")
Back to Journals » Veterinary Medicine: Research and Reports » Volume 10
Modeling the pasture-associated severe equine asthma bronchoalveolar lavage fluid proteome identifies molecular events mediating neutrophilic airway inflammation
Authors Bright LA, Dittmar W, Nanduri B, McCarthy FM, Mujahid N, Costa LRR , Burgess SC , Swiderski CE
Received 13 November 2018
Accepted for publication 12 February 2019
Published 2 May 2019 Volume 2019:10 Pages 43—63
DOI https://doi.org/10.2147/VMRR.S194427
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Young Lyoo
Lauren A Bright,1 Wellesley Dittmar,1 Bindu Nanduri,2 Fiona M McCarthy,3 Nisma Mujahid,2 Lais RR Costa,2 Shane C Burgess,3 Cyprianna E Swiderski1
1Department of Clinical Sciences, College of Veterinary Medicine, Mississippi State University, Starkville, MS 39762, USA; 2Department of Basic Sciences, College of Veterinary Medicine, Mississippi State University, Starkville, MS 39762, USA; 3School of Animal Comparative and Biomedical Sciences, College of Agriculture and Life Sciences, University of Arizona, Tucson, AZ 85721, USA
Background: Pasture-associated severe equine asthma is a warm season, environmentally-induced respiratory disease characterized by reversible airway obstruction, persistent and non-specific airway hyper-responsiveness, and chronic neutrophilic airway inflammation. During seasonal exacerbation, signs vary from mild to life-threatening episodes of wheezing, coughing, and chronic debilitating labored breathing.
Purpose: In human asthma, neutrophilic airway inflammation is associated with more severe and steroid-refractory asthma phenotypes, highlighting a need to decipher the mechanistic basis of this disease characteristic. We hypothesize that the collective biological activities of proteins in bronchoalveolar lavage fluid (BALF) of horses with pasture-associated severe asthma predict changes in neutrophil functions that contribute to airway neutrophilic inflammation.
Methods: Using shotgun proteomics, we identified 1,003 unique proteins in cell-free BALF from six horses experiencing asthma exacerbation and six control herdmates. Contributions of each protein to ten neutrophil functions were modeled using manual biocuration to determine each protein’s net effect on the respective neutrophil functions.
Results: A total of 417 proteins were unique to asthmatic horses, 472 proteins were unique to control horses (p<0.05), and 114 proteins were common in both groups. Proteins whose biological activities are responsible for increasing neutrophil migration, chemotaxis, cell spreading, transmigration, and infiltration, which would collectively bring neutrophils to airways, were over-represented in the BALF of asthmatic relative to control horses. By contrast, proteins whose biological activities support neutrophil activation, adhesion, phagocytosis, respiratory burst, and apoptosis, which would collectively shorten neutrophil lifespan, were under-represented in BALF of asthmatic relative to control horses. Interaction networks generated using Ingenuity® Pathways Analysis further support the results of our biocuration.
Conclusion: Congruent with our hypothesis, the collective biological functions represented in differentially expressed proteins of BALF from horses with pasture-associated severe asthma support neutrophilic airway inflammation. This illustrates the utility of systems modeling to organize functional genomics data in a manner that characterizes complex molecular events associated with clinically relevant disease.
Keywords: pasture-associated severe equine asthma, Equus caballus, horse, proteomics, neutrophil functional genomics
Introduction
Human asthma is a heterogenous disease with varied clinical phenotypes.1 Though Th2-dominated asthma characterized by eosinophilic inflammation2,3 has been well recognized for decades, Th17-mediated neutrophilic airway inflammation has been increasingly recognized in a sizeable group of asthmatic human patients.4–6 However, neutrophilic asthma and eosinophilic asthma are not mutually exclusive. Associations have been established between airway neutrophilic inflammation and increased severity of signs in severe asthma,7–9 corticosteroid-resistant asthma,10–12 asthma exacerbations,13 nocturnal asthma,14 ‘‘asthma in smokers”,15 occupational asthma,16 and “sudden onset” fatal asthma.17 These associations between neutrophils and asthma severity in humans highlight a need to understand how neutrophils contribute to asthma pathophysiology in order to identify new and effective therapeutics.18,19
Two naturally occurring, environmentally-induced asthma syndromes affect outbred populations of animals: severe equine asthma and feline asthma.20,21 Two forms of severe equine asthma are recognized: a barn-associated form, commonly referred to as “heaves”, is exacerbated by mold and dust when horses are maintained in stalls in continental climates,22 while a pasture-associated form (aka recurrent airway obstruction, summer pasture associated recurrent airway obstruction) affects horses grazing pastures during conditions of high heat and humidity and was originally described in the southeastern United States.23 Airway inflammation in both barn-associated, and pasture-associated equine asthma, may be devoid of eosinophils, and is consistently and predominantly neutrophilic.22,24–28 Other features shared with human asthma include genetic linkage,25 reversible bronchoconstriction triggered by environmental aeroallergen challenge, airway remodeling, airway hyper-responsiveness, chronic airway inflammation, and responsiveness to β2-adrenoceptor agonists and corticosteroids. These clinical parallels have led to the recognition of equine asthma as a unique spontaneous model of neutrophilic asthma.22,24–27,29–32
Neutrophilic inflammation has been demonstrated to correlate with poor outcome in other chronic human lung diseases in addition to asthma, including bronchiolitis obliterans in lung transplant recipients,33 and COPD,34 where the correlation is independent of viral or bacterial infection. Neutrophil elastase also correlates with declining lung function in cystic fibrosis.35 Neutrophilic inflammation has been linked to poor prognosis in other organs, for example, intracranial hemorrhage,36 alcoholic hepatitis,37 myocardial infarction,38–42 pulmonary embolism,43 peripheral vascular disease following angioplasty, tumor recurrence in hepatocellular carcinoma,44,45 colorectal tumors,46 and lung cancer.47 Accordingly, identifying proteins that contribute to neutrophilic airway inflammation in horses with severe pasture asthma contributes to the identification of molecular signatures that herald advancing disease, and identifies targets for investigating directed therapies to moderate neutrophilic inflammation.
In systems modeling, homeostasis within a network of eukaryotic proteins (proteostasis) protects the body against disease and allows for healthy cellular development and aging.48 Disease then reflects challenges to the integrity of this network of interacting proteins (proteome) that direct the generation, maintenance, and removal of proteins to achieve normal function. A growing area of airway research centers on the relationship between airway stress diseases, including COPD, emphysema, asthma, and disorders of proteostasis.49 Congruent with the concept of disordered proteostasis, we hypothesize that changes in the protein composition of fluid in the airways of horses with pasture-associated severe asthma are collectively predicted to alter functional attributes of neutrophils in ways that contribute to neutrophilic airway inflammation.
Building upon our prior evidence that protein functions in normal bronchoalveolar lavage fluid (BALF) are conserved across horse, human, and mouse,50 identifying how changes in the BALF proteome contribute to neutrophilic airway inflammation will enable recognition of shared regulatory events in human asthma and severe equine pasture asthma. This in turn informs our understanding of the similar roles of neutrophils in human asthma and severe equine pasture asthma. Here, we employed systems modeling of the BALF proteome of horses experiencing exacerbation of severe pasture asthma, and that of asymptomatic controls exposed to identical aeroallergen challenge and environmental conditions, to evaluate differences in the regulation of ten neutrophil functions: migration, activation, chemotaxis, adhesion, cell spreading, transmigration, infiltration, phagocytosis, respiratory burst, and apoptosis.
Methods and materials
Animals and BALF sample collection
Archived BALF in this study was collected from six horses with pasture-associated severe equine asthma, and six clinically normal control horses of mixed breeds using a bronchoalveolar lavage catheter (240cm, MILA International, Inc., Florence, KY, USA) under standing sedation. The BAL technique has been previously described.28 Samples were immediately placed on ice, transported to the laboratory, centrifuged (600x g for 10 minutes) to derive cell-free BALF, and aliquots of supernate were frozen at −80°C for subsequent proteomic analysis.51,52 All horses were co-housed on pasture, fed identical diets, had no medications within 7 days of sampling, and were sampled in the summer months in Louisiana. Experimental procedures were approved by the Animal Care and Use Committee of Louisiana State University, complying with all federal guidelines overseeing the use of research animals in the United States. All procedures utilized were considered veterinary care best practices.
