")
Back to Journals » Infection and Drug Resistance » Volume 12
Molecular characterization of para-aminosalicylic acid resistant Mycobacterium tuberculosis clinical isolates in southwestern China
Authors Luo M , Li K, Zhang H, Yan X, Gu J, Zhang Z, Chen Y, Li J, Wang J, Chen Y
Received 4 March 2019
Accepted for publication 22 May 2019
Published 24 July 2019 Volume 2019:12 Pages 2269—2275
DOI https://doi.org/10.2147/IDR.S207259
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Ming Luo,1 Kun Li,1 Huizheng Zhang,1 Xiaofeng Yan,2 Jing Gu,3 Zhen Zhang,4 Yu Chen,5 Jungang Li,1 Jing Wang,1 Yaokai Chen1
1Central Laboratory, Chongqing Public Health Medical Center, Chongqing 400036, People’s Republic of China; 2Department of Tuberculosis, Chongqing Public Health Medical Center, Chongqing 400036, People’s Republic of China; 3Key Laboratory of Special Pathogens and Biosafety, Wuhan Institute of Virology, Chinese Academy of Sciences, Wuhan 430071, People’s Republic of China; 4Department of Pharmacy, Chongqing Public Health Medical Center, Chongqing 400036, People’s Republic of China; 5Department of Medical Records, Chongqing Public Health Medical Center, Chongqing 400036, People’s Republic of China
Purpose: The aim of this study is to investigate para-aminosalicylic acid (PAS) resistance-related gene mutations in clinical Mycobacterium tuberculosis (MTB) isolates and analyze the associated risk factors in southwestern China.
Patients and methods: Total 122 PAS-resistant and 55 PAS-susceptible clinical isolates were obtained from Chongqing Public Health Medical Center between April 2014 and January 2018. Drug susceptibility test was performed, and the PAS resistance-related genes were sequenced.
Results: PAS-resistant strains were more likely to resist streptomycin (OR: 9.5, 95% CI: 3.87-23.3; P<0.01), isoniazid (OR: 5.98, 95% CI: 2.14–16.76; P<0.01), rifampin (OR: 5.01, 95% CI: 2.11–11.88; P<0.01), ethambutol (OR: 2.79, 95% CI: 1.44–5.4; P<0.01), levofloxacin (OR: 2.56, 95% CI: 1.33–4.93; P<0.01), and amikacin (OR: 4.29, 95% CI: 1.70–10.83; P<0.01). The sequencing results showed that 112 (91.8%) PAS-resistant strains harbored 30 different mutations in folC, thyA, and ribD. Mutations in folC were the most commonly observed in PAS-resistant isolates (54.5%, 61/112), followed by mutations in thyA and ribD. Residues I43 in folC, R235 in thyA, and −11G in upstream of ribD were hotspots for mutation sites.
Conclusion: PAS drug resistance in MTB in southwestern China is mainly caused by mutations in folC, thyA, and ribD, among which folC was the most frequent mutation. Some mutation hotspots exist in the three genes, which accounts for about 80% of total mutations. These results highlight the possibility of developing molecular diagnostic methods for PAS-resistant tuberculosis in the future.
