")
Back to Journals » OncoTargets and Therapy » Volume 12
microRNA-605 directly targets SOX9 to alleviate the aggressive phenotypes of glioblastoma multiforme cell lines by deactivating the PI3K/Akt pathway
Authors Jia J, Wang J, Yin M, Liu Y
Received 21 April 2019
Accepted for publication 17 June 2019
Published 8 July 2019 Volume 2019:12 Pages 5437—5448
DOI https://doi.org/10.2147/OTT.S213026
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr William C. Cho
This paper has been retracted.
Jianwu Jia, 1,* Jing Wang, 1,* Meifeng Yin, 1 Yongdong Liu 2
1Department of Neurosurgery, Yidu Central Hospital of Weifang, Weifang, Shandong 262500, People’s Republic of China; 2Department of Pediatrics, Weifang People’s Hospital, Weifang, Shandong 261041, People’s Republic of China
*These authors contributed equally to this work
Background: Aberrant microRNA (miRNA) expression has been widely reported to play a crucial role in the progression and development of glioblastoma (GBM). miR-605 has been identified as a tumor-suppressing miRNA in several types of human cancers. Nevertheless, the expression profile and detailed roles of miR-605 in GBM remain unclear and need to be further elucidated.
Materials and methods: RT-qPCR analysis was utilized for the determination of miR-605 expression in GBM tissues and cell lines. In addition, CCK-8 assay, transwell migration and invasion assays, as well as sub-cutaneous xenograft mouse models were utilized to evaluate the effects of miR-605 upregulation in GBM cells. Notably, the potential mechanisms underlying the activity of miR-605 in the malignant phenotypes of GBM were explored.
Results: We observed that expression of miR-605 was reduced in GBM tissues and cell lines. Decreased miR-605 expression exhibited significant correlation with KPS score. The overall survival rate in GBM patients with low miR-605 expression was lower than that of patients with high miR-605 expression. Increased miR-605 expression suppressed the proliferation, migration, and invasion of U251 and T98 cells. In addition, miR-605 upregulation impaired tumor growth in vivo. Furthermore, SRY-Box 9 (SOX9) was identified as a direct target gene of miR-605 in U251 and T98 cells. SOX9 expression was shown to exhibit an inverse correlation with miR-605 expression in GBM tissues. Moreover, silencing of SOX9 expression mimicked the tumor-suppressing roles of miR-605 in U251 and T98 cells, while SOX9 restoration rescued the suppressive effects of miR-605 overexpression in the same. Notably, miR-605 suppressed the PI3K/Akt pathway in GBM in vitro and in vivo.
Conclusion: These results demonstrated that miR-605 acts as a tumor suppressor in the development of GBM by directly targeting SOX9 and inhibiting the activation of the PI3K/Akt pathway, suggesting its potential role as a therapeutic target for GBM.
Keywords: glioblastoma multiforme, microRNA-605, malignant phenotypes, SRY-Box 9
Introduction
Glioma, a human malignant tumor that originates from neural cells, is the most prevalent and aggressive type of primary brain tumor in adults.1 Based on the pathological histology, the World Health Organization (WHO) classified all glioma into four tumor grades: two low-grade astrocytomas (WHO grade I-II), anaplastic astrocytomas (WHO grade III), and glioblastoma (GBM, WHO grade IV).2 In the past decade, there has been remarkable development in the comprehensive techniques used to treat patients with GBM, including surgical resection, immunotherapy, radiotherapy, and chemotherapy.3 Unfortunately, their therapeutic efficiencies are still poor, and their clinical prognosis is unsatisfactory with a 5-year survival rate of less than 5%.4 Dysregulation of gene expression and chromosomal anomalies have been demonstrated to be implicated in the occurrence and development of GBM.5–7 Nevertheless, the molecular mechanisms involved in crucial tumorigenic events remain elusive and need to be further investigated. It is, therefore, extremely urgent to explore the detailed molecular mechanisms associated with the multi-step process of GBM pathogenesis and identify effective therapeutic approaches for patients with this deadly disease.
microRNAs (miRNAs) are a series of endogenous and noncoding short RNAs about 18–23 nucleotides long.8 miRNAs act as regulatory molecules by directly binding to the target mRNAs in the 3′-untranslated regions (3′-UTRs), thereby resulting in either mRNA degradation or translational inhibition.9 So far, over 1,800 human miRNAs have been identified in the human genome, and these miRNAs play pivotal roles in the regulation of numerous biological processes, particularly human cancer.10 Aberrant expression of miRNAs has previously been reported in GBM.11–13 For instance, miR-129-3p,14 miR-135a,15 and miR-48516 are expressed at low levels in GBM, whereas miR-217,17 miR-595,18 and miR-128819 are highly expressed in GBM. miRNAs can play tumor-suppressive or oncogenic roles in the progression and development of GBM depending on the functional characterization of their target genes.20 Accordingly, further exploration of cancer-associated miRNAs in GBM may facilitate the identification of potential diagnostic indicators, prognostic factors, and therapeutic targets.
miR-605 has been identified as a tumor-suppressing miRNA in prostate cancer,21 melanoma,22 and intrahepatic cholangiocarcinoma.23 Nevertheless, the expression profile and detailed roles of miR-605 in GBM remain unclear and need to be further elucidated. Our results not only showed that miR-605 was downregulated in GBM but also suggested a crucial role for the aberrant expression of miR-605 in the development of malignant GBM, thus highlighting its potential as a therapeutic target in GBM patients.
