")
Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 13
miR-100 rs1834306 A>G Increases the Risk of Hirschsprung Disease in Southern Chinese Children
Authors Zhu Y, Lin A , Zheng Y, Xie X, He Q, Zhong W
Received 2 June 2020
Accepted for publication 27 July 2020
Published 10 August 2020 Volume 2020:13 Pages 283—288
DOI https://doi.org/10.2147/PGPM.S265730
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Yun Zhu,* Ao Lin,* Yi Zheng, Xiaoli Xie, Qiuming He, Wei Zhong
Department of Pediatric Surgery, Guangzhou Institute of Pediatrics, Guangdong Provincial Key Laboratory of Research in Structural Birth Defect Disease, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, Guangzhou, Guangdong 510623, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wei Zhong
Department of Pediatric Surgery, Guangzhou Institute of Pediatrics, Guangdong Provincial Key Laboratory of Research in Structural Birth Defect Disease, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, Guangzhou, Guangdong 510623, People’s Republic of China
Tel/ Fax +86-18902268667
Email [email protected]
Background: Hirschsprung disease (HSCR) is a rare congenital gastrointestinal disease characterized by the absence of intestinal submucosal and myometrial ganglion cells. Recently, researches indicated that miR-100 regulated the growth, differentiation and apoptosis of neurons, and affected the functions of HSCR-associated pathways. While miR-100 rs1834306 A>G polymorphism was shown to modify the susceptibility to tumors, the association between this polymorphism and HSCR susceptibility is still unknown.
Methods: This was a case–control study consisting of 1470 HSCR cases and 1473 controls from southern China. DNA was genotyped by TaqMan real-time PCR. Odds ratios (ORs) and 95% confidence intervals (CIs) were used as statistical indicators.
Results: We found that miR-100 rs1834306 G allele and GG genotype significantly increased HSCR susceptibility (GG vs AA: adjusted OR=1.31, 95% CI=1.04– 1.64, P=0.020; G vs A: adjusted OR=1.12, 95% CI=1.01– 1.25, P=0.041; GG vs AA/AG: adjusted OR=1.30, 95% CI=1.07– 1.59, P=0.010). In the stratified analysis, miR-100 rs1834306 GG genotype carriers had higher risk to develop HSCR in all clinical subtypes when compared with those with AA/AG genotypes, and OR was rising with HSCR aggravation (SHSCR: adjusted OR=1.28, 95% CI=1.03– 1.59, P=0.029; LHSCR: adjusted OR=1.48, 95% CI=1.06– 2.07, P=0.020; TCA: adjusted OR=2.12, 95% CI=1.22– 3.69, P=0.008).
Conclusion: Our findings suggested that miR-100 rs1834306 A>G polymorphism was associated with increased HSCR susceptibility in southern Chinese children. Furthermore, miR-100 rs1834306 GG genotype had a greater genetic pathopoiesis in severe HSCR.
Keywords: Hirschsprung disease, miR-100, polymorphism, susceptibility
Introduction
Hirschsprung disease (HSCR) is a rare congenital gastrointestinal disease in pediatric surgery. Approximately 80% of HSCR are sporadic and the global incidence is 1/5000. It is reported that males are more predisposed to HSCR than females.1 HSCR is characterized by the absence of intestinal submucosal and myometrial ganglion cells, which dues to the event that enteric neural crest-derived cells (ENCDCs) fail to invade, proliferate and migrate to distal colon during embryonic development.2,3 Severe HSCR will lead to death. Surgery is the only effective treatment. However, postoperative complications such as constipation, fecal incontinence, repeated enteritis and malnutrition continue to bother the survivors. People are actively exploring the intestinal nerve stem cell transplantation technology to improve the prognosis of HSCR, but the progress has been slow.4,5 Genetic factors play an important role in the pathogenesis and prognosis of HSCR. Any genes affecting cell proliferation, differentiation and migration may cause HSCR.6–8 Therefore, expounding the genetic etiologies of HSCR is of great significance for the diagnosis and treatment.
