")
Back to Journals » OncoTargets and Therapy » Volume 12
microRNA-605 inhibits the oncogenicity of non-small-cell lung cancer by directly targeting Forkhead Box P1
Received 6 November 2018
Accepted for publication 4 March 2019
Published 17 May 2019 Volume 2019:12 Pages 3765—3777
DOI https://doi.org/10.2147/OTT.S193675
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Geoffrey Pietersz
This paper has been retracted.
Wei Zhou, 1 Ruichao Li 2
1Department of Pneumology, Liyuan Hospital of Tongji Medical College of Huazhong University of Science and Technology, Wuhan, Hubei 430077, People’s Republic of China; 2Department of Gerontology, Tongji Hospital Tongji Medical College Huazhong University of Science and Technology, Wuhan, Hubei 430030, People’s Republic of China
Background and aims: microRNA-605 (miR-605) is dysregulated in multiple cancers and plays crucial roles in regulating cancer progression. However, little is known about the expression pattern and detailed roles of miR-605 in non-small-cell lung cancer (NSCLC). Thus, in this study, we evaluated miR-605 expression in NSCLC along with its clinical significance. More importantly, the detailed roles and the underlying molecular mechanisms of miR-605 in NSCLC were explored.
Material and methods: Quantitative reverse transcription polymerase chain reaction (RT-qPCR) was employed to detect miR-605 expression in NSCLC tissues and cell lines. A series of experiments were performed to determine the effects of miR-605 upregulation on NSCLC cell proliferation, apoptosis, migration and invasion in vitro and tumor growth in vivo. In addition, the downstream regulatory mechanisms of miR‐605 action in NSCLC cells were explored.
Results: Decreased expression of miR-605 was frequently detected in NSCLC tissues and cell lines. Low expression of miR-605 was significantly correlated with the tumor size, TNM stage, and distane metastasis in NSCLC patients. Exogenous miR-605 expression inhibited proliferation, increased apoptosis, and inhibited metastasis of NSCLC cells in vitro. Additionally, miR-605 overexpression hindered the growth of NSCLC cells in vivo. Furthermore, Forkhead Box P1 (FOXP1) was identified as a direct target gene of miR-605 in NSCLC cells. Moreover, FOXP1 was highly expressed in NSCLC cells and showed an inverse correlation with miR-605 expression levels. Besides, silencing of FOXP1 simulated roles similar to miR-605 upregulation in NSCLC cells. FOXP1 reintroduction partially abolished the anticancer effects of miR-605 in NSCLC cells.
Conclusion: Our results revealed that miR-605 inhibited the oncogenicity of NSCLC cells in vitro and in vivo by directly targeting FOXP1, suggesting the importance of the miR-605/FOXP1 pathway in the malignant development of NSCLC.
Keywords: non-small-cell lung cancer, microRNA-605, proliferation, apoptosis, metastasis, Forkhead Box P1
Introduction
Lung cancer is one of the most frequently diagnosed human malignancies and the leading cause of cancer-related deaths globally.1 Approximately 1.825 million newly diagnosed cases and 1.59 million mortalities caused by lung cancer are predicted every year all over the world.2 Lung cancer could be divided into two main subtypes, non-small-cell lung cancer (NSCLC) and small-cell lung cancer, which account for 85% and 15% of all lung cancer cases, respectively.3 Currently, the primary therapy for NSCLC is surgical resection, followed by chemotherapy, radiotherapy, and other anti-tumor therapy.4 With the notable development of therapeutic strategies in recent years, the treatment outcomes of patients with all stages of NSCLC have improved to a certain extent; however, the curative effects of cancer therapy remain unsatisfactory.5 Therefore, an in-depth elucidation of the mechanisms underlying the occurrence and development of NSCLC is imperative to the development of novel and effective therapeutic targets for patients with this fatal disease.
microRNAs (miRNAs) are a group of non-coding, short RNA molecules made up of 17–24 nucleotides.6 miRNAs function as endogenous RNA silencing molecules through complementary binding to the 3′-untranslated regions (3′-UTRs) of their target genes, resulting in mRNA degradation or promoting gene silencing by suppressing translation.7 Emerging studies have demonstrated that the expression of miRNA molecules is abnormal in most human malignancies and they play crucial roles in tumorigenesis and tumor development.8–10 A variety of miRNAs have been reported to be aberrantly expressed in NSCLC.11 For example, miR-212,12 miR-409,13 and miR-431714 are downregulated in NSCLC, whereas miR-21,15 miR-105,16 and miR-42117 are expressed at high levels. Dysregulation of miRNA is closely related with the progression and development of NSCLC through participating in the regulation of multiple biological behaviors, thus acting either as oncogenes or tumor suppressors.18 Hence, identifying new miRNAs and investigating their roles in NSCLC progression might facilitate the identification of novel therapeutic targets for treating patients with NSCLC in the future.
miR-605 is dysregulated in multiple cancers and play crucial roles in regulating cancer progression.19–21 However, little is known about the expression pattern and detailed roles of miR-605 in NSCLC. Thus, in this study, we detected the expression of miR-605 in NSCLC and investigated its clinical significance. More importantly, the detailed roles and the underlying molecular mechanisms of miR-605 in NSCLC were elucidated.
Materials and methods
Tissue specimens
Paired NSCLC tissues and adjacent normal lung tissues were collected from 45 patients who underwent surgical resection at Liyuan Hospital of Tongji Medical College of Huazhong University of Science and Technology between October 2015 and November 2017. None of the patients had received chemotherapy, radiotherapy or other anti-tumor therapy before surgical resection. Tissue specimens were resected, frozen in liquid nitrogen, and then stored at −80 °C until RNA extraction. This research was approved by the Ethics Committee of Liyuan Hospital of Tongji Medical College of Huazhong University of Science and Technology, and was performed in accordance with the Declaration of Helsinki. In addition, written informed consent was obtained from all participants prior to their enrolment in this study.
Cell lines and culture conditions
A non-tumorigenic bronchial epithelium cell line BEAS-2B and four human NSCLC cell lines (H460, SK-MES-1, H522, and H1299) were purchased from Shanghai Institute of Biochemistry and Cell Biology (Shanghai, China). The cells were routinely cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum (FBS) (both from Gibco, Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 1% v/v penicillin-streptomycin (Sigma-Aldrich, St. Louis, MO, USA). All cells were cultivated in a humidified incubator at 37 °C supplied with 5% CO2.
Cell transfection experiments
miR-605 mimics and negative control miRNA mimics (miR-NC) were ordered from Guangzhou RiboBio Co., Ltd (Guangzhou, China). Small interfering RNA (siRNA) targeting FOXP1 expression (si-FOXP1) and negative control siRNA (si-NC) were chemically synthesized by GenePharma Co. Ltd. (Shanghai, China). FOXP1 overexpression vector pCMV-FOXP1 and pCMV empty plasmid was generated by GeneCopoeia Co. Ltd. (Guangzhou, China). For transfection, cells were plated into 6-well plates with a density of 5×105 cells per well and allowed to adhere overnight. Lipofectamine™ 2000 (Invitrogen, Thermo Fisher Scientific, Inc., Waltham, MA, USA) was used for cell transfection, according to the manufacturer’s instructions. Cells were collected at different times after transfection and used for further analysis.