Keywords: Mycobacterium tuberculosis, para-aminosalicylic acid, resistance, molecular characteristic, southwestern China
Introduction
Tuberculosis (TB) is caused by Mycobacterium tuberculosis (MTB) and is a major public health problem globally. The emergence and transmission of multidrug-resistant tuberculosis (MDR-TB), which is resistant to both rifampin and isoniazid, have aggravated this problem.1 WHO estimated that there were 10.0 million new TB cases and 458,000 new MDR-TB cases in 2017.2 China is one of the countries with a high burden of both TB and MDR-TB in the world. In 2017, there were 778,000 new TB cases in China, of which MDR-TB cases were 58,000.2
Para-aminosalicylic acid (PAS) is the second drug used for TB treatment after streptomycin and now is used as a second-line anti-TB drug of MDR-TB.3–5 Although PAS has been used for more than 70 years for TB therapy, its mechanism of action is not fully understood. In recent years, studies have revealed that PAS is a prodrug that targets the folate synthesis in MTB.6–8 PAS acts as an alternative substrate for dihydropteroate synthase (DHPS/FolP1). The product hydroxy dihydropteroate (H2PtePAS), an analog of dihydropteroate (H2Pte), can be glutaminated by dihydrofolate synthase (DHFS/FolC) to form hydroxy dihydrofolate (H2PtePAS-Glu), which is an analog of dihydrofolate (H2Pte-Glu).6 H2PtePAS-Glu inhibits dihydrofolate reductase (DfrA) and arrests the growth of M. tuberculosis.7 Consistent with this model, mutations causing the overexpression of RibD, an alternate enzyme with dihydrofolate reductase activity, confer resistance to PAS.7 Mutations in dihydrofolate synthase (FolC) trigger resistance to this drug.7,9 In addition, loss-of-function mutations in thymidylate synthase ThyA trigger resistance to PAS through an undefined pathway.8,10
Chongqing is a municipal city located in southwestern China with 32 million residents and is a high-incidence area for both TB and MDR-TB in China.11 Little is known about the molecular characteristics of PAS resistance in M. tuberculosis isolates in this region. In this study, we analyzed PAS resistance-related mutations including ribD, folC, and thyA in clinical isolates of M. tuberculosis in Chongqing, as well as the risk factors related to PAS resistance of M. tuberculosis.
Materials and methods
Bacteria strains
122 PAS-resistant and 55 PAS-susceptible clinical isolates were obtained from Chongqing Public Health Medical Center, a specialist hospital for TB and other infectious diseases in Chongqing, between April 2014 and January 2018. Patient’s information was acquired from their medical records. For each patient, two sputum samples were collected for mycobacterium culture using Löwenstein–Jensen (L-J) medium and the BD Bactec MGIT 960. Positive cultures were examined by Zeihl–Nelson stain, and species identification was performed with a commercial MPB64 monoclonal antibody assay (Genesis, Hangzhou, China).12 Positive cultures were recultured with L-J for further drug susceptibility testing (DST) and DNA extraction.
Drug susceptibility test
Drug susceptibility test was performed according to previous literature.13 Concentrations of anti-TB drugs in L-J medium were as follows: streptomycin (STR), 4 μg/mL; rifampin (RIF), 40 μg/mL; isoniazid (INH), 0.2 μg/mL; ethambutol (EMB), 2 μg/mL; levofloxacin (LVX), 2 μg/mL; amikacin (AMK), 30 μg/mL; capromycin (CAP), 40 μg/mL; prothionamide (PTO), 40 μg/mL; and PAS, 1 μg/mL.
PCR amplification and sequencing
Genomic DNA was extracted from freshly cultured bacteria using Hipure mycobacterial DNA kit (Magen, Guangzhou, China) according to the manufacturer’s instruction and used as the template for gene amplification. Each PCR mixture was prepared in a volume of 50 μL containing 5 μL PCR buffer, 2 mM MgSO4, 200 μM of each deoxynucleoside triphosphate (dNTP), 2.5 μL of DMSO, 0.2 μM of each primer, 1 μL genomic DNA, and 1 μL KOD-plus DNA polymerase (TOYOBO, Osaka, Japan). The PCR program for amplification was initiated at 98°C for 10 mins followed by 5 cycles of 98°C for 1 min, 57°C for 1 min, and 72°C for 1 min. After that, there were 30 cycles of denaturation at 94°C for 30 s, annealing at different temperatures for 30 s for each gene, extension at 72°C for 30 s, and a final extension at 72°C for 5 mins. The annealing temperatures of the 30 cycles for each gene were as follows: 57°C was for 16 S DNA; 63.2°C was for thyA; 64.3°C was for folC; and 66.5°C was for ribD. 16S DNA was used to eliminate nontuberculosis mycobacteria (NTM) from the isolates. All amplicons were sent to Tsingke Company (Beiing, China) for sequencing. The sequence data were aligned with thyA, folC, and ribD of H37Rv (NC000962.3) by ClustalW2.14 The primers used to amplify thyA, folC, ribD, and 16S DNA are listed in Table 1.