Materials and methods
Patients and clinical tissues
Our study was carried out under the approval of the Ethics Committee of Yidu Central Hospital of Weifang, and was performed in accordance with the Declaration of Helsinki. Written informed consent was provided by all participators prior to surgical excision. In total, 44 pairs of GBM tissues and adjacent non-tumorous tissues were collected from patients who were diagnosed with GBM and underwent surgical resection at Yidu Central Hospital of Weifang. Patients were excluded from this research if they were treated with immunotherapy, radiotherapy, or chemotherapy before their enrollment in this study. After surgical resection, all tissue specimens were immediately snap-frozen in liquid nitrogen and stored at −80 °C for RNA or protein isolation.
Cell lines
A total of four GBM cell lines, including U138, U251, T98, and LN229, were obtained from the Shanghai Institute of Biochemistry and Cell Biology (Shanghai, China), and cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific), penicillin (100 U/mL), and streptomycin (100 mg/mL; Gibco; Thermo Fisher Scientific, Inc.). Normal human astrocytes (NHAs), bought from ScienCell Research Laboratories (Carlsbad, CA, USA), were maintained in astrocyte medium (ScienCell Research Laboratories) supplemented with 10% FBS. All cells were grown at 37 °C in a humidified chamber containing 5% CO2.
Oligonucleotides, plasmid, and cell transfection
To restore miR-605 expression, agomir-605 and its negative control, agomir-NC, was obtained from GenePharma (Shanghai, China). For the knockdown assay, small interfering (si) RNAs targeting the expression of SOX9 (si-SOX9) and its scramble control, siRNA (si-ctrl), were chemically synthesized by Ribobio (Guangzhou, China). The enforced expression plasmid (pcDNA3.1) for SOX9 (pcDNA3.1-SOX9; pc-SOX9) and empty plasmid, pcDNA3.1, were generated by the Chinese Academy of Sciences (Changchun, China). The transient transfection for oligonucleotides, siRNA, or plasmid was performed using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.) in accordance with the manufacturer’s protocols. The Cell Counting Kit-8 (CCK-8) assay, transwell migration and invasion assays, and sub-cutaneous xenograft mouse model were carried out at 24, 48 and 24 h after transfection, respectively. Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) was carried out to measure gene expression at 48 h post-transfection, while Western blotting was conducted in transfected cells after 72 h transfection.
RNA preparation and RT-qPCR
The isolation of total miRNA from tissue samples or cells was performed using mirVana™ miRNA Isolation kit (Ambion, Austin, TX, USA). Total miRNA was reverse transcribed into complementary DNA (cDNA) using the miScript Reverse Transcription kit (Qiagen GmbH, Hilden, Germany), and the cDNA was then used for quantitative PCR (qPCR) using the miScript SYBR Green PCR kit (Qiagen GmbH).
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) was used to extract total RNA from tissue samples or cells. To quantify SOX9 mRNA expression, the Prime-Script RT reagent Kit and the SYBR Premix Ex Taq (both from Takara Biotechnology Co., Ltd, Dalian, China) were employed to conduct reverse transcription and qPCR, respectively. U6 small nuclear RNA (snRNA) served as an internal control for miR-605 expression, while GAPDH was used as the internal reference for the expression level of SOX9 mRNA. Relative gene expression was calculated by 2−∆∆Cq method. The primer sequences were designed as follows: miR-605, 5′-CGCGGATCCAGAGTGCTGAGATTGCAG-3′ (forward) and 5′- CCGGAATTCCAGTTACGCCACATGAGG-3′ (reverse); U6, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ (forward) and 5′-CGCTTCACGAATTTGCGTGTCAT-3′ (reverse); SOX9, 5′-CTGGGAACAACCCGTCTA-3′ (forward) and 5′- GGGTAATGCGCTTGGATA-3′ (reverse); and GAPDH, 5′-CGGAGTCAACGGATTTGGTCGTAT-3′ (forward) and 5′-AGCCTTCTCCATGGTGGTGAAGAC-3′ (reverse).
CCK-8 assay
The CCK-8 reagent contains WST-8: chemical name: 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfonate Acid benzene)-2H-tetrazole monosodium salt. It is reduced by the dehydrogenase in the mitochondria of the cell to the highly water-soluble yellow formazan product Formazan under the action of the electron carrier 1-methoxy-5-methylphenazine dimethyl sulfate (1-Methoxy PMS). The amount of formazan produced is proportional to the number of living cells.