Horses in this study were a subset of those employed by Costa et al, for which maximal change in pleural pressure (ΔPplmax), BALF cytologic findings, and clinical scores of respiratory effort (CSRE) have been previously described.28 Asthmatic horses ranged from 10–20 years of age (mean ± SD, 16±4.3 years, two females and four castrated males). Control horses ranged from 7–20 years of age (mean 13.5±6.5 years, two females, three castrated males, and one stallion). Diagnosis of pasture-associated severe equine asthma was based on a history of seasonal remission of clinical disease during cool seasons, followed by episodic and reversible obstructive respiratory disease while grazing pasture during hot humid conditions. Overt respiratory distress was characterized by a CSRE ≥4.5, ΔPplmax ≥24cm H2O, audible expiratory wheezes in the lung fields, and neutrophilic airway inflammation (≥12%, mean =66%) in BALF. Control horses lacked historical episodic pasture-associated respiratory disease during the summer, had normal bronchovesicular sounds during lung auscultation with a re-breathing bag, CRSE <3, ΔPplmax ≤9cm H2O, and ≤3%–26% (mean =10%) neutrophils in BALF. Negative bacteriologic culture of BALF was a criterion for inclusion of asthmatic and control horses in this investigation.
Protein isolation, tryptic digestion, and liquid chromatography mass spectrometry/mass spectrometry (LC MS/MS)
Pooled samples containing 100 µg of cell-free BALF protein were created; one from the six control horses and another from the six asthmatic horses. For each pooled sample, 75 µg of protein was analyzed in triplicate using one dimensional LC nanospray ionization as previously described,54 except that we did not perform differential detergent fractionation. Precursor mass scans were performed using repetitive MS scans immediately followed by three MS/MS scans of the three most intense MS peaks.
Protein identification
Searches were performed using TurboSEQUESTTM (Bioworks Browser 3.3, ThermoElectron). Mass spectra and tandem mass spectra were searched against an in silico trypsin-digested database of equine non-redundant RefSeq proteins downloaded from the National Center for Biotechnology Institute (ver 47) as well as against a reversed decoy database. SEQUEST search results were filtered using a decoy search-based probabilistic method, in which only peptides with a probability <0.05 were considered to be correct. Proteins identified with peptides passing the filter criteria were evaluated for differential expression using an Xcorr resampling technique. Probability of differential expression was calculated for each protein and those proteins with a p-value <0.05 were considered differentially expressed.55
Go-based modeling of the effects of BALF proteins on neutrophil function
Systems modeling was performed to test the hypothesis that proteins in the airways of horses with severe pasture asthma are predicted to modify neutrophil functions in a manner that contributes to neutrophilic airway inflammation. Proteins identified in BALF from affected and control horses were biocurated as described in the following pagragraph for their contributions to ten neutrophil functions: migration, activation, chemotaxis, adhesion, cell spreading, transmigration, infiltration, phagocytosis, respiratory burst, and apoptosis. The definitions of these terms are detailed in Table1.
 | Table 1 Definitions of neutrophil terms employed for biocuration |
Proteins were individually biocurated according to the principles of the Gene Ontology (GO) consortium,56 using manual literature searches of each protein in PubMed to identify their effects on the ten neutrophil function terms. Search terms included each protein name paired with the word “neutrophil”. An example query to PubMed would be: “annexin A1” AND “neutrophil”. If more than 1,000 search results were found for any given protein, the search was refined by adding each of the ten neutrophil functions in turn (“annexin A1” AND “neutrophil”, apoptosis). Each relevant paper was read to identify the protein’s effect on the neutrophil term. The effect of each protein on each of the ten neutrophil functions was scored as pro (+1), anti (−1), or no net effect (0), as described in GoModeler.57 In order to model these biological data, the potential of each protein to modify the ten neutrophil functions was approximated by multiplying its expression value by effect (ie, −1, 0, +1), and the net effect of BALF proteins within each functional category was tabulated; similar to the method used for modeling effects on GO terms via GOModeler.57 These net effects are represented graphically for asthmatic and control horses in Figure 1. A list of proteins and their relative effects are provided in
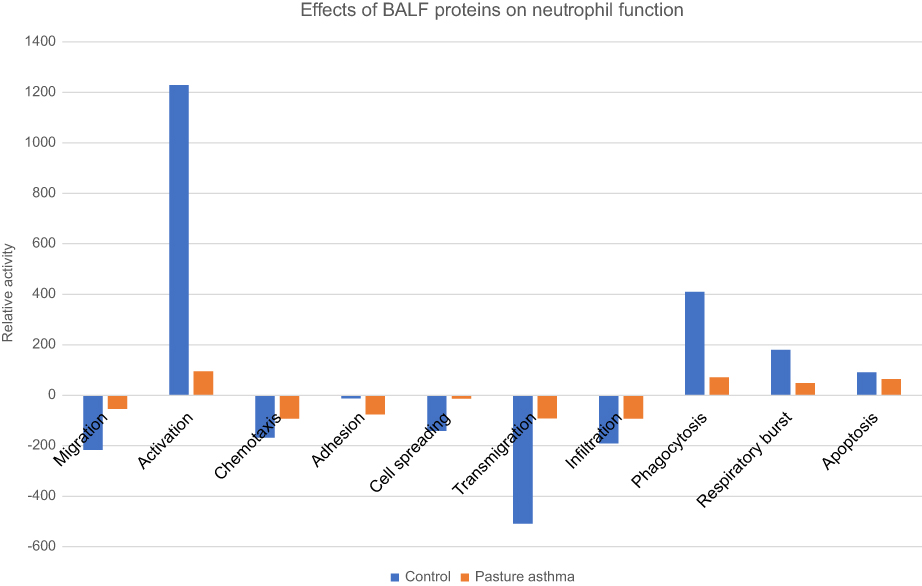 | Figure 1 Cumulative effect of bronchoalveolar lavage fluid (BALF) proteins on neutrophil functions in asthmatic vs control horses. A total of 1,003 proteins were reviewed to identify their effects on ten neutrophil functions. Peer-reviewed manuscripts were read and actions involving ten neutrophil functions were scored for each protein as pro (+1), anti (−1), no effect (0), or no data. The contribution of each protein to each neutrophil function was calculated as the product of magnitude of expression multiplied by its effect (ie, −1, 0, +1), then these values were tabulated for category of neutrophil function. Bars represent the predicted net effect of BALF proteins on the respective neutrophil function. |
Ingenuity® pathway analysis of the effects of BALF proteins on neutrophil functions
To identify central molecules in small sub-networks and support our manual biocuration findings using literature searching in PubMed, proteins were also analyzed for their contributions to neutrophil function using Ingenuity® Pathway Analysis (IPA, Qiagen NV, Venlo, the Netherlands). Neutrophil functions that were curated in IPA included migration, activation, chemotaxis, adhesion, infiltration, phagocytosis, and respiratory burst, and are defined in Table1. Because IPA does not accept equine gene products, human, mouse, and rat orthologs were first identified for each equine protein using Ensembl BioMart© (Ensembl 93 set)58 and then employed for analysis. Most proteins had human orthologs, but mouse, and then rat were used to find orthologs when orthologous human proteins could not be identified. IPA Core Analysis (Ingenuity® Version 01–13, July 2018) was chosen for analysis as all pathways, ontologies, and interactions are supported by manually curated literature, providing a robust and standardized platform for interpreting biologic function from lists of differentially expressed gene products.59
Results
Neutrophil relevant BAL proteome
We identified 1,003 proteins from horse BALF: 417 proteins unique to asthmatic BALF, 472 proteins unique to control BALF, and 114 proteins common to both groups. A total of 108 proteins were identified as significantly differentially expressed between asthmatic and control groups (p<0.05). Using manual biocuration, 93 of the 1,003 proteins identified in BALF were determined to have evidence of biological activities that regulate one or more of ten neutrophil functions. These proteins and their respective effects on neutrophil functions are listed in
Modeling effects of BALF proteins on neutrophil function
By modeling effects of the 93 proteins that were determined, by biocuration, to modify neutrophil functions, increases in neutrophil migration, chemotaxis, cell spreading, transmigration, and infiltration were predicted in asthmatic relative to control horses (Figure1). We found that proteins in control BALF exert a net negative regulation of migration, chemotaxis, cell spreading, transmigration, and infiltration while proteins in BALF of asthmatic horses facilitate a partial release of the negative regulation of these neutrophil functions, as compared to BALF proteins from control horses. This results in a protein milieu in BALF of asthmatic horses that is less inhibitory to migration, chemotaxis, transmigration, and infiltration than the proteins in BALF of control horses. By contrast, a net decrease in protein activities responsible for neutrophil activation, adhesion, phagocytosis, respiratory burst, and apoptosis was identified in the BALF of asthmatic relative to control horses. For these neutrophil functions, there is a net positive regulation of activation, phagocytosis, respiratory burst, and apoptosis by proteins in control BALF, while proteins in asthmatic BALF affect an incomplete decrease in the net positive modulation that characterizes homeostasis. Finally, neutrophil adhesion was decreased by the proteins in both asthmatic and control BALF, though this net effect was comparatively greater in asthmatic than control BALF. A summary of proteins contributing to each of the neutrophil functions segregated by increase, decrease, or no net effect is included within
Neutrophil interactive network
The 1,003 proteins identified in BALF were also modeled for their effects on neutrophil functions using IPA. Of the 1,003 proteins, 129 did not have human, mouse, or rat orthologs, resulting in 874 proteins for IPA analysis. Forty-six proteins were identified to have an influence on neutrophil migration, activation, chemotaxis, adhesion, infiltration, phagocytosis, and respiratory burst. An interaction network of proteins and their impact on neutrophil functions is presented in Figure 2. IPA modeling predicted a net increase in neutrophil migration, adhesion, infiltration, and respiratory burst, and a net decrease in neutrophil activation, chemotaxis, and phagocytosis. Differences in proteins annotated to each neutrophil function in IPA relative to manual curation are detailed in Table 2.