RNA extraction and quantitative reverse transcription polymerase chain reaction (RT-qPCR)
Total RNA was isolated from tissue specimens or cultured cells using TRIzol reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA) following the manufacturer’s protocol. The concentration of total RNA was determined using NanoDrop ND-1000 (Thermo Fisher Scientific, Inc., Waltham, MA, USA). For quantification of miR-605 expression, total RNA was reverse transcribed to cDNA using a miScript Reverse Transcription kit (Qiagen GmbH, Hilden, Germany). Next, quantitative PCR (qPCR) was performed using a miScript SYBR Green PCR kit (Qiagen GmbH, Hilden, Germany). To analyze FOXP1 mRNA expression, total RNA was reverse transcribed into cDNA using a PrimeScript RT Reagent Kit (TaKaRa, Tokyo, Japan). The PCR amplification for quantifying the expression of FOXP1 mRNA was carried out using a SYBR Premix Ex Taq (TaKaRa, Tokyo, Japan). U6 small nuclear RNA and GAPDH served as internal controls for miR-605 and FOXP1 mRNA levels, respectively. The 2−∆∆Cq method was used to calculate the relative gene expression.22
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) assay
Transfected cells were harvested after 24 h of incubation and inoculated into 96-well plates with a density of 2,500 cells/well. Cell proliferative potential was evaluated by an MTT assay at 0, 24, 48 and 72 h after inoculation. Briefly, 20 µl of MTT solution (5 mg/mL, Sigma-Aldrich, St. Louis, MO, USA) was added into each well prior to incubation at 37 °C with 5% CO2 for 4 h. Subsequently, the culture medium was replaced with 100 μl of dimethyl sulfoxide (DMSO). After the formazan was solubilized, the absorbance at 490 nm was detected using a microplate reader (Model 550, Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Cell apoptosis assay
Annexin V–fluorescein isothiocyanate (FITC) apoptosis detection kit (Biolegend, San Diego, CA, USA) was used to detect the percentage of apoptotic cells. In detail, transfected cells were collected by trypsinization, washed with ice-cold phosphate buffer saline (PBS) (both from Gibco, Thermo Fisher Scientific, Inc., Waltham, MA, USA) and then suspended in 100 µl of binding buffer. A volume of 5 µl of Annexin V-FITC and 5 µl of propidium iodide were added to the cell suspension and the cells were further incubated at room temperature for 15 min in darkness. Finally, a flow cytometer (FACScan™, BD Biosciences, Franklin Lakes, NJ, USA) was utilized to determine the cell apoptotic rate.
Transwell migration and invasion assays
Transwell inserts (Costar, Cambridge, MA, USA) pre-coated with Matrigel (BD Biosciences, Franklin Lakes, NJ, USA) on the upper surface were used for invasion assay or without Matrigel for migration assay. Following transfection for 48 h, cells were harvested and suspended in FBS-free DMEM. In total, 5×104 cells were plated into the upper compartments of the Transwell inserts and 200 µl of DMEM supplemented with 20% FBS was added in the lower compartments as a chemoattractant. After 24 h of incubation, the cells remaining in upper compartments were wiped using a cotton swab. Cells on the underside of the inserts were fixed with 4% paraformaldehyde, stained with 0.05% crystal violet, washed with PBS and air-dried. The number of cells that had passed through the membrane was counted under an inverted microscope (Olympus IX83; Olympus Corporation, Tokyo, Japan) and photographed.
Tumor xenograft assay
All animal experiments were approved by the Ethics Review Committee of Liyuan Hospital of Tongji Medical College of Huazhong University of Science and Technology, and were conducted in accordance with the Animal Protection Law of the People’s Republic of China-2009 for experimental animals. Animals were maintained following the guidelines for use and care of laboratory animals. 4–6-week-old BALB/c nude mice were purchased from Shanghai Laboratory Animal Center (Chinese Academy of Sciences, Shanghai, China). A total of 2×106 cells transfected with miR-605 mimics or miR-NC were subcutaneously injected into the rear flank of nude mice. The tumor width and length were measured every four days and the tumor volume was analyzed using the formula V (mm3) = width2 (mm2) × length (mm)/2. One month later, all nude mice were executed, and the xenografts were dissected out and weighed.
Target prediction and luciferase reporter assay
The putative targets of miR-605 were predicted using three miRNA target prediction software, including miRDB (
The 3ʹ-UTR fragments of FOXP1 containing the wild-type miR-605 binding sequences and mutant FOXP1 3ʹ-UTR were amplified by GenePharma Co. Ltd., and cloned into the pMIR-REPORT vector (Promega, Madison, WI, USA). These reporter plasmids were designated as wild-type pMIR-FOXP1-3ʹ-UTR and mutant pMIR-FOXP1-3ʹ-UTR, respectively. Cells were inoculated into 24-well plates one day prior to transfection. miR-605 mimics or miR-NC, in combination with wild-type pMIR-FOXP1-3ʹ-UTR or mutant pMIR-FOXP1-3ʹ-UTR, was introduced into cells using Lipofectamine™ 2000, in accordance with the manufacturer’s instructions. At 48 h after treatment, transfected cells were harvested and luciferase activity was detected using a Dual-Luciferase Reporter Assay system (Promega, Madison, WI, USA). Firefly luciferase activity was normalized against Renilla luciferase activity.
Western blot analysis
Cells or homogenized tissues were lysed using radioimmunoprecipitation assay lysis buffer (Beyotime Institute of Biotechnology, Shanghai, China) containing a protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO, USA). The concentration of total protein was quantified using a BCA Protein Assay Reagent kit (Pierce, Thermo Fisher Scientific, Inc., Waltham, MA, USA) according to the manufacturer’s protocol. Equal amounts of total protein were separated using a 10% SDS-PAGE gel and then transferred onto PVDF membranes (EMD Millipore, Billerica, MA, USA).
Subsequent to blocking at room temperature with 5% fat-free dried milk diluted in Tris-buffered saline with 0.1% Tween-20 (TBST) for 2 h, the membranes were incubated with the primary antibodies overnight at 4 °C. The primary antibodies used in this study were as follows: rabbit anti-human FOXP1 antibody (1:1,000, cat. no. ab196978) and rabbit anti-human GAPDH antibody (1:1,000, cat. no. ab181603, both from Abcam, Cambridge, UK). Horseradish peroxidase-conjugated goat anti-rabbit secondary antibody (cat. no. ab205718, Abcam, Cambridge, UK) was used at a dilution of 1:5,000 for 2 h at room temperature and the protein signals were detected using a Pierce ECL Western Blotting Substrate (Pierce, Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Statistical analysis
All results from at least three independent experiments were shown as mean ± standard deviation and analyzed using SPSS software, v. 19.0 for Windows (IBM Corp. Armonk, NY, USA). Student’s Two-tailed t-test was used for comparison of two treatment groups. One-way analysis of variance followed by the Bonferroni post hoc test was performed to evaluate the differences between multiple groups. Chi-square test was employed to determine the correlation of expression of miR-605 and clinico-pathological characteristics in NSCLC patients. The association between FOXP1 mRNA and miR-605 expression levels was analyzed by Spearman’s correlation analysis. P<0.05 was considered to indicate a statistically significant difference.
