")
Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 15
miR-17-5p Promotes Glucose Uptake of HTR8/SVneo Trophoblast Cells by Inhibiting TXNIP/NLRP3 Inflammasome Pathway
Authors Jiang Y , Wei L, Zhang H, Chen Y, Gao P, Zhang J, Zhou X, Zhu S, Du Y, Fang C, Li J, Feng L, He M, Wang S, Yu J
Received 10 August 2022
Accepted for publication 20 October 2022
Published 31 October 2022 Volume 2022:15 Pages 3361—3374
DOI https://doi.org/10.2147/DMSO.S385774
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Antonio Brunetti
Yi Jiang,1 Lijie Wei,1 Huiting Zhang,1 Yuting Chen,1 Peng Gao,1 Jingyi Zhang,1 Xuan Zhou,1 Shenglan Zhu,1 Yuanyuan Du,1 Chenyun Fang,1 Jiaqi Li,2 Ling Feng,1 Mengzhou He,1 Shaoshuai Wang,1 Jun Yu1
1Department of Obstetrics and Gynecology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, 430030, People’s Republic of China; 2Department of Obstetrics and Gynecology, The First Affiliated Hospital of Nanchang University, Nanchang, Jiangxi, 33006, People’s Republic of China
Correspondence: Jun Yu, Department of Obstetrics and Gynecology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, No. 1095 Jiefang Avenue, Qiaokou District, Wuhan, Hubei Province, People’s Republic of China, Email [email protected]
Introduction: Gestational diabetes mellitus (GDM) is one of the common metabolic disorders of pregnancy and results in poor pregnancy outcomes for both mother and fetus. MiR-17-5p is considered as the strongest predictor of metabolic syndrome status, but the relationship between GDM and miR-17-5p remains unclear. TXNIP, which leads to activation of NLRP3, is considered as a potential target of miR-17-5p, and the miR-17-5p/TXNIP/NLRP3 axis has been shown to play a major role in the occurrence and development of many metabolic diseases but has not been validated in GDM.
Methods: MiR-17-5p was detected by RT-qPCR. The expression of TXNIP and NLRP3 in placenta was detected by immunofluorescence, RT-qPCR and Western blot. To explore the effect of miR-17-5p on TXNIP and NLRP3 and glucose uptake of HTR8/SVneo cells, miR-17-5p mimic and miR-17-5p inhibitor were transfected to achieve overexpression and inhibition. The interaction between miR-17-5p and TXNIP was confirmed by dual-luciferase reporter assay. Besides, glucose consumption of trophoblast cells was detected by glucose assay kit.
Results: MiR-17-5p expression was down-regulated, while the expression of TXNIP and NLRP3 was up-regulated in GDM placental tissues. MiR-17-5p targeted TXNIP and inhibited its expression. MiR-17-5p also regulated NLRP3 expression and glucose uptake of HTR8/SVneo cells, which could be reversed by overexpression of TXNIP, suggesting that miR-17-5p improved glucose uptake of HTR8/SVneo cells by TXNIP/NLRP3 axis. The results were consistent with the above findings in high-glucose treated HTR8/SVneo cells.
Conclusion: Our results suggested that miR-17-5p ameliorates the glucose uptake of HTR8/SVneo cells by TXNIP/NLRP3 axis, which may provide a new idea for offspring health of GDM patients.
Keywords: diabetes, gestational, microRNAs, pregnancy, inflammation
Introduction
Gestational diabetes mellitus (GDM), which is defined as a state of glucose intolerance first detected during pregnancy, is one of the common metabolic disorders of pregnancy and results in poor pregnancy outcomes for both mother and fetus. According to the 15 centers involved in the Hyperglycemia and Adverse Pregnancy Outcomes (HAPO) study, the worldwide prevalence of GDM ranged from 9% to 26%, with an average of 18%.1 Lots of complications are associated with GDM, like offspring complications (macrosomia, preterm birth, neonatal hypoglycemia, and so on) and maternal complications (gestational hypertension, preeclampsia, and so on).2–7 Although the molecular mechanisms of GDM are still not clear, tissue insulin resistance is considered as a critical part of the pathophysiology of GDM,8 and the impaired glucose uptake of GDM is associated with insulin resistance. Compared with normal pregnancy, the rate of glucose uptake of GDM decreased by 54%,9 which results in a high-glucose environment in utero. The fetus exposed to high levels of intrauterine glucose may develop a variety of diseases, including preterm birth, miscarriage, macrosomia, intrauterine growth retardation, obesity and type 2 diabetes mellitus later in life.10 Therefore, ameliorating glucose uptake and alleviating insulin resistance of GDM play a key role in the treatment of GDM.
MicroRNA (miRNA) is a type of non-coding RNA, which is small (20–23nt long), highly conserved and endogenous. Via specifically binding to the 3’ untranslated region (3’-UTR), miRNA induces target gene mRNA translational inhibition or degradation,11 which has been shown to be closely related to the development of some diseases, including GDM. The role of some miRNAs in GDM has been studied. For example, miR-98 directly regulated Mecp2/Trpc3 axis, which was considered as a major signal of insulin-mediated glucose uptake.12 Besides, miR-96 was significantly down-regulated in GDM, resulting in decreased insulin concentration and cell viability.13 MiR-17-5p, considered as the strongest predictor of metabolic syndrome status (miR-17-5p displayed down-regulated expression in metabolic syndrome subjects),14 is a key contributor to the molecular mechanisms that regulate glucose-stimulated insulin secretion and pancreatic beta-cell adaptation under metabolic stress.15 However, the relationship between miR-17-5p and GDM remains unclear.
The Nod-like receptor protein 3 (NLRP3) inflammasome is a member of Nod-like receptors (NLRs), and lots of immune and metabolic disorders have been shown to be closely associated with its activation. In GDM pregnant women, previous study has proved the relationship between the NLRP3 inflammasome activation and insulin resistance,16 and in the placental tissues of GDM mice, the NLRP3 expression was elevated,17 which might be caused by high glucose levels and in turn lead to insulin resistance.18 Another study suggested that PLIN2 could activate NLRP3 inflammasome and via decreasing IRS-1 expression to inhibit the insulin-induced glucose uptake.19 Therefore, inhibiting activation of inflammasomes could be a feasible way to alleviate insulin resistance of GDM and ameliorate glucose uptake.
Thioredoxin-interacting protein (TXNIP), considered as an intracellular amplifier of oxidative stress and inflammasome activation, may play a role in insulin resistance.20 Prior researches have confirmed that NLRP3 interacted with TXNIP, and TXNIP deficiency led to impaired activation of NLRP3 inflammasome and affected subsequent IL-1β secretion.21 TXNIP was also found to be overexpressed in the placenta of GDM.22 Besides, TXNIP is thought to be a target of miR-17-5p,23 and the binding sites of miR-17-5p with TXNIP 3’UTR were predicted by Targetscan. The direct interaction of TXNIP with miR-17-5p in HEK293 cells and C17.2 mouse neural stem cells has been confirmed by dual luciferase reporter assays in previous studies.23,24
Our study focused on the relationship among miR-17-5p, TXNIP and NLRP3 in HTR8/SVneo cells and their effect on glucose uptake, which may provide a novel target in alleviating insulin resistance and improving prognosis of GDM offspring.