 |
Table 1 The primer pairs used in this study |
Statistical analysis
The Pearson Chi-square test was used to evaluate the associations among multiple categorical variables between PAS-susceptible and PAS-resistant groups. All statistical analysis was performed with SPSS 17.0 (IBM Corporation, Armonk, NY, USA). The results were summarized with ORs with 95% CIs. Differences were considered as statistically significant when the P-value was <0.05.
Ethics approval
Our study was conducted in accordance with the Declaration of Helsinki. The institutional review board of Chongqing Public Health Medical Center approved this study and waived the requirement for written informed consent. The institutional review board waived the need for informed consent because all patients’ data were analyzed in anonymity and no additional informed consent was required.
Results
Demographic characteristics
Among 177 clinical isolates included in this study, 116 (65.5%) were from male patients and 61 (34.5%) were from female patients. The mean age of patients was 40.2 years (range 13–75 years). There were 143 MDR-TB strains including 47 extensively drug-resistant tuberculosis (XDR-TB) strains (defined as MDR-TB strains resistant to at least one of the quinolone and one of the second-line anti-TB injectable drugs).15 A total of 30 strains were from new cases and 147 strains were from retreated cases. As shown in Table 2, of the 177 isolates, 147 (83.1%) were resistant to STR, 158 (89.3%) to INH, 150 (84.7%) to RIF, 95 (53.7%) to EMB, 111 (62.7%) to LVX, 48 (27.1%) to AMK, 49 (27.7%) to CAP, and 87 (49.2%) to PTO.
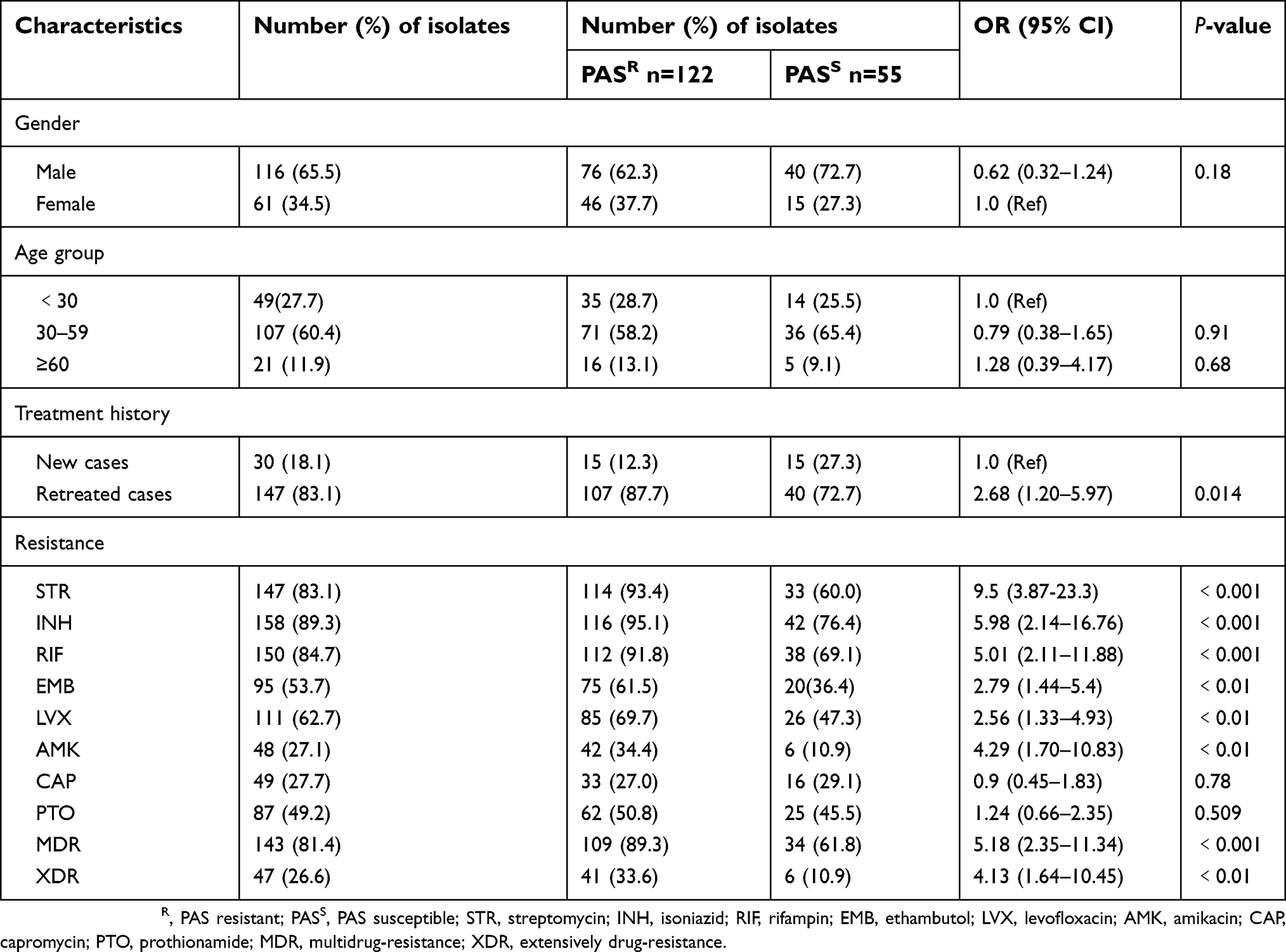 |
Table 2 Risk factor associated with PAS resistance in Chongqing, China |
Factors associated with PAS resistance