 |  |  |  |  |  |  | Table 2 Comparison of proteins curated by Ingenuity® Pathway Analysis (IPA) vs manual biocuration. Bolded proteins were common between the two methodologies |
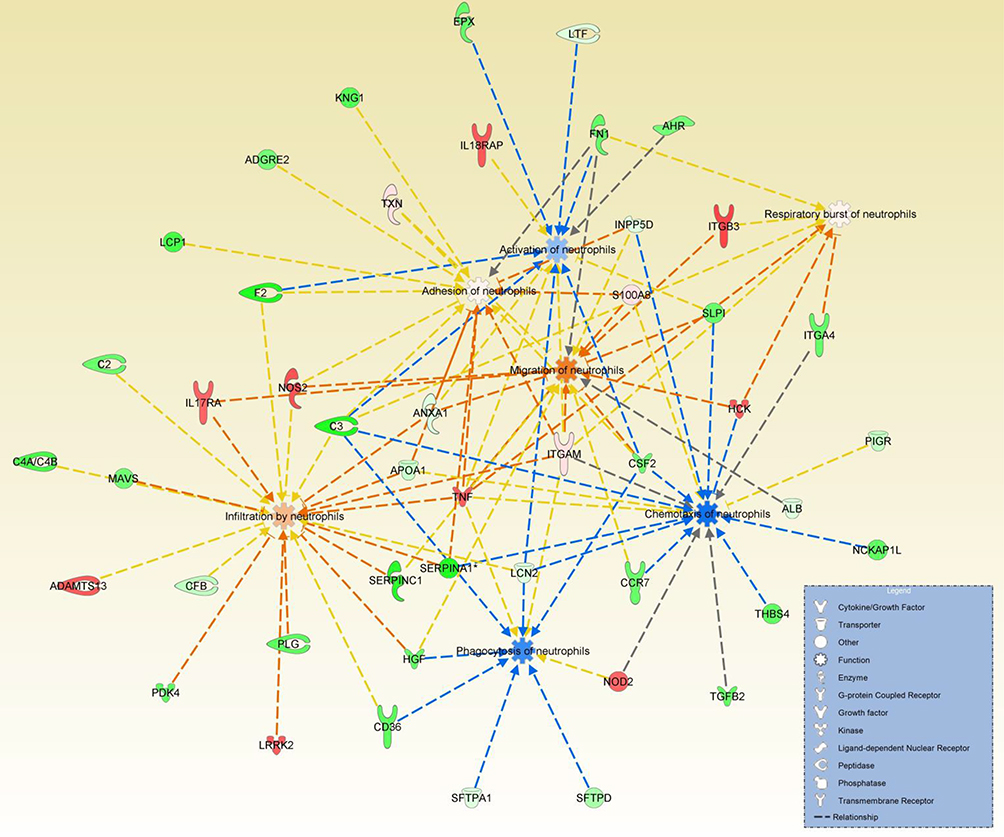 | Figure 2 Ingenuity® Pathway Analysis (IPA) predicted decreases in three and increases in four neutrophil functions. An interactive network was derived using IPA by screening 1,003 proteins in bronchoalveolar lavage fluid (BALF) of asthmatic vs control horses for their impact on neutrophil activity. Green shades indicate relative downregulation of proteins in asthmatic BALF (and relative upregulation in control BALF), while red shades indicate relative upregulation of proteins in asthmatic BALF (and relative downregulation of proteins in control BALF). Blue indicates predicted decreases in neutrophil functions, while orange shades indicate predicted increases. More intense (darker) colors indicate greater increases or decreases. Together, this network indicated net downregulation of neutrophil activation, chemotaxis, and phagocytosis, and upregulation of neutrophil migration, adhesion, infiltration, and respiratory burst. Broken lines indicate a relationship between the proteins and the neutrophil functions. |
Discussion
Though at least 50% of human asthmatics demonstrate neutrophilic inflammation in BALF, which correlates to increased clinical severity and resistance to corticosteroid therapy,1,4,60 regulatory mechanisms responsible for neutrophilic asthma are poorly characterized. Complicating this effort is the difficulty in attaining purified populations of airway neutrophils from human asthmatics, and the fact that bronchoscopy stimulates non-specific airway neutrophilia,61 resulting in a predominance of investigations that employ peripheral blood neutrophils. However, there is evidence that peripheral blood neutrophils do not behave in the same way as airway neutrophils.62–64 Deciphering regulatory mechanisms associated with neutrophilic airway inflammation and the major protein effectors responsible for these changes has the potential to identify molecular events that herald advancing airway disease, as well as avenues for pharmacologic intervention. Systems modeling provides a unique opportunity to translate a large amount of data from high throughput technologies, including proteomics and transcriptomics, into a better understanding of the mechanistic basis of complex biological phenomena and disordered proteostasis.