MicroRNAs (miRNAs) belong to non-coding RNAs that are about 22nt long. MiRNAs can specifically recognize the 3ʹUTRs of targeted mRNAs, and subsequently, lead to translation inhibition and mRNA degradation mediated by RNA Induced Silencing Complex (RISC). In this way, miRNAs regulate nearly 60% of human transcripts.9 Up to now, many miRNAs such as miR-150-5p, miR-143, miR-637, miR-483-3p, miR-1324, miR-215, miR-24-1, let-7a, miR-939, miR-770-5p, miR-369-3p, miR-483-5p, miR-206 and miR-214 have been shown to affect the occurrence and development of HSCR.10–21 However, no research has explored the role of miR-100 to HSCR.
MiR-100 is a popular cancer-associated molecule involved in the proliferation, differentiation, apoptosis and invasion of tumor cells.22 MiR-100 rs1834306 A>G polymorphism is located in pre-miR-100. In the past, Chang et al and Zhu et al observed that miR-100 rs1834306 G allele reduced the risk of esophageal squamous cell carcinoma and endometriosis.23,24 As for colorectal cancer, Boni et al found that the GG/GA genotypes lengthened the time to progression,25 while Lampropoulou et al pointed out that a similar effect was performed by the GG genotype.26 In addiction, some scholars supported a view that miR-100 rs1834306 A>G polymorphism was not associated with tumors.27–31
Recently, some researches indicated that miR-100 regulated the growth, differentiation and apoptosis of neurons and directly/indirectly affected HSCR-associated pathways.8,10,12,16,32-34 Considering that rs1834306 may be a potential function site of miR-100, we conducted a case–control study to explore the association between miR-100 rs1834306 A>G polymorphism and HSCR susceptibility in Southern Chinese population.
Patients and Methods
Study Population
During 2000 to 2015, a total of 1470 cases and 1473 controls were collected from Guangzhou Women and Children’s Medical Center. There were 240 females (16.33%) and 1230 males (83.67%) in the cases, 458 females (65.65%) and 1015 males (34.35%) in the controls. The frequency distribution of age and gender between cases and controls were significantly different (P<0.001), detailed demographic characteristics can refer to Supplemental Table 1. By histological examination of biopsy specimens, the children with the microscopic characteristics of ganglion cell disappearance and/or submucosal plexus hyperplasia were finally diagnosed with HSCR. The children traveling this hospital at the same period for health examination were recruited as controls. Exclusion criterion: (1) Non-southern Chinese; (2) familiar HSCR or syndromic HSCR; (3) owning the history of other congenital gastrointestinal malformations and endocrine disorders; (4) receiving therapy before blood drawing. According to the range of intestinal lesion, the patients were divided into the groups of short-segment HSCR (SHSCR), long-segment HSCR (LHSCR) and total colonic aganglionosis (TCA).35 Our study followed the Helsinki Declaration and was supported by the institutional review board of Guangzhou Women and Children’s Medical Center (ethic approve number: 201943800). Written informed consents were obtained from the participants or their guardians.
SNP Selection and Genotyping
NCBI dbSNP database (http://www.ncbi.nlm.nih.gov/projects/SNP) and SNPinfo (https://snpinfo.niehs.nih.gov/snpinfo/snpfunc.html) were used to screen SNPs. Detailed selection criteria were described in the previous article.36 Genomic DNA was extracted by the TIANamp Blood DNA Kit and TIANquick FFPE DNA Kit (TianGen Biotech Co., Ltd., Beijing, China). The TaqMan SNP Genotyping Assay (Applied Biosystems, Foster City, CA, USA) was used for genotyping.37–39 Experiments were conducted in strict accordance with the instructions. No false-positive result was found in negative controls (sterile water) and the repeatable rate of genotyping was 100%.