Cells were harvested one day after transfection and seeded with a density of 2 × 103 cells/well in 96-well plates. Cellular proliferation was measured using a WST-8 CCK-8 (CCK-8; Dojindo, Tokyo, Japan) at four time points: 0, 1, 2, and 3 days after inoculation. Ten microliters of CCK-8 solution were added into each well and the plates were incubated at 37 °C under 5% CO2 for another 2 h. The absorbance at 450 nm (A450) was read using a microplate reader (Bio-Tek Instruments, Winooski, VT, USA).
Transwell migration and invasion assays
For transwell invasion assay, transfected cells were suspended in FBS-free DMEM medium. In total, 200 µL of cell suspension containing 5×104 cells was placed into the upper compartments of the Matrigel pre-coated transwell chambers (BD Biosciences, Franklin Lakes, NJ, USA), followed by the addition of 600 μL DMEM supplemented with 20% FBS in the lower compartments. After being cultured for 24 h, the non-invasive cells remaining on the upper side of the Transwell filter were gently wiped away with cotton wool. The invasive cells were fixed with 100% methanol, stained with 0.5% crystal violet, and then, washed with phosphate buffer solution. The invasive ability was determined by counting the number of invasive cells in five randomly selected microscopic fields using an inverted light microscope (IX71; Olympus Corporation, Tokyo, Japan). In the transwell migration assay, the Matrigel was not used to precoat the transwell chambers. The remaining experimental procedures were similar to those used for the transwell invasion assay.
Sub-cutaneous xenograft mouse model
Animal experiments were approved by the Animal Ethics Committee of Yidu Central Hospital of Weifang, and performed in accordance with the protocols of Animal Ethics Committee of Yidu Central Hospital of Weifang and conducted in accordance with the guidance of Animal Protection Law of the People’s Republic of China-2009 for experimental animals. Female BALB/c nude mice (3–4 weeks of age) were bought from the Animal Center of Southern Medical University (Guangzhou, China) and maintained under specific pathogen-free conditions. All nude mice were divided into two groups: one was injected with agomir-605-transfected cells and the other with agomir-NC-transfected cells. The width and length of the tumor xenograft was recorded with Vernier calipers, and their volume was analyzed using the formula: 1/2× tumor length × tumor width2. After 4 weeks, all nude mice were sacrificed, and the tumor xenografts were obtained and used for further analysis.
Bioinformatics prediction
The potential targets of miR-605 were predicted using the software TargetScan (http://www.targetscan.org/vert_71/), miRDB (http://mirdb.org/), and microRNA (http://www.microrna.org/microrna/home.do).
Luciferase reporter assay
The 3′-UTR fragments of SOX9 containing the wild-type (wt) and mutant (mut) miR-605 binding site were amplified by GenePharma, and then, inserted in the pMIR-REPORT vector (Promega, Madison, WI, USA) to generate the pMIR-SOX9-3′-UTR wt and pMIR-SOX9-3′-UTR mut vectors, respectively. Cells were seeded into 24-well plates and incubated at 37 °C for one night. The synthetic luciferase reporter plasmids were co-transfected with either agomir-605 or agomir-NC into cells using Lipofectamine 2000, according to the manufacturer’s instructions. Forty-eight hours later, firefly luciferase activity was determined using the Dual-Luciferase Reporter Assay System (Promega) and normalized to the activity of Renilla luciferase.
Western blot analysis
Proteins were extracted from the tissue samples or cells by radioimmunoprecipitation assay lysis buffer (Beyotime Institute of Biotechnology Co., Ltd., Shanghai, China). Total protein was quantified using a Bradford protein assay kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Equivalent amounts of protein were separated using 10% SDS-PAGE, and then, electrophoretically transferred onto PVDF membranes. After blocking with 5% evaporated skimmed milk overnight at 4 °C, the primary antibodies were added, and the membranes were incubated at 4 °C overnight. Thereafter, the membranes were washed thrice with Tris-buffered saline containing 0.1% Tween-20 (TBST) followed by incubation with goat anti-rabbit (cat no. ab6721; Abcam, Cambridge, UK) or goat anti-mouse (cat no. ab6789; Abcam) horseradish peroxidase (HRP)-conjugated secondary antibody at room temperature for 2 h. Finally, the membranes were washed with TBST three times and protein signals were visualized using an enhanced chemiluminescence reagent (ECL; Pierce; Thermo Fisher Scientific, Inc.). The primary antibodies used were as follows: rabbit anti-human SOX9 antibody (cat. no. ab185966; Abcam), rabbit anti-human p-PI3K (cat. no. ab182651; Abcam,), rabbit anti-human PI3K (cat. no. ab191606; Abcam), mouse anti-human p-Akt (cat. no. sc-514032; Santa Cruz Biotechnology, CA, USA), mouse anti-human Akt (cat. no.sc-81434; Santa Cruz Biotechnology), and rabbit anti-human GAPDH antibody (cat. no. ab128915; 1:5,000 dilution; Abcam).