We report the predicted impact of changes in protein expression in BALF on neutrophil functions in horses with pasture-associated severe equine asthma. This study provides insight into the molecular events that contribute to neutrophilic airway inflammation, a characteristic of both equine asthma and human asthma that is not adequately addressed by rodent models. Our results indicate that, relative to the control BALF, proteins in BALF from asthmatic horses are predicted to increase neutrophil migration, chemotaxis, cell spreading, transmigration, and infiltration. These functions favor the movement of neutrophils into the airways, congruent with airway neutrophilic inflammation that is characteristic of pasture-associated severe equine asthma. Increased neutrophil migration,65 chemotaxis,66,67 cell spreading,68 and infiltration60,69 have similarly been reported in human asthma. Our findings are also consistent with increased neutrophil migration70 and chemotaxis71,72 reported in barn-associated equine asthma. Despite expanding knowledge of how individual proteins modify neutrophil movement in asthma, changes in neutrophil transmigration that occur during neutrophilic forms of asthma are not documented and the majority of knowledge is extrapolated from investigations of neutrophil transmigration across other epithelial surfaces.73 In asthma, increased airway neutrophils are considered a priori evidence of increased neutrophil transmigration.17,74,75 Increased transmigration of neutrophils into the airway is also a feature of several inflammatory diseases, including bronchitis and COPD.74
Apolipoprotein A-1, polymeric immunoglobulin receptor, and haptoglobin were highly expressed proteins in the BALF of control horses. Apolipoprotein A-1 decreases neutrophil migration and cell spreading,76,77 polymeric immunoglobulin receptor decreases neutrophil chemotaxis and transmigration,78 and haptoglobin decreases infiltration.79 Accordingly, decreased expression of these proteins in the BALF of horses with pasture-associated severe asthma relative to expression levels in control BALF had a strong influence on the predicted release of inhibition for these neutrophil functions by proteins in the BALF proteome of diseased horses. Apolipoprotein A-1 modulates lung health and contributes to protective pathways in lung biology, decreasing severity of respiratory diseases including asthma.80–82 This protein has been considered as a potential novel therapeutic agent for asthmatics.80,81,83 Congruent with our findings in the BALF of horses with severe asthma, BALF and serum samples from human asthmatics and murine models also demonstrate decreased levels of apolipoprotein A-1.82,84 Further, in agreement with our findings of increased polymeric immunoglobulin receptor in the BALF of horses with severe asthma, this protein, which transports dimeric IgA across mucosal epithelial cells,85 is also decreased in the bronchial epithelium of human asthma patients.86 Finally, haptoglobin, which was decreased in the BALF of horses with severe asthma, is an acute phase glycoprotein involved in tissue repair and airway remodeling in asthmatic patients.87 While some studies have demonstrated decreased expression in asthmatics relative to controls,87 others have shown expression levels that decrease after initial allergen challenge, but then increase 24 hours post-challenge, suggesting a replenishing of the protein over time.88
Protein expression changes in the BALF proteome of asthmatic horses are predicted, collectively, to decrease neutrophil activation, adhesion, phagocytosis, respiratory burst, and apoptosis. Similar decreases in neutrophil phagocytosis and apoptosis have been identified in asthmatic patients,89–93 and decreased apoptosis of airway neutrophils has been identified in horses with the barn-associated form of severe equine asthma.94,95 Decreased apoptosis contributes to airway neutrophilic inflammation that characterizes human and equine asthma by increasing neutrophil persistence in the airway. Similarly, neutrophil activation and phagocytosis program neutrophils for death,96–98 such that decreasing these activities contributes to neutrophil persistence and neutrophilic airway inflammation. In general, neutrophil adhesion is increased in asthmatic patients, primarily due to an increase in cellular adhesion molecules.99 Our findings, which indicate that the BALF proteome of asthmatic horses would reduce adhesion in asthmatic relative to control horses, contradict these findings in human asthma, but are congruent with the airway neutrophilic inflammation that characterizes equine pasture asthma. This difference may reflect changes in the neutrophilic phenotype induced by ex vivo manipulations or could reflect a compensatory mechanism in the BALF proteome designed to moderate increases in neutrophil adhesion that lead to tissue damage. Significantly, differences may reflect the relatively small sample size of horses included in this study.
While the proteins in BALF of asthmatic horses were predicted to decrease neutrophil activation relative to proteins in the BALF of controls, measures of neutrophil activation are reported to increase in asthma.100–104 However, neutrophil activation is a complex and multifaceted neutrophil function. For example, respiratory burst is one component of neutrophil activation. We identified BALF proteins that both increase and decrease respiratory burst, predicting a net negative effect on this neutrophil function in asthmatic relative to control horses. While characterization of the respiratory burst of airway neutrophils in asthma has been poorly characterized, our findings are congruent with those of Lacy et al (2003), which identified decreased respiratory burst activity in neutrophils from induced sputum of asthmatics relative to controls.63 However, this finding must be considered with the knowledge that ß2-agonists and corticosteroid therapy were not withheld from the asthmatic group, whereas our horses with asthma were not medicated. Other authors have demonstrated that respiratory burst of blood-derived neutrophils from asthmatics exceeds that from non-asthmatics.62,105,106 However, Marcal et al (2004) clarified that significant differences in measures of respiratory burst are influenced by disease severity and are not immediately evident in ex vivo neutrophils, occurring only in more severe asthmatics following ex vivo incubation periods of 25 minutes.62 Despite evidence that selected measures of neutrophil activation are increased in asthma,100–104 our modeling indicated that BALF proteome changes in asthmatic horses favor decreased neutrophil activation, which is consistent with neutrophil persistence in the airways and the airway neutrophilic inflammation that characterizes pasture-associated severe equine asthma. However, based upon aforementioned evidence that neutrophils are activated in human asthma, and that the net decrease in neutrophil activation was the most robust of the predicted effects of BALF proteins on neutrophil functions, we reason that proteins in BALF may work collectively to mitigate neutrophil activation that is driven by exogenous factors in neutrophilic asthma.
We identified four proteins, serotransferrin precursor, polymeric immunoglobulin receptor, surfactant protein A, and cytochrome C, which had strong contributions to the decreases in neutrophil activation, phagocytosis, respiratory burst, and apoptosis predicted by our modeling. Serotransferrin precursor and polymeric immunoglobulin receptor increase neutrophil activation,107,108 surfactant protein A increases neutrophil phagocytosis and respiratory burst,109 and cytochrome C increases apoptosis.110–112 In each case, these proteins demonstrated decreased expression in the BALF of asthmatic relative to control horses. Accordingly, decreases in expression of these four proteins had large effects on decreasing these neutrophil functions in association with asthma in our modeling. Increased expression of thioredoxin in the BALF of diseased horses, which is a negative regulator of neutrophil adhesion,113 was the largest contributor to the net decrease in neutrophil adhesion predicted by our modeling. While cytochrome C levels have not been specifically examined with regards to asthma, both serotransferrin and pulmonary surfactant-associated protein are decreased in the serum of asthmatic patients relative to controls, which is congruent with our equine findings.114–116 Pulmonary surfactant proteins have also been demonstrated to inhibit histamine release during allergen challenge and decrease lymphocyte proliferation in asthmatic children, providing a protective role against the pathogenesis of asthma.117 Thioredoxin, a redox-regulating protein with antioxidant activity, has been observed to be increased in human asthmatics, congruent with findings in asthmatic horses in this investigation.118,119
Functional analysis, performed using IPA, revealed 46 proteins with a known relationship with neutrophil functions (Figure 2). These proteins were predicted by IPA to cause an increase in neutrophil migration and infiltration, and a decrease in neutrophil activation and phagocytosis – predictions which directly corroborate our PubMed manual biocuration predictions. In contrast, IPA predicted an increase in neutrophil adhesion and respiratory burst, and a decrease in chemotaxis, while our manual biocuration findings predicted opposite trends for these functions. As previously indicated, chemotaxis is well documented to be increased in human asthma,66,67 while respiratory burst has been found to be decreased.63 These findings indicate that increased chemotaxis and decreased respiratory burst identified by the manual curation and associated data modeling in our approach are congruent with findings in human asthma. The IPA-predicted increase in neutrophil adhesion contradicts our manual biocuration findings but is congruent with human asthma literature.99 Several differences between the databases that support IPA vs our manual effort are likely relevant to these disparities. First, IPA identified only 46 proteins as relevant to modifying neutrophil functions, while 93 proteins were identified as relevant in our manual curation. Whereas IPA maintains an internal proprietary biocuration effort, our manual biocuration, which was limited to PubMed citations, identified more references that implicated BALF proteins in each neutrophil function, as demonstrated in Table2. Also, IPA relies upon the identification of orthologous human, mouse, or rat proteins from the equine proteins. In our case, 129 proteins that contributed to our curation effort did not, at the time of analysis, have identifiable human, mouse, or rat orthologs, so they were not included in IPA analysis. Of those 129 proteins without an ortholog, our manual biocuration found that eight of them affected neutrophil functions.
