")
Back to Journals » OncoTargets and Therapy » Volume 11
miRNA-641 inhibits the proliferation, migration, and invasion and induces apoptosis of cervical cancer cells by directly targeting ZEB1
Authors Yao R, Zheng H , Wu L, Cai P
Received 9 October 2018
Accepted for publication 27 October 2018
Published 11 December 2018 Volume 2018:11 Pages 8965—8976
DOI https://doi.org/10.2147/OTT.S190303
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjay Singh
This paper has been retracted
Rui Yao, Huzhong Zheng, Liqun Wu, Pingsheng Cai
Department of Obstetrics and Gynecology, Wenzhou Hospital of Integrated Traditional Chinese and Western Medicine, Wenzhou 325000, Zhejiang, People’s Republic of China
Background: miRNAs have been found to be dysregulated in cervical cancer. The dysregulation of miRNA has been implicated in cervical carcinogenesis and progression. Therefore, further studies of the specific roles of deregulated miRNAs in cervical cancer and underlying molecular mechanisms may facilitate the identification of novel therapeutic techniques for patients with this disease. miRNA-641 (miR-641) was previously reported to serve an important role in lung cancer. However, the expression pattern and roles of miR-641 in cervical cancer remain unclear.
Method: In this study, the expression level of miR-641 in cervical cancer tissues and cell lines was detected using RT-qPCR. The influence of miR-641 upregulation in cervical cancer cell proliferation, apoptosis, migration and invasion was evaluated using CCK-8 assay, flow cytometry assay, migration and invasion assays, respectively. In vivo tumor growth assay was utilized to determine the effect of miR-641 overexpression in the tumor growth of cervical cancer cells in vivo. The molecular mechanisms underlying the action of miR-641 in cervical cancer cells were also explored.
Results: We found that miR-641 expression was obviously decreased in cervical cancer tissues and cell lines, which strongly correlated with the International Federation of Gynecology and Obstetrics stage and lymph node metastasis. Upregulation of miR-641 inhibited cell proliferation, induced apoptosis, and reduced metastasis in cervical cancer. Additionally, bioinformatics analysis predicted ZEB1 as a novel target gene of miR-641. Notably, luciferase reporter assay, RT-qPCR, and Western blot analysis revealed that miR-641 decreased ZEB1 expression in cervical cancer cells by directly targeting its 3'-untranslated region. Furthermore, ZEB1 was upregulated in cervical cancer tissues, which was negatively correlated with miR-641 expression. Moreover, recovered ZEB1 expression attenuated the tumor suppressive action of miR-641 overexpression in the malignant phenotypes of cervical cancer cells. Besides, miR-641 could hinder cervical cancer tumor growth in vivo by inhibiting ZEB1.
Conclusion: These results indicate that miR-641 has tumor suppressive roles in the development of cervical cancer by directly targeting ZEB1, suggesting that miR-641 is a novel, effective therapeutic target for treating patients with this disease.
Keywords: microRNA-641, target therapy, zinc finger E-Box binding homeobox 1, aggressive behaviors
Introduction
Cervical cancer, one of the most common gynecological malignancies, is the fourth leading cause of cancer-related death worldwide.1 In the People’s Republic of China, the morbidity and mortality rates of cervical cancer in recent years have gradually increased each year.2 Approximately 98,900 novel cervical cancer cases were diagnosed and 30,500 deaths were caused by cervical cancer in 2015.3 Infection with high-risk human papillomavirus plays a crucial role in the pathogenesis of cervical cancer;4 however, the detailed molecular mechanisms remain unclear. Despite remarkable improvements in diagnosis and therapy, the prognosis of patients with cervical cancer remains unsatisfactory, with a 5-year survival rate of less than 40%.5,6 Recurrence and metastasis are the main factors affecting the poor therapeutic outcomes of patients with cervical cancer.7 Therefore, studies are needed to determine the mechanism responsible for cervical carcinogenesis and progression, which may be particularly useful for identifying attractive therapeutic targets for treating patients with this malignancy.
miRNAs are a subset of conserved, single-strand, non-coding, and short RNA molecules 18–24 nucleotides in length.8 miRNAs have been demonstrated to negatively regulate gene expression by directly interacting with the 3′-untranslated regions (3′-UTRs) of their target genes to cause mRNA degradation and/or translational suppression.9 More than 2,500 miRNAs have been validated in the human genome.10 Recently, miRNAs were found to be dysregulated in most human cancer types, such as lung cancer,11 colorectal cancer,12 breast cancer,13 and bladder cancer.14 Particularly, a variety of miRNAs are aberrantly expressed in cervical cancer.15 Aberrant expression of miRNAs is important in the malignant development of cervical cancer by dysregulation of multiple cellular biological processes, including cell proliferation, cell cycle, apoptosis, metastasis, motility, and angiogenesis.16,17 miRNAs may play oncogenic or tumor suppressive roles in the genesis and progression of cervical cancer, which are mainly attributable to the biological roles of their target genes.18 Therefore, miRNAs may be effective diagnostic and therapeutic targets for treating patients with cervical cancer.
miRNA-641 (miR-641) was found to play an important role in lung cancer.19,20 However, the expression pattern and roles of miR-641 in cervical cancer remain unclear. In this study, we detected miR-641 expression in cervical cancer, examined its role in malignant phenotypes, and investigated its underlying mechanisms. Our results indicate that miR-641 can be developed as a molecular target to treat patients with cervical cancer.
Material and methods
Clinical specimens and cell lines
Paired cervical cancer and adjacent non-tumor tissues were collected from 51 patients with cervical cancer who had undergone surgical resection at Wenzhou Hospital of Integrated Traditional Chinese and Western Medicine between June 2015 and April 2017. No patients had been treated with preoperative radiotherapy or chemotherapy. Fresh tissues were quickly snap-frozen in liquid nitrogen after surgical removal and then stored at −80°C until use. This study was granted approval by Ethics Committee of Wenzhou Hospital of Integrated Traditional Chinese and Western Medicine, and was performed in accordance with the Declaration of Helsinki and the guidelines of the Ethics Committee of Wenzhou Hospital of Integrated Traditional Chinese and Western Medicine. Written informed consent was provided by all individuals enrolled in this study.
A normal human cervix epithelial cell line (Ect1/E6E7) and four cervical cancer cell lines (C-33A, HeLa, SiHa, and CaSki) were purchased from American Type Culture Collection (Manassas, VA, USA). DMEM containing 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin (all from Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) was used for cell culture. Cultures were maintained in a humidified incubator at 37°C supplied with 5% CO2.
Oligonucleotide and plasmid transfection
miR-641 mimics and negative control miRNA mimics (miR-NC) were chemically synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, People’s Republic of China). siRNA against the expression of ZEB1 (ZEB1 siRNA) and negative control siRNA (NC siRNA) were obtained from Guangzhou RiboBio Co., Ltd. (Guangzhou, People’s Republic of China). The ZEB1 siRNA sequence was 5′-GCUGCCAAUAAGCAAACGA-3′ and the NC siRNA sequence was 5′-UUCUCCGAACGUGUCACGUTT-3′. The ZEB1 overexpression vector pcDNA3.1-ZEB1 (pc-ZEB1) and pcDNA3.1 blank vector were produced by the Chinese Academy of Sciences (Changchun, People’s Republic of China). For cell transfection, the cells were inoculated into 6-well plates at an initial density of 6×105 cells/well. Lipofectamine 2000 reagent (Invitrogen, Thermo Fisher Scientific) was used for all transfections in accordance with the manufacturer’s instructions.