Statistical analysis
All experiments were conducted at least three times independently, and the obtained results were presented as the mean ± standard deviation. Student’s t-test was utilized for comparison of the differences between two groups, while the differences among multiple groups were investigated using one-way analysis of variance followed by a Tukey’s post-hoc test. A χ2 test was performed to examine the correlation between miR-605 expression and the clinicopathological characteristics of patients with GBM. A Spearman’s correlation analysis was applied to explore the association between miR-605 and SOX9 expressions in GBM tissues. The association between miR-605 expression and overall survival rate of GBM patients was analyzed using a log-rank test. A value of P < 0.05 was considered to indicate statistically significant differences.
Results
miR-605 is downregulated in GBM and is associated with poor prognosis in GBM patients
We first analyzed the expression profile of miR-605 in GBM tissues. RT-qPCR was performed to measure miR-605 expression in 44 pairs of GBM tissues and adjacent non-tumorous tissues. The expression level of miR-605 in GBM tissues was significantly lower than that in non-tumorous tissues (Figure 1A, P<0.05). In addition, we detected the expression level of miR-605 in four GBM cell lines (U138, U251, T98, and LN229) and normal human astrocytes (NHAs). Compared with NHAs, miR-605 expression was decreased in all four GBM cell lines (Figure 1B, P<0.05).
 |
Figure 1 miR-605 is downregulated in GBM and indicates poor prognosis. (A) miR-605 expression in 44 pairs of GBM tissues and adjacent non-tumorous tissues was measured by RT-qPCR analysis. RT-qPCR was performed in triplicate and repeated three times. *P<0.05 vs non-tumorous tissues. (B) RT-qPCR was utilized for the determination of miR-605 expression in four GBM cell lines (U138, U251, T98, and LN229) and NHAs. RT-qPCR was performed in triplicate and repeated three times. *P<0.05 vs NHAs. (C) The Kaplan-Meier method and log-rank test was used to analyze the overall survival of GBM patients with high/low miR-605 expression. P=0.0134.Abbreviations: GBM, glioblastoma; NHAs, normal human astrocytes. |
All GBM patients were divided into two groups according to the median value of miR-605 expression level in GBM tissues, including a miR-605-low expression group and a miR-605-high expression group. Subsequently, we explored the associations between miR-605 expression level and the clinicopathological parameters of GBM patients. Decreased miR-605 expression was notably correlated with KPS score (P=0.033; Table 1). Furthermore, GBM patients having low miR-605 expression exhibited shorter overall survival rates compared with those with high miR-605 expression level (Figure 1C, P=0.0047). These results indicated that miR-605 expression was decreased in GBM tissue, as compared to normal brain tissue, and was associated with a poor prognosis.
 |
Table 1 The association between of miR-605 and clinicopathological characteristics in patients with GBM |
miR-605 inhibits cell proliferation, migration, and invasion of GBM cells
To explore the detailed roles of miR-605 in GBM cells, the U251 and T98 cell lines,which exhibited the lowest expression level of miR-605 among the four GBM cell lines, were selected for further experiments. To increase endogenous miR-605 expression, the U251 and T98 cell lines were transfected with agomir-605, whilethe control cells were transfected with agomir-NC. After transfection, miR-605 was markedly upregulated in agomir-605-transfected U251 and T98 cells compared to the cells transfected with agomir-NC (Figure 2A, P<0.05). The CCK-8 assay was performed to determine cellular proliferation, and it revealed that miR-605 upregulation significantly suppressed the proliferation of U251 and T98 cells (Figure 2B, P<0.05). Transwell migration and invasion assays were employed to assess the migratory and invasive abilities of U251 and T98 cells after miR-605 overexpression. The migration (Figure 2C, P<0.05) and invasion (Figure 2D, P<0.05) of U251 and T98 cells was significantly decreased upon miR-605 overexpression. Taken together, these findings suggested that miR-605 may play a tumor suppressive role in GBM progression by inhibiting cell growth and invasiveness in vitro.
