")
Back to Journals » Infection and Drug Resistance » Volume 12
Molecular and phenotypical characterization of two cases of antibiotic-driven ceftazidime-avibactam resistance in blaKPC-3-harboring Klebsiella pneumoniae
Authors Venditti C, Nisii C , D’Arezzo S , Vulcano A, Capone A , Antonini M, Ippolito G , Di Caro A
Received 8 March 2019
Accepted for publication 13 April 2019
Published 3 July 2019 Volume 2019:12 Pages 1935—1940
DOI https://doi.org/10.2147/IDR.S207993
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sahil Khanna
Carolina Venditti,1 Carla Nisii,1 Silvia D’Arezzo,1 Antonella Vulcano,1 Alessandro Capone,2 Mario Antonini,2 Giuseppe Ippolito,3 Antonino Di Caro1
1Laboratory of Microbiology, National Institute for Infectious Diseases "L. Spallanzani", Rome, Italy; 2Clinical Department, National Institute for Infectious Diseases "L.Spallanzani", Rome, Italy; 3Scientific Direction, National Institute for Infectious Diseases "L.Spallanzani", Rome, Italy
Background: For years, Klebsiella pneumoniae carbapenemase (KPC)-producing K. pneumoniae have represented a serious health problem in hospitals worldwide. Since its approval in 2015, ceftazidime-avibactam (CAZ-AVI) had been successfully used for treating complicated KPC-K. pneumoniae infections, until increasing reports of resistance began to emerge.
Methods: Phenotypic tests and molecular analysis were performed in four multidrug-resistant K. pneumoniae isolates, collected from two patients following treatment with CAZ-AVI.
Results: In this study, we report two cases of emergence of CAZ-AVI resistance in KPC-3-producing K. pneumoniae isolates, collected from two patients following treatment with CAZ-AVI. Molecular analysis highlighted the D179Y mutation in the blaKPC-3 gene, whose role in the loss of hydrolytic activity (resulting in decreased carpabenem minimum inhibitory concentrations and negative phenotypic tests) of the enzyme has already been shown.
Conclusion: Most surveillance schemes aimed at detecting carbapenem-resistant Enterobacteriaceae (CRE) rely on confirmatory phenotypic tests for detecting carbapenemase production. As reports of these treatment-induced, altered CRE phenotypes are increasing, the initial susceptibility testing should be followed by a combination of phenotypic and molecular methods, to make sure that no potential carbapenemase-producing bacteria are missed.
Keywords: Klebsiella pneumoniae, carbapenem-resistance, ceftazidime-avibactam
Introduction
Carbapenem-resistant Enterobacteriaceae (CRE) are among the greatest challenges to global health because of their limited treatment options and increased risk of adverse outcomes.1,2 In Italy, the dissemination of CRE has largely been due to the plasmid-mediated Klebsiella pneumoniae carbapenemase (KPC), which represents the most common serine-carbapenemase in K. pneumoniae.3 Few drugs are active against these strains, and a combination therapy is often necessary.
Avibactam (AVI) is a non-beta-lactam beta-lactamase inhibitor with broad-spectrum activity against serine-carbapenemases. In association with the 3rd generation cephalosporin ceftazidime (CAZ), AVI restores the activity of CAZ by inhibiting the Ambler class A carbapenemases (KPC included), class C, and some class D beta-lactamases, eg, the OXA-48 group.4 Since its approval in 2015 by the Food and Drug Administration, CAZ-AVI has been successfully used in patients with limited treatment options for complicated intra-abdominal infections and hospital-acquired, ventilator-associated pneumonia caused by CRE and other multidrug Gram-negative bacteria.5,6 The greatest clinical benefit of CAZ-AVI is its activity against KPC-producing CRE, which have become increasingly prevalent infectious organisms worldwide.7 After initial promising results, however, several studies began reporting the emergence of CAZ-AVI resistance either in the absence of prior treatment, or after short periods of exposure to CAZ-AVI. Mutations in the blaKPC gene, decreased permeability caused by mutations in outer member proteins and differences in susceptibilities of KPC subtypes are all mechanism of CAZ-AVI resistance observed in CRE, especially KPC-producing K. pneumoniae.8–11 The mechanism most often associated with the emergence of CAZ-AVI resistance after treatment has been linked to single amino acid substitutions in the KPC Ω-loop.12,13
The aim of this study was to investigate the emergence of resistance after a short course of CAZ-AVI by characterizing four KPC-producing K. pneumoniae isolates obtained from two patients, before and after treatment.