RT-qPCR
RT-qPCR was conducted to detect miR-641 and ZEB1 mRNA levels. Total RNA was extracted from tissue specimens or cells using TRIzol reagent (Invitrogen, Thermo Fisher Scientific) according to the manufacturer’s protocol. To evaluate miR-641 expression, cDNA was produced from total RNA using a miScript Reverse Transcription kit (Qiagen NV, Venlo, the Netherlands). Subsequently, quantitative PCR was conducted using a miScript SYBR-Green PCR kit (Qiagen NV). To measure ZEB1 mRNA expression, total RNA was reverse-transcribed into cDNA using a PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd., Shiga, Japan). Next, SYBR Premix Ex Taq™ (Takara Biotechnology Co., Ltd.) was used to perform qPCR according to the manufacturer’s instructions. Relative miR-641 and ZEB1 mRNA expression were normalized to that of U6 snRNA and GAPDH, respectively. The primers were designed as follows: miR-641, 5′-TTATACTCTCACCATTTGGATC-3′ (forward) and 5′-TGACAAGATTTTACATCAAGAA-3′ (reverse); U6, 5′-CTTCGGCAGCACATATACT-3′ (forward) and 5′-AAAATATGGAACGCTTCACG-3′ (reverse); ZEB1, 5′-TTGTAGCGACTGGATTTT-3′ (forward) and 5′-AGACGATAGTTGGGTCCCGGC-3′ (reverse); and GAPDH, 5′-CGGAGTCAACGGATTTGGTCGTAT-3′ (forward) and 5′-AGCCTTCTCCATGGTGGTGAAGAC-3′ (reverse). Relative gene expression was calculated using the 2−ΔΔCq method.21
Cell counting kit-8 (CCK-8) assay
Transfected cells were collected after 24 hours of incubation at 37°C with 5% CO2. Cells were resuspended and plated into 96-well plates at a density of 3×103 cells/well. Cellular proliferation was assessed by conducting a CCK-8 assay (Dojindo, Kumamoto, Japan) at four time points: 0, 1, 2, and 3 days after incubation. A total of 10 μL CCK-8 reagent was added to each well and incubated at 37°C with 5% CO2 for another 2 hours. Finally, the optical density of each well was measured at a wavelength of 450 nm using a microplate reader (Bio-Rad Laboratories Inc., Hercules, CA, USA).
Flow cytometry assay
Transfected cells were harvested at 48 hours post-transfection and washed twice with ice-cold PBS. An Annexin V fluorescein isothiocyanate (FITC) apoptosis detection kit (Biolegend, San Diego, CA, USA) was utilized to evaluate apoptosic rate. Briefly, transfected cells were stained in the dark with 5 μL of Annexin V FITC and 5 μL of propidium iodide diluted in 100 μL of binding buffer. Following incubation at room temperature for 20 minutes, the percentage of apoptotic cells was detected by flow cytometry (FACScan; BD Biosciences, San Jose, CA, USA).
Migration and invasion assays
For migration assay, cells were collected after 48 hours of transfection, suspended in FBS-free DMEM, and inoculated into the upper compartment of Transwell chambers (24-well insert; pore size, 8 μm; Corning Incorporated, Corning, NY, USA). The lower compartments were covered with 500 μL DMEM containing 20% FBS to serve as the chemoattractant. Following incubation for 24 hours, non-invading cells were carefully removed with a cotton swab, whereas invasive cells were fixed with 95% methanol and stained with 0.5% crystal violet. Stained cells were observed and counted under an inverted microscope (200× magnification; IX83; Olympus Corporation, Tokyo, Japan) in five randomly chosen fields in each chamber. The experimental procedure of the invasion assay was similar to that of the migration assay except that the Transwell chambers were precoated with Matrigel (BD Biosciences) before examination.
In vivo tumor growth assay
All procedures involving animals were approved by the Experimental Animal Ethics Committee of Wenzhou Hospital of Integrated Traditional Chinese and Western Medicine, and were carried out in accordance with the Declaration of Helsinki and the guidelines of the Experimental Animal Ethics Committee of Wenzhou Hospital of Integrated Traditional Chinese and Western Medicine. In total, 2×106 miR-NC- or miR-641 mimic-transfected cells were subcutaneously seeded into the flanks of nude mice (Shanghai Laboratory Animal Center; Chinese Academy of Sciences, Shanghai, People’s Republic of China). The mice were checked daily, and the tumor volume was measured for 4 days using the following formula: tumor volume =1/2 × tumor length × tumor width.2 Four weeks later, all nude mice were sacrificed under deep anesthesia. The formed tumor xenografts were excised and weighed.
Luciferase reporter assay
The 3′-UTR fragments of ZEB1 containing putative wild-type (wt) and mutant (mut) miR-641-binding sites were constructed by Shanghai GenePharma Co., Ltd., inserted into the pMIR-REPORT miRNA Expression Reporter vector (Ambion; Thermo Fisher Scientific), and named pMIR-ZEB1-3′-UTR wt and pMIR-ZEB1-3′-UTR mut, respectively. Cells were plated into 24-well plates 1 night before transfection. miR-641 mimics or miR-NC were co-transfected with pMIR-ZEB1-3′-UTR wt or pMIR-ZEB1-3′-UTR mut into cells using Lipofectamine 2000 reagent according to the manufacturer’s recommendations. Transfected cells were collected after 48 hours of incubation at 37°C and then luciferase activity was measured using a dual-luciferase reporter assay system (Promega Corporation, Fitchburg, WI, USA) in accordance with the manufacturer’s instructions. Firefly luciferase activity was normalized to that of Renilla luciferase.
Western blot analysis
Total protein was extracted from cultured cells or tissue samples using RIPA Lysis Buffer (Beyotime Institute of Biotechnology, Haimen, People’s Republic of China) in the presence of a protease inhibitor cocktail (Pierce Biotechnology, Inc., Rockford, IL, USA). A bicinchoninic acid protein assay (Aidlab Biotechnologies Co., Ltd., Beijing, People’s Republic of China) was conducted to determine the concentration of total protein. Equal amounts of protein were separated by 10% SDS-PAGE, transferred to polyvinylidene fluoride membranes (Sigma-Aldrich Co., St Louis MO, USA), and then blocked at room temperature for 1 hour in 5% fat-free milk diluted in Tris-buffered saline containing 0.1% Tween-20 (TBST). The membranes were incubated overnight at 4°C with the following primary antibodies: rabbit anti-human ZEB1 primary antibody (1:1,000 dilution; cat no: ab203829) or rabbit anti-human GAPDH primary antibody (1:1,000 dilution; cat no: ab181603; both from Abcam, Cambridge, MA, USA). After washing three times with TBST, horseradish peroxidase-conjugated goat anti-rabbit secondary antibodies (1:5,000 dilution; cat no: ab205718; Abcam) were incubated with the membranes at room temperature for 2 hours. Finally, an ECL Protein Detection kit (Pierce Biotechnology, Inc.) was applied to develop the protein bands.