Results
miR-605 expression is decreased in NSCLC tissues and cell lines
To examine the expression status of miR-605 in NSCLC, RT-qPCR was performed to detect miR-605 expression in 45 pairs of NSCLC tissues and adjacent normal lung tissues. The results showed that the expression level of miR-605 was notably lower in NSCLC tissues than that in adjacent normal lung tissues (Figure 1A, P<0.05). We next explored the relationship between miR-605 expression and clinico-pathological characteristics, to clarify the clinical value of miR-605 in NSCLC patients. Chi-square test indicated that decreased expression of miR-605 was significantly correlated with tumor size (P=0.026), TNM stage (P=0.004), and distant metastasis (P=0.023), but not with the gender, age, histological tumor type and tumor differentiation (all P>0.05; Table 1). Furthermore, the data obtained from RT-qPCR analysis revealed that miR-605 was frequently downregulated in all four NSCLC cell lines (H460, SK-MES-1, H522, and H1299) relative to its expression in a non-tumorigenic bronchial epithelium cell line BEAS-2B (Figure 1B, P<0.05). Collectively, these observations strongly implied that a change in miR-605 expression might be related to the progression of NSCLC.
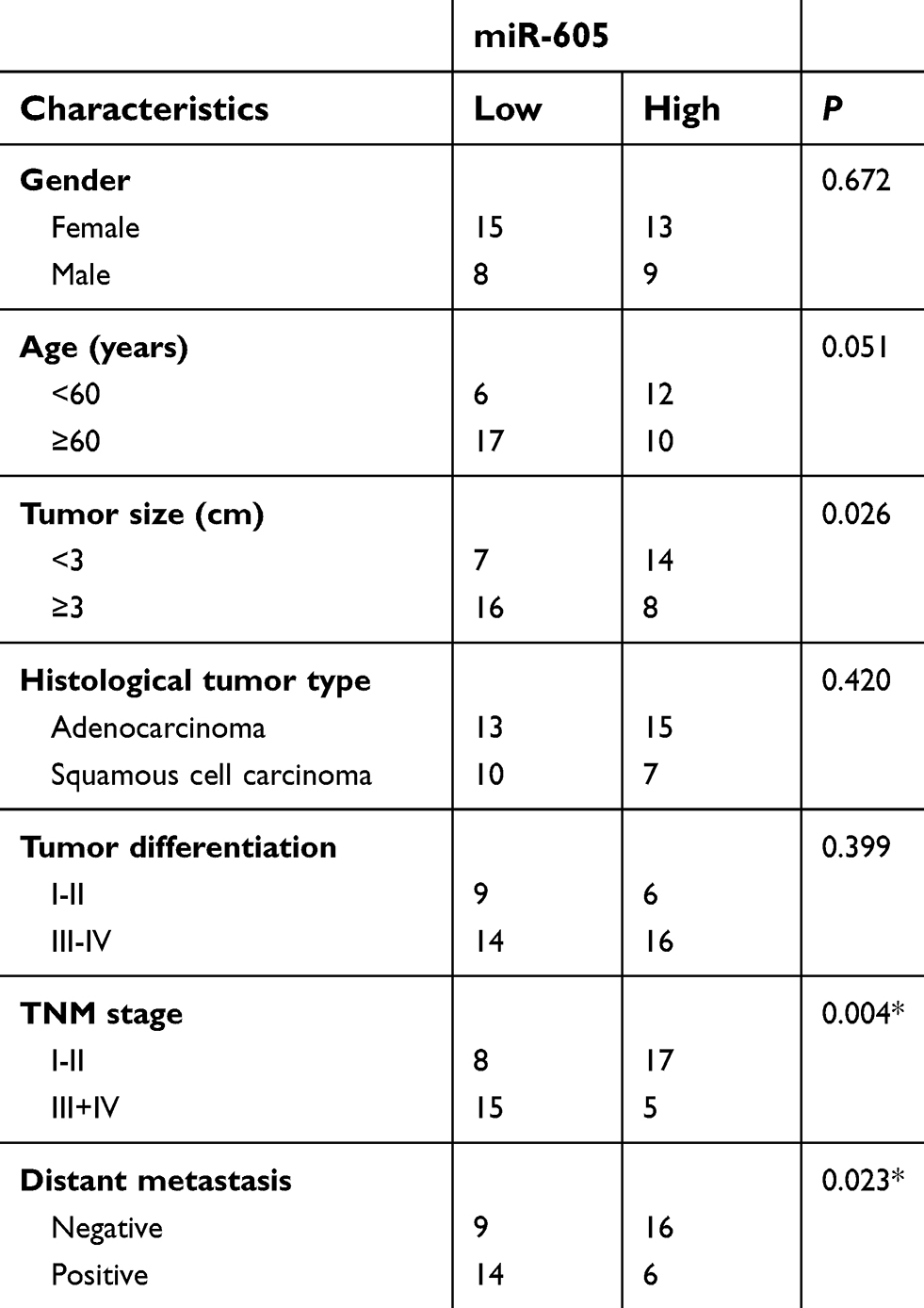 | Table 1 Associations between miR-605 expression and clinic-pathological characteristics in NSCLC patients |
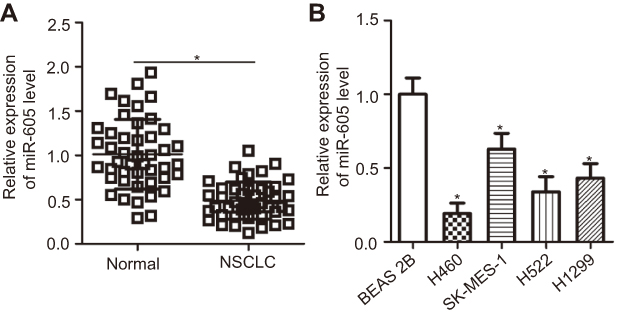 | Figure 1 miR-605 is downregulated in NSCLC tissues and cell lines. (A) The expression of miR-605 in 45 pairs of NSCLC tissues and adjacent normal lung tissues was detected using RT-qPCR. *P<0.05 vs normal lung tissues. (B) RT-qPCR was performed to determine miR-605 expression in a total of four NSCLC cell lines (H460, SK-MES-1, H522 and H1299). Non-tumorigenic bronchial epithelium cell line BEAS-2B was used as a control. *P<0.05 vs BEAS-2B. |
miR-605 inhibits the malignant phenotype of NSCLC in vitro
H460 and H522 cell lines showed relatively lower miR-605 expression among the four NSCLC cell lines and hence, these two NSCLC cell lines were selected for further functional assays. To illustrate the role of miR-605 in the aggressiveness of NSCLC cells, H460 and H522 cells were transfected with miR-605 mimics in order to increase endogenous miR-605 expression (Figure 2A, P<0.05). miR-NC served as a control for miR-605. MTT assay was performed to detect cellular proliferation, and it was demonstrated that the proliferative ability of H460 and H522 cells transfected with miR-605 mimics was significantly decreased in comparison to cells transfected with miR-NC (Figure 2B, P<0.05). In addition, the effect of upregulated miR-605 in the apoptosis of NSCLC cells was determined via cell apoptosis assay. As expected, ectopic miR-605 expression enhanced the percentages of apoptotic cells in H460 and H522 cell lines (Figure 2C, P<0.05). Transwell migration and invasion assays were further performed to explore whether miR-605 might affect the migration and invasion of NSCLC cells. The results showed that overexpression of miR-605 significantly decreased the migratory (Figure 2D, P<0.05) and invasive (Figure 2E, P<0.05) capacities of H460 and H522 cells compared to the miR-NC group. These results suggested that miR-605 might exert an inhibitory role in NSCLC cell growth and metastasis, in vitro.