Patients
The two patients included in the study were treated in the intensive care unit (ICU) at the “L. Spallanzani” Hospital in Rome, Italy, and the four strains analyzed were obtained from routine broncho-aspirate samples (BAS), before and after treatment with CAZ-AVI.
Patient-1, a 27-year-old female with HBV-related fulminant hepatic failure, underwent liver transplantation 5 days after admission. The patient required prolonged mechanical ventilation for a large-for-size graft syndrome treated with a bogotà-bag. Piperacillin-tazobactam had been administered since admission and was continued until 4 days after surgery. On post-operative day 4, because of the emergence of fever, meropenem (1 g every 8 hrs), linezolid and anidulafungin were added; after 12 days, meropenem was replaced by tigecycline. On day 22, carbapenem and colistin-resistant K. pneumoniae were grown from a BAS (strain Kpn1-T0) and from an intra-abdominal abscess after a further 9 days. Antimicrobial treatment with CAZ-AVI and fosfomycin (2.5 g every 8 hrs and 4 g every 4 hrs, respectively) was therefore started, and was continued for 30 days with a good clinical response. Ten days after the interruption of treatment, a CAZ-AVI-resistant K. pneumoniae isolate, with an minimum inhibitory concentration (MIC) of >256 µg/mL (strain Kpn1-T30) and lower carbapenem MICs (Table 1), was isolated from a surveillance BAS; no further antimicrobial treatment was initiated. The patient died 82 days after admission due to graft dysfunction and multiple organ failure.
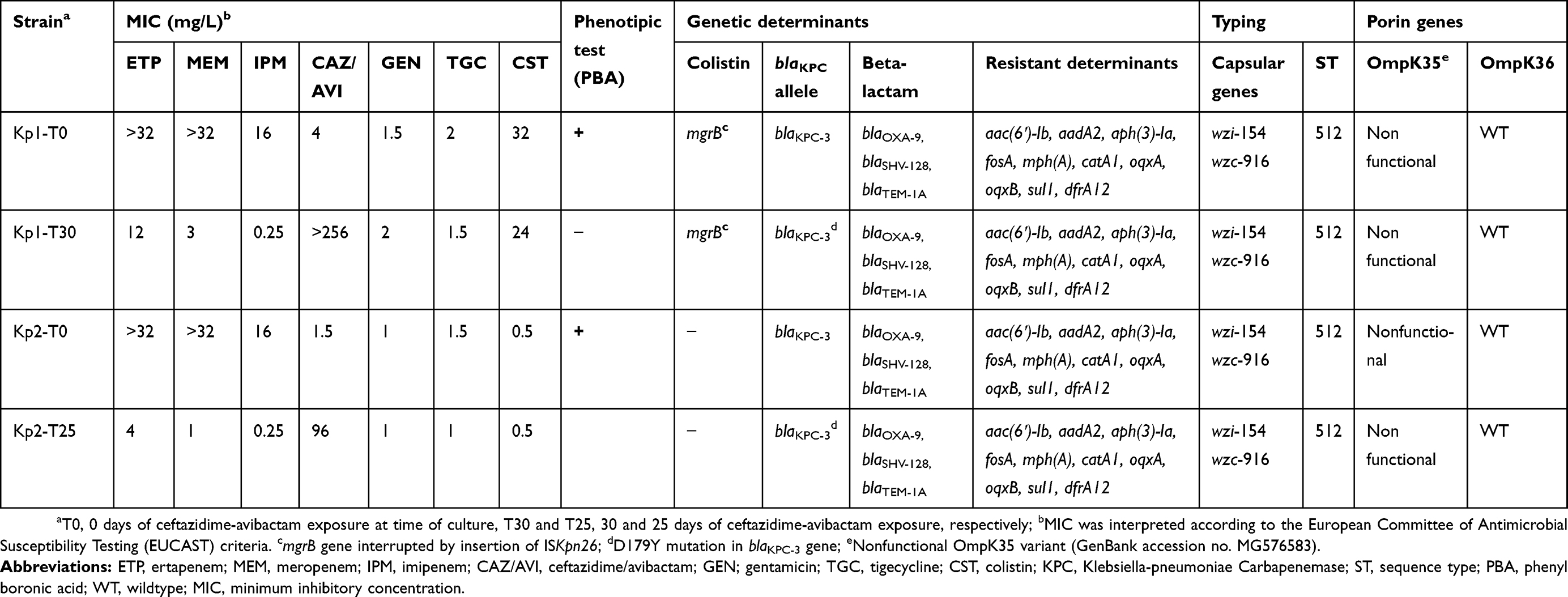 |
Table 1 Phenotypic and molecular analysis of KPC-K. pneumoniae clinical isolates described in this study |
Patient-2, a 53-year-old man with HIV-AIDS and kaposi sarcoma, was transferred to the ICU with a septic shock and respiratory failure following surgery. The patient had undergone a week of empiric antibiotic therapy with meropenem, vancomycin and voriconazole. A methicillin-resistant Staphylococcus aureus was grown from blood cultures, and the antibiotic therapy was switched to linezolid and daptomycin. A carbapenem-resistant K. pneumoniae was isolated from the BAS after 17 days (strain Kpn2-T0). At this point, the patient was treated with colistin plus tigecycline. Ten days later, when also the patient’s haemocultures became positive, CAZ-AVI (2.5 g every 8 hrs) was added in substitution of colistin, and was continued for 25 days. A CAZ-AVI resistant (MIC 96 µg/mL) and meropenem-susceptible K. pneumoniae (strain Kpn2-T25) was isolated from a surveillance BAS 3 days after the treatment was withdrawn. The patient did not develop any further acute infectious complications and was later discharged.