Statistical analysis
Data were expressed as mean ± SD from at least three independent experiments. The chi-squared test was used to assess the correlation among miR-641 expression and clinicopathological indices of cervical cancer patients. Student’s t-test was utilized to evaluate the differences between two groups, while the significance of multiple groups was determined by one-way ANOVA followed by a Tukey’s post hoc test. SPSS software (version 16.0; SPSS, Inc., Chicago, IL, USA) was applied for all statistical analysis. The association between miR-641 and ZEB1 mRNA levels was examined by Spearman’s correlation analysis. A P-value less than 0.05 was considered to indicate statistical significance.
Results
miR-641 expression is decreased in cervical cancer tissues and cell lines
To determine the expression status of miR-641 in cervical cancer, RT-qPCR was conducted to detect miR-641 expression in 51 pairs of cervical cancer and adjacent non-tumor tissues. Compared to that in adjacent non-tumor tissues, miR-641 showed low expression in cervical cancer tissues (Figure 1A, P<0.05). Additionally, the expression level of miR-641 in four cervical cancer cell lines (C-33A, HeLa, SiHa, and CaSki) and a normal human cervix epithelial cell line (Ect1/E6E7) was determined by RT-qPCR. miR-641 was downregulated in all four cervical cancer cell lines relative to that in Ect1/E6E7 (Figure 1B, P<0.05). These results suggest that downregulation of miR-641 may be closely related with the malignant progression of cervical cancer.
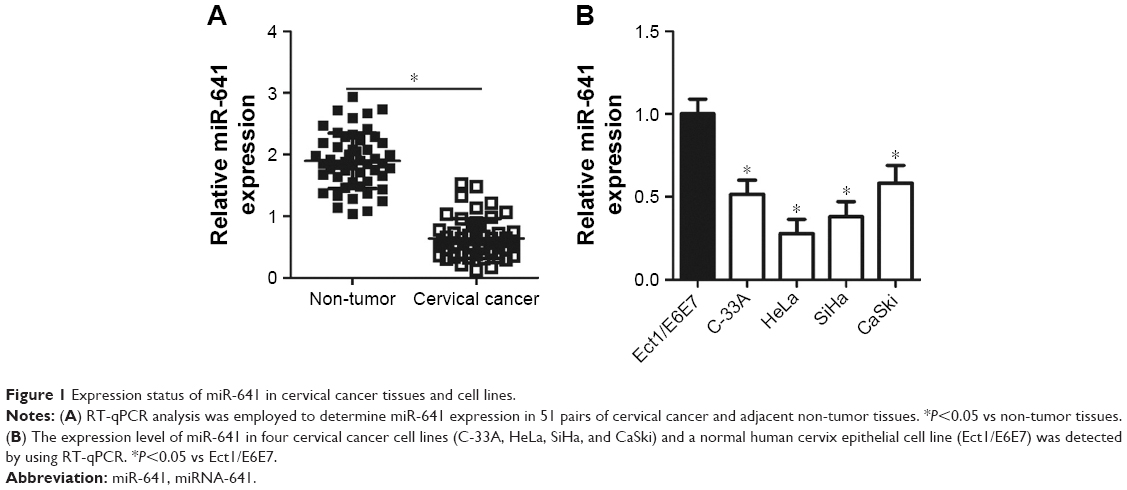 | Figure 1 Expression status of miR-641 in cervical cancer tissues and cell lines. |
Association of miR-641 expression with clinicopathological factors in cervical cancer patients
To determine the clinical significance of miR-641 in cervical cancer, all patients with cervical cancer enrolled in this study were divided into two groups: miR-641 low expression group or miR-641 high expression group. The median value of miR-641 was used as a cut-off point. Statistical analysis revealed that decreased miR-641 expression was clearly correlated with the Federation of Gynecology and Obstetrics (FIGO) stage (P=0.016) and lymph node metastasis (P=0.036), while no significant correlation with any other clinicopathological indices was observed (Table 1, all P>0.05).
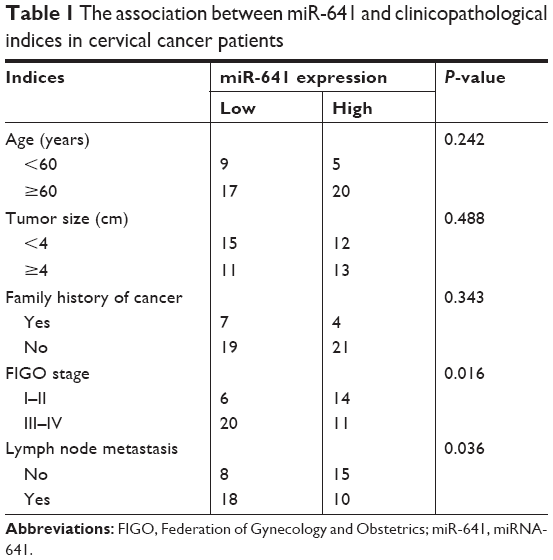 | Table 1 The association between miR-641 and clinicopathological indices in cervical cancer patients |
miR-641 inhibits cervical cancer cell proliferation, migration, and invasion, and promotes cell apoptosis in vitro
HeLa and SiHa cell lines showed relatively lower miR-641 expression among the four cervical cancer cell lines, and therefore the two cell lines were used in subsequent functional experiments. To explore the biological functions of miR-641 in cervical cancer, HeLa and SiHa cells were transfected with miR-641 mimics to increase endogenous miR-641 expression (Figure 2A, P<0.05). Subsequently, the regulatory influence of miR-641 on the proliferation of cervical cancer cells was examined. HeLa and SiHa cells transfected with miR-641 mimics exhibited obvious growth suppression compared to cells transfected with miR-NC (Figure 2B, P<0.05). The role of miR-641 in regulating cervical cancer cell apoptosis was then investigated. Flow cytometry analysis revealed that miR-641 upregulation significantly promoted the apoptosis of HeLa and SiHa cells (Figure 2C, P<0.05). Furthermore, migration and invasion assays were performed to determine the effect of miR-641 on the metastasis of cervical cancer cells. Ectopic miR-641 expression attenuated the migratory (Figure 2D, P<0.05) and invasive (Figure 2E, P<0.05) abilities of HeLa and SiHa cells. These results suggest that miR-641 plays a tumor suppressive role in the growth and metastasis of cervical cancer cells.