Materials and Methods
Participants and Samples
Pregnant women diagnosed at Tongji Hospital were recruited in this study. A total of 32 women who had routine prenatal examinations were included, in which 16 women were diagnosed with GDM and another 16 women were normal pregnancies (Normal). There were no other pregnancy complications among all participants and all of them underwent 75g oral glucose tolerance test (OGTT) during 24–28 weeks of gestation. The diagnosis of GDM is based on the International Association of Diabetes and Pregnancy Study Groups (IADPSG). Written informed consent was taken from all the participants. This research was approved by the Ethics Board of Tongji Hospital.
1 cm3 placental tissues of maternal side were collected from full-term patients at the time of cesarean. After washing with cold PBS, the tissues were immediately cryopreserved in liquid nitrogen and then stored at −80°C. Other tissues from placenta were fixed in formalin and paraffin embedded for following histological analysis.
Cell Culture and Treatment
HTR-8/SVneo cell line was purchased from Servicebio Technology (Wuhan, China). HTR-8/SVneo cells were cultured in RPMI 1640 medium (Gibco, USA) supplemented with 10% fetal bovine serum (FBS) at 37°C with 5% CO2.
miR-17-5p mimic, miR-17-5p mimic NC, miR-17-5p inhibitor and miR-17-5p inhibitor NC were purchased from RiboBio (Guangzhou, China). TXNIP Vector and Vector NC were purchased from Vigene Biosciences Inc. (Jinan, China). HTR8/SVneo cells were transfected with Lipofectamine® 3000 (Thermo Fisher Scientific, USA) transfection reagents when reached 50–60% density.
Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
Total RNA was extracted from all placental tissues and HTR8/SVneo cells by TRIzol reagent (Takara, Japan). For synthesizing the cDNA of miRNA, miRNA 1st Strand cDNA Synthesis Kit (by stem-loop) (Vazyme, China) was carried out, while HiScript® II Q RT SuperMix for qPCR (Vazyme, China) was used for cDNA synthesis of mRNA. U6 was used as an endogenous control for the analysis of miR-17-5p and ACTB was used to normalize the expression of TXNIP and NLRP3. RT-qPCR was carried out using the Bio-rad CFX CONNECT Real-Time System (Bio-rad, Hercules, California, USA). Gene-relative expression levels were calculated using the 2−ΔΔCT method and normalized to the above controls (U6 and ACTB). The sequences of primers were the following:
miR-17-5p F 5’-GGCCAAAGTGCTTACAGTGC-3’, R 5’-ACTGCAGGGTCCGAGGTATT-3’;
TXNIP F 5’- CAGCAGTGCAAACAGACTTCGG-3’, R 5’- CTGAGGAAGCTCAAAGCCGAAC-3’;
NLRP3 F 5’- GCCAGGAAGACAGCATTGAAGA-3’, R 5’- CTCTCACCCAGACGGGCATT-3’;
U6 F 5’- CTCGCTTCGGCAGCACA-3’, R 5’- AACGCTTCACGAATTTGCGT-3’;
ACTB F 5’- CACCATTGGCAATGAGCGGTTC-3’, R 5’- AGGTCTTTGCGGATGTCCACGT-3’.
Western Blot
Total proteins from placental tissues and HTR-8/SVneo cells were extracted with RIPA buffer (Thermo Fisher Scientific, USA) containing inhibitors of protease and phosphatase according to the protocol. The 20μg protein was separated by SDS-PAGE and then transferred to PVDF membranes. 5% non-fat milk was used to block the membranes at room temperature for 1 hour. Primary anti-bodies against β-Actin (66009-1-Ig; 1:5000; Proteintech, China), TXNIP (ab188865; 1:1000; Abcam) and NLRP3 (ab263899; 1:1000; Abcam) were used to incubate the membranes at 4°C overnight. The next day, the membranes were incubated with the secondary antibodies (HRP-conjugated secondary antibody) at room temperature for 1 hour, and the blots were detected using electrochemiluminescence assay with detection by G:BOX Chemi XRQ (Syngene, UK).
Immunofluorescence
The formalin-fixed paraffin-embedded placenta samples were sliced into 4 um thick sections. The sections were performed immunofluorescence analysis according to the standard protocol. All sections were first incubated with anti-TXNIP antibody (A9342; 1:100; ABclonal, China) or anti-NLRP3 antibody (A5652; 1:100; ABclonal, China) in combination with anti-CK7 (Cytokeratin 7) antibody (YM3054, Immunoway, Beijing, China) at 4°C overnight. The next day, the sections were incubated with secondary antibody. The DNA dye-4’,6’-diamidino-2-phenylindole (DAPI) was used for nucleus staining. All sections were imaged through an immunofluorescence microscope (Olympus BX53, Tokyo, Japan).
Glucose Consumption
In order to explore the glucose consumption of HTR8/SVneo cells, we used glucose assay kit (Sigma-Aldrich, USA) to detect the glucose levels of culture medium. Glucose consumption was calculated using the difference value between initial and final glucose content and normalized to corresponding amounts.
Dual Luciferase Reporter Gene Assay
pMIR-REPORT Luciferase-TXNIP 3’UTR (WT) and the pMIR-REPORT Luciferase-TXNIP 3’UTR (MUT) were designed and purchased from Obio (Shanghai, China). HTR8/SVneo cells were co-transfected with TXNIP-WT, TXNIP-MUT, miR-17-5p mimic or miR-17-5p mimic NC. Cells were cultured for 48h after transfection, and then, the luciferase activities of transfected cells were detected using Dual-Luciferase Reporter Assay System (Vazyme, China).
Statistical Analysis
For all analyses, continuous variables were expressed as mean ± standard deviation (SD). ImageJ was used for detecting intensity of protein band. Statistical analyses were conducted using GraphPad Prism 9.0. Student’s t-test or non-parametric test were used to compare two independent groups, while comparisons between multiple groups were performed using one-way ANOVA test. Data of each experiment came from at least 3 independent experiments and differences were considered significant at a P value with a two-side <0.05.
Results
The miR-17-5p Expression Was Decreased While TXNIP and NLRP3 Expression Were Increased in GDM Placental Tissues
The clinical characteristics are summarized in Table 1. For 32 pregnant women included in this study, half of pregnant women (16) were diagnosed as GDM with a mean age of 32.06 ± 2.98 years, while another half were identified as Normal group with an average age of 30.06 ± 2.74 years. Without doubt, pregnant women with GDM had higher plasma glucose compared with Normal group (P < 0.05), and we also found that the pre-pregnancy BMI of the GDM group was 21.75 ± 1.76 kg/m2, which was significantly higher than the Normal group (P < 0.05).