Phenotypic and molecular characterization of isolates
Species identification and antimicrobial susceptibility were obtained by MALDI-TOF MS (bioMérieux, Marcy l’Étoile, France) and Phoenix system (Becton Dickinson Diagnostics, CA, USA). As per EUCAST recommendations, MICs were confirmed by broth microdilution for colistin and CAZ-AVI (Sensititre, ThermoFisher Scientific, Waltham, MA, USA), while the E-test method (Liofilchem, Roseto degli Abruzzi, Italy) was used for tigecycline.14 Confirmatory phenotypic inhibition tests for detection of carbapenemase activity were performed on all isolates, using phenyl boronic acid (PBA) and EDTA (Liofilchem).
Multiplex PCRs were performed to identify the most frequently detected carbapenemase genes belonging to class A (blaKPC), class B (blaVIM and blaNDM) and class D (blaOXA-48-like) (RealCycler KPND and OXVI, Progenie Molecular, Spain). Identification of mutations in the blaKPC genes was performed using Sanger sequencing, mapping the reads of the CAZ-AVI-resistant sequences by Basic Local Alignment Search Tool (BLAST) analysis against the blaKPC-3 reference sequence (GenBank accession no. AF395881.1). Colistin resistance mechanisms were analyzed by standard PCR of the mgrB gene chromosome,15 as well as porin alterations (OmpK35 and OmpK36), as previously described.16
Sequence types (ST) were determined by Multi Locus Sequence Typing (MLST) of the traditional seven constitutive genes, as previously described17 while capsular types were identified by PCR and sequence analysis of the conserved wzi and wzc genes, present in the capsule polysaccharide locus (http://bigsdb.web.pasteur.fr/klebsiella/).
Genetic relationships were further investigated by the validated WGS-based core genome MLST (cgMLST) scheme v1.0, using the Ridom SeqSphere + software (Ridom GmbH, Münster, Germany) with default settings. Whole genome sequencing was performed using the Ion Torrent GSS5 platform (Life Technologies, Carlsbad, CA, USA) by constructing single-end libraries with average lengths of 200 bp according to manufacturer’s instructions. All raw reads generated were submitted to the Sequence Read Archive (SRA) under the BioProject accession number PRJNA510900.
