")
Back to Journals » Cancer Management and Research » Volume 11
MicroRNA-665 inhibits the oncogenicity of retinoblastoma by directly targeting high-mobility group box 1 and inactivating the Wnt/β-catenin pathway
Authors Wang S, Du S, Lv Y, Zhang F, Wang W
Received 6 January 2019
Accepted for publication 25 February 2019
Published 11 April 2019 Volume 2019:11 Pages 3111—3123
DOI https://doi.org/10.2147/CMAR.S200566
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Chien-Feng Li
This paper has been retracted
Shuai Wang, Shanshan Du, Yong Lv, Fengyan Zhang, Wenzhan Wang
Department of Ophthalmology, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan 450052, People’s Republic of China
Purpose: Previous studies have revealed that microRNA-665 (miR-665) is dysregulated in a variety of human cancers. However, little is known regarding its expression profiles and functions in retinoblastoma (RB). Therefore, the aims of our study were to evaluate miR-665 expression in RB and determine the precise roles of miR-665 in the progression of RB.
Patients and methods: Herein, RT-qPCR was used to determine miR-665 expression levels in RB tissues and cell lines, and a series of functional experiments were performed to explore the influence of miR-665 on RB cell proliferation, colony formation, apoptosis, migration, and invasion as well as tumor growth. The molecular mechanisms underlying the tumor-suppressive action of miR-665 in RB were also explored.
Results: We found that miR-665 was markedly reduced in RB tissues and cell lines and that lower miR-665 expression was strongly associated with tumor size, TNM stage, and differentiation in patients with RB. Exogenous expression of miR-665 suppressed cell proliferation, colony formation, migration, and invasion, and induced cell apoptosis in RB cells, while silencing miR-665 expression had the opposite effects. In addition, upregulation of miR-665 decreased the tumor growth of RB cells in vivo. High-mobility group box 1 (HMGB1) was identified as a direct target of miR-665 in RB cells, and decreasing the expression of HMGB1 simulated the regulatory effects of miR-665 overexpression in RB cells, while knockdown of HMGB1 expression counteracted the miR-665-mediated antitumor effects in RB cells. Moreover, miR-665 was shown to regulate the Wnt/β-catenin signaling pathway by targeting HMGB1 in vitro and in vivo.
Conclusion: Taken together, our in vitro and in vivo results suggest that miR-665 acts as a tumor-suppressive miRNA in RB by directly targeting HMGB1 and inactivating the Wnt/β-catenin pathway. Hence, this miRNA is a candidate prognostic biomarker and therapeutic target in patients with RB.
Keywords: microRNA-665, retinoblastoma, high-mobility group box 1, Wnt/β-catenin pathway, oncogenicity
Introduction
Retinoblastoma (RB) is an aggressive intraocular malignancy that arises from the primitive retinal layer.1 It accounts for 2–4% of all malignant tumors in children under 5 years of age.2 Approximately 9,000 cases of RB are diagnosed every year worldwide,3 and approximately 20% of these occur in China.4 In developing countries, most patients are diagnosed with RB at an advanced stage; therefore, their clinical outcomes are poorer than those of patients in developed countries.5,6 The primary treatment options for patients with RB are enucleation, laser photocoagulation, chemotherapy, and focal therapy.7 Although tremendous progress has been made in the diagnosis and treatment of RB over the past decade,8 the prognosis is still unsatisfactory due to its rapid progression. The allelic inactivation of the RB1 gene along with other oncogenes or tumor suppressors has been shown to play a crucial role in the development and progression of RB;9,10 however, the exact molecular mechanisms are poorly understood. Therefore, delineating the molecular events involved in the pathogenesis of RB is of great and urgent significance for the identification of effective molecular therapeutic strategies for patients with RB.
In recent years, our knowledge of the importance of microRNAs (miRNAs) in cancer has greatly increased.11 miRNAs are a group of short (19–23 nt), noncoding RNA molecules that function as novel gene expression regulators.12 miRNAs pair imperfectly with the 3′-untranslated region (3′-UTR) of their target genes, which leads to mRNA degradation and/or transcription silencing.13 Because miRNAs modulate approximately 60% of all human protein-coding genes, they have been implicated in various physiological functions and pathological conditions, including carcinogenesis and cancer progression.14–16 Numerous studies have shown that the expression profiles of various miRNAs are altered in RB.17–19 For example, miR-101-3p,20 miR-506-3p,21 and miR-87422 are expressed at low levels in RB and function as tumor-suppressive miRNAs. In contrast, miR-10b,23 miR-198,24 and miR-49825 are upregulated in RB and have a tumor-promoting functions. Therapeutic techniques targeting the miRNAs that contribute to the initiation and progression of RB may have potential applications.
Abnormal miR-665 expression has been reported in multiple human cancers.26–29 However, the expression profile and biological function of miR-665 in RB and the underlying molecular mechanisms are largely unknown. Hence, in this study, we aimed to detect miR-665 expression in RB and evaluate its clinical value in patients with RB. Additionally, we investigated the function of miR-665 in RB progression and explored the molecular mechanisms underlying the tumor-suppressive action of miR-665 in RB.
Material and methods
Patients and tissue samples
The study protocol was approved by the Ethics Committee of The First Affiliated Hospital of Zhengzhou University, and the study was performed in accordance with the guidelines of the Declaration of Helsinki. Written informed consent was obtained from the parents or guardians of all patients. A total of 34 RB tissues and 11 normal retinal tissues were collected at The First Affiliated Hospital of Zhengzhou University between March 2015 and October 2017. Normal retinal tissues were obtained from patients with ophthalmorrhexis who received enucleation. All enrolled patients were newly diagnosed and were being treated for the first time, and they had not been treated with laser photocoagulation, chemotherapy, or focal therapy before enucleation. All tissue specimens were quickly frozen in liquid nitrogen after enucleation and stored at −80 °C until use.
Cell lines and culture conditions
The normal retinal pigmented epithelial cell line ARPE-19 and three human RB cell lines, Y79, SO-RB50, and WERI-RB-1, were acquired from the American Type Culture Collection (Manassas, VA, USA). The cell lines were maintained at 37 °C in a humidified atmosphere containing 5% CO2 and grown in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Gibco) and 1% penicillin/streptomycin (Sigma-Aldrich, St. Louis, MO, USA).
Cell transfection
Human synthetic miR-665 mimics, a miR-665 inhibitor, negative control miRNA mimics (miR-NC), and a negative control miRNA inhibitor (NC inhibitor) were purchased from GenePharma (Shanghai, China). To knockdown HMGB1, an HMGB1-targeting small interfering RNA (siRNA; si-HMGB1) and a corresponding negative control (si-NC) were purchased from GeneCopoeia (Guangzhou, China). To increase HMGB1 expression, the full-length human HMGB1 cDNA was chemically synthesized by GenePharma and then inserted into the mammalian expression vector pcDNA3.1(+) (Invitrogen, Carlsbad, CA, USA) to generate the plasmid pcDNA3.1-HMGB1 (pc-HMGB1). Cells were plated in 6-well plates 12 h prior to transfection, and Lipofectamine 2000 (Invitrogen) was utilized for cell transfection. Transfected cells were collected at different time points and then used for subsequent experiments.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
TRIzol (Invitrogen) was used to extract total RNA from prepared tissue specimens and cells. Then, the extracted total RNA was reverse transcribed using the miScript Reverse Transcription kit (Qiagen, Hilden, Germany). The obtained cDNA and the miScript SYBR Green PCR kit (Qiagen) were used to quantify miR-665 expression by using quantitative PCR (qPCR). To determine HMGB1 mRNA expression, the reverse transcription reaction was performed using the PrimeScript™ RT reagent Kit (Takara, Kusatsu, Japan), followed by qPCR with SYBR-Green PCR Master Mix (Takara). The RT-qPCR was performed on an ABI 7900 thermocycler (Applied Biosystems, Foster City, CA, USA). Relative miR-665 and HMGB1 expression levels were normalized to the reference genes U6 small nuclear RNA and GAPDH, respectively, and relative gene expression was analyzed by the 2−ΔΔCt method.30
Cell counting kit-8 (CCK-8) assay
Transfected cells were collected after 24 h of culture, resuspended in culture medium, and seeded into 96-well plates at a density of 2,000 cells per well. Each group had six replicate wells. The cells were then incubated at 37 °C in a humidified atmosphere containing 5% CO2. After 0, 24, 48, and 72 h of incubation, 10 μL of CCK-8 solution (DOJINDO, Tokyo, Japan) was added to each well and incubated at 37 °C for an additional 2 h. Then, the plates were read on a microplate reader (Bio-Rad, Hercules, CA, USA) at a wavelength of 450 nm.