There was no significant difference in the distribution of the PAS-resistant isolates according to gender and age (P﹥0.05). However, the treatment history was clearly associated with the PAS resistance as the proportion of retreated patients in the PAS-resistant group was significantly higher (OR 95% CI: 2.68 [1.20–5.9], P<0.05). Further analysis of other drug-susceptibility data showed that PAS resistance was more likely to be related to the resistance of STR (OR: 9.5, 95% CI: 3.87-23.3; P<0.01), INH (OR: 5.98, 95% CI: 2.14–16.76; P<0.01), RIF (OR: 5.01, 95% CI: 2.11–11.88; P<0.01), EMB (OR: 2.79, 95% CI: 1.44–5.4; P<0.01), LVX (OR: 2.56, 95% CI: 1.33–4.93; P<0.01), and AMK (OR: 4.29, 95% CI: 1.70–10.83; P<0.01), but not correlated with CAP and PTO (P﹥0.05). In addition, the percentages of MDR (OR: 5.18, 95% CI: 2.35–11.34; P<0.01) and XDR (OR: 4.13, 95% CI: 1.64–10.45; P<0.01) in PAS-resistant group were significantly higher than those of PAS-susceptible group.
Mutations in three PAS resistance-related genes
Three genes (folC, thyA, and ribD) from the 177 MTB strains were sequenced. Among the 122 PAS-resistant isolates, 112 (91.8%) strains harbored 30 different mutation types in the three genes and 10 strains had no mutation in these three genes. No mutation was found in those genes in PAS-susceptible strains. As shown in Table 3, 59 strains harbored mutations in folC; 39 strains had mutations in thyA and 11 strains in ribD; 2 strains had mutations in both folC and thyA; 1 strain had mutations in both thyA and ribD. Mutations in folC were the most commonly found in PAS-resistant isolates (54.5%, 61/112) and 11 mutation types were found at 7 condons in folC. The most common mutation sites are residues 40, 43, 49, and 150. We found 18 mutation types at 16 different sites throughout the entire thyA gene and residues, 147 and 235, which were the most frequent mutation sites. Three types of mutation were identified in ribD including a synonymous mutation, and the dominant mutation is a G/A transition in the 11th nucleotide upstream of the start codon. In general, the most frequent mutations in PAS-resistant strains were I43T in folC (35.7, 40/112), followed by R235P in thyA (14.3%, 16/112) and −11G→A of ribD (8.0%, 9/112). The detailed information of all mutations is listed in Table S1.