Compared to the reference strain (GenBank accession no. NC_012731.1), 2,358 target genes were used to characterize the gene-by-gene allelic profile of the K. pneumoniae isolates. Antimicrobial resistance genes and plasmid replicons were extracted from the WGS data and identified by in silico analysis using the ResFinder v3.0 web server (http://www.genomicepidemiology.org). The minimum percentage of sequence identity was set at 100%, with an alignment length of >98%.
Results
For both patients, testing of susceptibility to CAZ-AVI and carbapenems showed two distinct susceptibility phenotypes (Table 1): at baseline (patient 1, strain Kpn1-T0 and patient 2, strain Kpn2-T0) and after 30 days (patient 1, strain Kpn1-T30) and 25 days (patient 2, strain Kpn2-T25) of CAZ-AVI treatment. Gene content analysis demonstrated that all four belonged to ST512 with no allele differences within the 2,358-gene targets detected using cgMLST analysis and with identical capsular polysaccharide and porin genes (Table 1).
Beta-lactamase gene variants and resistant determinants are also shown in Table 1. Antimicrobial susceptibility profiles showed that the isolates were resistant to: amoxicillin/clavulanic acid (MIC >32/2 mg/L), ampicillin, CAZ and cefepime (MIC >8 mg/L), cefotaxime and tobramycin (MIC >4 mg/L), ciprofloxacin (MIC >1 mg/L), amikacin (MIC >16 mg/L), trimethoprim/sulfamethoxazole (MIC >4/76 mg/L) and piperacillin/tazobactam (MIC >16/4 mg/L). All four strains were susceptible to gentamicin and tigecycline with varying MIC values (Table 1).
Both isolates of patient 1 showed resistance to colistin due to inactivation of the mgrB gene by insertion of ISKpn26 (an IS5-like element). The strains isolated from patient 2 were susceptible to colistin.
Plasmid content was the same in all strains. We observed the following plasmids of incompatibility groups (100% identity): FII(K), FIB(pQil), all known replicons of plasmids carrying the blaKPC-3 gene (pKpQil-IT, JN233705), FIB(K) and ColRNAI.
Following treatment, only CAZ-AVI-resistant K. pneumoniae isolates with decreased carbapenem MICs (strains Kpn1-T30 and Kpn2-T25 for patient 1 and patient 2, respectively) were cultured from both patients’ BAS. As shown in Table 1, the MICs of CAZ-AVI increased by at least six 2-fold serial dilutions, ie, from 4 to >256 mg/L and from 1.5 to 96 mg/L, for patients 1 and 2, respectively. Instead, the MICs of all three carbapenems were markedly decreased, for both patients’ isolates (Table 1). Phenotypic inhibition tests using PBA were positive for both “parent” strains, and negative for the isolates grown after CAZ-AVI treatment. Despite the decreased MIC values and the negative phenotypic inhibition tests, also the two “post-treatment” strains harboured the blaKPC gene, as detected by WGS data and confirmed by PCR (Table 1). BLAST analysis of the complete genome sequences revealed the presence of the blaKPC-31 variant (GenBank accession no. NG_055494.1) in the CAZ-AVI-resistant K. pneumoniae isolates. Analysis of the blaKPC gene showed the presence of a single nucleotide substitution (G to T), which causes a tyrosine/aspartic acid change at position 179 (D179Y). This mutation determines an alteration within the conserved blaKPC Ω loop of the blaKPC gene, which has been shown to have a central role in the hydrolytic stability of the enzyme.18
Discussion
In recent years, the increase in resistance seen in Gram-negative bacteria has led to strong advocacy for the research and development of new agents against infections caused by such organisms. CAZ-AVI has represented a promising therapeutic option to combat KPC-producing K. pneumoniae.5,6 However, the increased reporting of resistance, with or without prior exposure to the compound, is becoming a matter of concern.
