A, Hernot S, Toczek J, et al. Nanobodies targeting mouse/human VCAM1 for the nuclear imaging of atherosclerotic lesions. Circ Res. 2012;110(7):927–937. | ||
Vosjan MJWD, Perk LR, Roovers RC, et al. Facile labelling of an anti-epidermal growth factor receptor nanobody with Ga-68 via a novel bifunctional desferal chelate for immuno-PET. Eur J Nucl Med Mol Imaging. 2011;38(4):753–763. | ||
Movahedi K, Schoonooghe S, Laoui D, et al. Nanobody-based targeting of the macrophage mannose receptor for effective in vivo imaging of tumor-associated macrophages. Cancer Res. 2012;72(16):4165–4177. | ||
Kijanka M, Warnders F-J, El Khattabi M, et al. Rapid optical imaging of human breast tumour xenografts using anti-HER2 VHHs site-directly conjugated to IRDye 800CW for image-guided surgery. Eur J Nucl Med Mol Imaging. 2013;40(11):1718–1729. | ||
Oliveira S, van Dongen GAMS, Stigter-van Walsum M, et al. Rapid visualization of human tumor xenografts through optical imaging with a near-infrared fluorescent anti-epidermal growth factor receptor nanobody. Mol Imaging. 2012;11(1):33–46. | ||
Saerens D, Frederix F, Reekmans G, et al. Engineering camel single-domain antibodies and immobilization chemistry for human prostate-specific antigen sensing. Anal Chem. 2005;77(23):7547–7555. | ||
Harmsen MM, Fijten HPD. Improved functional immobilization of llama single-domain antibody fragments to polystyrene surfaces using small peptides. J Immunoassay Immunochem. 2012;33(3):234–251. | ||
Saerens D, Huang L, Bonroy K, Muyldermans S. Antibody fragments as probe in biosensor development. Sensors. 2008;8(8):4669–4686. | ||
Even-Desrumeaux K, Baty D, Chames P. Strong and oriented immobilization of single domain antibodies from crude bacterial lysates for high-throughput compatible cost-effective antibody array generation. Mol Biosyst. 2010;6(11):2241–2248. | ||
Abbady AQ, Al-Daoude A, Al-Mariri A, Zarkawi M, Muyldermans S. Chaperonin GroEL a Brucella immunodominant antigen identified using nanobody and MALDI-TOF-MS technologies. Vet Immunol Immunopathol. 2012;146(3–4):254–263. | ||
Deckers N, Saerens D, Kanobana K, et al. Nanobodies, a promising tool for species-specific diagnosis of Taenia solium cysticercosis. Int J Parasitol. 2009;39(5):625–633. | ||
Saerens D, Stijlemans B, Baral TN, et al. Parallel selection of multiple anti-infectome nanobodies without access to purified antigens. J Immunol Methods. 2008;329(1–2):138–150. | ||
Habib I, Smolarek D, Hattab C, et al. VHH (nanobody) directed against human glycophorin A: a tool for autologous red cell agglutination assays. Anal Biochem. 2013;438(1):82–89. | ||
Hmila I, Saerens D, Ben Abderrazek R, et al. A bispecific nanobody to provide full protection against lethal scorpion envenoming. FASEB J. 2010;24(9):3479–3489. | ||
Hussack G, Arbabi-Ghahroudi M, van Faassen H, et al. Neutralization of Clostridium difficile toxin A with single-domain antibodies targeting the cell receptor binding domain. J Biol Chem. 2011;286(11):8961–8976. | ||
Stewart CS, MacKenzie CR, Hall JC. Isolation, characterization and pentamerization of alpha-cobrotoxin specific single-domain antibodies from a naive phage display library: preliminary findings for antivenom development. Toxicon. 2007;49(5):699–709. | ||
Cook DAN, Samarasekara CL, Wagstaff SC, Kinne J, Wernery U, Harrison RA. Analysis of camelid IgG for antivenom development: immunoreactivity and preclinical neutralisation of venom-induced pathology by IgG subclasses, and the effect of heat treatment. Toxicon. 2010;56(4):596–603. | ||
Harrison RA, Hasson SS, Harmsen M, Laing GD, Conrath K, Theakston RDG. Neutralisation of venom-induced haemorrhage by IgG from camels and llamas immunised with viper venom and also by endogenous, non-IgG components in camelid sera. Toxicon. 2006;47(3):364–368. | ||
Pant N, Marcotte H, Hermans P, et al. Lactobacilli producing bispecific llama-derived anti-rotavirus proteins in vivo for rotavirus-induced diarrhea. Future Microbiol. 2011;6(5):583–593. | ||
Kruger C, Hultberg A, Marcotte H, et al. Therapeutic effect of llama derived VHH fragments against Streptococcus mutans on the development of dental caries. Appl Microbiol Biotechnol. 2006;72(4):732–737. | ||
Harmsen MM, Fijten HPD, Engel B, Dekker A, Eble PL. Passive immunization with llama single-domain antibody fragments reduces foot-and-mouth disease transmission between pigs. Vaccine. 2009;27(13):1904–1911. | ||