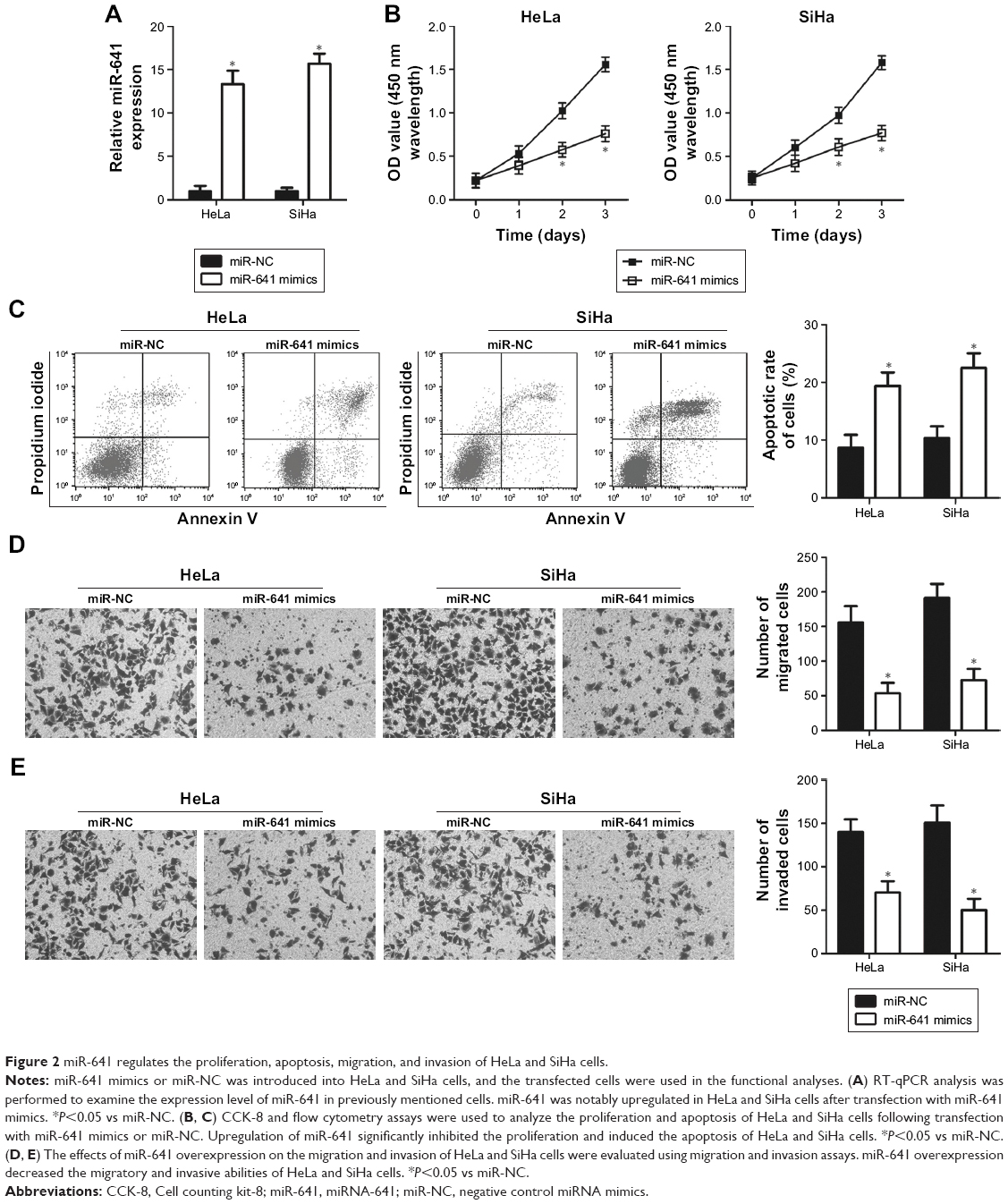 | Figure 2 miR-641 regulates the proliferation, apoptosis, migration, and invasion of HeLa and SiHa cells. |
ZEB1 is a direct target gene of miR-641 in cervical cancer cells
To clarify the molecular mechanism responsible for the tumor suppressor activity of miR-641 in cervical cancer, bioinformatics analysis was carried out to search for the putative target of miR-641. Two highly conserved putative binding sites were observed in the 3′-UTR of ZEB1 (Figure 3A). ZEB1 was selected for further verification because it is a well-known oncogene and has been implicated in the initiation and progression of cervical cancer.22–27 A luciferase reporter assay was then conducted to determine whether miR-641 directly binds to the 3′-UTR of ZEB1 in cervical cancer cells. As shown in Figure 3B, miR-641 overexpression noticeably reduced the luciferase activity of the plasmid harboring the wt (1 and 2) miR-641 binding site (P<0.05) and failed to affect that of the plasmid carrying mutated (1 and 2) ZEB1 3′-UTR in HeLa and SiHa cells (Figure 3B). Additionally, RT-qPCR and Western blot analysis revealed that restoration of miR-641 expression suppressed the mRNA (Figure 3C, P<0.05) and protein (Figure 3D, P<0.05) expression of ZEB1 in HeLa and SiHa cells. Collectively, these results demonstrate that ZEB1 is a direct target of miR-641 in cervical cancer cells.
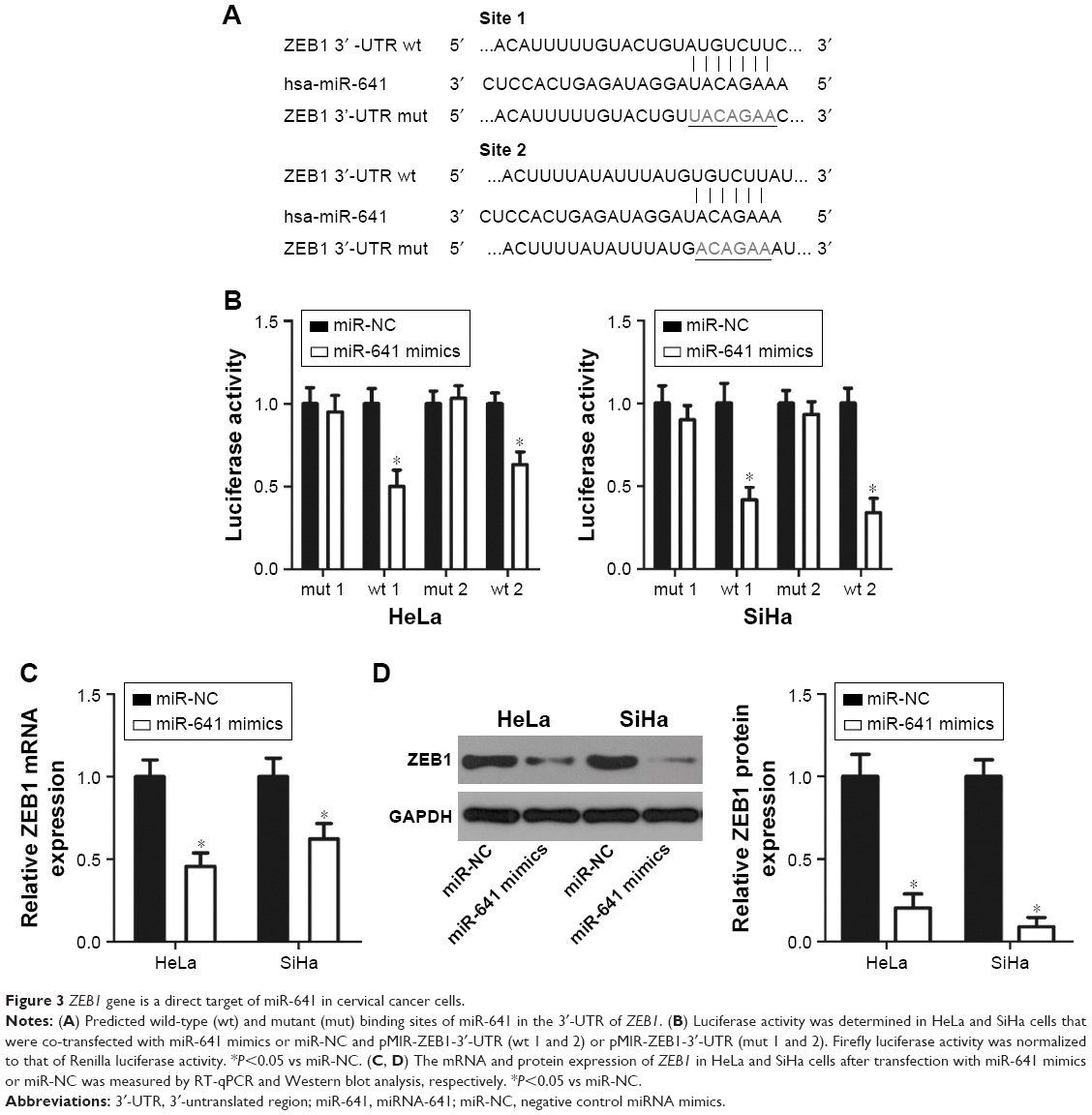 | Figure 3 ZEB1 gene is a direct target of miR-641 in cervical cancer cells. |
Upregulation of ZEB1 is negatively correlated with miR-641 expression in cervical cancer tissues
To further evaluate the relationship between miR-641 and ZEB1 in cervical cancer, the expression of ZEB1 in 51 pairs of cervical cancer and adjacent non-tumor tissues was determined by RT-qPCR. The expression level of ZEB1 mRNA was higher in cervical cancer tissues than in adjacent non-tumor tissues (Figure 4A, P<0.05). Additionally, Western blot analysis revealed that ZEB1 protein level was upregulated in cervical cancer tissues compared to that in adjacent non-tumor tissues (Figure 4B, P<0.05). Furthermore, an inverse correlation was found between miR-641 and ZEB1 mRNA levels in cervical cancer tissues (Figure 4C; r=−0.5231, P<0.0001). These results suggest that upregulation of ZEB1 expression in cervical cancer tissues is, at least partly, caused by low miR-641 expression.