Clonogenic assay
Transfected cells were collected at 24 h post-transfection and plated in 6-well plates at an initial density of 1,000 cells/well. Then, the cells were grown at 37 °C in a humidified incubator with 5% CO2 for 2 weeks. On day 15, the cells were fixed with 4% paraformaldehyde and then stained with methyl violet. Finally, the number of colonies (>50 cells) was counted under an inverted light microscope (Olympus, Tokyo, Japan).
Flow cytometric analysis of apoptotic cells
After 24 h of culture, transfected cells were collected, and the apoptosis rate was determined. After three washes with PBS, apoptotic cells were evaluated using the Annexin V-fluorescein isothiocyanate (FITC) apoptosis detection kit (BioLegend, San Diego, CA, USA). Cells were resuspended in 1× binding buffer, and the cell density was adjusted to 1×108 cells/mL. Then, the cells were stained with 5 μL of Annexin V-FITC and 5 μL of propidium iodide solution. After 15 min of incubation in the dark at 4 °C, the cells were evaluated with a flow cytometer (FACScan; BD Biosciences, San Jose, CA, USA). The data were analyzed with CellQuest software (BD Biosciences).
In vitro migration and invasion assays
The migration and invasion of RB cells were assessed using transwell inserts (8 μm pores; BD Biosciences). In the invasion assays, the upper side of the insert was precoated with Matrigel (BD Biosciences), while a non-Matrigel-coated insert was used for the migration assays. At 48 h post-transfection, cell suspensions were prepared using FBS-free DMEM. Then, 300 μL of a cell suspension containing 5×104 transfected cells was placed into the upper compartment of the insert, and 700 μL of DMEM supplemented with 20% FBS was added to the lower compartment. The Transwells were incubated at 37 °C in a humidified atmosphere containing 5% CO2 for 24 h, and then the cells were fixed with 4% paraformaldehyde and stained with 0.5% crystal violet. Finally, the number of migrated and invaded cells was counted under an inverted light microscope (magnification, 200×).
Tumor xenograft mouse model
All animal care and experimental protocols were approved by the Ethical Committee of The First Affiliated Hospital of Zhengzhou University, and the experiments were carried out in accordance with the Animal Protection Law of the People’s Republic of China-2009 for experimental animals. Seven-week-old BALB/c nude mice were obtained from the Chinese Academy of Sciences (Shanghai, China) and subcutaneously injected with miR-665 mimic- or miR-NC-transfected cells. Each group contained four mice. The tumors were monitored every 2 days, and tumor volume was calculated using the following formula: tumor volume = 0.5 × long diameter × short diameter2. Four weeks after injection, the mice were sacrificed, and the weight of the tumor xenografts was measured.
Bioinformatics prediction
The putative targets of miR-665 were predicted using three miRNA target prediction programs, TargetScan (
Luciferase reporter assay
The wild-type (wt) and mutant (mut) 3′-UTR of HMGB1, containing putative and mutated miR-665 binding sites, respectively, were chemically synthesized by GenePharma and inserted into the pmirGLO luciferase reporter vector (Promega, Madison, WI, USA). The luciferase plasmids were then co-transfected with miR-665 mimics or miR-665 inhibitor into cells and plated in 24-well plates. At 48 h after transfection, the cells were harvested and lysed, and luciferase activity was measured using the Dual Luciferase Reporter Assay System (Promega) according to the manufacturer’s instructions. Relative firefly luciferase activity was normalized to Renilla luciferase activity.
Western blot analysis
Total protein was extracted with the active protein extraction kit (KGP1050; Nanjing KeyGen Biotech, Nanjing, China), and protein levels were quantified with the BCA Protein Assay Kit (Beyotime Biotechnology, Shanghai, China). Equal amounts of protein were loaded onto 10% sodium dodecyl sulfate polyacrylamide gels, separated by electrophoresis, and transferred to polyvinylidene fluoride membranes (Beyotime Biotechnology). After blocking with 5% skimmed milk at room temperature for 2 h, the membranes were incubated overnight at 4 °C with primary antibodies against HMGB1 (ab77302; Abcam, Cambridge, MA, USA), p-β-catenin (sc-57534; Santa Cruz Biotechnology, Santa Cruz, CA, USA), β-catenin (sc-59737; Santa Cruz Biotechnology), cyclin D1 (ab40754; Abcam), and GAPDH (ab128915; Abcam), which were diluted to 1:1,000. The, the membranes were incubated with goat anti-rabbit (ab205718; Abcam) or goat anti-mouse (ab6789; Abcam) HRP-conjugated secondary antibodies at a dilution of 1:5,000 for detection. The bands were visualized using enhanced chemiluminescence solution (Pierce; Thermo Fisher Scientific, Inc.). GAPDH was used as a loading control.
Statistical analysis
All statistical analyses were performed using SPSS 19.0 (SPSS Inc., Chicago, IL, USA). Each assay was repeated at least three times, and all data were expressed as mean ± standard error. The chi-squared test was used to evaluate the correlation between miR-665 levels and clinicopathological parameters in patients with RB. Spearman’s correlation analysis was used for the correlation analysis between miR-665 and HMGB1 mRNA levels in tissues obtained from patients with RB. Two-tailed Student’s t-test and one-way analysis of variance with Student-Newman-Keuls tests were utilized for comparisons between two groups and multiple groups, respectively. P-values less than 0.05 were considered statistically significant.
Results
miR-665 expression is decreased in RB tissues and cell lines
To determine the expression profile of miR-665 in RB, we first detected its expression in 34 RB tissues and 11 normal retinal tissues. The results of the RT-qPCR analysis showed that miR-665 expression levels were lower in RB tissues than in normal retinal tissues (Figure 1A, P<0.05). We also measured miR-665 expression in the RB cell lines Y79, SO-RB50, and WERI-RB-1 and the normal retinal pigmented epithelial cell line ARPE-19. Consistently, lower miR-665 expression was observed in all three RB cell lines when compared with the level in ARPE-19 cells (Figure 1B, P<0.05).
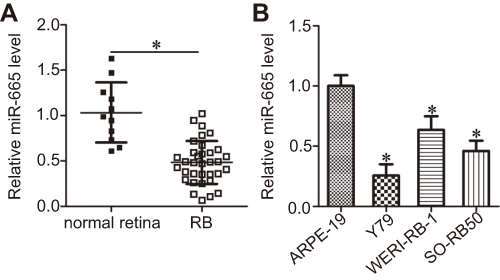 | Figure 1 Decreased miR-665 expression in RB tissues and cell lines. (A) RT-qPCR was used to determine the expression profile of miR-665 in 34 RB tissues and 11 normal retina tissues. *P<0.05 compared with normal retina tissues. (B) The miR-665 expression level was assessed in three RB cell lines (Y79, SO-RB50, and WERI-RB-1) and a normal retinal pigmented epithelial cell line (ARPE-19). *P<0.05 compared with ARPE-19. |
We next attempted to evaluate the clinical role of miR-665 in patients with RB. All enrolled patients were divided into either the low or high miR-665 expression group using the median value as the cutoff. The statistical analysis showed that low miR-665 expression was significantly related to tumor size (P=0.015), TNM stage (P=0.004), and differentiation (P=0.032, Table 1) in patients with RB. These results indicate that downregulation of miR-665 might play a critical role in the malignant development of RB.