Strokappe N, Szynol A, Aasa-Chapman M, et al. Llama antibody fragments recognizing various epitopes of the CD4bs neutralize a broad range of HIV-1 subtypes A, B and C. PLoS One. 2012;7(3):e33298. | ||
Koh WWL, Steffensen S, Gonzalez-Pajuelo M, et al. Generation of a family-specific phage library of llama single chain antibody fragments that neutralize HIV-1. J Biol Chem. 2010;285(25):19116–19124. | ||
Hinz A, Hulsik DL, Forsman A, et al. Crystal structure of the neutralizing llama V-HH D7 and its mode of HIV-1 gp120 interaction. PLoS One. 2010;5(5):e10482. | ||
De Haard HJW, Bezemer S, Ledeboer AM, et al. Llama antibodies against a lactococcal protein located at the tip of the phage tail prevent phage infection. J Bacteriol. 2005;187(13):4531–4541. | ||
Ibanez LI, De Filette M, Hultberg A, et al. Nanobodies with in vitro neutralizing activity protect mice against H5N1 influenza virus infection. J Infect Dis. 2011;203(8):1063–1072. | ||
Stijlemans B, Conrath K, Cortez-Retamozo V, et al. Efficient targeting of conserved cryptic epitopes of infectious agents by single domain antibodies – African trypanosomes as paradigm. J Biol Chem. 2004;279(2):1256–1261. | ||
Koch-Nolte F, Reyelt J, Schossow B, et al. Single domain antibodies from llama effectively and specifically block T cell ecto-ADP-ribosyltransferase ART2.2 in vivo. FASEB J. 2007;21(13):3490–3498. | ||
Camacho-Villegas T, Mata-Gonzalez T, Paniagua-Solis J, Sanchez E, Licea A. Human TNF cytokine neutralization with a vNAR from Heterodontus francisci shark: a potential therapeutic use. MAbs. 2013;5(1):80–85. | ||
Bojalil R, Mata-Gonzalez MT, Sanchez-Munoz F, et al. Anti-tumor necrosis factor (V)NAR single domains reduce lethality and regulate underlying inflammatory response in a murine model of endotoxic shock. BMC Immunol. 2013;14:17. | ||
Van Roy M, Ververken C, Beirnaert E, et al. The preclinical pharmacology of the high affinity anti-IL-6R Nanobody® ALX-0061 supports its clinical development in rheumatoid arthritis. Arthritis Res Ther. 2015;17:135. | ||
Holz J-B. The TITAN trial – assessing the efficacy and safety of an anti-von Willebrand factor nanobody in patients with acquired thrombotic thrombocytopenic purpura. Trans Apher Sci. 2012;46(3):343–346. | ||
Kontermann RE. Strategies for extended serum half-life of protein therapeutics. Curr Opin Biotechnol. 2011;22(6):868–876. | ||
Li T, Bourgeois J-P, Celli S, et al. Cell-penetrating anti-GFAP VHH and corresponding fluorescent fusion protein VHH-GFP spontaneously cross the blood-brain barrier and specifically recognize astrocytes: application to brain imaging. FASEB J. 2012;26(10):3969–3979. | ||
Perruchini C, Pecorari F, Bourgeois J-P, Duyckaerts C, Rougeon F, Lafaye P. Llama VHH antibody fragments against GFAP: better diffusion in fixed tissues than classical monoclonal antibodies. Acta Neuropathol. 2009;118(5):685–695. | ||
Iqbal U, Trojahn U, Albaghdadi H, et al. Kinetic analysis of novel mono- and multivalent VHH-fragments and their application for molecular imaging of brain tumours. Br J Pharmacol. 2010;160(4):1016–1028. | ||
Emmerson CD, van der Vlist EJ, Braam MR, et al. Enhancement of polymeric immunoglobulin receptor transcytosis by biparatopic VHH. PLoS One. 2011;6(10):e26299. | ||
Van de Broek B, Devoogdt N, D’Hollander A, et al. Specific cell targeting with nanobody conjugated branched gold nanoparticles for photothermal therapy. ACS Nano. 2011;5(6):4319–4328. | ||
Jobling SA, Jarman C, Teh MM, Holmberg N, Blake C, Verhoeyen ME. Immunomodulation of enzyme function in plants by single-domain antibody fragments. Nat Biotechnol. 2003;21(1):77–80. | ||
Verheesen P, De Jonghe C, inventor; AgroSavfe NV, assignee (Gent, BE). Manufacturing of specifically targeting microcapsules. US patent 20150087517. Mar 26, 2015. | ||
Ladenson RC, Crimmins DL, Landt Y, Ladenson JH. Isolation and characterization of a thermally stable recombinant anti-caffeine heavy-chain antibody fragment. Anal Chem. 2006;78(13):4501–4508. | ||
Turner KB, Alves NJ, Medintz IL, Walper SA. Improving the targeting of therapeutics with single-domain antibodies. Expert Opin Drug Del. 2016;13(4):561–570. | ||
Kijanka M, Dorresteijn B, Oliveira S, Henegouwen P. Nanobody-based cancer therapy of solid tumors. Nanomedicine (Lond). 2015;10(1):161–174. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.