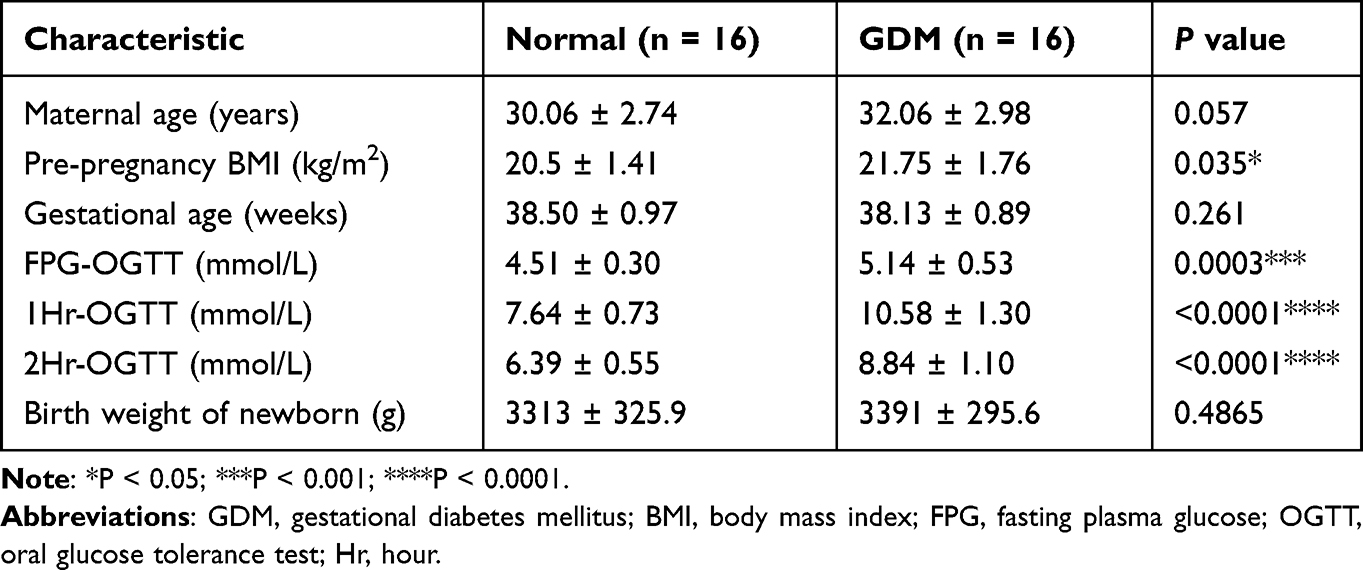 |
Table 1 Clinical Characteristics of Normal and GDM Groups |
Besides, we found that miR-17-5p expression in GDM placenta was significantly decreased compared with normal by RT-qPCR (Figure 1A). Then, we detected the expression of TXNIP and the expression of NLRP3 in the placental tissues by immunofluorescence in order to identify the expression and location of both proteins in the placenta. We observed that both TXNIP and NLRP3 staining (red) were strongly positive in CK7+ cells (green), and both TXNIP and NLRP3 expression in GDM group were increased (Figure 1B). Furthermore, the TXNIP mRNA and NLRP3 mRNA in GDM placental tissues were significantly higher than Normal group (Figure 1C and D), and Western blot showed that these two proteins’ expression in GDM group was also significantly increased than Normal group (Figure 1E–G). These results suggested that miR-17-5p, TXNIP and NLRP3 may play a role in the development of GDM.
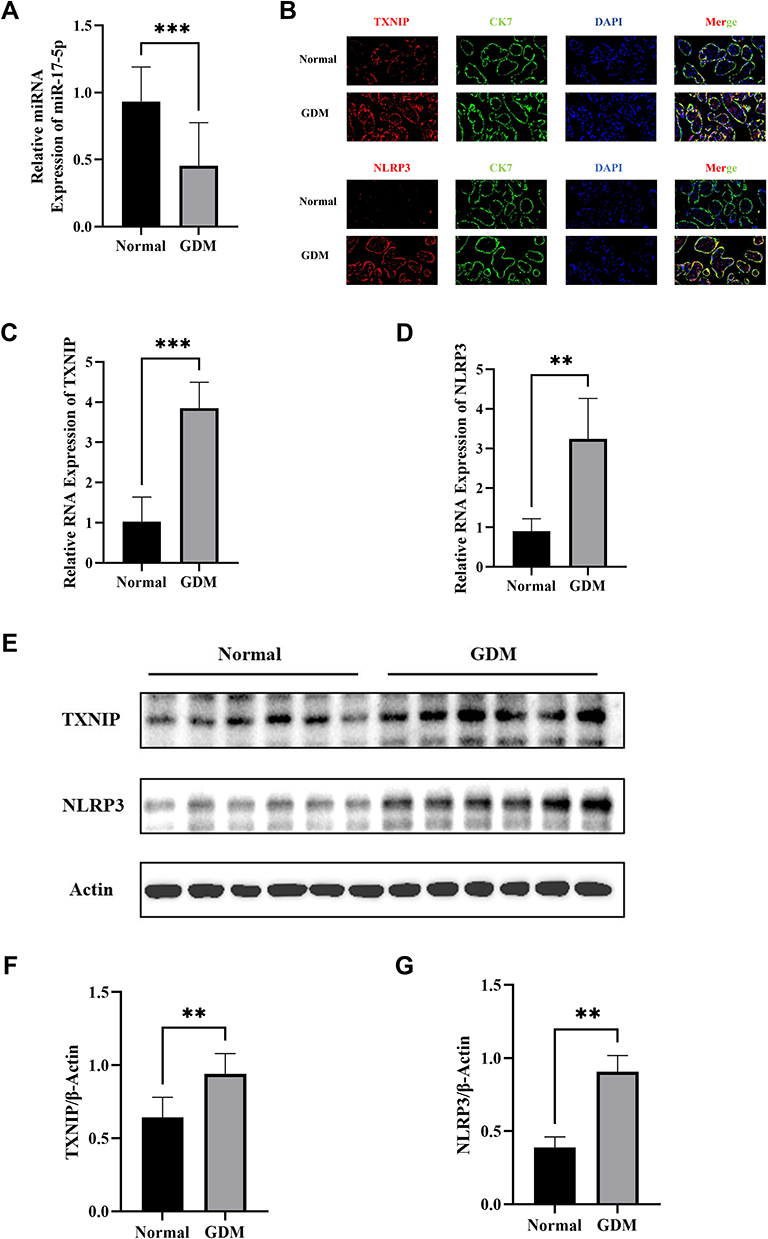 |
Figure 1 MiR-17-5p, TXNIP and NLRP3 expression in placental tissues in GDM group (n=16) and Normal group (n=16). (A) Quantitative analysis of miR-17-5p expression in placental tissues by qRT-PCR. (B) Representative images of immunofluorescence double staining of TXNIP (red) or NLRP3 (red) and CK7 (green) in placental tissues of GDM and Normal groups. (C and D) The mRNA expression of TXNIP (C) and NLRP3 (D) were detected in normal tissues and GDM tissues by qRT-PCR. (E–G) The protein expression of TXNIP (G) and NLRP3 (H) was detected in normal tissues and GDM tissues by Western blot. **P<0.01, ***P<0.001. |
MiR-17-5p Targets TXNIP Expression in HTR8/SVneo
To explore the regulatory effect of miR-17-5p on TXNIP in HTR8/SVneo cells, miR-17-5p mimic, miR-17-5p mimic NC, miR-17-5p inhibitor and miR-17-5p inhibitor NC were transfected, respectively. A promotion of expression of TXNIP mRNA and protein was detected when miR-17-5p inhibitor was transfected, while a suppression was detected with miR-17-5p mimic (Figure 2A–F). Moreover, the binding sites of miR-17-5p with TXNIP 3’UTR were found by Targetscan (Figure 2G), and luciferase reporter assay was used to explore the relationship between miR-17-5p and TXNIP directly. We constructed luciferase reporter vectors containing TXNIP wild type (TXNIP-WT) and mutant (TXNIP-MUT) sequences and co-transfected with miR-17-5p mimic or miR-17-5p mimic NC into HTR8/SVneo cells. The luciferase activity was significantly decreased by miR-17-5p mimic in TXNIP-WT group, while there was no significant difference among treatments in TXNIP-MUT group (Figure 2G). These results suggested that miR-17-5p directly targets TXNIP and negatively regulates its expression in HTR8/SVneo cells.