It has been reported that the period of exposure to CAZ-AVI varies from 12 to 19 days and that CAZ-AVI-resistant K. pneumoniae have been often identified as respiratory tract colonizers or a cause of recurrent pneumonia.8–13 The mechanism most often associated with resistance emerged after a CAZ-AVI-based therapy has been linked to single amino acid substitutions in the blaKPC gene.13,19 Our results are in accordance with these data, as the mutation we observed in the two strains isolated from BAS samples of two patients after a course of CAZ-AVI therapy was the single D179Y substitution, which has been shown to determine an alteration within the conserved blaKPC Ω loop.16 Such mutation produces the effect of preventing the binding of AVI, causing the simultaneous reduction of carbapenemase activity, and the decrease of carbapenem MICs. This results in an apparent phenotypic change from carbapenem-resistant to susceptible (although with elevated MICs), of KPC-producing K. pneumoniae, as we observed in the two cases we described in this study.
The spread of CAZ-AVI-resistant CRE would bring serious consequences for hospital surveillance and infection control, so much so that the ECDC recently issued a document calling for actions.20 Many microbiology laboratories often rely on susceptibility testing followed by phenotypic tests for carbapenemase production in their active CRE surveillance schemes, while others employ a molecular approach by screening for the presence of the blaKPC gene by PCR. Only rarely are both methods used, because of higher costs and workload. It is important to remember that when employing phenotypic tests alone, which demonstrate the production and hydrolytic activity of the enzymes, without prior knowledge of CAZ-AVI exposure a KPC-K. pneumoniae strain carrying a mutated blaKPC gene (which would yield a negative result) could be mistaken for an extended-spectrum β-lactamase (ESBL) producer, as has been reported before.10 On the other hand, PCR detection of the presence of the blaKPC gene would not imply resistance to carbapenems, owing to a possible alteration of the gene.
Taken together, our results as well as all the recent literature data on CAZ-AVI resistance are a reminder that we need to use all diagnostic means available, ie, the combination of phenotypic and molecular methods, to ensure the best possible sensitivity in the search of resistance determinants. Furthermore, with the increasing use of CAZ-AVI in hospitals worldwide, it can be expected that resistance will continue to occur. In this scenario, the risk of a rapid spread of these new resistance determinants by horizontal gene transfer is high and hospital surveillance schemes need to be improved and strengthened: new strategies are needed to meet new challenges.
Ethical statement
No samples were collected specifically for this research; only anonymized clinical residual samples collected during routine hospital procedures were used for this study.
Acknowledgment
This work was supported by “Ricerca corrente” and “5X1.000” research funds from the Italian Ministry of Health.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Tumbarello M, Trecarichi EM, De Rosa FG, et al. Infections caused by KPC-producing Klebsiella pneumoniae: differences in therapy and mortality in a multicentre study. J Antimicrob Chemother. 2015;70(7):2133–2143. doi:10.1093/jac/dkv086
2. Martin A, Fahrbach K, Zhao Q, Lodise T. Association between carbapenem resistance and mortality among adult, hospitalized patients with serious infections due to Enterobacteriaceae: results of a systematic literature review and meta-analysis. Open Forum Infect Dis. 2018;5:ofy150. doi:10.1093/ofid/ofy150
3. Giani T, Antonelli A, Caltagirone M, et al. Evolving beta-lactamase epidemiology in Enterobacteriaceae from Italian nationwide surveillance, October 2013: KPC-carbapenemase spreading among outpatients. Euro Surveill. 2017;22(31):pii:30583.
4. Falagas ME, Lourida P, Poulikakos P, Rafailidis PI, Tansarli GS. Antibiotic treatment of infections due to carbapenem-resistant Enterobacteriaceae: systematic evaluation of the available evidence. Antimicrob Agents Chemother. 2014;58(2):654–663. doi:10.1128/AAC.01222-13
5. Temkin E, Torre-Cisneros J, Beovic B, et al. Ceftazidime-avibactam as salvage therapy for infections caused by carbapenem-resistant organisms. Antimicrob Agents Chemother. 2017;61(2):pii:e01964-e02016.