 |
Figure 2 miR-605 suppresses the proliferation, migration, and invasion of U251 and T98 cells. (A) The expression level of miR-605 in U251 and T98 cells was determined by RT-qPCR after agomir-605 or agomir-NC transfection. RT-qPCR was performed in triplicate and repeated three times. *P<0.05 vs agomir-NC. (B) The proliferation of miR-605 overexpressing-U251 and T98 cells was detected using the CCK-8 assay, which was repeated three times.*P<0.05 vs agomir-NC. (C, D) Transwell migration and invasion assays were used to explore whether miR-605 is involved in the regulation of U251 and T98 cell migration and invasion. Representative images were shown at 200× magnification. Transwell migration and invasion assays were repeated three times.*P<0.05 vs agomir-NC. |
SOX9 is a direct target gene of miR-605 in GBM cells
Having demonstrated that miR-605 expression was decreased in GBM tissue compared to normal neural tissues, and that it inhibited the tumor cell proliferation, migration and invasion, we then explored the molecular mechanisms underlying the tumor-suppressive activity of miR-605 in GBM cells. Firstly, bioinformatics tools were used to search for potential targets of miR-605. A binding site for miR-605 was observed in the 3′-UTR of SOX9 (Figure 3A). Among these candidates, SOX9, frequently reported to be implicated in the formation and progression of GBM,24–30 was chosen for further verification. A luciferase reporter assay was performed to investigate whether miR-605 could directly interact with the 3′-UTR of SOX9. As indicated in Figure 3B, the luciferase reporter plasmid carrying the wt miR-605 binding site led to a significant decrease in the luciferase activity after transfection with agomir-605 (P<0.05), whereas the plasmid harboring the mut 3ʹ-UTR had no effect on luciferase activity.
 |
Figure 3 SOX9 is the direct target gene of miR-605 in GBM cells. (A) The sequences of miR-605 and predicted wild-type binding site of miR-605 in the 3ʹ-UTR of SOX9. The mutant binding site is also shown. (B) agomir-605 or agomir-NC in combination with the vector pMIR-SOX9-3′-UTR wt or pMIR-SOX9-3′-UTR mut was co-transfected into U251 and T98 cells. Luciferase activity was examined at 48 h post-transfection. The lucifease reporter assay was repeated three times *P<0.05 vs agomir-NC. (C) The expression of SOX9 mRNA in 44 pairs of GBM tissues and adjacent non-tumorous tissues was quantified using RT-qPCR. RT-qPCR was performed in triplicate and repeated three times.*P<0.05 vs non-tumorous tissues. (D) The correlation between miR-605 and SOX9 mRNA expression in the same GBM tissues (n=44) was explored using Spearman’s correlation analysis. R2=0.3616, P<0.0001. (E, F) RT-qPCR and Western blot analysis was employed to measure the SOX9 mRNA and protein levels in miR-605-overexpressing U251 and T98 cells. Both assays were repeated three times. *P<0.05 vs agomir-NC.Abbreviation: GBM, glioblastoma. |
We further detected SOX9 mRNA expression in 44 pairs of GBM tissues and adjacent non-tumorous tissues using RT-qPCR. The results revealed that the mRNA level of SOX9 in GBM tissues was significantly higher than that in adjacent non-tumorous tissues (Figure 3C, P<0.05). In addition, an inverse correlation was observed between the expression levels of miR-605 and SOX9 mRNA in the same GBM tissues (Figure 3D; n = 44, R2 = 0.3616, P < 0.001). Furthermore, the expression levels of SOX9 mRNA (Figure 3E; P<0.05) and protein (Figure 3F; P<0.05) were both reduced after agomir-605 transfection in U251 and T98 cells. These findings suggested that SOX9 is a direct target of miR-605 in GBM cells.
Downregulation of SOX9 inhibits GBM cell proliferation, migration, and invasion
To explore whether the specific roles of SOX9 silencing and miR-605 upregulation were similar, a siRNA against SOX9 (si-SOX9) was used to knockdown endogenous SOX9 expression in U251 and T98 cells. The SOX9 protein level was efficiently silenced in U251 and T98 cells that were transected with si-SOX9 (Figure 4A, P<0.05). The CCK-8 assay revealed that the proliferation of U251 and T98 cells was significantly decreased after SOX9 downregulation (Figure 4B, P<0.05). In addition, the transwell migration and invasion assays revealed that silenced SOX9 expression significantly suppressed the migratory (Figure 4C, P<0.05) and invasive (Figure 4D, P<0.05) abilities of U251 and T98 cells compared with that of cells transfected with si-ctrl. In summary, the functional roles of SOX9 silencing in GBM cells were similar to those induced by miR-605 upregulation, indicating SOX9 as downstream target of miR-605 in GBM cells.