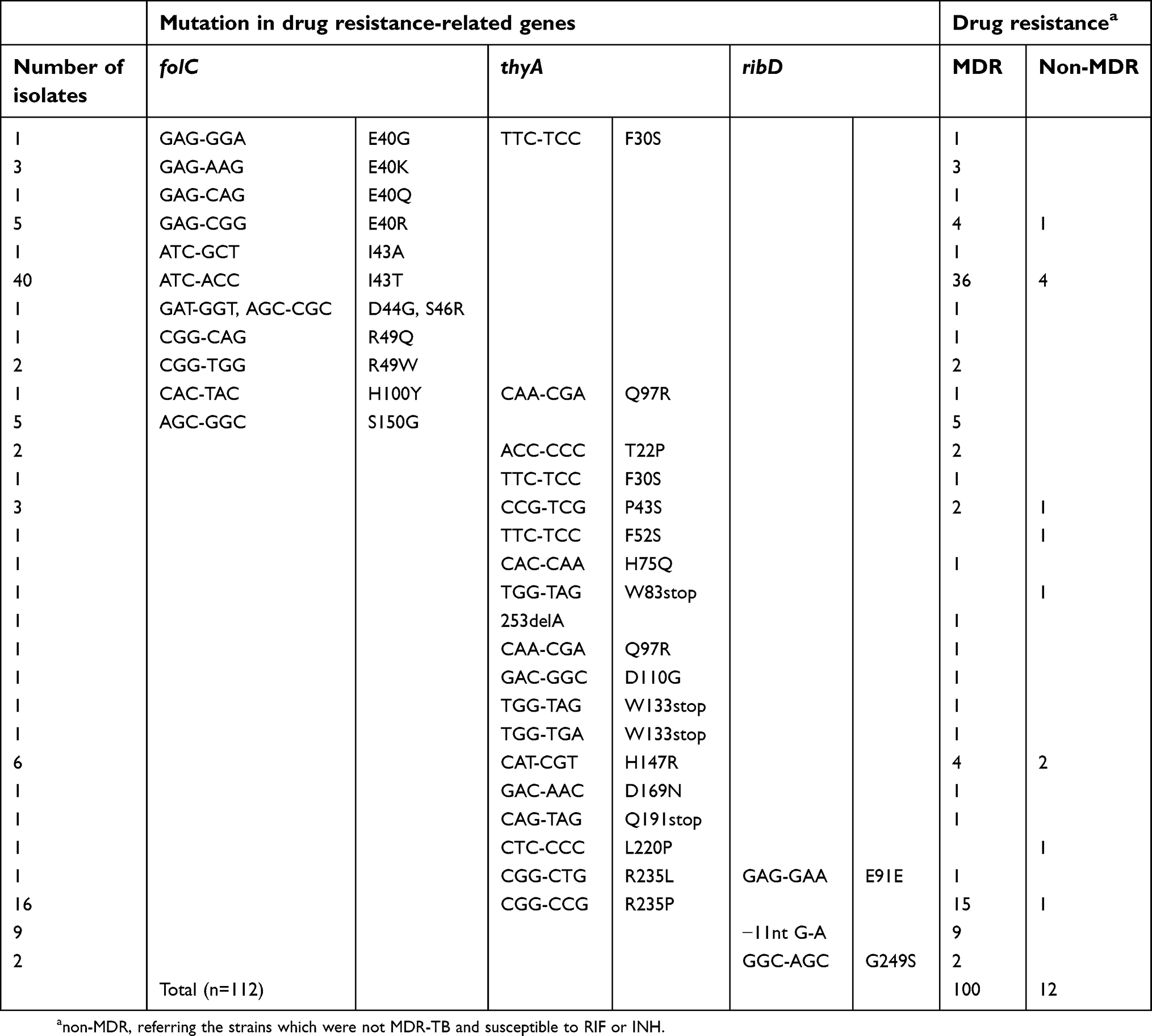 |
Table 3 Features of PAS-resistant isolates in Chongqing, China |
Discussion
Although PAS has been used to treat TB for decades, its mechanisms of resistance are still not fully understood. Previously, mutations of three genes, folC, thyA, and ribD, were found to be related to PAS resistance in MTB clinical isolates.7–10 However, there are few studies reporting on the distribution of mutations in these three genes in clinical PAS-resistant MTB isolates. Furthermore, the molecular characteristics of PAS-resistant MTB in southwestern China are still unclear.