Conclusion
Changes in the protein composition of BALF from horses with pasture-associated severe asthma are predicted to increase neutrophil migration, chemotaxis, cell spreading, transmigration, and infiltration, and to decrease neutrophil activation, adhesion, phagocytosis, respiratory burst, and apoptosis. With the exception of adhesion and activation, these predictions mirror changes in neutrophil functions described in both human asthma and barn-associated equine asthma, suggesting that BALF proteins may have a role in moderating the neutrophil activation and adhesion that are reported in human asthma. This investigation confirms our hypothesis that changes in the protein composition of fluid in the airways of horses with pasture-associated severe equine asthma predict functional attributes of neutrophils in the airways of asthmatic horses that contribute to neutrophilic airway inflammation. These data also support the value of pasture-associated severe equine asthma as an animal model of neutrophilic asthma, while demonstrating utility of systems modeling to identify complex molecular events associated with clinical disease in genome scale datasets.
Highlights
Pasture-associated severe equine asthma is characterized by neutrophilic airway inflammation.
Modeling differentially expressed protein functions in BALF from asthmatic horses identifies biologic events that account for neutrophilic airway inflammation.
Systems modeling can organize high throughput data from clinical disease processes in a manner that defines a biologically relevant phenomenon.
Acknowledgments
The authors would like to thank Santosh Kumar TK, Leslie A Shack, Savannah Mack, Courtney Hunter, and Ken Pendarvis for their technical assistance in the laboratory. This work was supported by the College of Veterinary Medicine, Mississippi State University, Mississippi State, MS 39762. Lauren A Bright's present address is Comparative Medicine Resources, Rutgers, The State University of New Jersey, Piscataway, NJ 08854, USA.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wenzel SE. Asthma phenotypes: the evolution from clinical to molecular approaches. Nat Med. 2012;18(5):716–725. doi:10.1038/nm.2678
2. Coleman RA. Current animal models are not predictive for clinical asthma. Pulm Pharmacol Ther. 1999;12(2):87–89. doi:10.1006/pupt.1999.0193
3. Jatakanon A, Lim S, Barnes PJ. Changes in sputum eosinophils predict loss of asthma control. Am J Respir Crit Care Med. 2000;161(1):64–72. doi:10.1164/ajrccm.161.1.9809100
4. Douwes J, Gibson P, Pekkanen J, Pearce N. Non-eosinophilic asthma: importance and possible mechanisms. Thorax. 2002;57(7):643–648.
5. Chang HS, Lee TH, Jun JA, et al. Neutrophilic inflammation in asthma: mechanisms and therapeutic considerations. Expert Rev Respir Med. 2017;11(1):29–40. doi:10.1080/17476348.2017.1268919
6. Lambrecht BN, Hammad H. The immunology of asthma. Nat Immunol. 2015;16(1):45–56. doi:10.1038/ni.3049
7. Jatakanon A, Uasuf C, Maziak W, Lim S, Chung KF, Barnes PJ. Neutrophilic inflammation in severe persistent asthma. Am J Respir Crit Care Med. 1999;160(5 Pt 1):1532–1539. doi:10.1164/ajrccm.160.5.9806170
8. Little SA, MacLeod KJ, Chalmers GW, Love JG, McSharry C, Thomson NC. Association of forced expiratory volume with disease duration and sputum neutrophils in chronic asthma. Am J Med. 2002;112(6):446–452.
9. Wenzel SE, Szefler SJ, Leung DY, Sloan SI, Rex MD, Martin RJ. Bronchoscopic evaluation of severe asthma. Persistent inflammation associated with high dose glucocorticoids. Am J Respir Crit Care Med. 1997;156(3 Pt 1):737–743. doi:10.1164/ajrccm.156.3.9610046
10. Green RH, Brightling CE, Woltmann G, Parker D, Wardlaw AJ, Pavord ID. Analysis of induced sputum in adults with asthma: identification of subgroup with isolated sputum neutrophilia and poor response to inhaled corticosteroids. Thorax. 2002;57(10):875–879.
11. Pavord ID, Brightling CE, Woltmann G, Wardlaw AJ. Non-eosinophilic corticosteroid unresponsive asthma. Lancet. 1999;353(9171):2213–2214. doi:10.1016/S0140-6736(99)01813-9
12. Wenzel SE, Schwartz LB, Langmack EL, et al. Evidence that severe asthma can be divided pathologically into two inflammatory subtypes with distinct physiologic and clinical characteristics. Am J Respir Crit Care Med. 1999;160(3):1001–1008. doi:10.1164/ajrccm.160.3.9812110
13. Fahy JV, Kim KW, Liu J, Boushey HA. Prominent neutrophilic inflammation in sputum from subjects with asthma exacerbation. J Allergy Clin Immunol. 1995;95(4):843–852.
14. Martin RJ, Cicutto LC, Smith HR, Ballard RD, Szefler SJ. Airways inflammation in nocturnal asthma. Am Rev Respir Dis. 1991;143(2):351–357. doi:10.1164/ajrccm/143.2.351
15. Chalmers GW, MacLeod KJ, Thomson L, Little SA, McSharry C, Thomson NC. Smoking and airway inflammation in patients with mild asthma. Chest. 2001;120(6):1917–1922.
16. Anees W, Huggins V, Pavord ID, Robertson AS, Burge PS. Occupational asthma due to low molecular weight agents: eosinophilic and non-eosinophilic variants. Thorax. 2002;57(3):231–236.
17. Sur S, Crotty TB, Kephart GM, et al. Sudden-onset fatal asthma. A distinct entity with few eosinophils and relatively more neutrophils in the airway submucosa? Am Rev Respir Dis. 1993;148(3):713–719. doi:10.1164/ajrccm/148.3.713
18. Bruijnzeel PL, Uddin M, Koenderman L. Targeting neutrophilic inflammation in severe neutrophilic asthma: can we target the disease-relevant neutrophil phenotype? J Leukoc Biol. 2015;98(4):549–556. doi:10.1189/jlb.3VMR1214-600RR
19. Chesne J, Braza F, Mahay G, Brouard S, Aronica M, Magnan A. IL-17 in severe asthma. Where do we stand? Am J Respir Crit Care Med. 2014;190(10):1094–1101. doi:10.1164/rccm.201405-0859PP
20. Fernandes DJ, Xu KF, Stewart AG. Anti-remodelling drugs for the treatment of asthma: requirement for animal models of airway wall remodelling. Clin Exp Pharmacol Physiol. 2001;28(8):619–629.
21. Sagar S, Akbarshahi H, Uller L. Translational value of animal models of asthma: challenges and promises. Eur J Pharmacol. 2015;759:272–277. doi:10.1016/j.ejphar.2015.03.037
22. Robinson NE, Derksen FJ, Olszewski MA, Buechner-Maxwell VA. The pathogenesis of chronic obstructive pulmonary disease of horses. Br Vet J. 1996;152(3):283–306.
23. Seahorn TL, Groves MG, Harrington KS, Beadle RE. Chronic obstructive pulmonary disease in horses in Louisiana. J Am Vet Med Assoc. 1996;208(2):248–251.
24. Derksen FJ, Robinson NE, Armstrong PJ, Stick JA, Slocombe RF. Airway reactivity in ponies with recurrent airway obstruction (heaves). J Appl Physiol (1985). 1985;58(2):598–604. doi:10.1152/jappl.1985.58.2.598
25. Jost U, Klukowska-Rotzler J, Dolf G, et al. A region on equine chromosome 13 is linked to recurrent airway obstruction in horses. Equine Vet J. 2007;39(3):236–241.
26. Kaup FJ, Drommer W, Damsch S, Deegen E. Ultrastructural findings in horses with chronic obstructive pulmonary disease (COPD). II: pathomorphological changes of the terminal airways and the alveolar region. Equine Vet J. 1990;22(5):349–355.