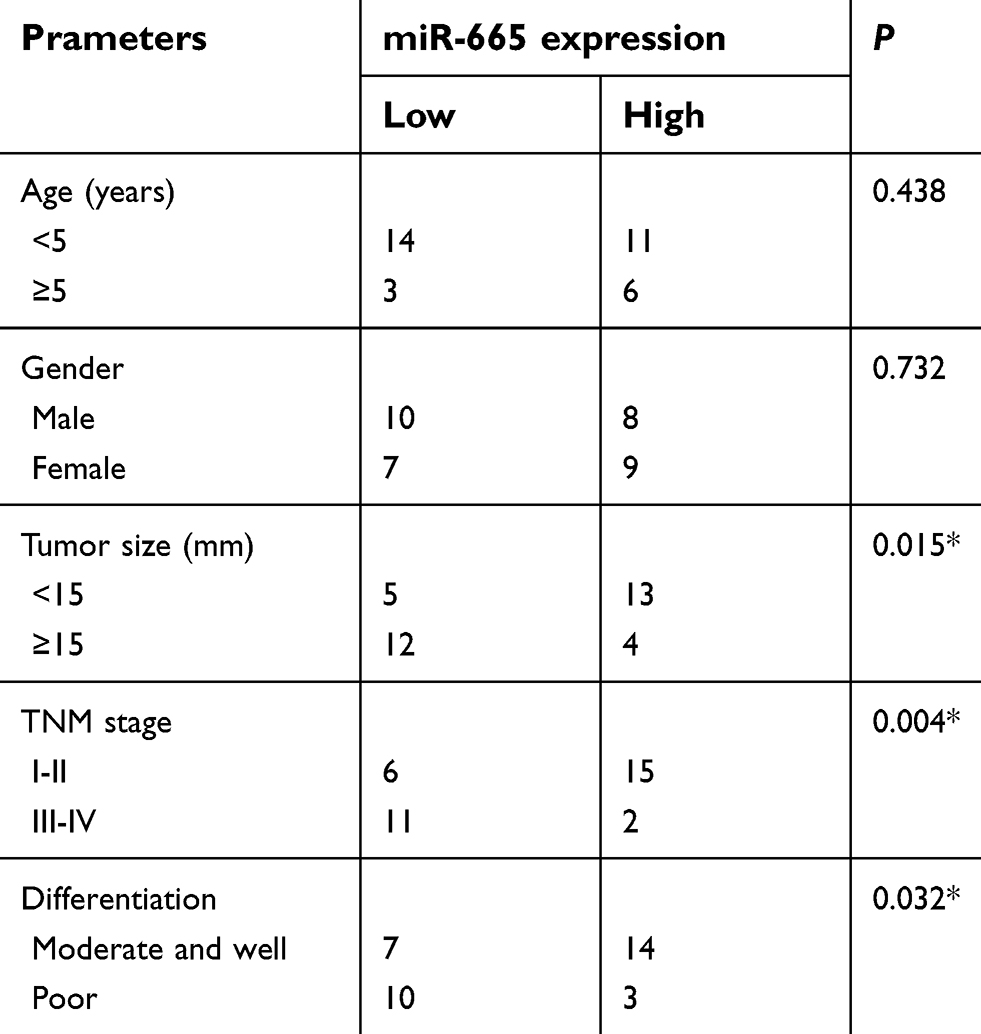 | Table 1 The association between clinicopathological parameters and miR-665 expression in 34 patients with RB. RB patients were divided into a low miR-665 or high miR-665 expression group based on median value of miR-665 expression in RB tissues |
miR-665 attenuates the malignant phenotypes of RB cells in vitro
To explore the biological functions of miR-665 in RB, gain- and loss-of-function assays were performed to determine the effect of altering miR-665 expression on the phenotypes of RB cells. Y79 cells, which exhibited the lowest miR-665 expression among the three tested RB cell lines, were transfected with miR-665 mimics, while WERI-RB-1 cells, which showed the highest miR-665 expression level, were transfected with a miR-665 inhibitor. Transfection efficiency was validated by RT-qPCR (Figure 2A, P<0.05). CCK-8 and clonogenic assays revealed that transfection of Y79 cells with the miR-665 mimics decreased cell proliferation and colony formation, respectively, while transfection of WERI-RB-1 cells with the miR-665 inhibitor led to enhancement of cell proliferation and colony formation (Figure 2B and C, P<0.05). Since miR-665 inhibited RB cell proliferation, we next examined whether miR-665 regulated apoptosis in RB cells. As shown in Figure 2D, ectopic miR-665 expression in Y79 cells led to an obvious increase in apoptosis, while silencing miR-665 expression in WERI-RB-1 cells led to a marked reduction in apoptosis (P<0.05). We also investigated the effects of miR-665 on the migration and invasion of RB cells in vitro. The results from the in vitro migration and invasion assays showed that restoration of miR-665 expression restricted the migration and invasion of Y79 cells. In contrast, downregulation of miR-665 promoted the migration and invasion of WERI-RB-1 cells (Figure 2E and F, P<0.05). Taken together, these results indicate that miR-665 may function as a tumor-suppressing miRNA in RB.
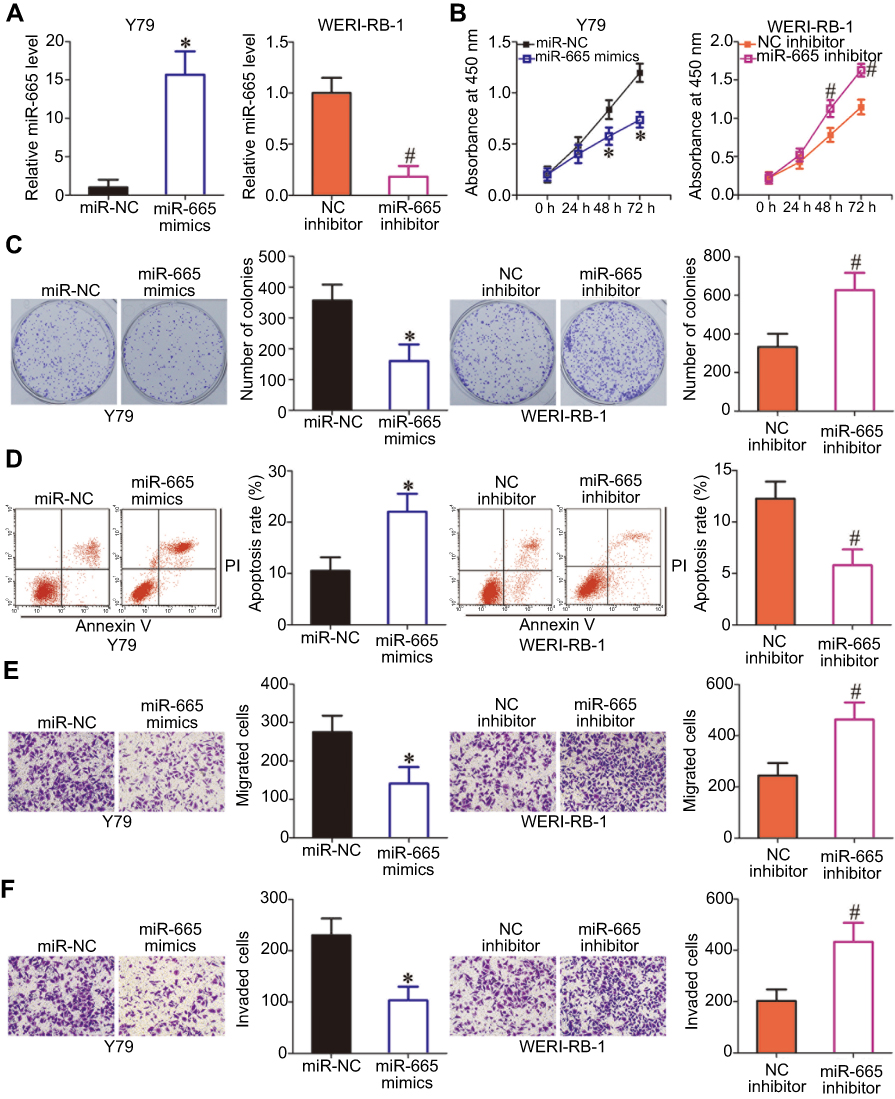 | Figure 2 Effects of miR-665 on RB cell proliferation, colony formation, apoptosis, migration, and invasion in vivo. (A) Y79 cells were transfected with miR-665 mimics or miR-NC, and WERI-RB-1 cells were transfected with a miR-665 inhibitor or NC inhibitor. After transfection, miR-665 expression was assessed by RT-qPCR. *P<0.05 compared with miR-NC. #P<0.05 compared with NC inhibitor. (B, C) CCK-8 and clonogenic assays were performed to assess cell proliferation and colony formation, respectively, in Y79 cells overexpressing miR-665 and in WERI-RB-1 cells with knocked down miR-665. On day 15, a photo of the colonies formed was taken. *P<0.05 compared with miR-NC. #P<0.05 compared with NC inhibitor. (D) Apoptosis was examined by flow cytometric analysis in Y79 cells transfected with miR-665 mimics and WERI-RB-1 cells transfected with a miR-665 inhibitor. *P<0.05 compared with miR-NC. #P<0.05 compared with NC inhibitor. (E, F) Cell migration and invasion were evaluated after transfection of Y79 cells with miR-665 mimics or the transfection of WERI-RB-1 cells with miR-665 inhibitor. *P<0.05 compared with miR-NC. #P<0.05 compared with NC inhibitor. |
HMGB1 is a direct target of miR-665 in RB cells
Bioinformatics analysis was performed to search for putative targets of miR-665 and determine the mechanisms responsible for the effects of miR-665 in RB cells. The 3′-UTR of HMGB1 contained a highly conserved binding site for miR-665 (Figure 3A), suggesting that HMGB1 is a direct target of miR-665. Thus, a luciferase reporter assay was conducted to verify this prediction. The luciferase activity of a plasmid harboring the wild-type HMGB1 3′-UTR was decreased by upregulation of miR-665 expression in Y79 cells, whereas inhibition of miR-665 in WERI-RB-1 cells increased luciferase activity (Figure 3B, P<0.05). However, the luciferase activity of a plasmid containing a mutant HMGB1 3′-UTR was unaffected by a change in miR-665 expression.