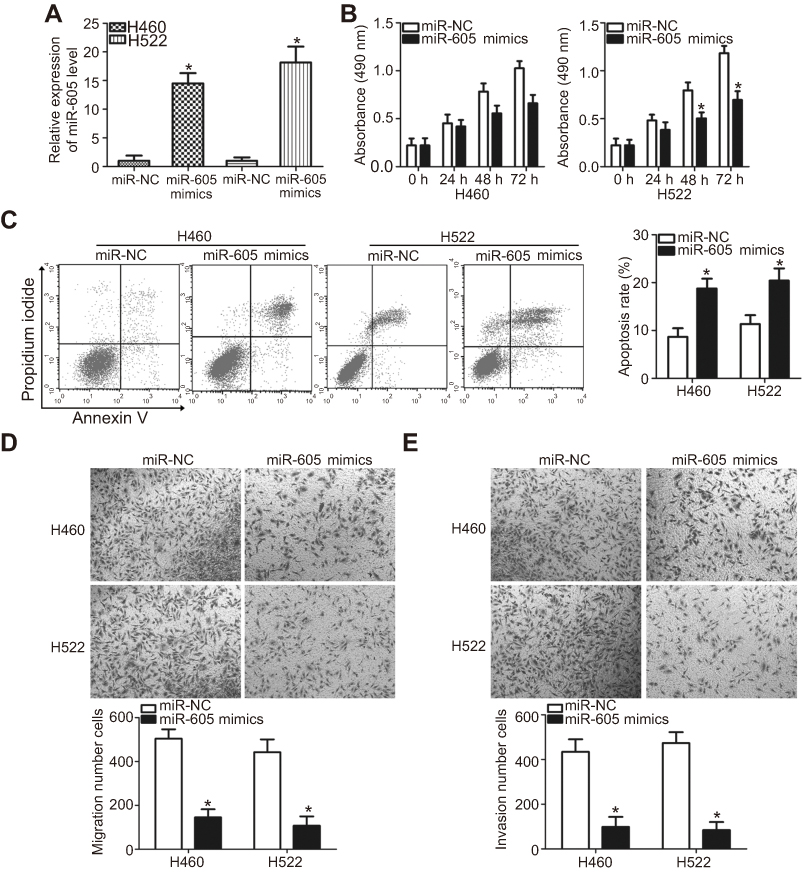 | Figure 2 miR-605 exerts tumor-suppressing roles in NSCLC cells. (A) Quantitation of the expression of miR-605 through RT-qPCR in H460 and H522 cells after transfection with miR-605 mimics or miR-NC . *P<0.05 vs miR-NC. (B) MTT assay showed the proliferative ability of H460 and H522 cells transfected with miR-605 mimics or miR-NC. *P<0.05 vs miR-NC. (C) The percentage of apoptotic H460 and H522 cells with indicated treatments was evaluated by cell apoptosis assay. *P<0.05 vs miR-NC. (D, E) Representative images and quantitation of the transwell migration and invasion assays carried out in H460 and H522 cells that were treated with miR-605 mimics or miR-NC. *P<0.05 vs miR-NC. |
FOXP1 is a direct target gene of miR-605 in NSCLC cells
To elucidate the downstream regulatory mechanism of action of miR-605 in NSCLC cells, bioinformatics analysis was performed to search for the putative target of miR-605. Based on the results, FOXP1 (Figure 3A) ignited our interest since this gene plays crucial roles in the progression and development of NSCLC.23 Luciferase reporter assay was performed to determine whether miR-605 was able to directly target the 3′-UTR of FOXP1 in NSCLC cells. Restoring the expression of miR-605 significantly decreased the luciferase activity of wild-type pMIR-FOXP1-3ʹ-UTR in both H460 and H522 cells (P<0.05), whereas the inhibitory effect was abrogated when the binding sequences of miR-605 in the 3ʹ-UTR of FOXP1 were mutated (Figure 3B). To further confirm whether FOXP1 is a direct target of miR-605, we detected its expression in NSCLC tissues and examined its relationship with miR-605. RT-qPCR analysis demonstrated that both FOXP1 mRNA (Figure 3C, P<0.05) and protein (Figure 3D, P<0.05) levels were notably elevated in NSCLC tissues compared to that in adjacent normal lung tissues. Furthermore, an inverse association between miR-605 and FOXP1 mRNA levels was identified in NSCLC tissues (Figure 3E; r=−0.5480, P<0.0001). Moreover, decreased FOXP1 mRNA (Figure 3F, P<0.05) and protein (Figure 3G, P<0.05) levels were observed in miR-605-overexpressing H460 and H522 cells. Therefore, we drew a conclusion that FOXP1 is the direct target gene of miR-605 in NSCLC cells.