In this study, we found that PAS resistance was significantly associated with resistance to most of the anti-TB drugs (STR, INH, RIF, EMB, LVX, and AMK), as well as the treatment history and clinical types of TB (MDR-TB and XDR-TB). These results are consistent with previous researches on PTO and pyrazinamide (PZA).16,17 Previous studies have shown that the exposure to antimicrobial agents such as RIF, fluoroquinolone (FQ), and aminoglycosides induces the production of oxygen radicals, which would then cause high-frequency mutagenesis in M. tuberculosis resulting in cross-resistance between PZA and other drugs.17 Considering that retreated, MDR and XDR cases required longer durations of anti-TB therapy than new and non-MDR cases, we speculated the mechanism for this cross-resistance between PAS and other anti-TB agents should be the same as that of PZA cross-resistance.
Another finding of our study is that we observed a very high proportion (91.8%, 112/122) of mutations in PAS-resistant MTB isolates, and some mutation hotspots did exist in PAS-resistant TB isolates from Chongqing. So far, there has been limited data about the genotypic characteristics of PAS-resistant MTB. Recently, Zhang X et al examined gene mutations in PAS-resistant MTB from northern China.18 Compared to the results from northern China (61.1%, 127/208), the mutation rate in PAS-resistant MTB isolates from Chongqing is much higher. In our study, mutations in folC can be found in half of the PAS-resistant isolates (50.0%, 61/122), which is much higher than that from northern China (29.3%, 61/208). We also found that mutation hotspots did exist, such as residues E40 (10 isolates), I43 (41 isolates), and S150 (5 isolates) in folC and H47 (6 isolates) and R235 (17 isolates) in thyA, −11G (9 isolates) upstream of ribD. Mutations in these loci accounted for 79.1% (88/112) of total mutations. Although I43T is the dominant mutation in folC in PAS-resistant MTB from both Chongqing and northern China, it seems to be more common in isolates from Chongqing (64.5%, 40/62 from Chongqing; vs 23.0%, 14/61 from northern China). Previously, E153A was found to be the most frequently identified mutation type in folC in PAS-resistant isolates from Hongkong, which is also common in isolates from northern China. However, in this study, we did not find any E150 mutation in PAS-resistant isolates from Chongqing.18,19
Three new mutations were found in folC, including two single mutations (E40R and H100Y) and a double mutation (E44G: S46R). To our best knowledge, this is the first report showing that double mutations exist in folC of PAS-resistant TB. Most mutations in folC such as E40, I43, R49, S150, and E153 are located in the substrate binding pocket, which leads to reduced efficiency of H2PtePAS glutamination.9,19,20 Since E44 and S46 were adjacent to I43 and R49, H100 was located in the Ω-loop just adjacent to the Ser-cis-Pro motif in the active site of FolC.20 These results suggest that the mutations in these residues may also affect the glutamination process.