Statistical Analysis
SAS 9.1 was used for data analysis. Hardy-Weinberg equilibrium (HWE) in controls was estimated by the good-of-fit test. The differences in demographic characteristics between patients and controls were evaluated by the T-test and chi-square test. To adjust the confounding effect caused by age and gender, we utilized the unconditional multiple logistic regression model to estimate the association between miR-100 rs1834306 A>G polymorphism and HSCR susceptibility. Stratified analysis was performed by HSCR clinical subtypes (SHSCR, LHSCR and TCA). Odds ratios (ORs) and 95% confidence intervals (CIs) were used as statistical indicators. All tests were two-sided. When P<0.05, results were considered statistically significant.
Results
miR-100 rs1834306 A>G Polymorphisms and HSCR Susceptibility
As shown in Table 1, a total of 1398 cases and 1445 controls were successfully genotyped, the genotype frequency of miR-100 rs1834306 in controls was conformed to HWE (P>0.05). After adjusting for age and gender, we found that G allele and GG genotype significantly increased HSCR susceptibility (GG vs AA: adjusted OR=1.31, 95% CI=1.04–1.64, P=0.020; additive model: adjusted OR=1.12, 95% CI=1.01–1.25, P=0.041; GG vs AA/AG: adjusted OR=1.30, 95% CI=1.07–1.59, P=0.010).
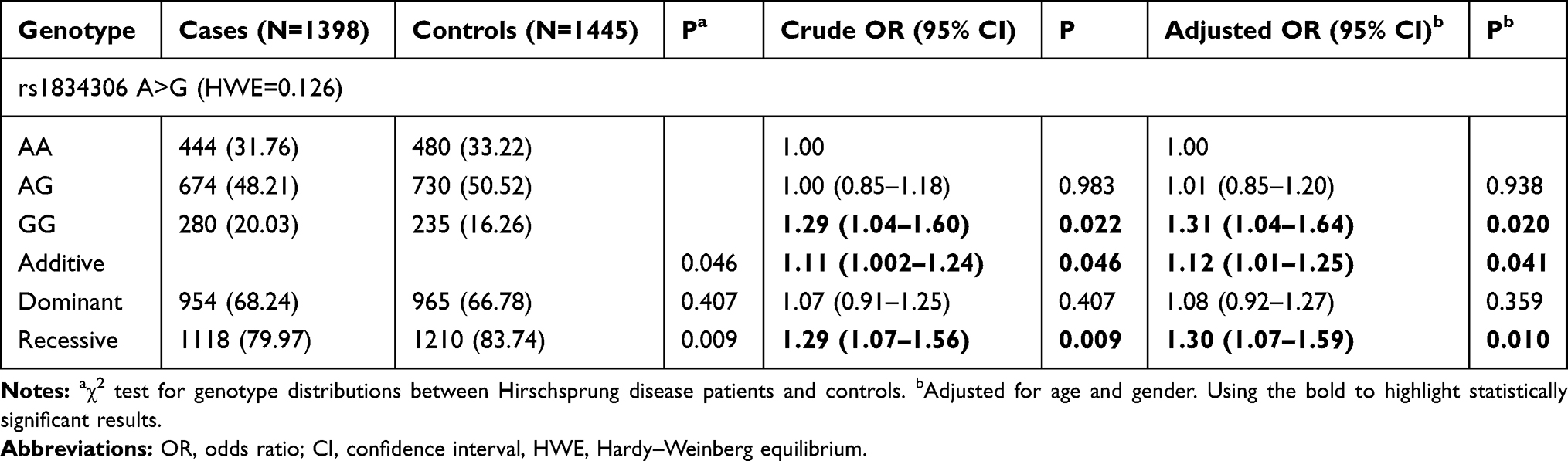 |
Table 1 Association Between miR-100 rs1834306 A>G Polymorphism and Hirschsprung Disease Susceptibility |
Stratification Analysis
As shown in Table 2, stratified analysis revealed that miR-100 rs1834306 GG genotype carriers had higher risk to develop HSCR in all clinical subtypes (SHSCR, LHSCR and TCA) when compared with those with AA/AG genotypes. Further, OR was rising with HSCR aggravation (SHSCR: adjusted OR=1.28, 95% CI=1.03–1.59, P=0.029; LHSCR: adjusted OR=1.48, 95% CI=1.06–2.07, P=0.020; TCA: adjusted OR=2.12, 95% CI=1.22–3.69, P=0.008).