27. Pirie RS, Collie DD, Dixon PM, McGorum BC. Inhaled endotoxin and organic dust particulates have synergistic proinflammatory effects in equine heaves (organic dust-induced asthma). Clin Exp Allergy. 2003;33(5):676–683.
28. Costa LR, Seahorn TL, Moore RM, Taylor HW, Gaunt SD, Beadle RE. Correlation of clinical score, intrapleural pressure, cytologic findings of bronchoalveolar fluid, and histopathologic lesions of pulmonary tissue in horses with summer pasture-associated obstructive pulmonary disease. Am J Vet Res. 2000;61(2):167–173.
29. Leclere M, Lavoie-Lamoureux A, Gelinas-Lymburner E, David F, Martin JG, Lavoie JP. Effect of antigenic exposure on airway smooth muscle remodeling in an equine model of chronic asthma. Am J Respir Cell Mol Biol. 2011;45(1):181–187. doi:10.1165/rcmb.2010-0300OC
30. Leclere M, Lavoie-Lamoureux A, Lavoie JP. Heaves, an asthma-like disease of horses. Respirology. 2011;16(7):1027–1046. doi:10.1111/j.1440-1843.2011.02033.x
31. Bullone M, Lavoie JP. Asthma “of horses and men"–how can equine heaves help us better understand human asthma immunopathology and its functional consequences? Mol Immunol. 2015;66(1):97–105. doi:10.1016/j.molimm.2014.12.005
32. Ferrari CR, Cooley J, Mujahid N, et al. Horses with pasture asthma have airway remodeling that is characteristic of human asthma. Vet Pathol. 2018;55(1):144–158. doi:10.1177/0300985817741729
33. Neurohr C, Huppmann P, Samweber B, et al. Prognostic value of bronchoalveolar lavage neutrophilia in stable lung transplant recipients. J Heart Lung Transplant. 2009;28(5):468–474. doi:10.1016/j.healun.2009.01.014
34. Papi A, Bellettato CM, Braccioni F, et al. Infections and airway inflammation in chronic obstructive pulmonary disease severe exacerbations. Am J Respir Crit Care Med. 2006;173(10):1114–1121. doi:10.1164/rccm.200506-859OC
35. Cohen-Cymberknoh M, Kerem E, Ferkol T, Elizur A. Airway inflammation in cystic fibrosis: molecular mechanisms and clinical implications. Thorax. 2013;68(12):1157–1162. doi:10.1136/thoraxjnl-2013-203204
36. Leira R, Davalos A, Silva Y, et al. Early neurologic deterioration in intracerebral hemorrhage: predictors and associated factors. Neurology. 2004;63(3):461–467.
37. Dominguez M, Miquel R, Colmenero J, et al. Hepatic expression of CXC chemokines predicts portal hypertension and survival in patients with alcoholic hepatitis. Gastroenterology. 2009;136(5):1639–1650. doi:10.1053/j.gastro.2009.01.056
38. Chia S, Nagurney JT, Brown DF, et al. Association of leukocyte and neutrophil counts with infarct size, left ventricular function and outcomes after percutaneous coronary intervention for ST-elevation myocardial infarction. Am J Cardiol. 2009;103(3):333–337. doi:10.1016/j.amjcard.2008.09.085
39. Papa A, Emdin M, Passino C, Michelassi C, Battaglia D, Cocci F. Predictive value of elevated neutrophil-lymphocyte ratio on cardiac mortality in patients with stable coronary artery disease. Clin Chim Acta. 2008;395(1–2):27–31. doi:10.1016/j.cca.2008.04.019
40. Takahashi T, Hiasa Y, Ohara Y, et al. Relationship of admission neutrophil count to microvascular injury, left ventricular dilation, and long-term outcome in patients treated with primary angioplasty for acute myocardial infarction. Circ J. 2008;72(6):867–872.
41. Tamhane UU, Aneja S, Montgomery D, Rogers EK, Eagle KA, Gurm HS. Association between admission neutrophil to lymphocyte ratio and outcomes in patients with acute coronary syndrome. Am J Cardiol. 2008;102(6):653–657. doi:10.1016/j.amjcard.2008.05.006
42. Nunez J, Nunez E, Bodi V, et al. Usefulness of the neutrophil to lymphocyte ratio in predicting long-term mortality in ST segment elevation myocardial infarction. Am J Cardiol. 2008;101(6):747–752. doi:10.1016/j.amjcard.2007.11.004
43. Kayrak M, Erdogan HI, Solak Y, et al. Prognostic value of neutrophil to lymphocyte ratio in patients with acute pulmonary embolism: a restrospective study. Heart Lung Circ. 2014;23(1):56–62. doi:10.1016/j.hlc.2013.06.004
44. Halazun KJ, Hardy MA, Rana AA, et al. Negative impact of neutrophil-lymphocyte ratio on outcome after liver transplantation for hepatocellular carcinoma. Ann Surg. 2009;250(1):141–151. doi:10.1097/SLA.0b013e3181a77e59
45. Gomez D, Farid S, Malik HZ, et al. Preoperative neutrophil-to-lymphocyte ratio as a prognostic predictor after curative resection for hepatocellular carcinoma. World J Surg. 2008;32(8):1757–1762. doi:10.1007/s00268-008-9552-6
46. Kishi Y, Kopetz S, Chun YS, Palavecino M, Abdalla EK, Vauthey JN. Blood neutrophil-to-lymphocyte ratio predicts survival in patients with colorectal liver metastases treated with systemic chemotherapy. Ann Surg Oncol. 2009;16(3):614–622. doi:10.1245/s10434-008-0267-6
47. Teramukai S, Kitano T, Kishida Y, et al. Pretreatment neutrophil count as an independent prognostic factor in advanced non-small-cell lung cancer: an analysis of Japan multinational trial organisation LC00-03. Eur J Cancer. 2009;45(11):1950–1958. doi:10.1016/j.ejca.2009.01.023
48. Balch WE, Morimoto RI, Dillin A, Kelly JW. Adapting proteostasis for disease intervention. Science. 2008;319(5865):916–919. doi:10.1126/science.1141448
49. Bouchecareilh M, Balch WE. Proteostasis, an emerging therapeutic paradigm for managing inflammatory airway stress disease. Curr Mol Med. 2012;12(7):815–826.
50. Bright LA, Mujahid N, Nanduri B, et al. Functional modelling of an equine bronchoalveolar lavage fluid proteome provides experimental confirmation and functional annotation of equine genome sequences. Anim Genet. 2011;42(4):395–405. doi:10.1111/j.1365-2052.2010.02158.x
51. Costa LR, Eades SC, Venugopal CS, Moore RM. Plasma and pulmonary fluid endothelin in horses with seasonal recurrent airway obstruction. J Vet Intern Med. 2009;23(6):1239–1246. doi:10.1111/j.1939-1676.2009.0385.x
52. Bright LA, Burgess SC, Chowdhary B, Swiderski CE, McCarthy FM. Structural and functional-annotation of an equine whole genome oligoarray. BMC Bioinformatics. 2009;10(Suppl 11):S8. doi:10.1186/1471-2105-10-S11-S8
53. Seahorn TL, Beadle RE, McGorum BC, Marley CL. Quantification of antigen-specific antibody concentrations in tracheal lavage fluid of horses with summer pasture-associated obstructive pulmonary disease. Am J Vet Res. 1997;58(12):1408–1411.