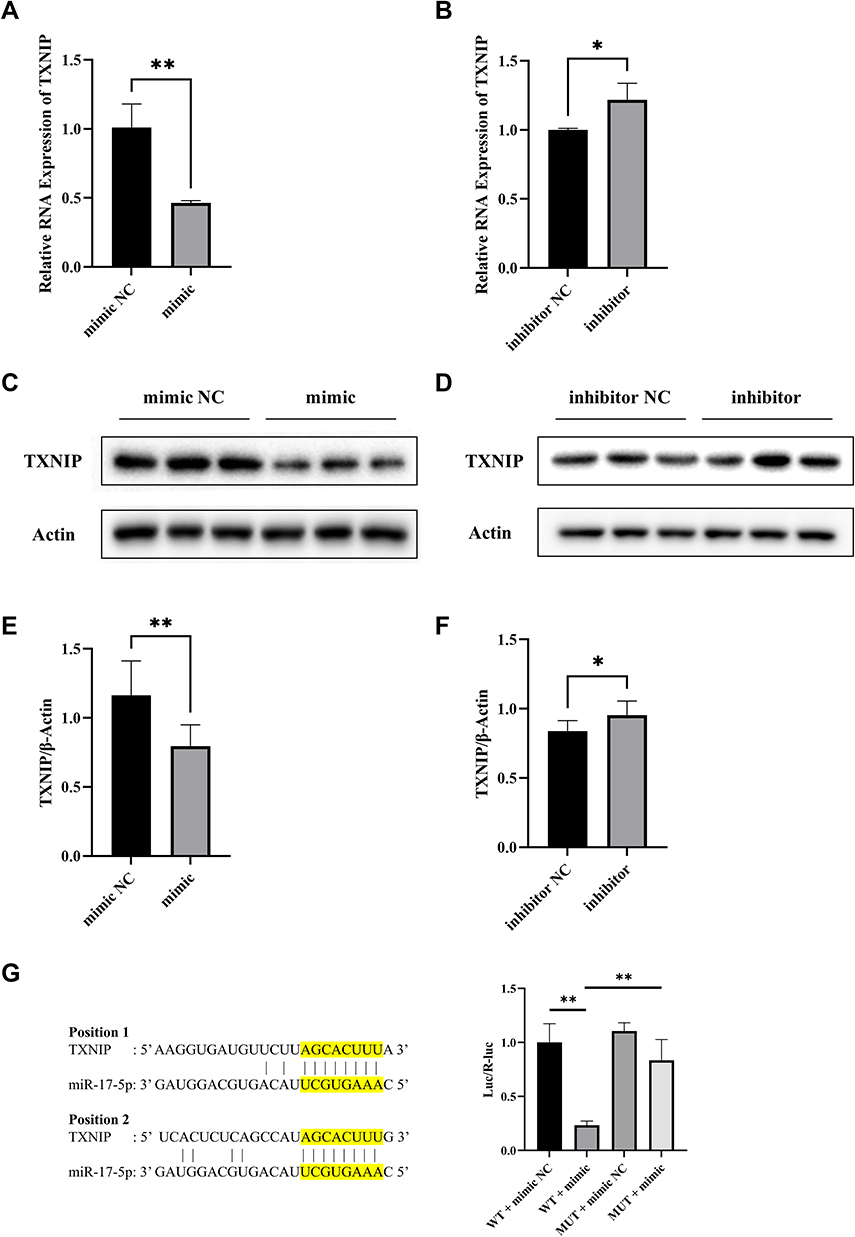 |
Figure 2 MiR-17-5p regulates TXNIP expression in HTR8/SVneo cells. (A and B) The mRNA expression of TXNIP was detected by qRT-PCR. (C–F) Representative images and quantitative analysis of TXNIP protein expression. (G) The binding sites between miR-17-5p and TXNIP 3’UTR predicted by Targetscan. TXNIP-WT or TXNIP-MUT was co-transfected with miR-17-5p mimic or miR-17-5p mimic NC into HTR8/SVneo cells. Luciferase activity was measured using luciferase reporter assay. *P<0.05, **P<0.01. |
MiR-17-5p Regulates NLRP3 Expression and Cellular Glucose Uptake in HTR8/SVneo
We found that NLRP3 mRNA expression was inhibited when miR-17-5p mimic was transfected and promoted when miR-17-5p inhibitor was transfected in HTR8/SVneo cells (Figure 3A and B). The same conclusion was obtained at the protein expression level (Figure 3C–F). Moreover, to explore the effect of miR-17-5p on glucose uptake, we measured the glucose consumption and the results showed that glucose consumption was significantly increased in miR-17-5p mimic-treated cells compared with NC group (Figure 3G), which suggested that miR-17-5p inhibits the expression of NLRP3 and promotes the glucose uptake of HTR8/SVneo cells. However, no direct binding site between miR-17-5p and NLRP3 was predicted in Targetscan, which implied that the regulatory effect of miR-17-5p on NLRP3 may be mediated by other mediators.