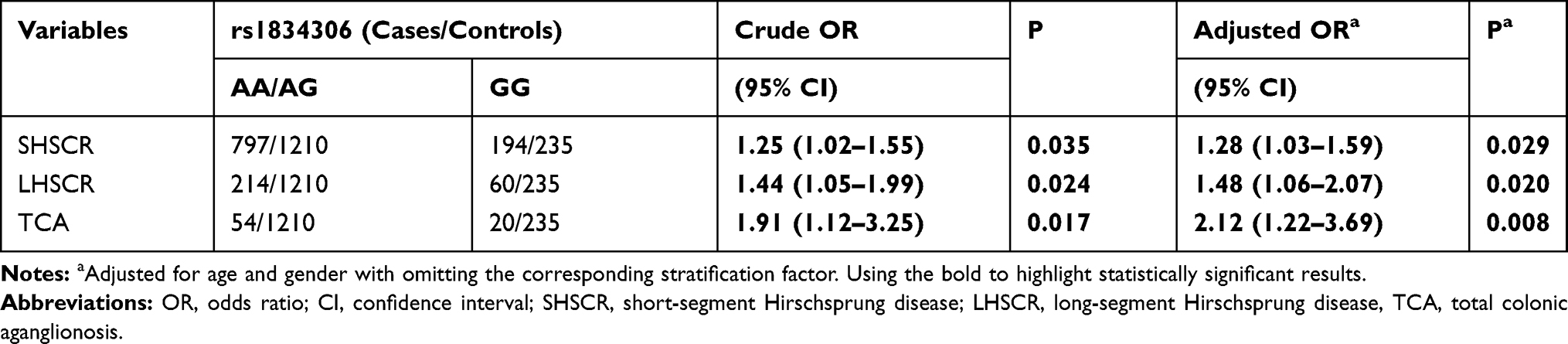 |
Table 2 Stratification Analysis for the Association Between miR-100 rs1834306 A>G and Hirschsprung Disease Susceptibility (by Subtype) |
Discussion
Both enteric nervous system (ENS) and central nervous system (CNS) originate from neural crest, and show high similarities in structure and neurochemistry.40 In fact, most HSCR patients with Waardenburg syndromes type 4 or Goldberg-Shprintzen syndrome exhibit some CNS symptoms such as cochlear nerve deafness, seizures or developmental delay.41,42 Therefore, ENS regulates gastrointestinal function independently of CNS, but we cannot conclude that the developments of ENS and CNS are completely independent. Genetic etiologies that lead to CNS disease may also cause ENS dysfunction.40 Recently, Kong et al found that reducing the expression of miR-100 blocked hydroperoxide-induced apoptosis, promoted the growth and differentiation of retinal ganglion cells, and activated the TrkB-AKT-ERK pathway through phosphorylation, suggesting that miR-100 not only regulated the development of tumor cells, but also that of neurons.32 It is noting that the PI3/ATK and RAS/ERK pathways have been found to help the RET pathway mediate signals which is one of the most important signal pathways implicated in HSCR pathogenesis.8
In addition, miR-100 was found to block the metastasis of hepatoma cells by inhibiting ICMT-RAC1 pathway, while RAC1 was found to activate PI3K/ATK pathway.33,34 Interestingly, Tang et al observed that RAC1/2 were inactivated after miR-241 and Let-7a inhibiting ARP2/3, thereby suppressing the migration and proliferation of the 293T and SH-SY5Y cell lines, and proposed that the mir-241/let-7a-ARP2/3 complex-RAC isoforms pathway was a potential pathogenic mechanism for HSCR.16
Above evidences suggested that miR-100 may take part in the development of HSCR by a complex regulatory network. We hypothesized that increased expression of miR-100 inhibited AKT and ERK by the TrkB-AKT-ERK or ICMT-RAC1 pathways to block the signal mediation of the RET pathway, besides it can also inhibit the downstream gene of the mir-241/let-7a-ARP2/3 complex-RAC isoforms pathway (RAC1) by ICMT-RAC1 pathway, thus promoting the occurrence and progression of HSCR.