 |
Figure 4 SOX9 knockdown inhibits U251 and T98 cell proliferation, migration, and invasion in vitro. (A) U251 and T98 cells were transfected with si-SOX9 or si-ctrl. After culture for 72 h, the transfection efficiency was evaluated using Western blot analysis. Western blotting was repeated three times. *P<0.05 vs si-ctrl. (B) Effect of silencing SOX9 expression on U251 and T98 cell proliferation was determined by the CCK-8 assay. The CCK-8 assay was repeated three times.*P<0.05 vs si-ctrl. (C, D) Transwell migration and invasion assays were performed for the determination of migratory and invasive abilities of U251 and T98 cells after transfection with si-SOX9 or si-ctrl. Transwell migration and invasion assays were repeated three times.*P<0.05 vs si-ctrl. |
SOX9 expression restoration eliminates the effects of miR-605 overexpression on the malignant phenotypes of GBM cells
A series of rescue experiments were carried out to validate whether miR-605 acted by decreasing SOX9 expression in GBM cells. Since SOX9 expression decreased in agomir-605-transfected U251 and T98 cells, we rescued the protein expression of SOX9 in miR-605 expressing cells by co-transfecting with the SOX9 overexpression plasmid pc-SOX9. Western blot analysis revealed that co-transfection with pc-SOX9 successfully recovered the suppressive effect of miR-605 overexpression on the SOX9 protein level (Figure 5A, P<0.05). Further experiments showed that restoration of SOX9 expression reversed the reduction in proliferation (Figure 5B, P<0.05), migration (Figure 5C, P<0.05), and invasion (Figure 5D, P<0.05) of U251 and T98 cells that was caused by miR-605 upregulation. These results indicated that miR-605 exerts its anticancer activity in the malignant phenotypes of GBM, at least partially, by decreasing SOX9 expression.
 |
Figure 5 Restoring SOX9 expression abolishes the miR-605-induced effects on the malignant phenotypes of U251 and T98 cells. (A) Western blot analysis was performed to detect the protein level of SOX9 in U251 and T98 cells transfected with pc-SOX9 or pcDNA3.1 in the presence of agomir-605. Western blotting was repeated three times. *P<0.05 vs agomir-NC. #P<0.05 vs agomir-605+ pcDNA3.1. (B-D) The proliferation, migration, and invasion of the aforementioned cells was investigated using the CCK-8, transwell migration, and invasion assays, respectively. Assays were repeated three times. *P<0.05 vs agomir-NC. #P<0.05 vs agomir-605+ pcDNA3.1. |
miR-605 inhibits the PI3K/Akt pathway in GBM by downregulating SOX9 expression
Previous studies have reported that SOX9 is implicated in the regulation of the PI3K/Akt pathway.31,32 Hence, we next attempted to determine whether miR-605 inhibited the activation of the PI3K/Akt pathway in GBM cells by decreasing SOX9 expression. Western blotting indicated that the protein levels of p-PI3K and p-Akt in U251 and T98 cells were downregulated by miR-605 overexpression, which were reversed by co-transfection with pc-SOX9 (Figure 6). Thus, miR-605 suppressed the PI3K/Akt pathway in GBM cells by decreasing SOX9 expression.
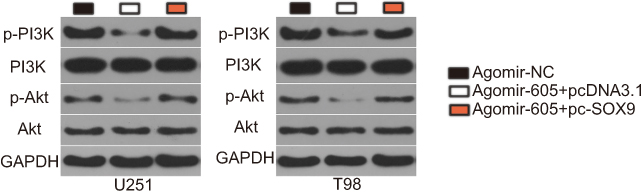 |
Figure 6 miR-605 suppresses the activation of the PI3K/Akt pathway in GBM cells. agomir-605 was co-transfected with pc-SOX9 or pcDNA3.1 into U251 and T98 cells. Seventy two hours after transfection, Western blot analysis was conducted to measure p-PI3K, PI3K, p-Akt, and Akt protein levels. Western blotting was repeated three times.Abbreviation: GBM, glioblastoma. |
miR-605 suppresses the tumor growth of GBM cells in vivo
Finally, the influence of miR-605 on GBM tumorigenicity in vivo was explored using a xenograft mouse model. Tumor xenografts from the agomir-605-treated group showed a significantly decreased volume (Figure 7A and B, P<0.05) and weight (Figure 7C, P<0.05) compared with those in the agomir-NC-treated group. We then determined the expression level of miR-605 in the tumor xenografts and investigated whether the suppression of GBM tumor growth in vivo was caused by miR-605 upregulation. The RT-qPCR data confirmed a significant miR-605 overexpression in the tumor xenografts derived from the agomir-605-treated group (Figure 7D, P<0.05). In addition, Western blot analysis revealed that SOX9, p-PI3K, and p-Akt expression were significantly downregulated in the miR-605 overexpressing group compared with the agomir-NC group (Figure 7E). These results suggested that miR-605 impaired the GBM growth in vivo by downregulating SOX9 expression and inhibiting the PI3K/Akt pathway.