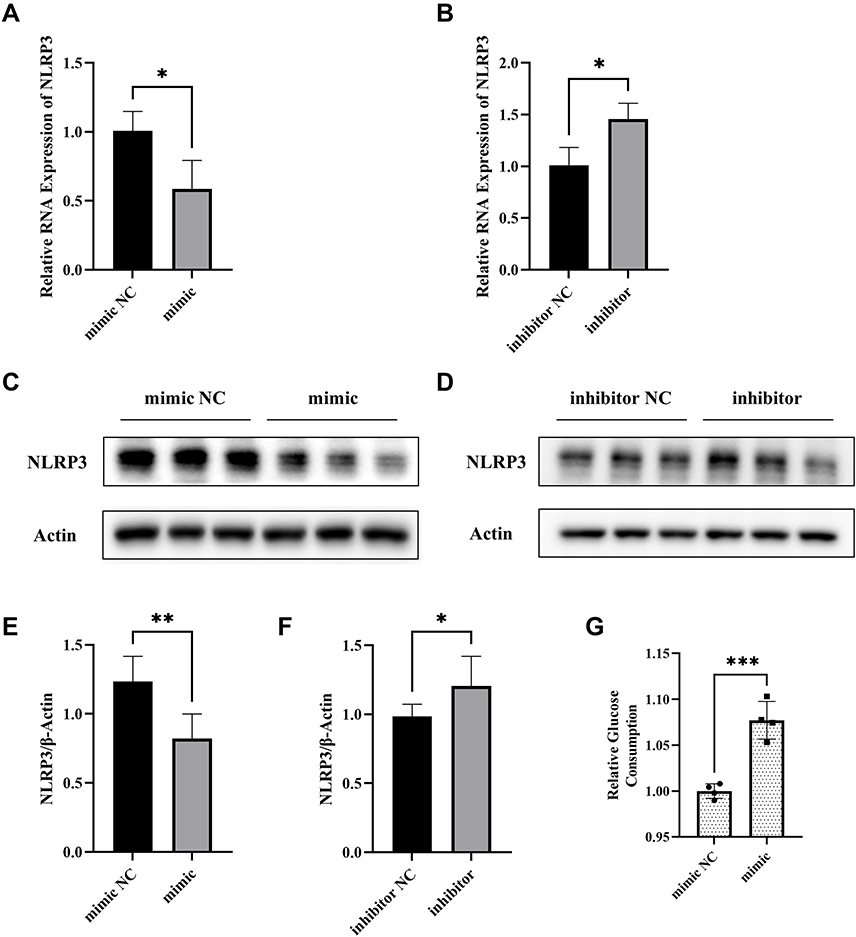 |
Figure 3 MiR-17-5p regulates NLRP3 expression and glucose uptake in HTR8/SVneo cells. (A and B) The expression of NLRP3 mRNA of transfected HTR8/SVneo cells detected by qRT-PCR. (C–F) Representative images and quantitative analysis of NLRP3 protein expression. (G) Relative quantitative analyses of glucose consumption. *P<0.05, **P<0.01, ***P<0.001. |
Effects of TXNIP on Regulation of NLRP3 Expression and Cellular Glucose Uptake in HTR8/SVneo
Next, we explored the function of TXNIP on NLRP3 expression. TXNIP Vector and Vector Blank were transfected into HTR8/SVneo, respectively. The TXNIP mRNA and protein expression were significantly increased when TXNIP Vector was transfected (Figure 4A and D). Interestingly, the NLRP3 mRNA and protein expression were also significantly increased under the same condition (Figure 4B and E). However, the relative glucose consumption of HTR8/SVneo cells was decreased (Figure 4F), which suggested that the cellular ability of glucose uptake was impaired. These results suggested a correlation in gene expression between TXNIP and NLRP3 in HTR8/SVneo. Given that we have concluded that miR-17-5p can target TXNIP and inhibit its expression, the regulatory effect of miR-17-5p on NLRP3 may be mediated by TXNIP.
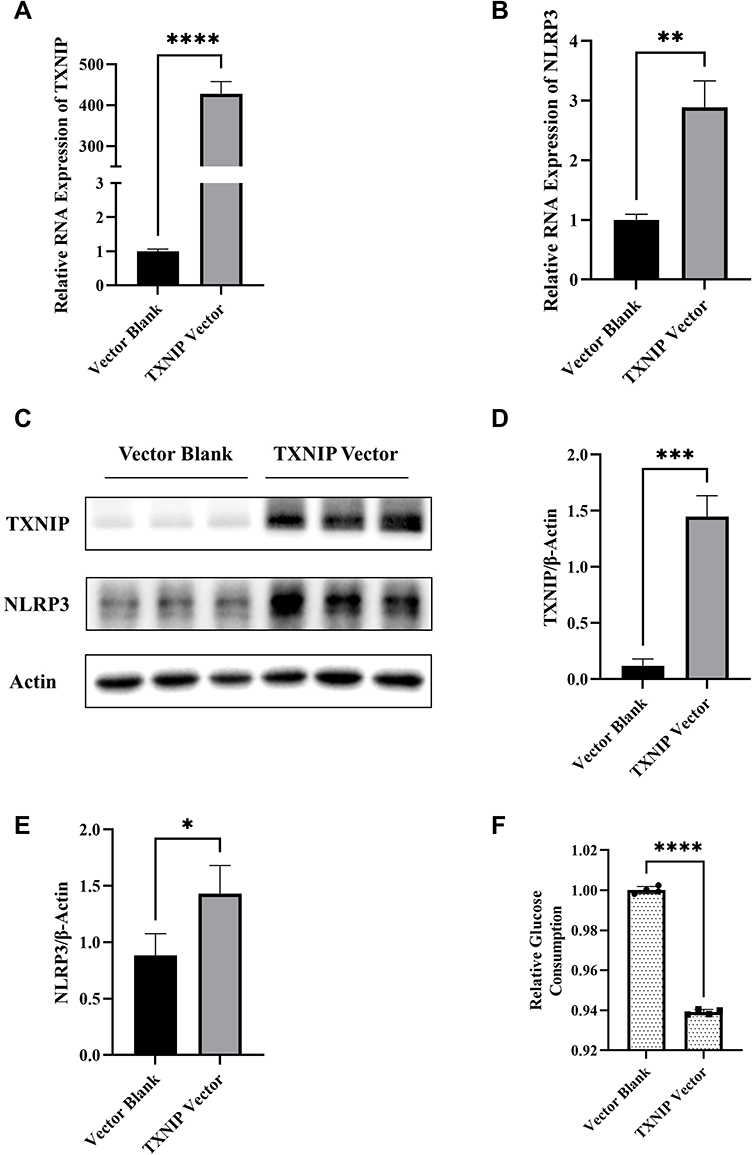 |
Figure 4 Effects of TXNIP on regulation of NLRP3 expression and cellular glucose uptake in HTR8/SVneo cells. (A and B) The mRNA expression of TXNIP and NLRP3 in HTR8/SVneo cells transfected with TXNIP Vector or Blank. (C–E) Representative images and quantitative analysis of TXNIP and NLRP3 protein expression detected by Western blot. (F) Relative quantitative analyses of glucose consumption. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. |
MiR-17-5p Inhibits NLRP3 Expression and Improves Cellular Glucose Uptake by Targeting TXNIP
In order to explore the regulatory relationship among miR-17-5p, TXNIP and NLRP3, miR-17-5p mimic, miR-17-5p mimic NC, miR-17-5p mimic + TXNIP Vector or miR-17-5p mimic + Vector Blank was transfected into HTR8/SVneo cells. The NLRP3 mRNA and protein expression were inhibited by miR-17-5p, which could be reversed by TXNIP overexpression (Figure 5A–E). Moreover, transfection of mimic alone increased glucose consumption, which was impaired by mimic and TXNIP Vector co-transfection (Figure 5F). These results suggested that miR-17-5p inhibited NLRP3 expression and improved cellular glucose uptake by targeting TXNIP in HTR8/SVneo.