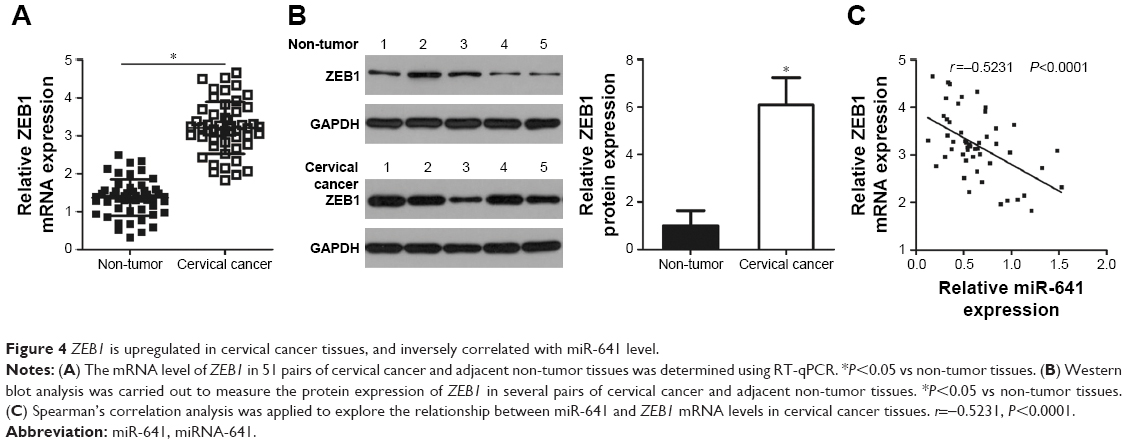 | Figure 4 ZEB1 is upregulated in cervical cancer tissues, and inversely correlated with miR-641 level. |
Knockdown of ZEB1 simulates the tumor suppressing effects of miR-641 overexpression in cervical cancer cells
Because ZEB1 was identified as a direct target gene of miR-641 in cervical cancer, we then investigated the functional roles of ZEB1 in cervical cancer.
HeLa and SiHa cells were transfected with ZEB1 siRNA to knock down endogenous ZEB1 levels. ZEB1 siRNA efficiently decreased ZEB1 protein expression in HeLa and SiHa cells compared to cells transfected with NC siRNA (Figure 5A, P<0.05). The CCK-8 assay revealed that downregulation of ZEB1 led to a decreased proliferative ability of HeLa and SiHa cells (Figure 5B, P<0.05). Flow cytometry analysis indicated that inhibition of ZEB1 increased the apoptosis rate of HeLa and SiHa cells compared to the NC siRNA groups (Figure 5C, P<0.05). Furthermore, the number of migrated (Figure 5D, P<0.05) and invaded (Figure 5E, P<0.05) cells was significantly reduced in ZEB1 siRNA transfectants compared to that in NC siRNA-transfected HeLa and SiHa cells. These results demonstrate that the effects of ZEB1 knockdown on cervical cancer cells were similar to that of miR-641 overexpression, further suggesting that ZEB1 is a functional downstream target of miR-641 in cervical cancer cells.
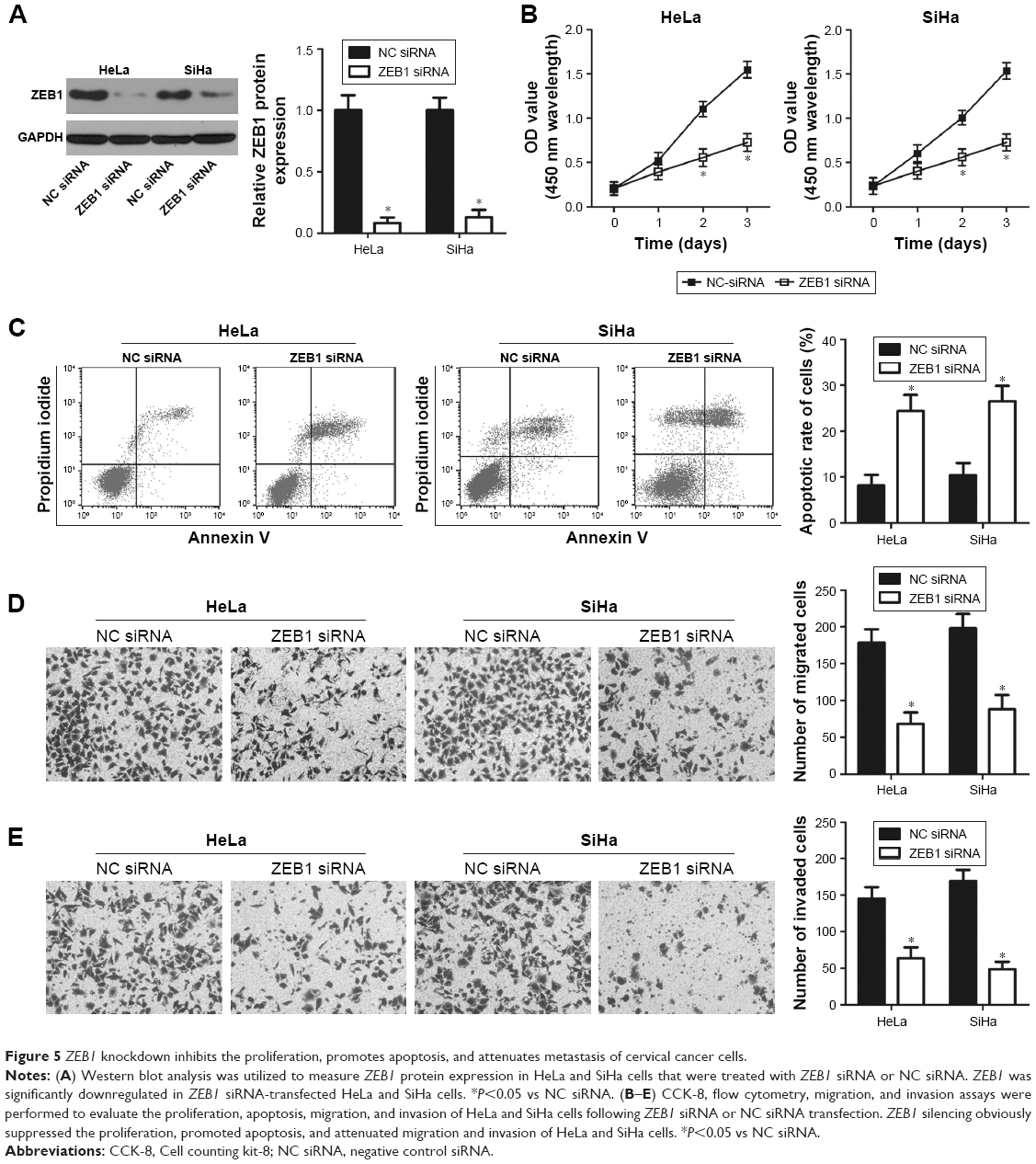 | Figure 5 ZEB1 knockdown inhibits the proliferation, promotes apoptosis, and attenuates metastasis of cervical cancer cells. |
Restored ZEB1 expression attenuates the suppressive effects of miR-641 overexpression in malignant phenotypes of cervical cancer cells
A series of rescue experiments were performed to confirm that ZEB1 mediates the tumor suppressive action of miR-641 in cervical cancer cells. Thus, miR-641-overexpressing HeLa and SiHa cells were transfected with ZEB1 overexpression plasmid (pc-ZEB1) or pcDNA3.1 blank vector. After transfection, miR-641 