Pri-miRNAs and pre-miRNAs are important precursors of miRNAs. SNPs in the precursors will lead to abnormal expression and secondary structure of miRNAs, which further changes the regulation of miRNAs to targeted mRNAs, eventually cause disease.43 In our study, miR-100 rs1834306 G allele and GG genotype were associated with increased HSCR susceptibility. The same conclusion was observed in all the HSCR subtypes. Furthermore, OR increased gradually with HSCR aggravation, suggesting that GG genotype had a greater genetic pathopoiesis in severe HSCR.
As predicted by SNPinfo, miR-100 rs1834306 is a transcription factor binding site. In the past, Motawi et al verified that miR-100 rs1834306 G allele reduced the expression of miR-100 in hepatitis B.44 It seemed to conflict with our prediction that GG genotype would increase miR-100 expression to promote HSCR by directly/indirectly suppressing the functions of HSCR-associated pathways. The explanations for this contradiction were as follows. First, the same polymorphism may play different roles in diverse diseases. For example, pre-miR-146a rs2910164 CC genotype or C allele was found to reduce the expression of miR-146a and associated with lower risk to develop prostate cancer and hepatocellular carcinoma.45,46 However, Shen et al observed that pre-miR-146a rs2910164 C allele increased the expression of miR-146a and associated with increased susceptibility to earlier familial breast and ovarian cancer.47 Therefore, we could not rule out the possibility that rs1834306 G allele or GG genotype upregulated the expression of miR-100 in HSCR. Second, a research observed that MIR143HG, the precursor of miR-143, acted as a molecular sponge which could enhance the expression of RBM24 by competitively binding to miR-143.11 The similar mechanism may exist between pre-miR-100 and miR-100. We guessed that rs1834306 A>G polymorphism would lead to down-regulated miR-100 by decreasing the expression or changing the secondary structure of pre-miR-100. Subsequently, the dysfunction of pre-miR-100 could weaken its combination with miR-100, thereby enhancing the inhibition of mir-100 to HSCR-associated pathways. Finally, the pathogenesis of HSCR and the regulation of miR-100 are complex, down-regulated miR-100 may restore a more advantageous pathway that inhibits the proliferation and migration of ENCDCs.3 These hypotheses need further verification.
This is the first research about miR-100 and HSCR. However, there are some limitations in our study. 1. Our study is limited to southern Chinese and the conclusion may not be extrapolated to other populations. 2. Although we have found a significant association between mir-100 rs1834306 A>G polymorphism and HSCR susceptibility, we do not conduct any functional experiment to verify this finding.
Conclusion
In conclusion, we suggested that miR-100 rs1834306 A>G polymorphism was associated with increased HSCR susceptibility in southern Chinese children. Furthermore, miR-100 rs1834306 GG genotype had a greater genetic pathopoiesis in severe HSCR.
Funding
This study was funded by grants from the Natural Science Foundation of Guangdong Province, China (No: 2019A1515010971), and Guangdong Provincial Key Laboratory of Research in Structural Birth Defect Disease (No: 2019B030301004).
Disclosure
The authors have no conflicts of interest to declare for this work.