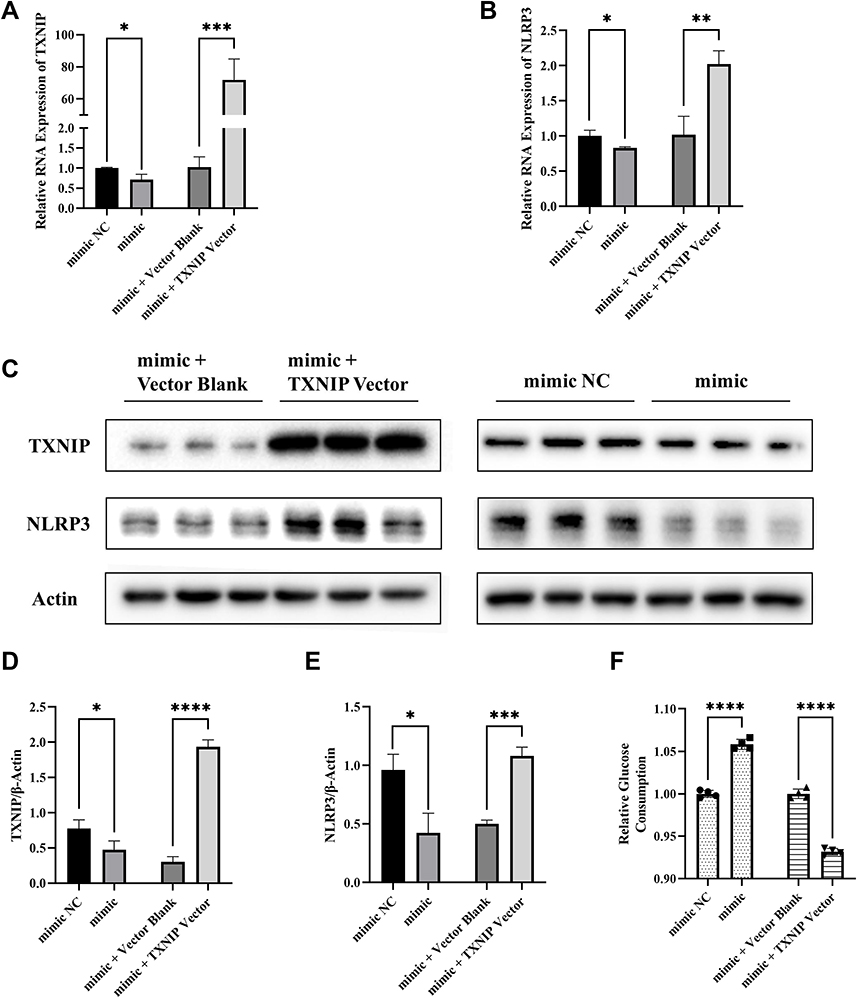 |
Figure 5 MiR-17-5p inhibits the expression of NLRP3 and improves cellular glucose uptake by targeting TXNIP. (A and B) The mRNA expression of TXNIP and NLRP3 in HTR8/SVneo cells transfected by mimic + TXNIP Vector, mimic + Vector Blank, mimic or mimic NC respectively. (C–E) Representative images and quantitative analysis of TXNIP and NLRP3 protein expression. (F) Relative quantitative analyses of glucose consumption. *P<0.05, **P<0.01, ***P<0.01, ****P<0.0001. |
Effects of miR-17-5p on TXNIP, NLRP3 Expression and Glucose Uptake in HTR8/SVneo Cells with High-Glucose
In order to simulate the high glucose environment in GDM, we used high glucose medium (25mmol/L) to treat HTR8/SVneo cells. We found that the expression of TXNIP and NLRP3 was significantly increased in high-glucose treated cells compared with the normal cells (Figure 6A–E). However, when cells were transfected by miR-17-5p mimic and stimulated with high-glucose at the same time, the expression of TXNIP and NLRP3 returned to normal (Figure 6A–E). Besides, the glucose consumption of high-glucose treated HTR8/SVneo cells was less than that of normal cells, which was reversed by miR-17-5p mimic transfection (Figure 6F).
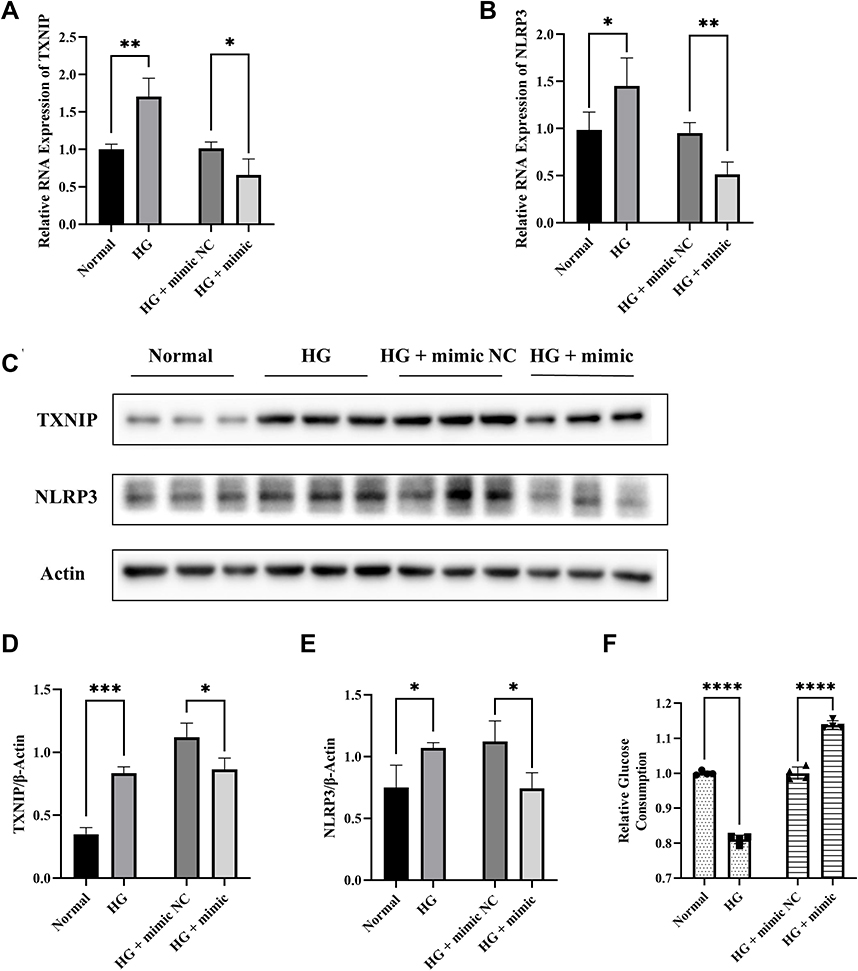 |
Figure 6 Effects of miR-17-5p on TXNIP, NLRP3 expression and glucose uptake in HTR8/SVneo cells treated with high-glucose. (A and B) The mRNA expression of TXNIP and NLRP3 in HTR8/SVneo cells. (C–E) Representative images and quantitative analysis of TXNIP and NLRP3 protein expression. (F) Relative quantitative analyses of glucose consumption. *P<0.05, **P<0.01, ***P<0.01, ****P<0.0001. |
Discussion
The causal mechanisms of GDM are complex and still not completely elucidated, but what we do know is that GDM is not a simple placenta-derived disease.25 However, placenta is still considered as an important organ because it is the hub connecting the maternity and offspring. Although insulin resistance occurs in most organs in GDM, the placental insulin resistance appears to be more closely associated with offspring health.26 Reduced glucose uptake is a major consequence of insulin resistance, and the decreased glucose uptake capacity of placenta leads to the increase in cord blood glucose level, which further promotes the high glucose environment in offspring, leading to the occurrence of long-term metabolic diseases in offspring. Therefore, improving the placental glucose uptake capacity is the key to ameliorating the high glucose environment of offspring and preventing long-term metabolic diseases in offspring.