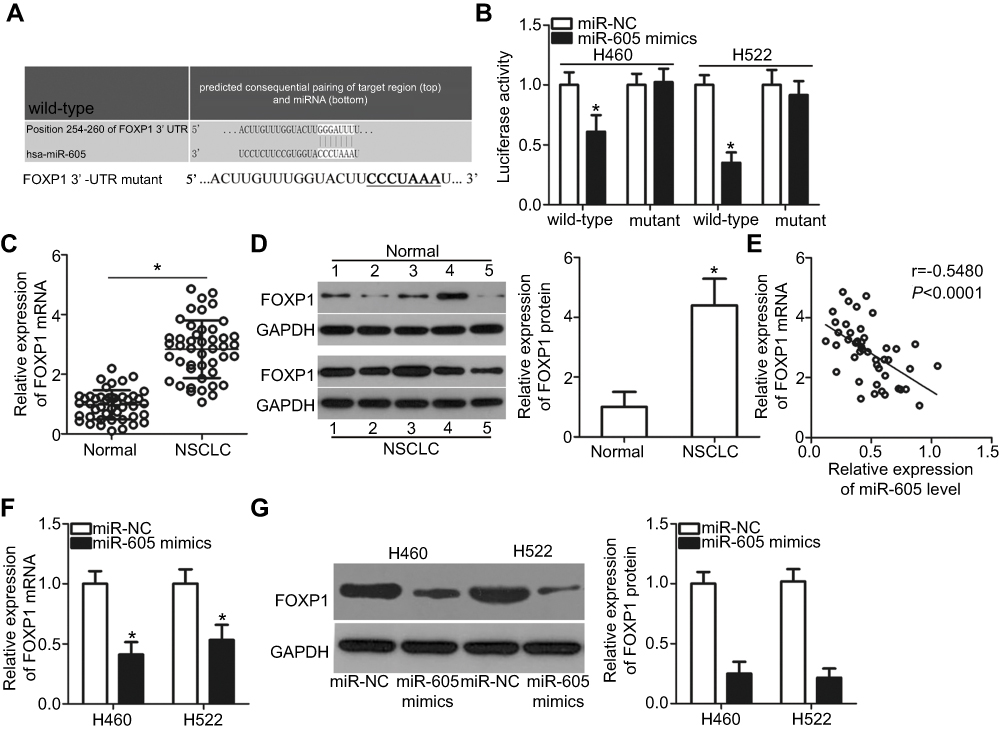 | Figure 3 FOXP1 is a direct target gene of miR-605 in NSCLC cells. (A) Comparison of sequences of miR-605 with wild-type or mutant putative binding sites in the 3′-UTR of FOXP1 gene. (B) Luciferase reporter assay was employed to determine that miR-605 directly targets the 3ʹ-UTR of FOXP1. Luciferase activity was detected in H460 and H522 cells co-transfected with miR-605 mimics or miR-NC and wild-type pMIR-FOXP1-3ʹ-UTR or mutant pMIR-FOXP1-3ʹ-UTR. *P<0.05 vs miR-NC. (C, D) The FOXP1 mRNA and protein levels in NSCLC tissues and adjacent normal lung tissues were detected via RT-qPCR and western blot analysis, respectively. *P<0.05 vs normal lung tissues. (E) Spearman’s correlation analysis showed an inverse correlation between miR-605 and FOXP1 mRNA levels in NSCLC tissues. r=−0.5480, P<0.0001. (F, G) RT-qPCR and western blot analysis was adopted to measure the mRNA and protein levels of FOXP1 in H460 and H522 cells transfected with miR-605 mimics or miR-NC. *P<0.05 vs miR-NC. |
FOXP1 inhibition shows effects similar to miR-605 overexpression in NSCLC cells
To determine the role of FOXP1 in the malignant progression of NSCLC, small interfering RNA targeting FOXP1 expression (si-FOXP1) and negative control siRNA (si-NC) were transfected into H460 and H522 cells. The protein level of FOXP1 was notably downregulated in H460 and H522 cells after si-FOXP1 transfection, as measured by western blot analysis (Figure 4A, P<0.05). MTT and cell apoptosis assays revealed that knockdown of FOXP1 restricted proliferation (Figure 4B, P<0.05) and promoted apoptosis (Figure 4C, P<0.05) of H460 and H522 cells compared to cells transfected with si-NC. Furthermore, the migration (Figure 4D, P<0.05) and invasion (Figure 4E, P<0.05) of H460 and H522 cells was suppressed by FOXP1 knockdown. These results indicated that FOXP1 silencing exerted an impact in NSCLC cells similar to that caused by miR-605 overexpression, further suggesting that FOXP1 is a direct target of miR-605.
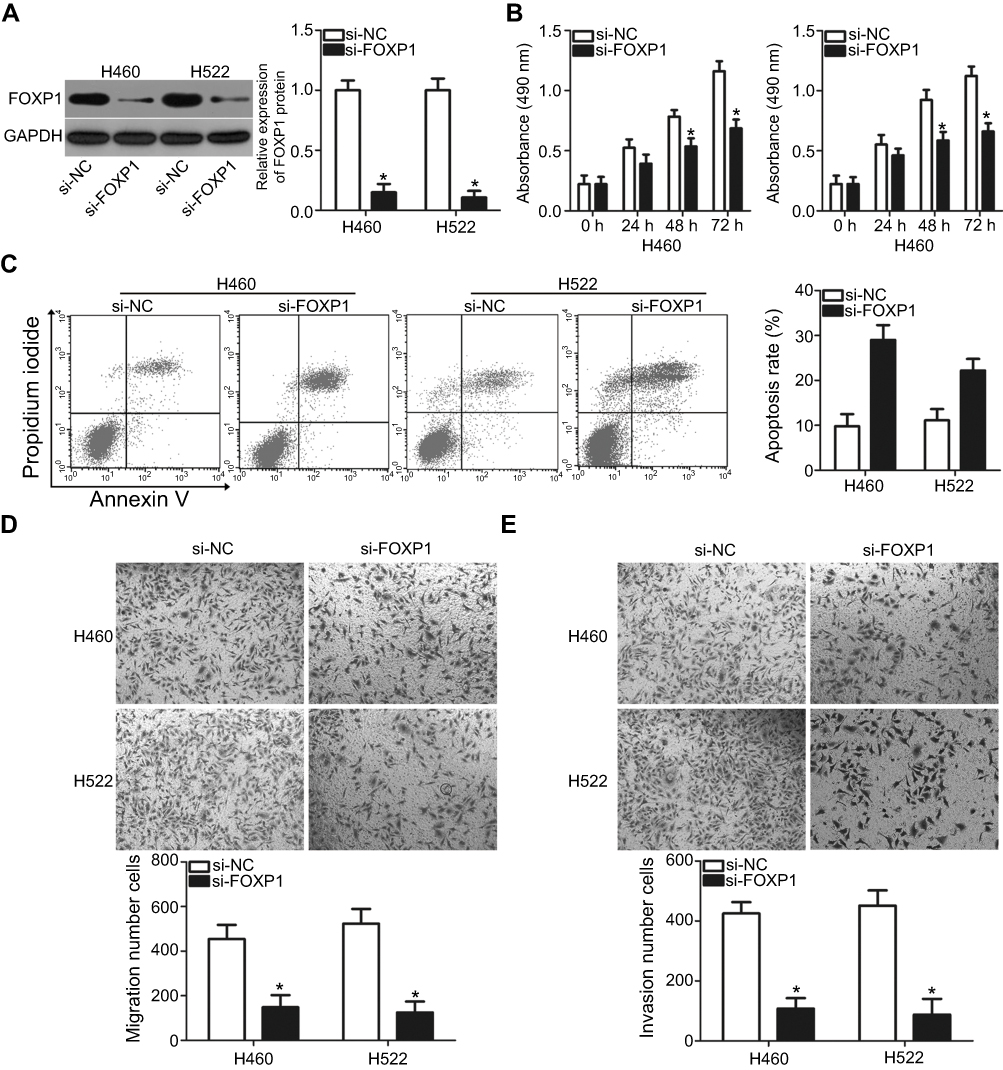 | Figure 4 Inhibition of FOXP1 is able to imitate the anticancer effects of miR-605 upregulation in NSCLC cells. (A) H460 and H522 cells were transfected with si-FOXP1 or si-NC. After 72 h of transfection, knockdown of FOXP1 was efficient in H460 and H522 cells, as demonstrated by western blot analysis. *P<0.05 vs si-NC. (B, C) MTT and cell apoptosis assays were conducted to examine the effects of FOXP1 silencing on NSCLC cell proliferation and apoptosis, respectively. *P<0.05 vs si-NC. (D, E) The migratory and invasive capacities of H460 and H522 cells after si-FOXP1 or si-NC transfection were evaluated through transwell migration and invasion assays. *P<0.05 vs si-NC. |
FOXP1 restoration abrogates the antitumor effects of miR-605 in NSCLC cells
A series of rescue experiments were further performed to verify whether FOXP1 contributes to miR-605-mediated tumor-suppressing activity in NSCLC cells. To this end, miR-605-overexpressing H460 and H522 cells were further transfected with FOXP1 overexpression vector pCMV-FOXP1 or pCMV empty plasmid. Transfection of pCMV-FOXP1 partially restored the decreased FOXP1 protein level in miR-605 mimics-transfected H460 and H522 cells (Figure 5A, P<0.05). As expected, the restored FOXP1 expression reversed the suppressive effects of miR-605 overexpression in H460 and H522 cells, with respect to proliferation (Figure 5B, P<0.05), apoptosis (Figure 5C, P<0.05), migration (Figure 5D, P<0.05) and invasion (Figure 5E, P<0.05), in vitro. Overall, these results clearly demonstrated that miR-605 conferred its antitumor effect in NSCLC cells by directly targeting and downregulating FOXP1 and the downregulation of FOXP1 by miR-605 was essential for the tumor suppressive roles of miR-605 in NSCLC cells.