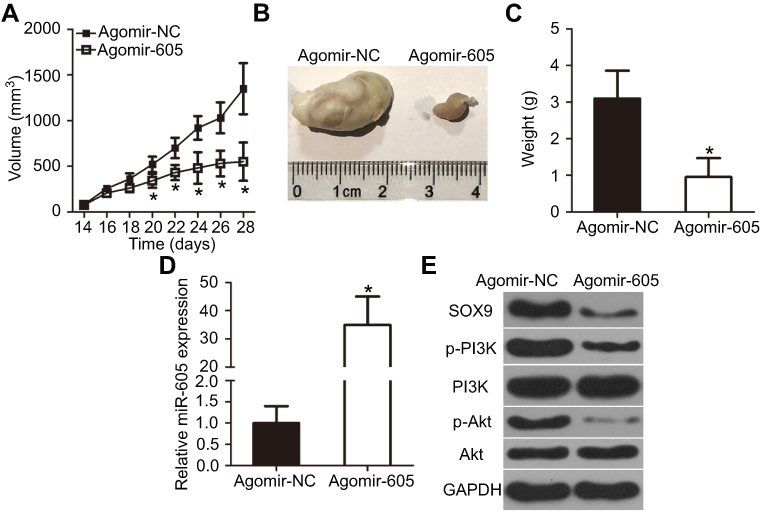 |
Figure 7 miR-605 upregulation impairs GBM growth in vivo. (A) U251 cells transfected with agomir-605 or agomir-NC. The transfected cells were injected subcutaneously into nude mice. Tumor growth curves after injection of nude mice with agomir-605 or agomir-NC-transfected U251 cells. *P<0.05 vs agomir-NC. (B) Representative images of tumor xenografts derived from nude mice implanted with agomir-NC- or agomir-605-transfected U251 cells. (C) Tumor xenografts were isolated and weighed. *P<0.05 vs agomir-NC. (D) Quantification of miR-605 expression in the tumor xenografts derived from agomir-605 or agomir-NC groups. RT-qPCR was performed in triplicate and repeated three times.*P<0.05 vs agomir-NC. (E) Total protein was extracted from tumor xenografts, and subjected to Western blot analysis for the measurement of SOX9, p-PI3K, PI3K, p-Akt, and Akt protein expression levels. Western blotting was repeated three times.Abbreviation: GBM, glioblastoma. |
Discussion
Aberrant miRNA expression has been widely reported to play crucial roles in the progression and development of GBM.33–35 Therefore, determining the specific roles of miRNAs in GBM may facilitate the identification of therapeutic targets and prognostic biomarkers for GBM patients. In this study, for the first time, we evaluated the expression profile of miR-605 in GBM patients and examined its clinical significance. Notably, the roles and potential underlying mechanisms of miR-605 in the malignant phenotypes of GBM in vitro and in vivo were explored in detail. These findings provided novel evidence of the tumor-suppressing action of miR-605 in GBM cell lines by directly targeting SOX9 and inhibiting the activation of the PI3K/Akt pathway, suggesting that this miRNA may be utilized as a promising therapeutic target for GBM patients.
miR-605 is expressed at low levels in prostate cancer tissues and cell lines.21 Downregulation of miR-605 is also observed in melanoma22 and intrahepatic cholangiocarcinoma.23 However, the expression profile of miR-605 in GBM remains to be elucidated. In this study, RT-qPCR was used to determine miR-605 expression in GBM tissues and cell lines. The results showed that expression of miR-605 was decreased in both GBM tissues and cell lines. The decrease in miR-605 expression was significantly associated with KPS score. GBM patients harboring low miR-605 expression exhibited shorter overall survival rates compared with those with high miR-605 expression. These results suggested that miR-605 may be a predictor for the therapeutic outcomes of GBM patients.
miR-605 is identified as a tumor suppressor in several types of human cancer. For instance, resumption of miR-605 expression suppresses the proliferation and invasion of prostate cancer cells in vitro.21 Ectopic miR-605 expression inhibited the growth of melanoma cells in vitro and in vivo.22 Restoring miR-605 expression inhibited intrahepatic cholangiocarcinoma cell proliferation, induced cell apoptosis in vitro, and reduced tumor growth in vivo.23 In our current study, we explored the detailed roles of miR-605 in GBM progression; it was revealed that miR-605 exerted an inhibitory role in GBM cell proliferation, migration, invasion in vitro, and tumor growth in vivo. These findings suggested that miR-605 may be a promising therapeutic target for GBM patients.However, a limitation of our study is that the characteristic infiltrative nature of GBM is lost in sub-cutaneous xenografts. This loss is well illustrated in Figure 7B showing a sub-cutaneous tumor totally encapsulated. Thus, we will use a xenograft mouse model of orthotopic GBM tumors to further confirm our findings in the future.