References
1. Amiel J, Sproat-Emison E, Garcia-Barcelo M, et al. Hirschsprung disease, associated syndromes and genetics: a review. J Med Genet. 2008;45(1):1–14. doi:10.1136/jmg.2007.053959
2. Obermayr F, Hotta R, Enomoto H, Young HM. Development and developmental disorders of the enteric nervous system. Nat Rev Gastroenterol Hepatol. 2013;10(1):43–57. doi:10.1038/nrgastro.2012.234
3. Lake JI, Heuckeroth RO. Enteric nervous system development: migration, differentiation, and disease. Am J Physiol Gastrointest Liver Physiol. 2013;305(1):G1–24. doi:10.1152/ajpgi.00452.2012
4. Shu X, Meng Q, Jin H, et al. Treatment of aganglionic megacolon mice via neural stem cell transplantation. Mol Neurobiol. 2013;48(3):429–437. doi:10.1007/s12035-013-8430-x
5. Gustafson E, Larsson T, Danielson J. Controlled outcome of Hirschsprung’s disease beyond adolescence: a single center experience. Pediatr Surg Int. 2019;35(2):181–185. doi:10.1007/s00383-018-4391-5
6. Heuckeroth RO, Schafer KH. Gene-environment interactions and the enteric nervous system: neural plasticity and Hirschsprung disease prevention. Dev Biol. 2016;417(2):188–197. doi:10.1016/j.ydbio.2016.03.017
7. Badner JA, Sieber WK, Garver KL, Chakravarti A. A genetic study of Hirschsprung disease. Am J Hum Genet. 1990;46(3):568–580.
8. Sergi CM, Caluseriu O, McColl H, Eisenstat DD. Hirschsprung’s disease: clinical dysmorphology, genes, micro-RNAs, and future perspectives. Pediatr Res. 2017;81(1–2):177–191. doi:10.1038/pr.2016.202
9. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/s0092-8674(04)00045-5
10. Peng L, Chen G, Zhu Z, et al. Circular RNA ZNF609 functions as a competitive endogenous RNA to regulate AKT3 expression by sponging miR-150-5p in Hirschsprung’s disease. Oncotarget. 2017;8(1):808–818. doi:10.18632/oncotarget.13656
11. Du C, Shen Z, Zang R, et al. Negative feedback circuitry between MIR143HG and RBM24 in Hirschsprung disease. Biochim Biophys Acta. 2016;1862(11):2127–2136. doi:10.1016/j.bbadis.2016.08.017
12. Li Y, Zhou L, Lu C, et al. Long non-coding RNA FAL1 functions as a ceRNA to antagonize the effect of miR-637 on the down-regulation of AKT1 in Hirschsprung’s disease. Cell Prolif. 2018;51(5):e12489. doi:10.1111/cpr.12489
13. Zhi Z, Zhu H, Lv X, et al. IGF2-derived miR-483-3p associated with Hirschsprung’s disease by targeting FHL1. J Cell Mol Med. 2018;22(10):4913–4921. doi:10.1111/jcmm.13756
14. Zhou L, Li Y, Jiang W, et al. Down-regulation of circ-PRKCI inhibits cell migration and proliferation in Hirschsprung disease by suppressing the expression of miR-1324 target PLCB1. Cell Cycle. 2018;17(9):1092–1101. doi:10.1080/15384101.2018.1480210
15. Lei H, Li H, Xie H, et al. Role of MiR-215 in Hirschsprung’s Disease Pathogenesis by Targeting SIGLEC-8. Cell Physiol Biochem. 2016;40(6):1646–1655. doi:10.1159/000453214
16. Tang W, Cai P, Huo W, et al. Suppressive action of miRNAs to ARP2/3 complex reduces cell migration and proliferation via RAC isoforms in Hirschsprung disease. J Cell Mol Med. 2016;20(7):1266–1275. doi:10.1111/jcmm.12799
17. Chen G, Du C, Shen Z, et al. MicroRNA-939 inhibits cell proliferation via targeting LRSAM1 in Hirschsprung’s disease. Aging. 2017;9(12):2471–2479. doi:10.18632/aging.101331