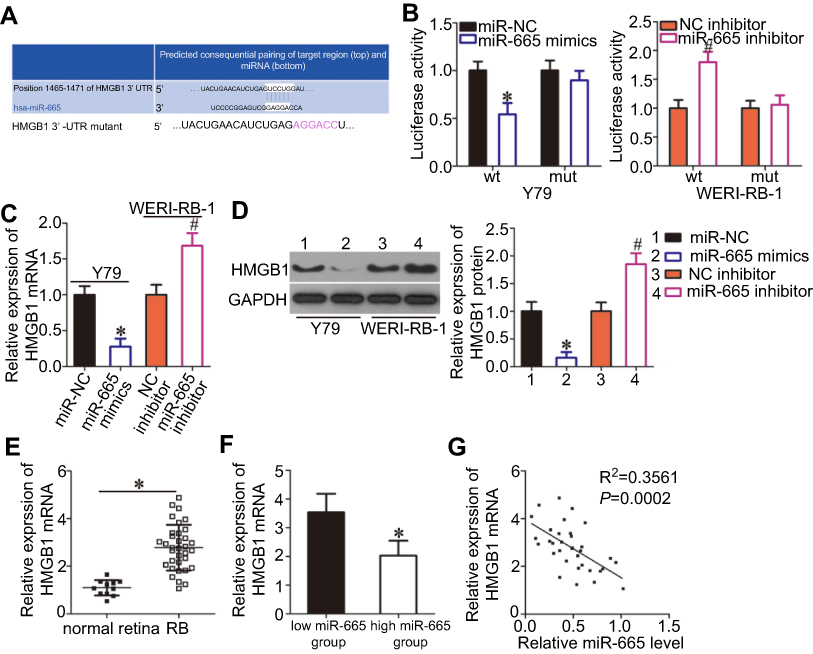 | Figure 3 Identification of HMGB1 as a direct target gene of miR-665 in RB cells. (A) Bioinformatics prediction revealed a highly conserved miR-665 binding site in the 3′-UTR of HMGB1. (B) Y79 cells were treated with either miR-665 mimics or miR-NC and either wild-type (wt) or mutant (mut) 3′-UTR reporter plasmid. miR-665 inhibitor or NC inhibitor along with the wt or mut 3′-UTR reporter plasmid were transfected into WERI-RB-1 cells. Luciferase activity was measured at 48 h post-transfection and normalized to that of the Renilla luciferase activity. *P<0.05 compared with miR-NC. #P<0.05 compared with NC inhibitor. (C, D) Expression levels of HMGB1 mRNA and protein in miR-665-overexpressing Y79 cells and miR-665-knocked down WERI-RB-1 cells were measured by using RT-qPCR and Western blot analysis, respectively. On day 15, a photo of colonies formed was taken. *P<0.05 compared with miR-NC. #P<0.05 compared with NC inhibitor. (E) HMGB1 mRNA expression in 34 RB tissues and 11 normal retina tissues was analyzed by RT-qPCR. *P<0.05 compared with normal retina tissues. (F) HMGB1 mRNA expression in the high miR-665 expression group was significantly lower than that in the low miR-665 expression group. *P<0.05 compared with low miR-665 expression group. (F) Correlation analysis of the expression levels of miR-665 and HMGB1 in RB tissues was performed using Spearman’s correlation analysis. R2=0.3561 P=0.0002. |
RT-qPCR and Western blot analysis were employed to determine the effects of miR-665 on the endogenous expression levels of HMGB1 mRNA and protein. The results showed that the mRNA and protein levels of HMGB1 were suppressed by exogenous miR-665 expression in Y79 cells and were induced by silencing miR-665 expression in WERI-RB-1 cells (Figure 3C and D, P<0.05). Furthermore, we found that HMGB1 mRNA expression was notably upregulated in RB tissues relative to the level in normal retinal tissues (Figure 3E, P<0.05). RB tissues with high miR-665 expression showed lower HMGB1 mRNA levels than RB tissues with low miR-665 expression (Figure 3F, P<0.05). Moreover, Spearman’s correlation analysis indicated that the HMGB1 mRNA level was negatively correlated with the miR-665 level in RB tissues (Figure 3G; R2= 0.3561, P= 0.0002). These results suggest that HMGB1 is a direct target gene of miR-665 in RB.
HMGB1 knockdown can simulate the effect of miR-665 upregulation in RB cells
Loss-of-function assays were performed to investigate the biological functions of HMGB1 in RB progression. Western blot analysis was used to validate the knockdown efficiency of HMGB1 siRNA (si-HMGB1). Transfection of si-HMGB1 efficiently decreased HMGB1 protein levels in Y79 and WERI-RB-1 cells compared with the levels in cells transfected with si-NC (Figure 4A, P<0.05). Cell proliferation and colony formation in the si-HMGB1-transfected cells were attenuated when compared with those in si-NC-transfected cells for both the Y79 and WERI-RB-1 cell lines (Figure 4B and C, P<0.05). In addition, transfection of both Y79 and WERI-RB-1 cells with si-HMGB1 promoted apoptosis (Figure 4D, P<0.05). In vitro migration and invasion assays showed that HMGB1 knockdown also restricted the migration and invasion of Y79 and WERI-RB-1 cells (Figure 4E and F, P<0.05). These results demonstrated that suppression of HMGB1 had effects similar to those of miR-665 overexpression in RB cells, confirming HMGB1 as a downstream target of miR-665 in RB cells.