6. Tumbarello M, Trecarichi EM, Corona A, et al. Efficacy of ceftazidime-avibactam salvage therapy in patients with infections caused by KPC-producing Klebsiella pneumoniae. Clin Infect Dis. 2018;18;68(3):355–364
7. Grundmann H, Glasner C, Albiger B, et al. Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): a prospective, multinational study. Lancet Infect Dis. 2017;17(2):153–163. doi:10.1016/S1473-3099(16)30257-2
8. Humphries RM, Yang S, Hemarajata P, et al. First report of ceftazidime-avibactam resistance in a KPC-3-expressing Klebsiella pneumoniae isolate. Antimicrob Agents Chemother. 2015;59:6605–6607. doi:10.1128/AAC.01165-15
9. Shields RK, Potoski BA, Haidar G, et al. Clinical outcomes, drug toxicity, and emergence of ceftazidime-avibactam resistance among patients treated for carbapenem-resistant enterobacteriaceae infections. Clin Infect Dis. 2016;63(12):1615–1618. doi:10.1093/cid/ciw636
10. Haidar G, Clancy CJ, Shields RK, Hao B, Cheng S, Nguyen MH. Mutations in bla (KPC-3) that confer ceftazidime-avibactam resistance encode novel KPC-3 variants that function as extended-spectrum β-lactamases. Antimicrob Agents Chemother. 2017;61(5). doi:10.1128/AAC.02534-16
11. Shields RK, Chen L, Cheng S, et al. Emergence of ceftazidime-avibactam resistance due to plasmid-borne bla(KPC-3) mutations during treatment of carbapenem-resistant Klebsiella pneumoniae infections. Antimicrob Agents Chemother. 2017;61(3). doi:10.1128/AAC.02097-16.
12. Gaibani P, Campoli C, Lewis RE, et al. In vivo evolution of resistant subpopulations of KPC-producing Klebsiella pneumoniae during ceftazidime/avibactam treatment. J Antimicrob Chemother. 2018;73(6):1525–1529. doi:10.1093/jac/dky082
13. Hemarajata P, Humphries RM. Ceftazidime/avibactam resistance associated with L169P mutation in the omega loop of KPC-2. J Antimicrob Chemother. 2019;74:1241–1243. doi:10.1093/jac/dkz026
14. European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint tables for interpretation of MICs and zone diameters. Version 8.1. Örebro: EUCAST; 2018. Available from: http://www.eucast.org.
15. Cannatelli A, D’Andrea MM, Giani T, et al. In vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob Agents Chemother. 2013;57(11):5521–5526. doi:10.1128/AAC.01480-13
16. Kaczmarek FM, Dib-Hajj F, Shang W, Gootz TD. High-level carbapenem resistance in a Klebsiella pneumoniae clinical isolate is due to the combination of bla(ACT-1) beta-lactamase production, porin OmpK35/36 insertional inactivation, and down-regulation of the phosphate transport porin phoe. Antimicrob Agents Chemother. 2006;50(10):3396–3406. doi:10.1128/AAC.00285-06
17. Diancourt L, Passet V, Verhoef J, Grimont PA, Brisse S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J Clin Microbiol. 2005;43(8):4178–4182. doi:10.1128/JCM.43.8.4178-4182.2005
18. Winkler ML, Papp-Wallace KM, Bonomo RA. Activity of ceftazidime/avibactam against isogenic strains of Escherichia coli containing KPC and SHV beta-lactamases with single amino acid substitutions in the omegaloop. J Antimicrob Chemother. 2015;70:2279–2286. doi:10.1093/jac/dku445
19. Giddins MJ, Macesic N, Annavajhala MK, et al. Successive emergence of ceftazidime-avibactam resistance through distinct genomic adaptations in bla(KPC-2)-Harboring Klebsiella pneumoniae sequence type 307 isolates. Antimicrob Agents Chemother. 2018;62(3):pii: e02101-e02117.
20. European Centre for Disease Prevention and Control. Emergence of resistance to ceftazidime-avibactam in carbapenem-resistant Enterobacteriaceae – 12 June 2018. Stockholm; ECDC; 2018
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.