18. Li H, Li B, Zhu D, et al. Downregulation of lncRNA MEG3 and miR-770-5p inhibit cell migration and proliferation in Hirschsprung’s disease. Oncotarget. 2017;8(41):69722–69730. doi:10.18632/oncotarget.19207
19. Pan W, Yu H, Zheng B, et al. Upregulation of MiR-369-3p suppresses cell migration and proliferation by targeting SOX4 in Hirschsprung’s disease. J Pediatr Surg. 2017;52(8):1363–1370. doi:10.1016/j.jpedsurg.2017.04.002
20. Wang G, Guo F, Wang H, et al. Downregulation of microRNA-483-5p Promotes Cell Proliferation and Invasion by Targeting GFRA4 in Hirschsprung’s Disease. DNA Cell Biol. 2017;36(11):930–937. doi:10.1089/dna.2017.3821
21. Wu L, Yuan W, Chen J, et al. Increased miR-214 expression suppresses cell migration and proliferation in Hirschsprung disease by interacting with PLAGL2. Pediatr Res. 2019;86(4):460–470. doi:10.1038/s41390-019-0324-9
22. Li C, Gao Y, Zhang K, et al. Multiple Roles of MicroRNA-100 in human cancer and its therapeutic potential. Cell Physiol Biochem. 2015;37(6):2143–2159. doi:10.1159/000438572
23. Chang CY, Lai M-T, Chen Y, et al. Up-regulation of ribosome biogenesis by MIR196A2 genetic variation promotes endometriosis development and progression. Oncotarget. 2016;7(47):76713–76725. doi:10.18632/oncotarget.11536
24. Zhu J, Yang L, You W, et al. Genetic variation in miR-100 rs1834306 is associated with decreased risk for esophageal squamous cell carcinoma in Kazakh patients in northwest China. Int J Clin Exp Pathol. 2015;8(6):7332–7340.
25. Boni V, Zarate R, Villa JC, et al. Role of primary miRNA polymorphic variants in metastatic colon cancer patients treated with 5-fluorouracil and irinotecan. Pharmacogenomics J. 2011;11(6):429–436. doi:10.1038/tpj.2010.58
26. Lampropoulou DI, Aravantinos G, Laschos K, et al. MiR-218 and miR-100 polymorphisms as markers of irinotecan-based chemotherapy response in metastatic colorectal cancer. Int J Colorectal Dis. 2019;34(11):1871–1877. doi:10.1007/s00384-019-03401-3
27. Stenholm L, Stoehlmacher-Williams J, Al-Batran SE, et al. Prognostic role of microRNA polymorphisms in advanced gastric cancer: a translational study of the Arbeitsgemeinschaft Internistische Onkologie (AIO). Ann Oncol. 2013;24(10):2581–2588. doi:10.1093/annonc/mdt330
28. Hu Y, Yu CY, Wang JL, et al. MicroRNA sequence polymorphisms and the risk of different types of cancer. Sci Rep. 2014;4(1):3648. doi:10.1038/srep03648
29. Chuanyin L, Xiaona W, Zhiling Y, et al. The association between polymorphisms in microRNA genes and cervical cancer in a Chinese Han population. Oncotarget. 2017;8(50):87914–87927. doi:10.18632/oncotarget.21235
30. Danesh H, Hashemi M, Bizhani F, Hashemi SM, Bahari G. Association study of miR-100, miR-124-1, miR-218-2, miR-301b, miR-605, and miR-4293 polymorphisms and the risk of breast cancer in a sample of Iranian population. Gene. 2018;647:73–78. doi:10.1016/j.gene.2018.01.025
31. Xiong XD, Luo, XP, Cheng, J, et al. A genetic variant in pre-miR-27a is associated with a reduced cervical cancer risk in southern Chinese women. Gynecol Oncol. 2014;132(2):450–454. doi:10.1016/j.ygyno.2013.12.030