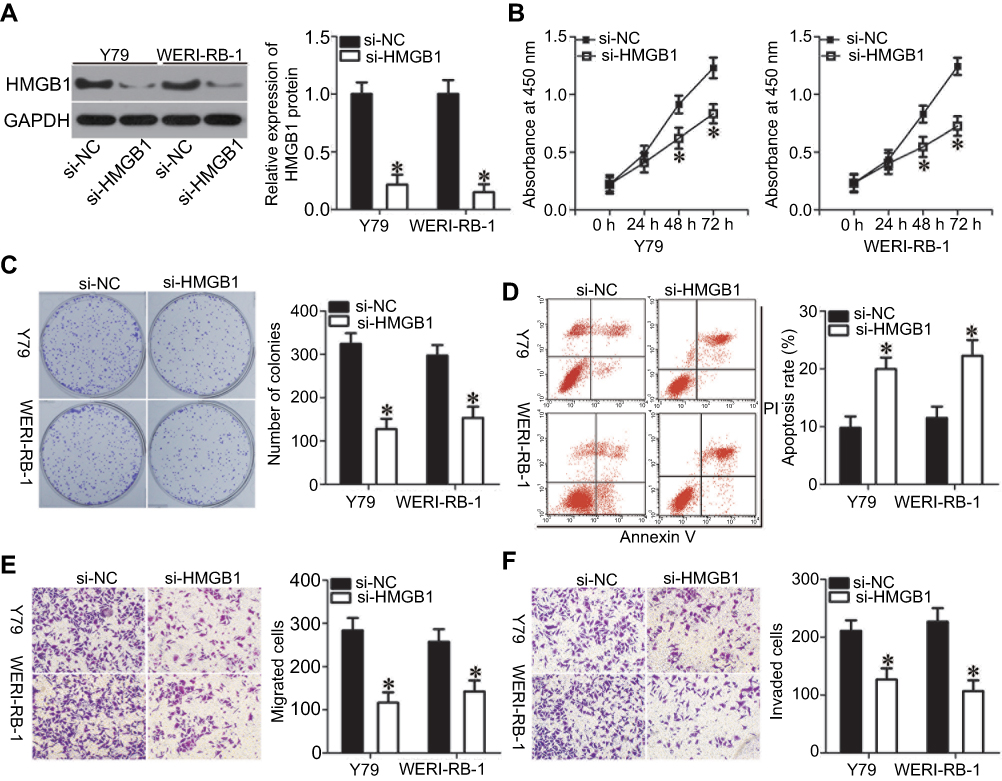 | Figure 4 Silencing of HMGB1 expression inhibited RB cell proliferation, colony formation, migration, and invasion, and promoted apoptosis in vitro. Y79 and WERI-RB-1 cells were transfected with si-HMGB1 or si-NC, and then used in the following assays. (A) Western blot analysis to detect HMGB1 protein levels. *P<0.05 compared with si-NC. (B, C) The effects of HMGB1 knockdown on proliferation and colony formation of Y79 and WERI-RB-1 cells were examined by using the CCK-8 and clonogenic assays, respectively. On day 15, a photo of colonies formed was taken. *P<0.05 compared with si-NC. (D) Flow cytometric analysis was applied to detect apoptosis in HMGB1-silenced Y79 and WERI-RB-1 cells. *P<0.05 compared with si-NC. (E, F) In vitro migration and invasion assays were used to investigate the migration and invasion of Y79 and WERI-RB-1 cells after transfection with si-HMGB1 or si-NC. *P<0.05 compared with si-NC. |
The functional roles of miR-665 in RB cells depend on HMGB1 expression
To investigate whether HMGB1 expression affects the functions of miR-665 in RB cells and the underlying mechanism, rescue experiments were performed by restoring HMGB1 expression in Y79 and WERI-RB-1 cells. To this end, miR-665-overexpressing Y79 cells were transfected with pc-HMGB1, and si-HMGB1 was introduced into WERI-RB-1 cells that were transfected with a miR-665 inhibitor. Restoration of HMGB1 protein expression was corroborated by Western blot analysis (Figure 5A, P<0.05). Functional experiments revealed that cell growth (Figure 5B–D, P<0.05) and metastasis (Figure 6A and B, P<0.05) in Y79 cells were inhibited by miR-665 overexpression; however, these inhibitory effects were abolished by co-transfection with pc-HMGB1. Similarly, co-transfection with si-HMGB1 partially counteracted the miR-665 inhibitor-induced promotion of WERI-RB-1 cell growth and metastasis. These results suggest that miR-665 exerts its anticancer effects in RB, at least in part, through the negative regulation of HMGB1.
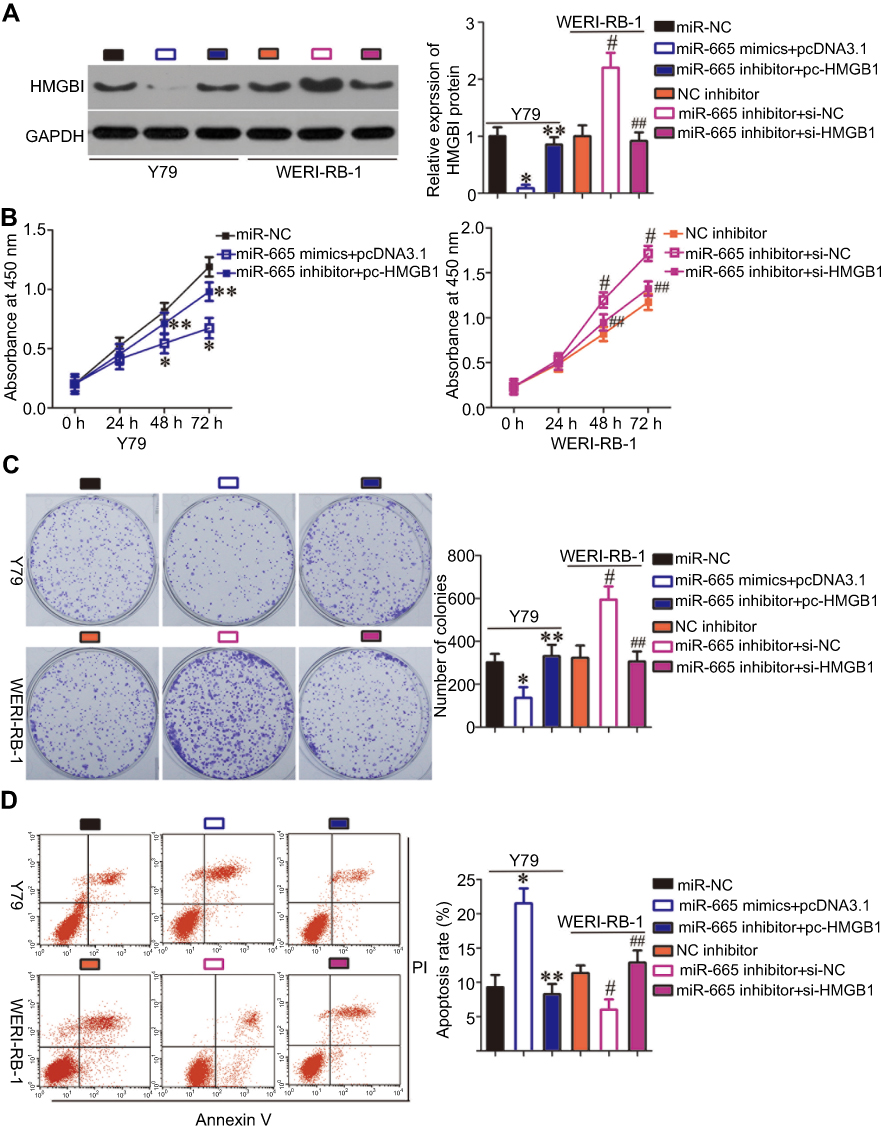 | Figure 5 Rescue experiments were performed to confirm that HMGB1 is the functional target of miR-665 in RB cells. Y79 cells were co-transfected with miR-665 mimics and either pc-HMGB1 or pcDNA3.1. WERI-RB-1 cells were co-transfected with miR-665 inhibitor and either si-HMGB1 or si-NC. After different incubation times, the cells were collected and used in the following experiments. (A) At 72 h post transfection, HMGB1 protein was measured by Western blot analysis. *P<0.05 compared with miR-NC. **P<0.05 compared with miR-665 mimics + pcDNA3.1. #P<0.05 compared with NC inhibitor. ##P<0.05 compared with miR-665 inhibitor + si-NC. (B-D) Cell proliferation, colony formation, and apoptosis were examined by using the CCK-8, clonogenic, and flow cytometric analysis, respectively. On day 15, a photo of colonies formed was taken. *P<0.05 compared with miR-NC. **P<0.05 compared with miR-665 mimics + pcDNA3.1. #P<0.05 compared with NC inhibitor. ##P<0.05 compared with miR-665 inhibitor + si-NC. |
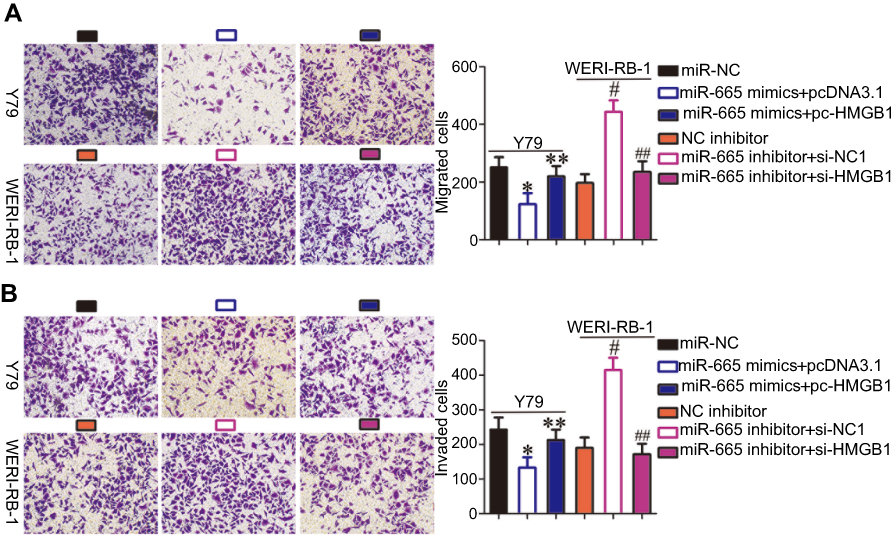 | Figure 6 Restoration of HMGB1 expression abolished the effects of miR-665 on the migration and invasion of RB cells. (A, B) Y79 cells were cotransfected with miR-665 mimics and either pc-HMGB1 or pcDNA3.1. WERI-RB-1 cells were cotransfected with miR-665 inhibitor and either si-HMGB1 or si-NC into. In vitro migration and invasion assays were conducted to examine cell migration and invasion. *P<0.05 compared with miR-NC. **P<0.05 compared with miR-665 mimics + pcDNA3.1. #P<0.05 compared with NC inhibitor. ##P<0.05 compared with miR-665 inhibitor + si-NC. |
miR-665 suppresses the Wnt/β-catenin signaling pathway in RB cells via regulation of HMGB1