thyA was the first gene reported to be associated with PAS resistance.8 Previous reports showed that about a quarter to one-third of the PAS-resistant strains contain thyA mutations, and our results (34.4%, 42/122) are consistent with those findings.18,21 We identified 18 mutation types at 16 different sites throughout the entire thyA. Similar to the findings of northern China, most of the mutations are located in the active or dimeric domains.18 Nevertheless, the most common mutation in thyA in our study was R235P, whereas the most frequent mutation in northern China, H75N, was found only in one strain in our study.18 R235 is located on the outer surface of ThyA (PDB code 4FQS), and it is unreasonable to explain the impact of this mutation on thymidylate synthase activity by proximity of position. Therefore, how the mutation at this site affects thymidylate synthase activity needs further investigation. We observed that −11G→A transition was the most common mutation in ribD, which is also consistent with the previous finding in northern China.18 This mutation leads to the overexpression of ribD and finally causes PAS resistance.7 Interestingly, a G249S mutation was identified in ribD. Notwithstanding, further studies are warranted in order to verify the relationship between this mutation and PAS-resistance.
As far as we know, this is the first investigation of phenotypic and molecular characteristics of PAS-resistance in southwestern China. It is well understood that the accuracy of molecular diagnosis of drug-resistant TB depends on understanding the mechanisms of drug resistance and the prevalence of drug-resistant mutations in TB. Thus, our study will expand our understanding of the mechanisms of PAS-resistance, and benefit both the molecular diagnosis of PAS resistant MTB and clinical treatment of PAS-resistant TB patients in south-western China. Nevertheless, there were some obvious limitations in our research. For example, the sample size was small, which limits the representativeness of our results. What is more, all isolates in our study were from a specialist hospital for TB, which may lead to deviations in high TB drug resistance. Therefore, multicenter and larger sample studies are still needed.
Conclusion
PAS drug resistance in southwestern China is mainly caused by mutations in folC, thyA, and ribD, among which folC was the most frequent mutation. Some mutation hotspots exist in the three genes, which accounts for about 80% of total mutations. Our results highlight the possibility of developing molecular diagnostic methods for PAS-resistant TB in the future.
Acknowledgments
This work is funded by the National Science and Technology Major Project of China for the 13th five-year plan period (2018ZX10302104) and the Open Research Fund Program of Wuhan National Bio-Safety Level 4 Lab of Chinese Academy of Science (2017SPCAS004).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Zhao Y, Xu S, Wang L, et al. National survey of drug-resistant tuberculosis in China. N Engl J Med. 2012;366(23):2161–2170. doi:10.1056/NEJMoa1108789
2. World Health Organization (WHO). Global Tuberculosis Report 2018. Geneva: WHO; 2018.
3. Lehmann J. Para-aminosalicylic acid in the treatment of tuberculosis. Lancet. 1946;247(6384):15–16. doi:10.1016/S0140-6736(46)91185-3
4. Lehmann J. Twenty years afterward historical notes on the discovery of the antituberculosis effect of para-aminosalicylic acid (PAS) and the first clinical trials. Am Rev Respir Dis. 1964;90:953–956. doi:10.1164/arrd.1964.90.6.953
5. Dookie N, Rambaran S, Padayatchi N, Mahomed S, Naidoo K. Evolution of drug resistance in Mycobacterium tuberculosis: a review on the molecular determinants of resistance and implications for personalized care. J Antimicrob Chemother. 2018;73(5):1138–1151. doi:10.1093/jac/dkx506