overexpression significantly reduced the protein level of ZEB1 in HeLa and SiHa cells; however, the ZEB1 protein expression was recovered by co-transfection with pc-ZEB1 (Figure 6A, P<0.05). Functional analyses revealed that restoration of ZEB1 expression rescued the effects of miR-641 overexpression on the proliferation (Figure 6B, P<0.05), apoptosis (Figure 6C, P<0.05), migration (Figure 6D, P<0.05), and invasion (Figure 6E, P<0.05) of HeLa and SiHa cells. These data confirm that the tumor suppressor activity of miR-641 in the malignant phenotypes of cervical cancer cells is, at least in part, attributable to downregulation of ZEB1.
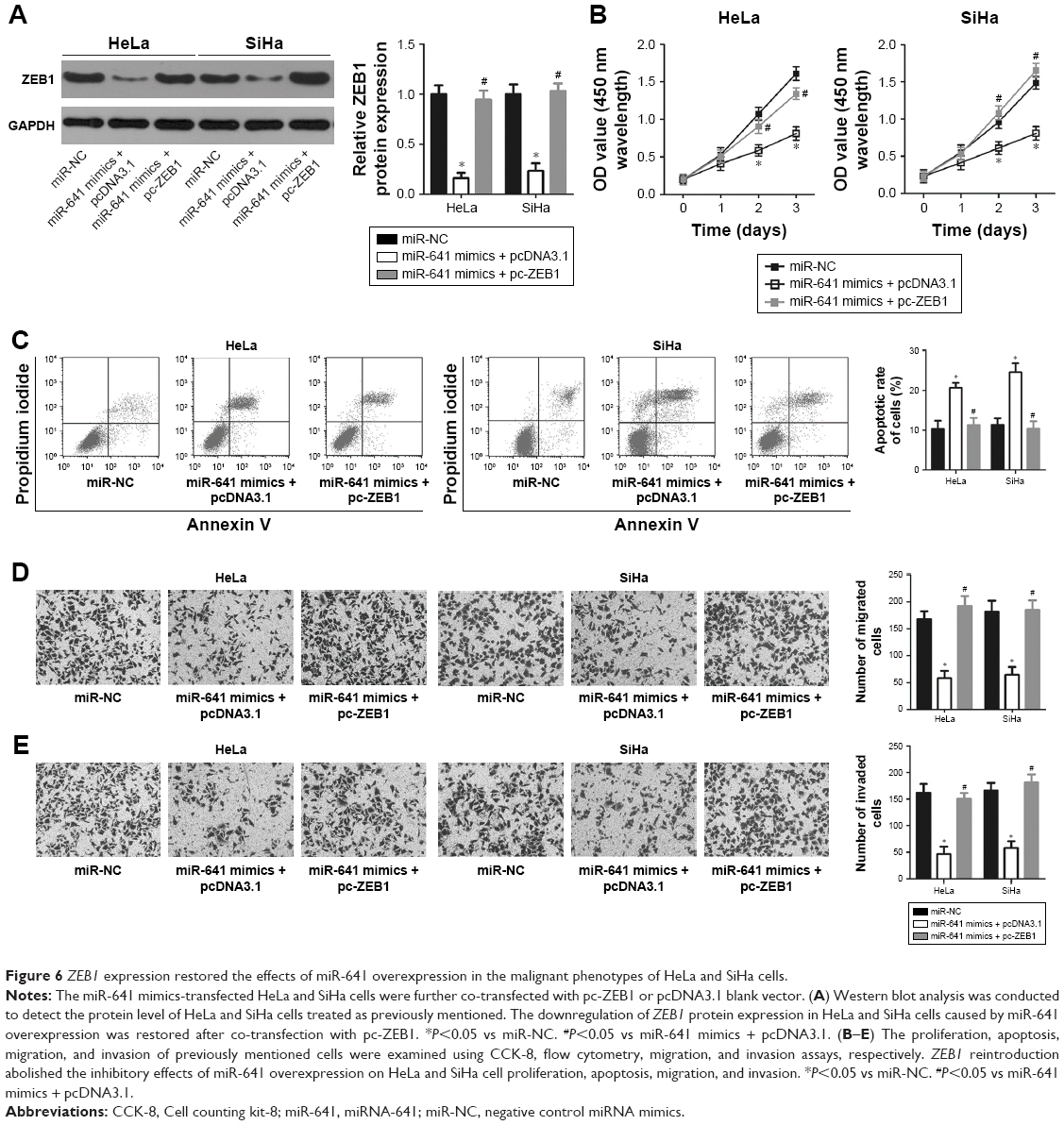 | Figure 6 ZEB1 expression restored the effects of miR-641 overexpression in the malignant phenotypes of HeLa and SiHa cells. |
miR-641 inhibits cervical cancer tumor growth in vivo
To determine the effect of miR-641 on tumor growth in vivo, in vivo tumor growth assay was carried out by subcutaneous implantation of miR-641 mimics- or miR-NC-transfected HeLa cells in nude mice. The miR-641-overexpressing tumor xenografts had an obvious reduction of tumor volume relative to that in miR-NC groups (Figure 7A and B, P<0.05). On day 29, all nude mice were sacrificed, and the tumor xenografts were excised and weighed. It was observed that the weight of tumor xenografts in miR-641 group was significantly lower than that of miR-NC group (Figure 7C, P<0.05). Additionally, RT-qPCR analysis revealed that the expression level of miR-641 in tumor xenografts from miR-641 mimic-transfected HeLa cells was markedly overexpressed (Figure 7D, P<0.05). Furthermore, Western blot analysis was applied to detect ZEB1 protein expression, and confirmed that ZEB1 protein expression was noticeably reduced in the tumor xenografts derived from the HeLa cells transfected with miR-641 mimics (Figure 7E). Together, these results suggest that miR-641 can hinder cervical cancer tumor growth in vivo by inhibiting ZEB1.