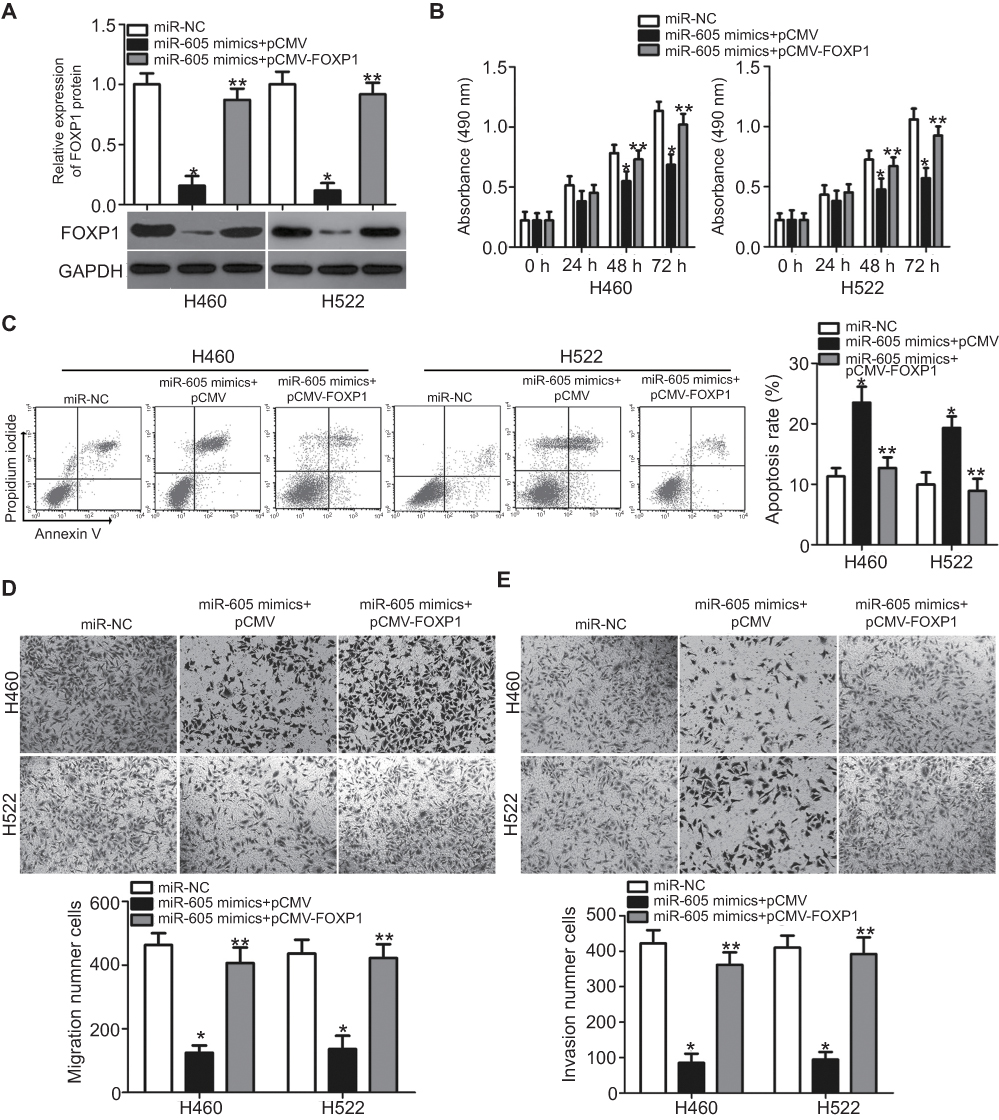 | Figure 5 Restoration of FOXP1 expression reverses the tumor-suppressing effects of miR-605 in NSCLC cells. miR-605 mimics in combination with pCMV-FOXP1 or pCMV was co-transfected into H460 and H522 cells. (A) Western blot analysis was performed at 72 h post-transfection to measure FOXP1 protein expression. *P<0.05 vs miR-NC. **P<0.05 vs miR-605 mimics + pCMV. (B–E) The proliferation, apoptosis, migration and invasion of above mentioned cells was determined through MTT, cell apoptosis, transwell migration and invasion assays, respectively. *P<0.05 vs miR-NC. **P<0.05 vs miR-605 mimics + pCMV. |
Overexpression of miR-605 hinders tumor growth of NSCLC cells in vivo
Tumor xenograft assay was performed to assess the effect of miR-605 overexpression on the growth of NSCLC cells in vivo. H460 cells were transfected with miR-605 mimics or miR-NC. Cells were collected after 24 h of transfection and subcutaneously injected into nude mice to generate transplanted tumors of BALB/c nude mice. The tumor volume was measured every four days. One month after implantation, the xenografts were dissected and the exact volume and weights were evaluated. The tumor volume (Figure 6A) and weights (Figure 6B, P<0.05) of the xenografts from mice injected with miR-605 mimics were significantly suppressed relative to those that received the miR-NC. The tumor growth curve indicated an obvious suppression in the miR-605 mimics group compared to the miR-NC group (Figure 6C, P<0.05). Meanwhile, RT-qPCR analysis was carried out to detect the expression of miR-605 in the xenografts. The data showed that the expression level of miR-605 was higher in the xenografts derived from the miR-605-expressing H460 cells (Figure 6D, P<0.05). Moreover, there was a significant decrease in FOXP1 protein expression in the miR-605 mimics group compared to that in the miR-NC group (Figure 6E). These observations demonstrated that miR-605 inhibits the growth of NSCLC cells, in vivo.