The Wnt/β-catenin signaling pathway has previously been reported to be regulated by HMGB1.31,32 To investigate whether a change in miR-665 expression affects the Wnt/β-catenin pathway in RB cells, the expression levels of several important molecules in the pathway, including p-β-catenin, β-catenin, and cyclin D1, were measured in RB cells following upregulation or downregulation of miR-665. Transfection of Y79 cells with miR-665 mimics downregulated the protein levels of p-β-catenin and cyclin D1, while silencing miR-665 expression in WERI-RB-1 cells had the opposite effects (Figure 7). However, recovery of HMGB1 expression partially alleviated the change in p-β-catenin and cyclin D1 levels caused by miR-665 upregulation or downregulation. These results indicate that miR-665 deactivates the Wnt/β-catenin pathway in RB cells via negative regulation of HMGB1.
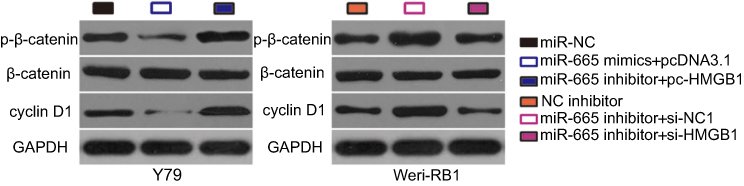 | Figure 7 miR-665 targets HMGB1 in RB cells to inactivate the Wnt/β-catenin signaling pathway Y79 cells were cotransfected with miR-665 mimics and either pc-HMGB1 or pcDNA3.1, while WERI-RB-1 cells were cotransfected with miR-665 inhibitor and either si-HMGB1 or si-NC. Western blot analysis was conducted to examine the expression levels of p-β-catenin, β-catenin, and cyclin D1. |
miR-665 inhibits tumor growth in vivo via HMGB1 and the Wnt/β-catenin pathway
To further illustrate the effects of miR-665 on RB cell growth, a tumor xenograft mouse model was established by inoculating Y79 cells transfected with the miR-665 mimics or miR-NC into the flanks of nude mice. The tumor xenografts generated from the miR-665 mimic-transfected cells were significantly smaller (Figure 8A and B, P<0.05) and lighter (Figure 8C, P<0.05) than the xenografts in mice of the miR-NC group. We then detected miR-665 expression in xenografts by RT-qPCR. The data showed that the expression level of miR-665 was significantly higher in xenografts originating from miR-665 mimic-transfected Y79 cells (Figure 8D, P<0.05). Western blot analysis was performed to determine the levels of HMGB1, p-β-catenin, β-catenin, and cyclin D1 in the xenografts, and the results were consistent with those of the in vitro experiments. The expression levels of HMGB1, p-β-catenin, and cyclin D1 proteins were significantly downregulated in the tumor xenografts of mice in the miR-665 mimics group (Figure 8E). These results show that miR-665 inhibits tumor growth by targeting HMGB1 via deactivation of the Wnt/β-catenin pathway.
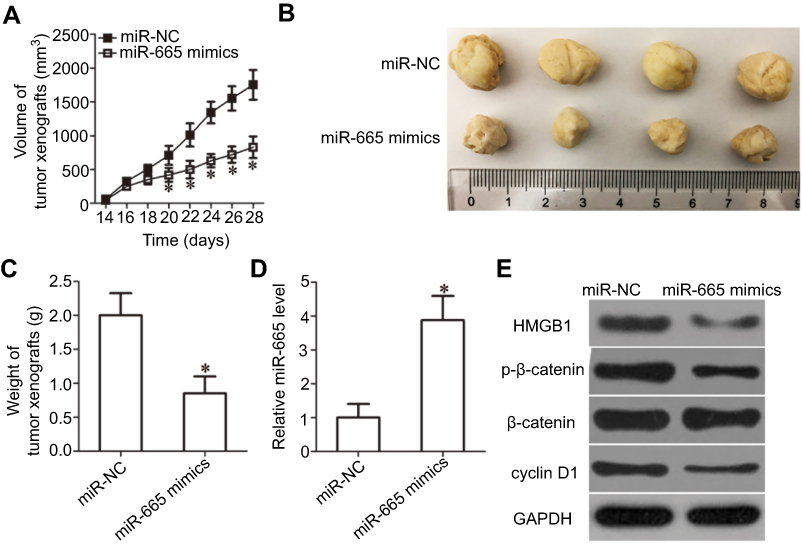 | Figure 8 miR-665 decreases HMGB1 to inhibit RB cell growth in vivo via the Wnt/β-catenin pathway. (A) The volume of tumor xenografts was measured every 2 days. The growth of the tumor xenografts in the miR-665 mimics group was notably slower than that in the miR-NC group. *P<0.05 compared with miR-NC. (B) Representative images of tumor xenografts excised from nude mice obtained 4 weeks after subcutaneous injection of Y79 cells transfected with miR-665 mimics or miR-NC. (C) All nude mice were sacrificed 30 days after injection, and the tumor xenografts were excised. The weight of the tumor xenografts in the miR-665 group was notably lower than that in the miR-NC group. *P<0.05 compared with miR-NC. (D) miR-665 expression in the tumor xenografts was analyzed by RT-qPCR. *P<0.05 compared with miR-NC. (E) The expression levels of HMGB1 and the central members of the Wnt/β-catenin pathway (p-β-catenin, β-catenin, and cyclin D1) were detected by Western blot analysis. |
Discussion
Dysregulation of miRNAs has been shown to be closely correlated with the malignant development of RB.17–19 Multiple miRNAs have been implicated in the aggressive behavior of RB by acting as tumor suppressors or oncogenes.33–35 Accordingly, further investigation of the specific miRNAs related to the development and progression of RB might facilitate the identification of promising therapeutic strategies for patients with RB. In the present study, we detected miR-665 expression in RB for the first time and subsequently investigated the roles of miR-665 in RB progression. More importantly, we explored the molecular mechanisms responsible for the tumor suppressor activity of miR-665 in RB cells.