Our study demonstrates that the increased expression of TXNIP and NLRP3 may be related to the downregulation of miR-17-5p in GDM placenta. MiR-17-5p inhibited the activation of the NLRP3 inflammasome in HTR8/SVneo cells, which could be reversed by TXNIP overexpression. Furthermore, miR-17-5p could improve the glucose uptake of HTR8/SVneo cells treated with high-glucose.
Some studies confirmed the effect of abnormal miRNA expression on the pathogenesis of GDM.27 For example, miR-518d up-regulation may be related to the pathogenesis of GDM by regulating PPARα expression28 and miR-98 up-regulation in the placenta not only induced DNA methylation but also regulated Mecp2, which targeted Trpc3, a major regulator of cellular glucose uptake.12 Our study has found that miR-17-5p expression was decreased in GDM placenta. Down-regulation of miR-17-5p has been found in a variety of metabolic diseases,29 and this miRNA is considered as the strongest predictor of metabolic syndrome status.14 We also found that miR-17-5p overexpression reversed the decrease in the ability of HTR8/SVneo cellular glucose uptake induced by high-glucose stimulation, which might highlight the potential therapeutic role of miR-17-5p on GDM. Thus, we propose the possibility that miR-17-5p could improve the health of GDM offspring by ameliorating high glucose environment in utero, and we further investigate its mechanism.
NLRP3 inflammasome participates in the regulation of interleukin (IL)-1β and IL-18,30 which have been considered as important inflammatory cytokines in the development of insulin resistance during GDM.31 Wu et al found the excessive activation of NLRP3 inflammasome in GDM placental tissues,32 which was consistent with our results. Besides, NLRP3 inflammasome activation can impair insulin sensitivity by promoting macrophage-T cell activation in adipose tissues.33 Our study also found that when NLRP3 expression was inhibited in HTR8/SVneo cells, the glucose uptake capacity of cells was increased. This conclusion further suggests that NLRP3 inflammasome plays a critical role in insulin resistance.
TXNIP is an endogenous negative regulator of cellular TRX, which can block its antioxidative function and may be separated from TRX and bind to NLRP3, leading to activation of NLRP3 inflammasome.34,35 Previous study has confirmed that TXNIP was increased in placental tissues of GDM patients, and this result was consistent with ours.22 The regulatory relationship between TXNIP and NLRP3 has been confirmed in several studies,21,36 but never in HTR8/SVneo cells and GDM. Our study found a positive correlation between TXNIP and NLRP3 expression in GDM placenta and the regulatory role of TXNIP in NLRP3 in HTR8/SVneo cells. Besides, TXNIP is a target of miR-17-5p, which has been confirmed in hypoxic-ischaemic rat models and in PC12 cells.37 Lerner et al used dual luciferase reporter assays to confirm the direct interaction of miR-17-5p and TXNIP,24 which was supported by our results. Since the inhibitory effect of miR-17-5p on NLRP3 expression can be reversed by TXNIP overexpression, we concluded that the regulation of NLRP3 by miR-17-5p is TXNIP-mediated. Liu et al reported that miR-17-5p improved glucose tolerance and suppressed pancreatic β-cell pyroptosis in diabetic mice, which might be caused by inhibiting TXNIP/NLRP3 inflammasome pathway.38
Based on our experimental results, we propose the following hypothesis which is similar to positive feedback: High glucose environment in GDM leads to a decrease of miR-17-5p in the placenta (the decrease of miR-17-5p is the result of high glucose environment, rather than the initiating cause), which results in the increased expression of TXNIP, leading to activation of NLRP3 inflammasome. NLRP3 inflammasome participates in the regulation of Interleukin (IL)-1β and IL-18 and impairs insulin sensitivity by promoting macrophage-T cell activation in adipose tissues, further contributing to insulin resistance and impaired glucose uptake and exacerbating the high glucose environment in utero. We demonstrated that miR-17-5p could terminate this positive feedback, underscoring its therapeutic effect. We also verified the effect of TXNIP/NLRP3 axis on insulin resistance and glucose uptake of HTR8/SVneo cells, providing a new idea for offspring health of GDM patients.
There are two obvious limitations in our study. First, we only detected the expression of NLRP3 inflammasome which can result in the generation of cleaved caspase-1, and produces IL-1β and IL-18, but we did not detect the expression of caspase-1, IL-1β and IL-18. Therefore, to further explore the correlation between inflammasome pathway and glucose uptake or insulin resistance in HTR8/SVneo cells is our next key point. Second, we only detected the glucose uptake capacity of HTR8/SVneo cells but did not detect the factors related to insulin resistance, like p-AKT and p-PI3K, so it is hard to say that improved glucose uptake is related to insulin resistance alleviation. Detection of these insulin resistance-related factors is also our next step, and animal experiment is also underway. Besides, the mechanism of miR-17-5p decrease caused by high glucose needs to be further studied.
Clinical Samples
Our study complies with the declaration of Helsinki.
Ethics Statement
Approval of the research protocol: The research protocol was approved by the Medical Ethics Committee of the Tongji Hospital of Huazhong University of Science and Technology.
Informed Consent
All participants signed their written informed consent.
Acknowledgments
This work was supported by the Health Commission of Hubei Province scientific research project (WJ2021M129) and the National Key Research and Development Program of China (2021YFC2701502).
Disclosure
The authors declare no conflict of interest.
References
1. Sacks DA, Hadden DR, Maresh M, et al. Frequency of gestational diabetes mellitus at collaborating centers based on IADPSG consensus panel-recommended criteria: the Hyperglycemia and Adverse Pregnancy Outcome (HAPO) study. Diabetes Care. 2012;35(3):526–528. doi:10.2337/dc11-1641
2. Wendland EM, Torloni MR, Falavigna M, et al. Gestational diabetes and pregnancy outcomes--a systematic review of the World Health Organization (WHO) and the International Association of Diabetes in Pregnancy Study Groups (IADPSG) diagnostic criteria. BMC Pregnancy Childbirth. 2012;12:23.
3. O’Sullivan EP, Avalos G, O’Reilly M, et al. Atlantic Diabetes in Pregnancy (DIP): the prevalence and outcomes of gestational diabetes mellitus using new diagnostic criteria. Diabetologia. 2011;54(7):1670–1675.
4. Mortier I, Blanc J, Tosello B, et al. Is gestational diabetes an independent risk factor of neonatal severe respiratory distress syndrome after 34 weeks of gestation? A prospective study. Arch Gynecol Obstet. 2017;296(6):1071–1077.