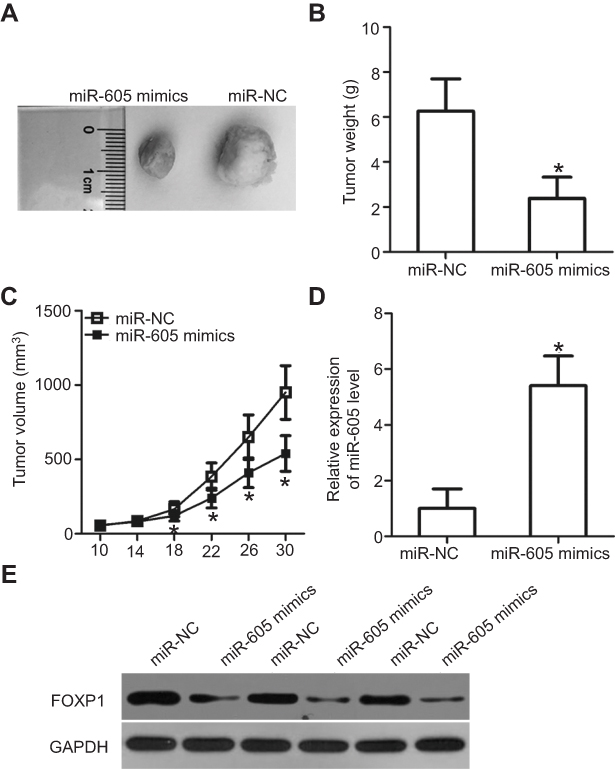 | Figure 6 miR-605 suppresses tumor growth of NSCLC cells in vivo. (A) Photographs of tumor xenografts were obtained from nude mice that were injected with miR-605 mimics or miR-NC transfected H460 cells. (B) The weight of tumor xenografts in miR-605 mimics and miR-NC groups was detected. *P<0.05 vs miR-NC. (C) The tumor volume was determined using the formula V (mm3) = width2 (mm2) × length (mm)/2. Growth curve of tumor volumes was calculated. *P<0.05 vs miR-NC. (D) Expression level of miR-605 in xenografts was detected by RT-qPCR. *P<0.05 vs miR-NC. (E) FOXP1 protein expression in the tumor xenografts was assessed by western blot analysis. |
Discussion
Recently, the changes in miRNA expression are currently a hot research area.24,25 The dysregulation of miRNAs in NSCLC has been widely reported in accumulating studies.26–28 Aberrantly expressed miRNAs are closely correlated with the malignant progression of NSCLC and participate in the regulation of various biological behaviors.11 Hence, in-depth studies of the effects of crucial miRNAs in NSCLC progression might provide a novel insight into their use as potential therapeutic targets for treating patients with this deadly disease. This study, for the first time, detected miR-605 expression in NSCLC, investigated the regulatory roles of this miRNA with respect to the aggressive behaviors of NSCLC and explored the possible underlying mechanisms.
miR-605 has been well-studied in multiple types of malignant tumors. For example, miR-605 is downregulated in melanoma tissues and cell lines. Restoring miR-605 expression decreased melanoma cell growth in vitro and in vivo.19 Expression level of miR-605 was also lower in prostate cancer tissues and cell lines. The upregulation of miR-605 inhibited the proliferation and invasion of prostate cancer cells.20 A study by Li et al revealed that miR-605 expression was decreased in intrahepatic cholangiocarcinoma. Restoration of miR-605 expression restricted cell progression in vitro and in vivo.21 However, the expression pattern and specific roles of miR-605 in NSCLC remain to be elucidated. Herein, RT-qPCR analysis indicated that miR-605 was expressed at low levels in NSCLC tissues and cell lines. Low expression of miR-605 was observed to be correlated with tumor size, TNM stage and distant metastasis in NSCLC patients. Functionally, miR-605 overexpression was able to inhibit the proliferation, induce apoptosis and suppress metastasis of NSCLC cells in vitro, as well as decrease tumor growth in vivo. These findings suggest that miR-605 might be a potential diagnostic biomarker and therapeutic target for patients with the above specific cancer type.
Three human genes, including INPP4B in melanoma,19 EN2 in prostate cancer,20 and PSMD10/Gankyrin in intrahepatic cholangiocarcinoma,21 have been demonstrated to be direct targets of miR-605. Hence, we next attempted to investigate the underlying mechanisms by which miR-605 might affect the oncogenicity of NSCLC cells. First, FOXP1 was predicted to be a potential target of miR-605, by all three miRNA target prediction software. Second, miR-605 could directly bind to the 3ʹ-UTR of FOXP1 in NSCLC cells, as demonstrated by luciferase reporter assay. Third, highly expressed FOXP1 in NSCLC tissues was inversely correlated with miR-605 expression. Fourth, the mRNA and protein levels of FOXP1 were notably downregulated in NSCLC cells upon miR-605 upregulation. Fifth, inhibition of FOXP1 exhibited similar effects on miR-605 in NSCLC cells. Finally, FOXP1 restoration partially attenuated the suppression phenotype driven by miR-605 upregulation in NSCLC cells. These results provided unequivocal evidence to support that miR-605 suppressed the aggressive progression of NSCLC by directly targeting FOXP1 and that downregulation of FOXP1 by miR-605 was essential for miR-605-induced antitumor roles in NSCLC.
FOXP1 was first identified by Shu et al,29 and it was considered as a glutamine rich factor. FOXP1 is a member of the forkhead box transcription factor family.30 It is a transcription inhibitor and has been reported to be dysregulated in multiple human cancers.31–33 One previous study reported that FOXP1 was upregulated in NSCLC at both mRNA and protein levels.23 High FOXP1 expression was significantly correlated with gender and histologic type in NSCLC patients. These patients with high FOXP1 expression had shorter five-year survival rate than patients with low FOXP1 expression.23 Furthermore, Kaplan-Meier survival and cox regression analyses identified FOXP1 expression as an independent biomarker to predict the poor prognosis of patients with NSCLC.23 In this study, we revealed that inhibition of FOXP1 suppressed NSCLC cell proliferation, promoted cell apoptosis, and decreased cell migration and invasion, in vitro. Notably, miR-605 directly targeted FOXP1, thereby inhibiting the malignant progression of NSCLC cells. Accordingly, targeting FOXP1 by restoring the expression of miR-605 might be an effective therapeutic approach for NSCLC patients.
Conclusion
To our knowledge, this is the first study to confirm that the downregulation of miR-605 is a common phenomenon in NSCLC tissues and cell lines and that this downregulation of miR-605 is significantly correlated with the tumor size, TNM stage, and distant metastasis. In addition, increased miR-605 expression can prohibit the progression of NSCLC in vitro and in vivo by directly targeting FOXP1. These findings might provide a new insight into NSCLC carcinogenesis and miR-605 could be developed as a potential therapeutic target for this cancer type.
Abbreviation list
NSCLC, Non-small-cell lung cancer; RT-qPCR, Quantitative reverse transcription-quantitative polymerase chain reaction; miRNA, microRNA; DMEM, Dulbecco’s modified Eagle’s medium; FBS, Fetal bovine serum; miRNA, microRNA; miR-NC, negative control miRNA mimics; siRNA, Small interfering RNA; si-NC, negative control siRNA; MTT, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide; DMSO, dimethyl sulfoxide; FITC, Annexin V–fluorescein isothiocyanate; PBS, phosphate buffer saline; TBST, Tris-buffered saline with 0.1% Tween-20; 3ʹ-UTR, 3ʹ-Untranslated region.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21262
2. Ferlay J, Soerjomataram I, Dikshit R, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136(5):E359386. doi:10.1002/ijc.29210
3. Torre LA, Sauer AM, Chen MS
4. Burdett S, Pignon JP, Tierney J, et al. Adjuvant chemotherapy for resected early-stage non-small cell lung cancer. Cochrane Database Syst Rev. 2015;(3):CD011430.