32. Kong N, Lu X, Li B. Downregulation of microRNA-100 protects apoptosis and promotes neuronal growth in retinal ganglion cells. BMC Mol Biol. 2014;15(1):25. doi:10.1186/s12867-014-0025-1
33. Zhou HC, Fang J-H, Luo X, et al. Downregulation of microRNA-100 enhances the ICMT-Rac1 signaling and promotes metastasis of hepatocellular carcinoma cells. Oncotarget. 2014;5(23):12177–12188. doi:10.18632/oncotarget.2601
34. Tolias KF, Cantley LC, Carpenter CL. Rho family GTPases bind to phosphoinositide kinases. J Biol Chem. 1995;270(30):17656–17659. doi:10.1074/jbc.270.30.17656
35. Zhang Y, He, Q, Zhang, R, et al. Large-scale replication study identified multiple independent SNPs in RET synergistically associated with Hirschsprung disease in Southern Chinese population. Aging. 2017;9(9):1996–2009. doi:10.18632/aging.101294
36. He J, Zou Y, Liu X, et al. Association of common genetic variants in pre-microRNAs and neuroblastoma susceptibility: a two-center study in chinese children. Mol Ther Nucleic Acids. 2018;11:1–8. doi:10.1016/j.omtn.2018.01.003
37. He J, Wang F, Zhu J, et al. Association of potentially functional variants in the XPG gene with neuroblastoma risk in a Chinese population. J Cell Mol Med. 2016;20(8):1481–1490. doi:10.1111/jcmm.12836
38. He J, Yang T, Zhang R, et al. Potentially functional polymorphisms in the LIN28B gene contribute to neuroblastoma susceptibility in Chinese children. J Cell Mol Med. 2016;20(8):1534–1541. doi:10.1111/jcmm.12846
39. He J, Zhang X, Zhang J, et al. LMO1 super-enhancer polymorphism rs2168101 G>T correlates with decreased neuroblastoma risk in Chinese children. J Cancer. 2018;9(9):1592–1597. doi:10.7150/jca.24326
40. Rao M, Gershon MD. The bowel and beyond: the enteric nervous system in neurological disorders. Nat Rev Gastroenterol Hepatol. 2016;13(9):517–528. doi:10.1038/nrgastro.2016.107
41. Shahar E, Shinawi M. Neurocristopathies presenting with neurologic abnormalities associated with Hirschsprung’s disease. Pediatr Neurol. 2003;28(5):385–391. doi:10.1016/s0887-8994(03)00010-9
42. Lecerf L, Kavo A, Ruiz-Ferrer M, et al. An impairment of long distance SOX10 regulatory elements underlies isolated Hirschsprung disease. Hum Mutat. 2014;35(3):303–307. doi:10.1002/humu.22499
43. Hrovatin K, Kunej T. Classification of miRNA-related sequence variations. Epigenomics. 2018;10(4):463–481. doi:10.2217/epi-2017-0126
44. Motawi TK, Mady AE, Shaheen S, et al. Genetic variation in microRNA-100 (miR-100) rs1834306 T/C associated with Hepatitis B virus (HBV) infection: correlation with expression level. Infect Genet Evol. 2019;73:444–449. doi:10.1016/j.meegid.2019.06.009
45. Xu B, et al. A functional polymorphism in Pre-miR-146a gene is associated with prostate cancer risk and mature miR-146a expression in vivo. Prostate. 2010;70(10):467–472. doi:10.1002/pros.21080
46. Xu T, Zhu Y, Wei Q-K, et al. A functional polymorphism in the miR-146a gene is associated with the risk for hepatocellular carcinoma. Carcinogenesis. 2008;29(11):2126–2131. doi:10.1093/carcin/bgn195
47. Shen J, Ambrosone CB, DiCioccio RA, et al. A functional polymorphism in the miR-146a gene and age of familial breast/ovarian cancer diagnosis. Carcinogenesis. 2008;29(10):1963–1966. doi:10.1093/carcin/bgn172
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.