54. McCarthy FM, Cooksey AM, Wang N, Bridges SM, Pharr GT, Burgess SC. Modeling a whole organ using proteomics: the avian bursa of Fabricius. Proteomics. 2006;6(9):2759–2771. doi:10.1002/pmic.200500648
55. Pendarvis K, Kumar R, Burgess SC, Nanduri B. An automated proteomic data analysis workflow for mass spectrometry. BMC Bioinformatics. 2009;10(Suppl 11):S17. doi:10.1186/1471-2105-10-S11-S17
56. McCarthy FM, Bridges SM, Burgess SC. GOing from functional genomics to biological significance. Cytogenet Genome Res. 2007;117(1–4):278–287. doi:10.1159/000103189
57. Manda P, Freeman MG, Bridges SM, et al. GOModeler–a tool for hypothesis-testing of functional genomics datasets. BMC Bioinformatics. 2010;11(Suppl 6):S29. doi:10.1186/1471-2105-11-S6-S29
58. Durinck S, Moreau Y, Kasprzyk A, et al. BioMart and Bioconductor: a powerful link between biological databases and microarray data analysis. Bioinformatics. 2005;21(16):3439–3440. doi:10.1093/bioinformatics/bti525
59. Kramer A, Green J, Pollard J
60. Nakagome K, Matsushita S, Nagata M. Neutrophilic inflammation in severe asthma. Int Arch Allergy Immunol. 2012;158(Suppl 1):96–102. doi:10.1159/000337801
61. Ndukwu IM, Naureckas ET, Maxwell C, Waldman M, Leff AR. Relationship of cellular transmigration and airway response after allergen challenge. Am J Respir Crit Care Med. 1999;160(5 Pt 1):1516–1524. doi:10.1164/ajrccm.160.5.9812126
62. Marcal LE, Rehder J, Newburger PE, Condino-Neto A. Superoxide release and cellular gluthatione peroxidase activity in leukocytes from children with persistent asthma. Braz J Med Biol Res. 2004;37(11):1607–1613.S0100-879X2004001100003
63. Lacy P, Abdel-Latif D, Steward M, Musat-Marcu S, Man SF, Moqbel R. Divergence of mechanisms regulating respiratory burst in blood and sputum eosinophils and neutrophils from atopic subjects. J Immunol. 2003;170(5):2670–2679.
64. Bedi P, Davidson DJ, McHugh BJ, Rossi AG, Hill AT. Blood neutrophils are reprogrammed in bronchiectasis. Am J Respir Crit Care Med. 2018. doi:10.1164/rccm.201712-2423OC
65. Oreo KM, Gibson PG, Simpson JL, Wood LG, McDonald VM, Baines KJ. Sputum ADAM8 expression is increased in severe asthma and COPD. Clin Exp Allergy. 2014;44(3):342–352. doi:10.1111/cea.12223
66. Radeau T, Chavis C, Damon M, Michel FB. Crastes de Paulet A, Godard PH. Enhanced arachidonic acid metabolism and human neutrophil migration in asthma. Prostaglandins Leukot Essent Fatty Acids. 1990;41(2):131–138.
67. Lavinskiene S, Bajoriuniene I, Malakauskas K, Jeroch J, Sakalauskas R. Sputum neutrophil count after bronchial allergen challenge is related to peripheral blood neutrophil chemotaxis in asthma patients. Inflamm Res. 2014;63(11):951–959. doi:10.1007/s00011-014-0770-0
68. Ozier A, Allard B, Bara I, et al. The pivotal role of airway smooth muscle in asthma pathophysiology. J Allergy (Cairo). 2011;2011:742710.
69. Nissen G, Hollaender H, Tang FSM, et al. Tumstatin fragment selectively inhibits neutrophil infiltration in experimental asthma exacerbation. Clin Exp Allergy. 2018. doi:10.1111/cea.13236
70. Bullone M, Moran K, Lavoie-Lamoureux A, Lavoie JP. PI3K and MAPKs regulate neutrophil migration toward the airways in heaves. J Vet Intern Med. 2013;27(1):164–170. doi:10.1111/jvim.12008
71. Franchini M, Gill U, von Fellenberg R, Bracher VD. Interleukin-8 concentration and neutrophil chemotactic activity in bronchoalveolar lavage fluid of horses with chronic obstructive pulmonary disease following exposure to hay. Am J Vet Res. 2000;61(11):1369–1374.
72. Tessier L, Cote O, Clark ME, et al. Impaired response of the bronchial epithelium to inflammation characterizes severe equine asthma. BMC Genomics. 2017;18(1):708. doi:10.1186/s12864-016-3396-5
73. Kumar SD, Krishnamurthy K, Manikandan J, Pakeerappa PN, Pushparaj PN. Deciphering the key molecular and cellular events in neutrophil transmigration during acute inflammation. Bioinformation. 2011;6(3):111–114.
74. Kidney JC, Proud D. Neutrophil transmigration across human airway epithelial monolayers: mechanisms and dependence on electrical resistance. Am J Respir Cell Mol Biol. 2000;23(3):389–395. doi:10.1165/ajrcmb.23.3.4068
75. Martin TR, Raghu G, Maunder RJ, Springmeyer SC. The effects of chronic bronchitis and chronic air-flow obstruction on lung cell populations recovered by bronchoalveolar lavage. Am Rev Respir Dis. 1985;132(2):254–260. doi:10.1164/arrd.1985.132.2.254
76. Murphy AJ, Woollard KJ, Suhartoyo A, et al. Neutrophil activation is attenuated by high-density lipoprotein and apolipoprotein A-I in in vitro and in vivo models of inflammation. Arterioscler Thromb Vasc Biol. 2011;31(6):1333–1341. doi:10.1161/ATVBAHA.111.226258
77. Montecucco F, Vuilleumier N, Pagano S, et al. Anti-Apolipoprotein A-1 auto-antibodies are active mediators of atherosclerotic plaque vulnerability. Eur Heart J. 2011;32(4):412–421. doi:10.1093/eurheartj/ehq521
78. Marshall LJ, Perks B, Ferkol T, Shute JK. IL-8 released constitutively by primary bronchial epithelial cells in culture forms an inactive complex with secretory component. J Immunol. 2001;167(5):2816–2823.
79. Arslan F, Smeets MB, Buttari B, et al. Lack of haptoglobin results in unbalanced VEGFalpha/angiopoietin-1 expression, intramural hemorrhage and impaired wound healing after myocardial infarction. J Mol Cell Cardiol. 2013;56:116–128. doi:10.1016/j.yjmcc.2012.12.012
80. Yao X, Gordon EM, Figueroa DM, Barochia AV, Levine SJ. Emerging roles of Apolipoprotein E and Apolipoprotein A-I in the pathogenesis and treatment of lung disease. Am J Respir Cell Mol Biol. 2016;55(2):159–169. doi:10.1165/rcmb.2016-0060TR
81. Yao X, Gordon EM, Barochia AV, Remaley AT, Levine SJ. The A‘s have it: developing apolipoprotein A-I mimetic peptides into a novel treatment for asthma. Chest. 2016;150(2):283–288. doi:10.1016/j.chest.2016.05.035
82. Barochia AV, Kaler M, Cuento RA, et al. Serum apolipoprotein A-I and large high-density lipoprotein particles are positively correlated with FEV1 in atopic asthma. Am J Respir Crit Care Med. 2015;191(9):990–1000. doi:10.1164/rccm.201411-1990OC
83. Gordon EM, Figueroa DM, Barochia AV, Yao X, Levine SJ. High-density lipoproteins and Apolipoprotein A-I: potential new players in the prevention and treatment of lung disease. Front Pharmacol. 2016;7:323. doi:10.3389/fphar.2016.00323
84. Park SW, Lee EH, Lee EJ, et al. Apolipoprotein A1 potentiates lipoxin A4 synthesis and recovery of allergen-induced disrupted tight junctions in the airway epithelium. Clin Exp Allergy. 2013;43(8):914–927.
85. Asano M, Komiyama K. Polymeric immunoglobulin receptor. J Oral Sci. 2011;53(2):147–156.
86. Ladjemi MZ, Gras D, Dupasquier S, et al. Bronchial epithelial IgA secretion is impaired in asthma. Role of IL-4/IL-13. Am J Respir Crit Care Med. 2018;197(11):1396–1409. doi:10.1164/rccm.201703-0561OC
87. Larsen K, Macleod D, Nihlberg K, et al. Specific haptoglobin expression in bronchoalveolar lavage during differentiation of circulating fibroblast progenitor cells in mild asthma. J Proteome Res. 2006;5(6):1479–1483. doi:10.1021/pr050462h
88. Kim CK, Chung CY, Koh YY. Changes in serum haptoglobin level after allergen challenge test in asthmatic children. Allergy. 1998;53(2):184–189.