5. Metzger BE, Coustan DR, Trimble ER, et al. Hyperglycemia and adverse pregnancy outcomes. N Engl J Med. 2008;358(19):1991–2002.
6. Fadl HE, Ostlund IK, Magnuson AF, et al. Maternal and neonatal outcomes and time trends of gestational diabetes mellitus in Sweden from 1991 to 2003. Diabet Med. 2010;27(4):436–441.
7. Billionnet C, Mitanchez D, Weill A, et al. Gestational diabetes and adverse perinatal outcomes from 716,152 births in France in 2012. Diabetologia. 2017;60(4):636–644.
8. Plows JF, Stanley JL, Baker PN, et al. The pathophysiology of gestational diabetes mellitus. Int J Mol Sci. 2018;19:11.
9. Catalano PM. Trying to understand gestational diabetes. Diabet Med. 2014;31(3):273–281.
10. Silva-Zolezzi I, Samuel TM, Spieldenner J. Maternal nutrition: opportunities in the prevention of gestational diabetes. Nutr Rev. 2017;75(suppl 1):32–50. doi:10.1093/nutrit/nuw033
11. Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight?. Nat Rev Genet. 2008;9(2):102–114. doi:10.1038/nrg2290
12. Cao JL, Zhang L, Li J, et al. Up-regulation of miR-98 and unraveling regulatory mechanisms in gestational diabetes mellitus. Sci Rep. 2016;6(1):32268. doi:10.1038/srep32268
13. Li L, Wang S, Li H, et al. microRNA-96 protects pancreatic beta-cell function by targeting PAK1 in gestational diabetes mellitus. Biofactors. 2018;44(6):539–547. doi:10.1002/biof.1461
14. Ramzan F, D’Souza RF, Durainayagam BR, et al. Circulatory miRNA biomarkers of metabolic syndrome. Acta Diabetol. 2020;57(2):203–214. doi:10.1007/s00592-019-01406-6
15. Chen Y, Tian L, Wan S, et al. MicroRNA-17-92 cluster regulates pancreatic beta-cell proliferation and adaptation. Mol Cell Endocrinol. 2016;437:213–223. doi:10.1016/j.mce.2016.08.037
16. Stienstra R, van Diepen JA, Tack CJ, et al. Inflammasome is a central player in the induction of obesity and insulin resistance. Proc Natl Acad Sci U S A. 2011;108(37):15324–15329. doi:10.1073/pnas.1100255108
17. Zhang R, Zhang X, Xing B, et al. Astragaloside IV attenuates gestational diabetes mellitus via targeting NLRP3 inflammasome in genetic mice. Reprod Biol Endocrinol. 2019;17(1):77. doi:10.1186/s12958-019-0522-7
18. Ward R, Ergul A. Relationship of endothelin-1 and NLRP3 inflammasome activation in HT22 hippocampal cells in diabetes. Life Sci. 2016;159:97–103.
19. Cho KA, Kang PB. PLIN2 inhibits insulin-induced glucose uptake in myoblasts through the activation of the NLRP3 inflammasome. Int J Mol Med. 2015;36(3):839–844.
20. Nasoohi S, Parveen K, Ishrat T. Metabolic syndrome, brain insulin resistance, and Alzheimer’s disease: Thioredoxin Interacting Protein (TXNIP) and inflammasome as core amplifiers. J Alzheimers Dis. 2018;66(3):857–885.
21. Zhou R, Tardivel A, Thorens B, et al. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat Immunol. 2010;11(2):136–140.
22. Sarina LDF, Feng ZQ, et al. Mechanism of placenta damage in gestational diabetes mellitus by investigating TXNIP of patient samples and gene functional research in cell line. Diabetes Ther. 2019;10(6):2265–2288.
23. Dong D, Fu N, Yang P. MiR-17 downregulation by high glucose stabilizes thioredoxin-interacting protein and removes thioredoxin inhibition on ASK1 leading to apoptosis. Toxicol Sci. 2016;150(1):84–96.
24. Lerner AG, Upton JP, Praveen PV, et al. IRE1alpha induces thioredoxin-interacting protein to activate the NLRP3 inflammasome and promote programmed cell death under irremediable ER stress. Cell Metab. 2012;16(2):250–264.
25. Kampmann U, Knorr S, Fuglsang J, et al. Determinants of maternal insulin resistance during pregnancy: an updated overview. J Diabetes Res. 2019;2019:5320156.
26. Kelly AC, Powell TL, Jansson T. Placental function in maternal obesity. Clin Sci (Lond). 2020;134(8):961–984.
27. Cai M, Kolluru GK, Ahmed A. Small molecule, big prospects: microRNA in pregnancy and its complications. J Pregnancy. 2017;2017:6972732.
28. Zhao C, Zhang T, Shi Z, et al. MicroRNA-518d regulates PPARalpha protein expression in the placentas of females with gestational diabetes mellitus. Mol Med Rep. 2014;9(6):2085–2090.
29. Heneghan HM, Miller N, McAnena OJ, et al. Differential miRNA expression in omental adipose tissue and in the circulation of obese patients identifies novel metabolic biomarkers. J Clin Endocrinol Metab. 2011;96(5):E846–850.
30. Fang X, Wang Y, Zhang Y, et al. NLRP3 inflammasome and its critical role in gynecological disorders and obstetrical complications. Front Immunol. 2020;11:555826.
31. Gomes CP, Torloni MR, Gueuvoghlanian-Silva BY, et al. Cytokine levels in gestational diabetes mellitus: a systematic review of the literature. Am J Reprod Immunol. 2013;69(6):545–557.
32. Wu W, Tan QY, Xi FF, et al. NLRP3 inflammasome activation in gestational diabetes mellitus placentas is associated with hydrogen sulfide synthetase deficiency. Exp Ther Med. 2022;23(1):94.
33. Vandanmagsar B, Youm YH, Ravussin A, et al. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med. 2011;17(2):179–188.
34. Mohamed IN, Hafez SS, Fairaq A, et al. Thioredoxin-interacting protein is required for endothelial NLRP3 inflammasome activation and cell death in a rat model of high-fat diet. Diabetologia. 2014;57(2):413–423.
35. Yang C, Xia W, Liu X, et al. Role of TXNIP/NLRP3 in sepsis-induced myocardial dysfunction. Int J Mol Med. 2019;44(2):417–426.
36. Tschopp J, Schroder K. NLRP3 inflammasome activation: the convergence of multiple signalling pathways on ROS production?. Nat Rev Immunol. 2010;10(3):210–215.
37. Gamdzyk M, Doycheva DM, Kang R, et al. GW0742 activates miR-17-5p and inhibits TXNIP/NLRP3-mediated inflammation after hypoxic-ischaemic injury in rats and in PC12 cells. J Cell Mol Med. 2020;24(21):12318–12330.
38. Liu S, Tang G, Duan F, et al. MiR-17-5p inhibits TXNIP/NLRP3 inflammasome pathway and suppresses pancreatic beta-cell pyroptosis in diabetic mice. Front Cardiovasc Med. 2021;8:768029.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.