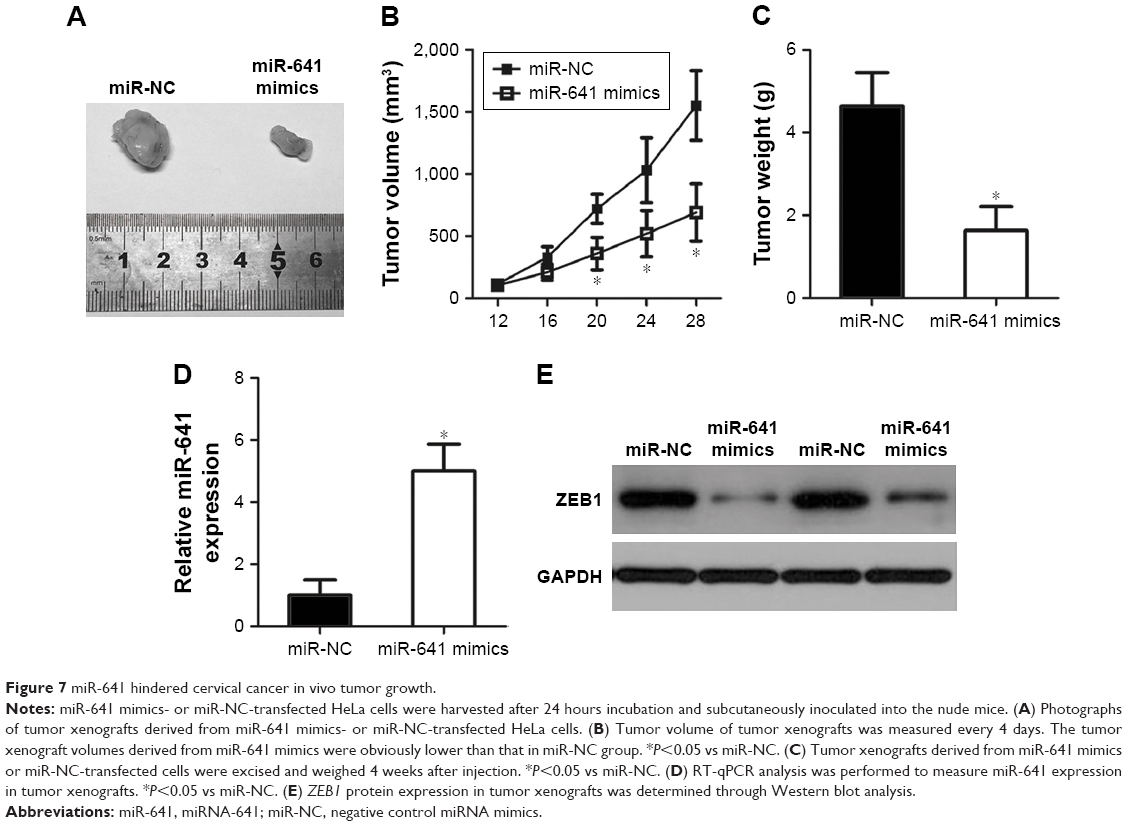 | Figure 7 miR-641 hindered cervical cancer in vivo tumor growth. |
Discussion
miRNAs were found to be dysregulated in cervical cancer, and their dysregulation has been implicated in cervical carcinogenesis and progression by regulating major tumor-related biological behaviors.28,29 Notably, exploring the underlying mechanisms of cervical cancer formation and progression may be helpful for early diagnosis and effective treatment.30 Therefore, further studies on the specific roles of deregulated miRNAs in cervical cancer and underlying molecular mechanisms may facilitate the identification of novel therapeutic techniques for patients with this disease. In this study, we detected miR-641 expression in cervical cancer tissues and determined its clinical significance in this disease. Additionally, the functional roles and underlying mechanisms of miR-641 in cervical cancer were investigated. Our results indicate that miR-641 can be used as a diagnostic biomarker and therapeutic agent in patients with cervical cancer.
miR-641 has been widely studied in lung cancer. For example, miR-641 expression is decreased in lung cancer tissues and cell lines. Ectopic miR-641 expression suppressed the proliferation and induced the apoptosis of lung cancer cells by directly targeting MDM2.19 Additionally, the expression level of miR-641 was increased in the serum of patients with non-small-cell lung cancer showing acquired resistance to erlotinib treatment. miR-641 induces erlotinib resistance by directly targeting NF1 and regulating ERK signaling in non-small-cell lung cancer.20 However, the expression status and specific roles of miR-641 in cervical cancer remain largely unknown. Here, we found that miR-641 was downregulated in cervical cancer, and the decreased miR-641 expression was significantly correlated with FIGO stage and lymph node metastasis. Functional analyses demonstrated that miR-641 overexpression restricted cervical cancer cell proliferation, promoted apoptosis, and attenuated migration and invasion in vitro as well as hindered tumor growth in vivo. Hence, this miRNA may be developed as a diagnostic biomarker and valuable therapeutic agent for patients with lung and cervical cancers.
Validation of the direct targets of miR-641 is important for understanding its roles in the progression of cervical cancer and may be helpful for identifying promising therapeutic approaches. Therefore, we investigated the molecular mechanisms responsible for the tumor suppressive actions of miR-641 in cervical cancer. First, bioinformatics analysis was conducted to search for potential targets of miR-641. One highly conserved putative binding site was observed at the 3′-UTR of ZEB1. Second, a luciferase reporter assay, RT-qPCR, and Western blot analysis revealed that miR-641 directly binds to the 3′-UTR of ZEB1 and inhibits endogenous ZEB1 expression in cervical cancer cell lines. Third, ZEB1 was upregulated in cervical cancer tissues, which is negatively correlated with miR-641 expression. Fourth, inhibition of ZEB1 mimicked the tumor suppressing roles of miR-641 overexpression in cervical cancer cell lines. Finally, rescue experiments confirmed that the downregulation of ZEB1 was essential for miR-641 to be effective in the malignant phenotypes of cervical cancer cells. These results clearly demonstrate that ZEB1 is a direct and functional downstream target of miR-641 in cervical cancer.
ZEB1 is a member of the deltaEF1 family of two-handed zinc-finger factors.31 ZEB1 was reported to be overexpressed in a variety of human malignant tumors, such as thyroid cancer,32 lung cancer,33 colorectal cancer,34 and endometrial cancer.35 Its expression is high in cervical cancer tissues and cell lines. Increased expression of ZEB1 is strongly correlated with the differentiation status, FIGO stages, lymph node metastasis, and occurrence of vascular invasion in cervical cancer patients.22,23 Emerging studies have shown that ZEB1 is closely related to the carcinogenesis and progression of cervical cancer by affecting cell proliferation, migration, invasion, epithelial–mesenchymal transition, and motility.24–27 Here, we revealed that ZEB1 is upregulated in cervical cancer and serves oncogenic roles in the genesis and development of cervical cancer. miR-641 directly targeted ZEB1 to inhibit the malignant phenotypes of cervical cancer in vitro and in vivo. Thus, ZEB1 knockdown using miR-641-based targeted therapy may be a suitable therapeutic strategy for the prevention and treatment of patients with cervical cancer.