5. Thomas A, Liu SV, Subramaniam DS, Giaccone G. Refining the treatment of NSCLC according to histological and molecular subtypes. Nat Rev Clin Oncol. 2015;12(9):511–526. doi:10.1038/nrclinonc.2015.90
6. Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer. 2006;6(11):857–866. doi:10.1038/nrc1997
7. Behm-Ansmant I, Rehwinkel J, Izaurralde E. MicroRNAs silence gene expression by repressing protein expression and/or by promoting mRNA decay. Cold Spring Harb Symp Quant Biol. 2006;71:523–530. doi:10.1101/sqb.2006.71.013
8. Dias F, Morais M, Teixeira AL, Medeiros R. Involving the microRNA targetome in esophageal-cancer development and behavior. Cancers. 2018;10(10). doi:10.3390/cancers10110400
9. Zhu Z, Zhang X, Wang G, Zheng H. Role of MicroRNAs in hepatocellular carcinoma. Hepat Mon. 2014;14(8):e18672. doi:10.5812/hepatmon.18672
10. Yu H, Guan Z, Cuk K, Brenner H, Zhang Y. Circulating microRNA biomarkers for lung cancer detection in Western populations. Cancer Med. 2018;7:4849–4862. doi:10.1002/cam4.2018.7.issue-10
11. Han Y, Li H. miRNAs as biomarkers and for the early detection of non-small cell lung cancer (NSCLC). J Thorac Dis. 2018;10(5):3119–3131. doi:10.21037/jtd.2018.05.32
12. Chen W, Huang Y, Zhang S, et al. MicroRNA-212 suppresses nonsmall lung cancer invasion and migration by regulating ubiquitin-specific protease-9. J Cell Biochem. 2019;120(4):6482–6489.
13. Song Q, Ji Q, Xiao J, et al. miR-409 inhibits human non-small-cell lung cancer progression by directly targeting SPIN1. Mol Ther Nucleic Acids. 2018;13:154–163. doi:10.1016/j.omtn.2018.08.020
14. He X, Chen SY, Yang Z, et al. miR-4317 suppresses non-small cell lung cancer (NSCLC) by targeting fibroblast growth factor 9 (FGF9) and cyclin D2 (CCND2). J Exp Clinl Cancer Res. 2018;37(1):230. doi:10.1186/s13046-018-0882-4
15. Li S, Zeng X, Ma R, Wang L. MicroRNA-21 promotes the proliferation, migration and invasion of non-small cell lung cancer A549 cells by regulating autophagy activity via AMPK/ULK1 signaling pathway. Exp Ther Med. 2018;16(3):2038–2045. doi:10.3892/etm.2018.6370
16. Jin X, Yu Y, Zou Q, et al. MicroRNA-105 promotes epithelial-mesenchymal transition of nonsmall lung cancer cells through upregulating Mcl-1. J Cell Biochem. 2019;120(4):5880–5888.
17. Li Y, Cui X, Li Y, Zhang T, Li S. Upregulated expression of miR-421 is associated with poor prognosis in non-small-cell lung cancer. Cancer Manag Res. 2018;10:2627–2633. doi:10.2147/CMAR.S167432
18. Boeri M, Pastorino U, Sozzi G. Role of microRNAs in lung cancer: microRNA signatures in cancer prognosis. Cancer J. 2012;18(3):268–274. doi:10.1097/PPO.0b013e318258b743
19. Chen L, Cao Y, Rong D, Wang Y, Cao Y. MicroRNA-605 functions as a tumor suppressor by targeting INPP4B in melanoma. Oncol Rep. 2017;38(2):1276–1286. doi:10.3892/or.2017.5740
20. Zhou YJ, Yang HQ, Xia W, et al. Down-regulation of miR-605 promotes the proliferation and invasion of prostate cancer cells by up-regulating EN2. Life Sci. 2017;190:7–14. doi:10.1016/j.lfs.2017.09.028
21. Li J, Tian F, Li D, et al. MiR-605 represses PSMD10/Gankyrin and inhibits intrahepatic cholangiocarcinoma cell progression. FEBS Lett. 2014;588(18):3491–3500. doi:10.1016/j.febslet.2014.08.008
22. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262
23. Feng J, Zhang X, Zhu H, Wang X, Ni S, Huang J. High expression of FoxP1 is associated with improved survival in patients with non-small cell lung cancer. Am J Clin Pathol. 2012;138(2):230–235. doi:10.1309/AJCPDHQFNYJZ01YG
24. Song CJ, Chen H, Chen LZ, Ru GM, Guo JJ, Ding QN. The potential of microRNAs as human prostate cancer biomarkers: A meta-analysis of related studies. J Cell Biochem. 2018;119(3):2763–2786. doi:10.1002/jcb.26445
25. Hajizamani S, Shahjahani M, Shahrabi S, Saki N. MicroRNAs as prognostic biomarker and relapse indicator in leukemia. Clin Transl Oncol. 2017;19(8):951–960. doi:10.1007/s12094-017-1638-x
26. Lu J, Zhan Y, Feng J, Luo J, Fan S. MicroRNAs associated with therapy of non-small cell lung cancer. Int J Biol Sci. 2018;14(4):390–397. doi:10.7150/ijbs.22243
27. Uddin A, Chakraborty S. Role of miRNAs in lung cancer. J Cell Physiol. 2018. doi:10.1002/jcp.26607
28. Zhou Q, Huang SX, Zhang F, et al. MicroRNAs: A novel potential biomarker for diagnosis and therapy in patients with non-small cell lung cancer. Cell Prolif. 2017;50(6). doi:10.1111/cpr.12368
29. Shu W, Yang H, Zhang L, Lu MM, Morrisey EE. Characterization of a new subfamily of winged-helix/forkhead (Fox) genes that are expressed in the lung and act as transcriptional repressors. J Biol Chem. 2001;276(29):27488–27497. doi:10.1074/jbc.M100636200
30. Hu Z, Zhu L, Gao J, et al. Expression of FOXP1 in epithelial ovarian cancer (EOC) and its correlation with chemotherapy resistance and prognosis. Tumor Biol. 2015;36(9):7269–7275. doi:10.1007/s13277-015-3383-5
31. Xiao J, He B, Zou Y, et al. Prognostic value of decreased FOXP1 protein expression in various tumors: a systematic review and meta-analysis. Sci Rep. 2016;6:30437. doi:10.1038/srep30437
32. Mizunuma M, Yokoyama Y, Futagami M, Horie K, Watanabe J, Mizunuma H. FOXP1 forkhead transcription factor is associated with the pathogenesis of endometrial cancer. Heliyon. 2016;2(5):e00116. doi:10.1016/j.heliyon.2016.e00116
33. Choi EJ, Seo EJ, Kim DK, et al. FOXP1 functions as an oncogene in promoting cancer stem cell-like characteristics in ovarian cancer cells. Oncotarget. 2016;7(3):3506–3519. doi:10.18632/oncotarget.6510
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.