6. Chakraborty S, Gruber T, Barry CE
7. Zheng J, Rubin EJ, Bifani P, et al. para-aminosalicylic acid is a prodrug targeting dihydrofolate reductase in Mycobacterium tuberculosis. J Biol Chem. 2013;288(32):23447–23456. doi:10.1074/jbc.M113.475798
8. Rengarajan J, Sassetti CM, Naroditskaya V, Sloutsky A, Bloom BR, Rubin EJ. The folate pathway is a target for resistance to the drug para-aminosalicylic acid (PAS) in mycobacteria. Mol Microbiol. 2004;53(1):275–282. doi:10.1111/j.1365-2958.2004.04120.x
9. Zhao F, Wang XD, Erber LN, et al. Binding pocket alterations in dihydrofolate synthase confer resistance to para-aminosalicylic acid in clinical isolates of Mycobacterium tuberculosis. Antimicrob Agents Chemother. 2014;58(3):1479–1487. doi:10.1128/AAC.01775-13
10. Fivian-Hughes AS, Houghton J, Davis EO. Mycobacterium tuberculosis thymidylate synthase gene thyX is essential and potentially bifunctional, while thyA deletion confers resistance to p-aminosalicylic acid. Microbiology. 2012;158(Pt 2):308–318. doi:10.1099/mic.0.053983-0
11. Zhang D, An J, Wang J, et al. Molecular typing and drug susceptibility of Mycobacterium tuberculosis isolates from Chongqing municipality, China. Infect Genet Evol. 2013;13:310–316. doi:10.1016/j.meegid.2012.10.008
12. Pang Y, Dong H, Tan Y, et al. Rapid diagnosis of MDR and XDR tuberculosis with the MeltPro TB assay in China. Sci Rep. 2016;6:25330. doi:10.1038/srep25330
13. World Health Organization. WHO Guidelines Approved by the Guidelines Review Committee. Policy Guidance on Drug-Susceptibility Testing (DST) of Second-Line Antituberculosis Drugs. Geneva: World Health Organization; 2008.
14. Larkin MA, Blackshields G, Brown NP, et al. Clustal W and clustal X version 2.0. Bioinformatics. 2007;23(21):2947–2948. doi:10.1093/bioinformatics/btm404
15. Pang Y, Zhou Y, Zhao B, et al. Spoligotyping and drug resistance analysis of Mycobacterium tuberculosis strains from national survey in China. PLoS One. 2012;7(3):e32976. doi:10.1371/journal.pone.0032976
16. Tan Y, Su B, Zheng H, Song Y, Wang Y, Pang Y. Molecular characterization of prothionamide-resistant Mycobacterium tuberculosis isolates in Southern China. Front Microbiol. 2017;8:2358. doi:10.3389/fmicb.2017.02358
17. Pang Y, Zhu D, Zheng H, et al. Prevalence and molecular characterization of pyrazinamide resistance among multidrug-resistant Mycobacterium tuberculosis isolates from Southern China. BMC Infect Dis. 2017;17(1):711. doi:10.1186/s12879-017-2761-6
18. Zhang X, Liu L, Zhang Y, Dai G, Huang H, Jin Q. Genetic determinants involved in p-aminosalicylic acid resistance in clinical isolates from tuberculosis patients in northern China from 2006 to 2012. Antimicrob Agents Chemother. 2015;59(2):1320–1324. doi:10.1128/AAC.03695-14
19. Cheng VW, Leung KS, Kwok JS, et al. Phylogenetic and structural significance of dihydrofolate synthase (folc) mutations in drug-resistant Mycobacterium tuberculosis. Microb Drug Resist. 2016;22(7):545–551. doi:10.1089/mdr.2015.0193
20. Mathieu M, Debousker G, Vincent S, Viviani F, Bamas-Jacques N, Mikol V. Escherichia coli FolC structure reveals an unexpected dihydrofolate binding site providing an attractive target for anti-microbial therapy. J Biol Chem. 2005;280(19):18916–18922. doi:10.1074/jbc.M413799200
21. Mathys V, Wintjens R, Lefevre P, et al. Molecular genetics of para-aminosalicylic acid resistance in clinical isolates and spontaneous mutants of Mycobacterium tuberculosis. Antimicrob Agents Chemother. 2009;53(5):2100–2109. doi:10.1128/AAC.01197-08
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.