89. Huynh ML, Malcolm KC, Kotaru C, et al. Defective apoptotic cell phagocytosis attenuates prostaglandin E2 and 15-hydroxyeicosatetraenoic acid in severe asthma alveolar macrophages. Am J Respir Crit Care Med. 2005;172(8):972–979. doi:10.1164/rccm.200501-035OC
90. Uddin M, Nong G, Ward J, et al. Prosurvival activity for airway neutrophils in severe asthma. Thorax. 2010;65(8):684–689. doi:10.1136/thx.2009.120741
91. Parfrey H, Farahi N, Porter L, Chilvers ER. Live and let die: is neutrophil apoptosis defective in severe asthma? Thorax. 2010;65(8):665–667. doi:10.1136/thx.2009.134270
92. Alexis NE, Eldridge MW, Peden DB. Effect of inhaled endotoxin on airway and circulating inflammatory cell phagocytosis and CD11b expression in atopic asthmatic subjects. J Allergy Clin Immunol. 2003;112(2):353–361.
93. Saffar AS, Alphonse MP, Shan L, Hayglass KT, Simons FE, Gounni AS. IgE modulates neutrophil survival in asthma: roleof mitochondrial pathway. J Immunol. 2007;178(4):2535–2541.
94. Niedzwiedz A, Jaworski Z, Tykalowski B, Smialek M. Neutrophil and macrophage apoptosis in bronchoalveolar lavage fluid from healthy horses and horses with recurrent airway obstruction (RAO). BMC Vet Res. 2014;10:29. doi:10.1186/1746-6148-10-1
95. Brazil TJ, Dagleish MP, McGorum BC, Dixon PM, Haslett C, Chilvers ER. Kinetics of pulmonary neutrophil recruitment and clearance in a natural and spontaneously resolving model of airway inflammation. Clin Exp Allergy. 2005;35(7):854–865. doi:10.1111/j.1365-2222.2005.02231.x
96. Zhang B, Hirahashi J, Cullere X, Mayadas TN. Elucidation of molecular events leading to neutrophil apoptosis following phagocytosis: cross-talk between caspase 8, reactive oxygen species, and MAPK/ERK activation. J Biol Chem. 2003;278(31):28443–28454. doi:10.1074/jbc.M210727200
97. Coxon A, Rieu P, Barkalow FJ, et al. A novel role for the beta 2 integrin CD11b/CD18 in neutrophil apoptosis: a homeostatic mechanism in inflammation. Immunity. 1996;5(6):653–666.
98. Watson RW, Redmond HP, Wang JH, Condron C, Bouchier-Hayes D. Neutrophils undergo apoptosis following ingestion of Escherichia coli. J Immunol. 1996;156(10):3986–3992.
99. Gao H, Ying S, Dai Y. Pathological roles of neutrophil-mediated inflammation in asthma and its potential for therapy as a target. J Immunol Res. 2017;2017:3743048. doi:10.1155/2017/5974574
100. Mann BS, Chung KF. Blood neutrophil activation markers in severe asthma: lack of inhibition by prednisolone therapy. Respir Res. 2006;7:59. doi:10.1186/1465-9921-7-59
101. Ciepiela O, Ostafin M, Demkow U. Neutrophils in asthma–a review. Respir Physiol Neurobiol. 2015;209:13–16. doi:10.1016/j.resp.2014.12.004
102. Durham SR, Carroll M, Walsh GM, Kay AB. Leukocyte activation in allergen-induced late-phase asthmatic reactions. N Engl J Med. 1984;311(22):1398–1402. doi:10.1056/NEJM198411293112202
103. Ward C, Kelly CA, Stenton SC, Duddridge M, Hendrick DJ, Walters EH. The relative contribution of bronchoalveolar macrophages and neutrophils to lucigenin- and luminol-amplified chemiluminescence. Eur Respir J. 1990;3(9):1008–1014.
104. Kallenbach J, Baynes R, Fine B, Dajee D, Bezwoda W. Persistent neutrophil activation in mild asthma. J Allergy Clin Immunol. 1992;90(2):272–274.
105. Monteseirin J, Camacho MJ, Bonilla I, et al. Respiratory burst in neutrophils from asthmatic patients. J Asthma. 2002;39(7):619–624.
106. Teramoto S, Shu CY, Ouchi Y, Fukuchi Y. Increased spontaneous production and generation of superoxide anion by blood neutrophils in patients with asthma. J Asthma. 1996;33(3):149–155.
107. Sakai C, Miura R, Shibata A. [Activation of neutrophil alkaline phosphatase of chronic myelogenous leukemia in vitro liquid culture: transferrin as a NAP-activating factor]. Nihon Ketsueki Gakkai Zasshi. 1987;50(3):582–588.
108. Wettero J, Bengtsson T, Tengvall P. C1q-independent activation of neutrophils by immunoglobulin M-coated surfaces. J Biomed Mater Res. 2001;57(4):550–558.
109. Madan T, Eggleton P, Kishore U, et al. Binding of pulmonary surfactant proteins A and D to Aspergillus fumigatus conidia enhances phagocytosis and killing by human neutrophils and alveolar macrophages. Infect Immun. 1997;65(8):3171–3179.
110. Hokari Y, Seki T, Nakano H, et al. Isolation and identification of novel neutrophil-activating cryptides hidden in mitochondrial cytochrome C. Protein Pept Lett. 2012;19(6):680–687.
111. Murphy BM, O‘Neill AJ, Adrain C, Watson RW, Martin SJ. The apoptosome pathway to caspase activation in primary human neutrophils exhibits dramatically reduced requirements for cytochrome C. J Exp Med. 2003;197(5):625–632.
112. Pryde JG, Walker A, Rossi AG, Hannah S, Haslett C. Temperature-dependent arrest of neutrophil apoptosis. Failure of Bax insertion into mitochondria at 15 degrees C prevents the release of cytochrome c. J Biol Chem. 2000;275(43):33574–33584. doi:10.1074/jbc.M001008200
113. Nakamura H, Herzenberg LA, Bai J, et al. Circulating thioredoxin suppresses lipopolysaccharide-induced neutrophil chemotaxis. Proc Natl Acad Sci U S A. 2001;98(26):15143–15148. doi:10.1073/pnas.191498798
114. Bakkeheim E, Mowinckel P, Carlsen KH, Burney P, Carlsen KC. Altered oxidative state in schoolchildren with asthma and allergic rhinitis. Pediatr Allergy Immunol. 2011;22(2):178–185. doi:10.1111/j.1399-3038.2010.01092.x
115. Brigham EP, McCormack MC, Takemoto CM, Matsui EC. Iron status is associated with asthma and lung function in US women. PLoS One. 2015;10(2):e0117545. doi:10.1371/journal.pone.0117545
116. Widmeier S, Bernard A, Tschopp A, et al. Surfactant protein A, exposure to endotoxin, and asthma in garbage collectors and in wastewater workers. Inhal Toxicol. 2007;19(4):351–360. doi:10.1080/08958370601144456
117. Wang JY, Shieh CC, You PF, Lei HY, Reid KB. Inhibitory effect of pulmonary surfactant proteins A and D on allergen-induced lymphocyte proliferation and histamine release in children with asthma. Am J Respir Crit Care Med. 1998;158(2):510–518. doi:10.1164/ajrccm.158.2.9709111
118. Yamada Y, Nakamura H, Adachi T, et al. Elevated serum levels of thioredoxin in patients with acute exacerbation of asthma. Immunol Lett. 2003;86(2):199–205.
119. Jeong HC, Lee SY, Lee EJ, et al. Proteomic analysis of peripheral T-lymphocytes in patients with asthma. Chest. 2007;132(2):489–496. doi:10.1378/chest.06-2980
120.
121. Marks RM, Todd RF
122. Reinhardt PH, Ward CA, Giles WR, Kubes P. Emigrated rat neutrophils adhere to cardiac myocytes via alpha 4 integrin. Circ Res. 1997;81(2):196–201.
123.
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.