miR-665 is upregulated in hepatocellular carcinoma, and its upregulation is significantly correlated with tumor size, vascular invasion, local invasion, Edmondson grading, and clinical stage.26,27 Hepatocellular carcinoma patients with high miR-665 levels show shorter survival times than patients with low miR-665 levels.26 In contrast, miR-665 is expressed at low levels in osteosarcoma tissues and cell lines, and osteosarcoma patients with low miR-665 expression have poorer prognoses than patients with high miR-665 expression.28 miR-665 expression is also downregulated in ovarian cancer29 and pancreatic cancer.36 However, the expression profile of miR-665 in RB had not been investigated previously. Herein, we showed that miR-665 expression was decreased in RB tissues and cell lines and that decreased miR-665 expression was markedly correlated with tumor size, TNM stage, and differentiation in patients with RB. These observations suggest miR-665 might be a novel marker of prognosis in patients with RB.
Aberrantly expressed miR-665 is involved in the malignant phenotypes of multiple human cancers. For instance, miR-665 has been identified as a tumor-promoting miRNA in hepatocellular carcinoma, and upregulation of miR-665 promotes the growth, metastasis, and epithelial-mesenchymal transition of hepatocellular carcinoma cells in vitro and in vivo.27 In contrast, miR-665 plays an inhibitory role in the growth and invasion of ovarian cancer cells.29 A study by Dong et al revealed that upregulation of miR-665 expression attenuated the proliferation, invasion, and epithelial-mesenchymal transition of osteosarcoma cells.28 However, the functional roles of miR-665 in RB cells have remained largely unknown. In the present study, exogenous miR-665 expression inhibited RB cell proliferation and colony formation, promoted cell apoptosis, and decreased cell migration and invasion in vitro. Conversely, silencing miR-665 expression showed the opposite effects. Further, in vivo experiments showed that miR-665 overexpression was sufficient to impair tumor growth. These results suggest that miR-665 is a potential therapeutic target for patients with RB.
Multiple studies have indicated that miRNAs are tightly correlated with carcinogenesis and cancer progression and function by directly regulating the expression levels of target genes.37 Three genes, PTPRB27 in hepatocellular carcinoma, RAB2328 in osteosarcoma, and homeobox A1029 in ovarian cancer, have been identified as direct targets of miR-665. Considering this, we attempted to identify the direct target gene involved in the anticancer roles of miR-665 in RB cells. HMGB1, a highly conserved DNA-binding protein, was found to be a direct and functional downstream target of miR-665 in RB. HMGB1 is located on chromosome 8q22 and has been found to be upregulated in various human malignancies, including endometrial carcinoma,38 gastric cancer,39 hepatocellular carcinoma,40 and renal cell carcinoma.41 It is also highly expressed in RB, and its overexpression is closely associated with poor tumor differentiation and optic nerve invasion.42 HMGB1 increases oncogene activity in the genesis and development of RB, and it is involved in the regulation of RB cell proliferation, autophagy, apoptosis, the cell cycle, viability, metastasis, and chemotherapy sensitivity.43–46 In the present study, we demonstrated that miR-665 directly targets HMGB1 and inactivates the Wnt/β-catenin pathway to inhibit the various malignant behaviors of RB in vitro and in vivo. These findings suggest that miR-665-mediated silencing of HMGB1 and inactivation of the Wnt/β-catenin pathway may represent an effective therapeutic approach for patients with RB.
Conclusion
In summary, this is the first study to demonstrate that miR-665 is downregulated in RB tissues and cell lines and that low miR-665 expression is significantly associated with tumor size, TNM stage, and differentiation. Upregulation of miR-665 suppresses the development and progression of RB, likely by directly targeting HMGB1 and inhibiting the activation of the Wnt/β-catenin pathway. A better understanding of the relationship between miR-665, HMGB1, and the Wnt/β-catenin pathway in RB may elucidate the molecular pathogenesis of RB and provide a potential target for therapy.
Disclosure
The authors report no conflicts of interest for this work.
References
1. Dimaras H, Kimani K, Dimba EA, et al. Retinoblastoma. Lancet. 2012;379(9824):1436–1446. doi:10.1016/S0140-6736(11)61137-9
2. MacCarthy A, Draper GJ, Steliarova-Foucher E, Kingston JE. Retinoblastoma incidence and survival in European children (1978–1997). Report from the Automated Childhood Cancer Information System project. Eur J Cancer. 2006;42(13):2092–2102. doi:10.1016/j.ejca.2006.06.003
3. Abramson DH, Shields CL, Munier FL, Chantada GL. Treatment of retinoblastoma in 2015: agreement and disagreement. JAMA Ophthalmol. 2015;133(11):1341–1347. doi:10.1001/jamaophthalmol.2015.3108
4. He MY, An Y, Gao YJ, Qian XW, Li G, Qian J. Screening of RB1 gene mutations in Chinese patients with retinoblastoma and preliminary exploration of genotype-phenotype correlations. Mol Vis. 2014;20:545–552.
5. Chantada GL, Qaddoumi I, Canturk S, et al. Strategies to manage retinoblastoma in developing countries. Pediatr Blood Cancer. 2011;56(3):341–348. doi:10.1002/pbc.22843
6. Canturk S, Qaddoumi I, Khetan V, et al. Survival of retinoblastoma in less-developed countries impact of socioeconomic and health-related indicators. Br J Ophthalmol. 2010;94(11):1432–1436. doi:10.1136/bjo.2009.168062
7. Errico A. Cancer therapy: retinoblastoma–chemotherapy increases the risk of secondary cancer. Nat Rev Clin Oncol. 2014;11(11):623. doi:10.1038/nrclinonc.2014.155
8. Shields CL, Say EA, Pointdujour-Lim R, Cao C, Jabbour PM, Shields JA. Rescue intra-arterial chemotherapy following retinoblastoma recurrence after initial intra-arterial chemotherapy. J Fr Ophtalmol. 2015;38(6):542–549. doi:10.1016/j.jfo.2015.03.004
9. Busch M, Grosse-Kreul J, Wirtz JJ, et al. Reduction of the tumorigenic potential of human retinoblastoma cell lines by TFF1 overexpression involves p53/caspase signaling and miR-18a regulation. Int J Cancer. 2017;141(3):549–560. doi:10.1002/ijc.30768
10. Chu WK, Law KS, Chan SO, et al. Antagonists of growth hormone-releasing hormone receptor induce apoptosis specifically in retinoblastoma cells. Proc Natl Acad Sci U S A. 2016;113(50):14396–14401. doi:10.1073/pnas.1617427113
11. Saliminejad K, Khorram Khorshid HR, Soleymani Fard S, Ghaffari SH. An overview of microRNAs: biology, functions, therapeutics, and analysis methods. J Cell Physiol. 2019 May;234(5):5451–5465. doi:10.1002/jcp.27486.
12. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297.
13. Chandra S, Vimal D, Sharma D, Rai V, Gupta SC, Chowdhuri DK. Role of miRNAs in development and disease: lessons learnt from small organisms. Life Sci. 2017;185:8–14. doi:10.1016/j.lfs.2017.07.017
14. Massillo C, Dalton GN, Farre PL, De Luca P, De Siervi A. Implications of microRNA dysregulation in the development of prostate cancer. Reproduction. 2017;154(4):R81–R97. doi:10.1530/REP-17-0322
15. Fang Y, Zhang L, Li Z, Li Y, Huang C, Lu X. MicroRNAs in DNA damage response, carcinogenesis, and chemoresistance. Int Rev Cell Mol Biol. 2017;333:1–49. doi:10.1016/bs.ircmb.2017.03.001
16. Mohammadi A, Mansoori B, Baradaran B. The role of microRNAs in colorectal cancer. Biomed Pharmacother. 2016;84:705–713. doi:10.1016/j.biopha.2016.09.099
17. Delsin LEA, Salomao KB, Pezuk JA, Brassesco MS. Expression profiles and prognostic value of miRNAs in retinoblastoma. J Cancer Res Clin Oncol. 2019 Jan;145(1):1–10.