In conclusion, we showed that miR-641 expression was decreased in cervical cancer and significantly associated with FIGO stage and lymph node metastasis. Recovery of miR-641 expression suppressed the development and progression of cervical cancer both in vitro and in vivo. Mechanistically, ZEB1 was validated as a direct target of miR-641 in cervical cancer cells. These results highlight that ZEB1 is a potential therapeutic target in cervical cancer. However, in this study, we did not explore whether miR-641 has an impact on cervical cancer epithelial–mesenchymal transition, cancer stem cell properties in vitro, and metastasis in vivo. These limitations of the present study may be resolved in future investigations.
Acknowledgment
This study was supported by the Natural Science Foundation of Zhejiang Province of China (2013C33098).
Disclosure
The authors report no conflicts of interest in this work.
References
Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. | ||
Thapa N, Maharjan M, Xiong Y, et al. Impact of cervical cancer on quality of life of women in Hubei, China. Sci Rep. 2018;8(1):11993. | ||
Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. | ||
Hildesheim A, Wang SS. Host and viral genetics and risk of cervical cancer: a review. Virus Res. 2002;89(2):229–240. | ||
Smith RA, Brooks D, Cokkinides V, Saslow D, Brawley OW. Cancer screening in the United States, 2013: a review of current American Cancer Society guidelines, current issues in cancer screening, and new guidance on cervical cancer screening and lung cancer screening. CA Cancer J Clin. 2013;63(2):88–105. | ||
Wright JD, Huang Y, Ananth CV, et al. Influence of treatment center and hospital volume on survival for locally advanced cervical cancer. Gynecol Oncol. 2015;139(3):506–512. | ||
Waggoner SE. Cervical cancer. Lancet. 2003;361(9376):2217–2225. | ||
Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. | ||
He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5(7):522–531. | ||
Kozomara A, Griffiths-Jones S. miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014;42(Database issue):D68–D73. | ||
Jiang W, Wei K, Pan C, et al. MicroRNA-1258 suppresses tumour progression via GRB2/Ras/Erk pathway in non-small-cell lung cancer. Cell Prolif. 2018;e12502. | ||
Gao X, Xu W, Lu T, Zhou J, Ge X, Hua D. MicroRNA-142-3p Promotes Cellular Invasion of Colorectal Cancer Cells by Activation of RAC1. Technol Cancer Res Treat. 2018;17:153303381879050. | ||
Li C, Zhang J, Ma Z, Zhang F, Yu W. miR-19b serves as a prognostic biomarker of breast cancer and promotes tumor progression through PI3K/AKT signaling pathway. Onco Targets Ther. 2018;11:4087–4095. | ||
Chen X, Jia C, Jia C, Jin X, Gu X. MicroRNA-374a Inhibits Aggressive Tumor Biological Behavior in Bladder Carcinoma by Suppressing Wnt/β-Catenin Signaling. Cell Physiol Biochem. 2018;48(2):815–826. | ||
Shu L, Zhang Z, Cai Y. MicroRNA-204 inhibits cell migration and invasion in human cervical cancer by regulating transcription factor 12. Oncol Lett. 2018;15(1):161–166. | ||
Laengsri V, Kerdpin U, Plabplueng C, Treeratanapiboon L, Nuchnoi P. Cervical Cancer Markers: Epigenetics and microRNAs. Lab Med. 2018;49(2):97–111. | ||
Satapathy S, Batra J, Jeet V, Thompson EW, Punyadeera C. MicroRNAs in HPV associated cancers: small players with big consequences. Expert Rev Mol Diagn. 2017;17(7):711–722. | ||
González-Quintana V, Palma-Berré L, Campos-Parra AD, et al. MicroRNAs are involved in cervical cancer development, progression, clinical outcome and improvement treatment response (Review). Oncol Rep. 2016;35(1):3–12. | ||
Kong Q, Shu N, Li J, Xu N. miR-641 Functions as a Tumor Suppressor by Targeting MDM2 in Human Lung Cancer. Oncol Res. 2018;26(5):735–741. | ||
Chen J, Cui JD, Guo XT, Cao X, Li Q. Increased expression of miR-641 contributes to erlotinib resistance in non-small-cell lung cancer cells by targeting NF1. Cancer Med. 2018;7(4):1394–1403. | ||
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402–408. | ||
Ma Y, Zheng X, Zhou J, Zhang Y, Chen K. ZEB1 promotes the progression and metastasis of cervical squamous cell carcinoma via the promotion of epithelial-mesenchymal transition. Int J Clin Exp Pathol. 2015;8(9):11258–11267. | ||
Chen Z, Li S, Huang K, et al. The nuclear protein expression levels of SNAI1 and ZEB1 are involved in the progression and lymph node metastasis of cervical cancer via the epithelial-mesenchymal transition pathway. Hum Pathol. 2013;44(10):2097–2105. | ||
Ran J, Lin DL, Wu RF, et al. ZEB1 promotes epithelial-mesenchymal transition in cervical cancer metastasis. Fertil Steril. 2015;103(6):1606–1614. | ||
Zhu H, Zeng Y, Zhou CC, Ye W. SNHG16/miR-216-5p/ZEB1 signal pathway contributes to the tumorigenesis of cervical cancer cells. Arch Biochem Biophys. 2018;637:1–8. | ||
Hu Y, Xie H, Liu Y, Liu W, Liu M, Tang H. miR-484 suppresses proliferation and epithelial-mesenchymal transition by targeting ZEB1 and SMAD2 in cervical cancer cells. Cancer Cell Int. 2017;17:36. | ||
Wang Y, Dong X, Hu B, Wang XJ, Wang Q, Wang WL. The effects of Micro-429 on inhibition of cervical cancer cells through targeting ZEB1 and CRKL. Biomed Pharmacother. 2016;80:311–321. | ||
Wang F, Li B, Xie X. The roles and clinical significance of microRNAs in cervical cancer. Histol Histopathol. 2016;31(2):131–139. | ||
Srivastava SK, Ahmad A, Zubair H, et al. MicroRNAs in gynecological cancers: Small molecules with big implications. Cancer Lett. 2017;407:123–138. | ||
Díaz-González SM, Deas J, Benítez-Boijseauneau O, et al. Utility of microRNAs and siRNAs in cervical carcinogenesis. Biomed Res Int. 2015;2015:374924. | ||
Shen A, Zhang Y, Yang H, Xu R, Huang G. Overexpression of ZEB1 relates to metastasis and invasion in osteosarcoma. J Surg Oncol. 2012;105(8):830–834. | ||
Zhang Y, Liu G, Wu S, Jiang F, Xie J, Wang Y. Zinc finger E-box-binding homeobox 1: its clinical significance and functional role in human thyroid cancer. Onco Targets Ther. 2016;9:1303–1310. | ||
Larsen JE, Nathan V, Osborne JK, et al. ZEB1 drives epithelial-to-mesenchymal transition in lung cancer. J Clin Invest. 2016;126(9):3219–3235. | ||
Zhang GJ, Zhou T, Tian HP, Liu ZL, Xia SS. High expression of ZEB1 correlates with liver metastasis and poor prognosis in colorectal cancer. Oncol Lett. 2013;5(2):564–568. | ||
Singh M, Spoelstra NS, Jean A, et al. ZEB1 expression in type I vs type II endometrial cancers: a marker of aggressive disease. Mod Pathol. 2008;21(7):912–923. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.