18. Golabchi K, Soleimani-Jelodar R, Aghadoost N, et al. MicroRNAs in retinoblastoma: potential diagnostic and therapeutic biomarkers. J Cell Physiol. 2018;233(4):3016–3023. doi:10.1002/jcp.26070
19. Mirakholi M, Mahmoudi T, Heidari M. MicroRNAs horizon in retinoblastoma. Acta Med Iran. 2013;51(12):823–829.
20. Jin Q, He W, Chen L, Yang Y, Shi K, You Z. MicroRNA-101-3p inhibits proliferation in retinoblastoma cells by targeting EZH2 and HDAC9. Exp Ther Med. 2018;16(3):1663–1670. doi:10.3892/etm.2018.6405
21. Wu L, Chen Z, Xing Y. MiR-506-3p inhibits cell proliferation, induces cell cycle arrest and apoptosis in retinoblastoma by directly targeting NEK6. Cell Biol Int. 2018. doi:10.1002/cbin.11041
22. Zhang Y, Wang X, Zhao Y. MicroRNA874 prohibits the proliferation and invasion of retinoblastoma cells by directly targeting metadherin. Mol Med Rep. 2018;18(3):3099–3105. doi:10.3892/mmr.2018.9295
23. Yang G, Fu Y, Zhang L, Lu X, Li Q. miR106b regulates retinoblastoma Y79 cells through Runx3. Oncol Rep. 2017;38(5):3039–3043. doi:10.3892/or.2017.5931
24. Wei D, Miao Y, Yu L, Wang D, Wang Y. Downregulation of microRNA198 suppresses cell proliferation and invasion in retinoblastoma by directly targeting PTEN. Mol Med Rep. 2018;18(1):595–602. doi:10.3892/mmr.2018.8979
25. Yang L, Wei N, Wang L, Wang X, Liu QH. miR-498 promotes cell proliferation and inhibits cell apoptosis in retinoblastoma by directly targeting CCPG1. Childs Nerv Syst. 2018;34(3):417–422. doi:10.1007/s00381-017-3622-8
26. Qu Z, Wu J, Wu J, et al. Exosomal miR-665 as a novel minimally invasive biomarker for hepatocellular carcinoma diagnosis and prognosis. Oncotarget. 2017;8(46):80666–80678. doi:10.18632/oncotarget.20881
27. Hu Y, Yang C, Yang S, Cheng F, Rao J, Wang X. miR-665 promotes hepatocellular carcinoma cell migration, invasion, and proliferation by decreasing Hippo signaling through targeting PTPRB. Cell Death Dis. 2018;9(10):954. doi:10.1038/s41419-018-0978-y
28. Dong C, Du Q, Wang Z, Wang Y, Wu S, Wang A. MicroRNA-665 suppressed the invasion and metastasis of osteosarcoma by directly inhibiting RAB23. Am J Transl Res. 2016;8(11):4975–4981.
29. Liu J, Jiang Y, Wan Y, Zhou S, Thapa S, Cheng W. MicroRNA665 suppresses the growth and migration of ovarian cancer cells by targeting HOXA10. Mol Med Rep. 2018;18(3):2661–2668. doi:10.3892/mmr.2018.9252
30. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262
31. Zhou X, Hu X, Xie J, Xu C, Xu W, Jiang H. Exogenous high-mobility group box 1 protein injection improves cardiac function after myocardial infarction: involvement of Wnt signaling activation. J Biomed Biotechnol. 2012;2012:743879. doi:10.1155/2012/743879
32. Jin X, Rong S, Yuan W, et al. High mobility group box 1 promotes aortic calcification in chronic kidney disease via the Wnt/beta-catenin pathway. Front Physiol. 2018;9:665. doi:10.3389/fphys.2018.00665
33. Bu W, Wang Y, Min X. MicroRNA-106b promotes the proliferation, migration and invasion of retinoblastoma cells by inhibiting the expression of ZBTB4 protein. Exp Ther Med. 2018;16(6):4537–4545. doi:10.3892/etm.2018.6811
34. Liu Y, Li H, Liu Y, Zhu Z. MiR-22-3p targeting alpha-enolase 1 regulates the proliferation of retinoblastoma cells. Biomed Pharmacother. 2018;105:805–812. doi:10.1016/j.biopha.2018.06.038
35. Miao X, Wang Z, Chen B, et al. miR-140-5p suppresses retinoblastoma cell proliferation, migration, and invasion by targeting CEMIP and CADM3. Cell Mol Biol. 2018;64(6):42–47. doi:10.14715/cmb/2018.64.6.8
36. Zhou B, Guo W, Sun C, Zhang B, Zheng F. Linc00462 promotes pancreatic cancer invasiveness through the miR-665/TGFBR1-TGFBR2/SMAD2/3 pathway. Cell Death Dis. 2018;9(6):706. doi:10.1038/s41419-018-0724-5
37. Harries LW. Long non-coding RNAs and human disease. Biochem Soc Trans. 2012;40(4):902–906. doi:10.1042/BST20120020
38. Luan X, Ma C, Wang P, Lou F. HMGB1 is negatively correlated with the development of endometrial carcinoma and prevents cancer cell invasion and metastasis by inhibiting the process of epithelial-to-mesenchymal transition. Onco Targets Ther. 2017;10:1389–1402. doi:10.2147/OTT.S123085
39. Zhang J, Zhang R, Lu WW, et al. Clinical significance of hmgb1 expression in human gastric cancer. Int J Immunopathol Pharmacol. 2014;27(4):543–551. doi:10.1177/039463201402700410
40. Liu F, Zhang Y, Peng Z, Gao H, Xu L, Chen M. High expression of High Mobility Group Box 1 (HMGB1) predicts poor prognosis for hepatocellular carcinoma after curative hepatectomy. J Transl Med. 2012;10:135. doi:10.1186/1479-5876-10-233
41. Wu CZ, Zheng JJ, Bai YH, Xia P, Zhang HC, Guo Y. HMGB1/RAGE axis mediates the apoptosis, invasion, autophagy, and angiogenesis of the renal cell carcinoma. Onco Targets Ther. 2018;11:4501–4510. doi:10.2147/OTT.S167197
42. Singh MK, Singh L, Pushker N, et al. Correlation of High Mobility Group Box-1 protein (HMGB1) with clinicopathological parameters in primary retinoblastoma. Pathol Oncol Res. 2015;21(4):1237–1242. doi:10.1007/s12253-015-9951-6
43. Liu K, Huang J, Xie M, et al. MIR34A regulates autophagy and apoptosis by targeting HMGB1 in the retinoblastoma cell. Autophagy. 2014;10(3):442–452. doi:10.4161/auto.27418
44. Wang LL, Feng YQ, Cheng YH. Effect on proliferation and apoptosis of retinoblastoma cell by RNA inhibiting high mobility group protein box-1 expression. Int J Ophthalmol. 2017;10(1):30–34. doi:10.18240/ijo.2017.01.05
45. Chai Y, Xiao J, Zhang S, et al. High-mobility group protein B1 silencing promotes susceptibility of retinoblastoma cells to chemotherapeutic drugs through downregulating nuclear factor-κB. Int J Mol Med. 2018;41(3):1651–1658. doi:10.3892/ijmm.2018.3379
46. Liu M, Wang SM, Jiang ZX, Lauren H, Tao LM. Effects of miR-22 on viability, migration, invasion and apoptosis in retinoblastoma Y79 cells by targeting high-mobility group box 1. Int J Ophthalmol. 2018;11(10):1600–1607. doi:10.18240/ijo.2018.10.05
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.