Multiple genes, such as engrailed-2 in prostate cancer;21 inositol polyphosphate-4-phosphatase, type II in melanoma;22 and proteasome 26S subunit, non-ATPase 10 in intrahepatic cholangiocarcinoma,23 have been reported as direct targets of miR-605. SOX9, a member of the sex-determining region Y box family, was validated to be a direct target of miR-605 in GBM. SOX9 is overexpressed in various types of human malignant tumors, including hepatocellular carcinoma,36 thyroid cancer,37 gastric cancer,38 esophageal squamous cell carcinoma,39 and non-small cell lung cancer.40 Expression of SOX9 is also increased in GBM cells, and the increase of SOX9 expression is significantly associated with WHO grade and KPS.24 GBM patients with high SOX9 expression exhibit shorter overall survival and disease-free survival rates than those with low SOX9 expression.24,29 In addition, a high SOX9 expression was identified as an independent prognostic indicator for GBM patients.24 The dysregulation of SOX9 is implicated in the genesis and development of GBM, and regulates several aggressive processes, including cell proliferation, cell cycle, colony formation, motility, metastasis, epithelial-mesenchymal transition, and chemosensitivity.25–30 Previous studies reported that SOX9 could be directly targeted by multiple miRNAs. For instance, miR-14541 and miR-30c42 directly interacted with SOX9 and decreased its expression in GBM cells, thus resulting in the suppression of GBM progression. Accordingly, SOX9 knockdown using miRNAs-based gene therapy might represent a potential therapeutic approach for the management of GBM patients.
Conclusion
Taken together, our results revealed that miR-605 decreased the malignancy of GBM in vitro and in vivo by directly targeting SOX9 and inhibiting the PI3K/Akt pathway. Our findings highlighted the importance of the miR-605/SOX9/PI3K/Akt axis in GBM progression and provided promising biomarkers and targets for the diagnosis, prognosis, and treatment of GBM patients.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Seliger C, Hau P. Drug repurposing of metabolic agents in malignant glioma. Int J Mol Sci. 2018;19(9):2768. doi:10.3390/ijms19092768
2. Louis DN, Perry A, Reifenberger G, et al. The 2016 World Health Organization classification of tumors of the central nervous system: a summary. Acta Neuropathol. 2016;131(6):803–820. doi:10.1007/s00401-016-1545-1
3. Li Z, Meng X, Jin L. Icaritin induces apoptotic and autophagic cell death in human glioblastoma cells. Am J Transl Res. 2016;8(11):4628–4643.
4. Hwang JH, Smith CA, Salhia B, Rutka JT. The role of fascin in the migration and invasiveness of malignant glioma cells. Neoplasia. 2008;10(2):149–159. doi:10.1593/neo.07909
5. Geraldo LHM, Garcia C, Da Fonseca ACC, et al. Glioblastoma therapy in the age of molecular medicine. Trends Cancer. 2019;5(1):46–65. doi:10.1016/j.trecan.2018.11.002
6. Chin C, Lunking ES, de la Fuente M, Ayad NG. Immunotherapy and epigenetic pathway modulation in glioblastoma multiforme. Front Oncol. 2018;8:521. doi:10.3389/fonc.2018.00521
7. Romani M, Pistillo MP, Banelli B. Epigenetic targeting of glioblastoma. Front Oncol. 2018;8:448. doi:10.3389/fonc.2018.00448
8. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. doi:10.1016/j.cell.2009.01.002
9. Sun K, Lai EC. Adult-specific functions of animal microRNAs. Nat Rev Genet. 2013;14(8):535–548. doi:10.1038/nrg3471
10. Banelli B, Forlani A, Allemanni G, Morabito A, Pistillo MP, Romani M. MicroRNA in glioblastoma: an overview. Int J Genomics. 2017;2017:7639084. doi:10.1155/2017/7639084
11. Zhu XY, Li GX, Liu ZL. MiR-599 as a potential biomarker for prognosis of glioma. Eur Rev Med Pharmacol Sci. 2018;22(2):294–298. doi:10.26355/eurrev_201801_14171
12. Wang N, Zhang Y, Liang H. MicroRNA-598 inhibits cell proliferation and invasion of glioblastoma by directly targeting metastasis associated in colon cancer-1 (MACC1). Oncol Res. 2018;26(8):1275–1283. doi:10.3727/096504018X15185735627746
13. Tan Z, Zhao J, Jiang Y. MiR-634 sensitizes glioma cells to temozolomide by targeting CYR61 through Raf-ERK signaling pathway. Cancer Med. 2018;7(3):913–921. doi:10.1002/cam4.1351
14. Fang DZ, Wang YP, Liu J, et al. MicroRNA-129-3p suppresses tumor growth by targeting E2F5 in glioblastoma. Eur Rev Med Pharmacol Sci. 2018;22(4):1044–1050. doi:10.26355/eurrev_201802_14387
15. Gomez Zubieta DM, Hamood MA, Beydoun R, Pall AE, Kondapalli KC. MicroRNA-135a regulates NHE9 to inhibit proliferation and migration of glioblastoma cells. Cell Commun Signal. 2017;15(1):55. doi:10.1186/s12964-017-0209-7
16. Mao K, Lei D, Zhang H, You C. MicroRNA-485 inhibits malignant biological behaviour of glioblastoma cells by directly targeting PAK4. Int J Oncol. 2017;51(5):1521–1532. doi:10.3892/ijo.2017.4122
17. Wang H, Zhi H, Ma D, Li T. MiR-217 promoted the proliferation and invasion of glioblastoma by repressing YWHAG. Cytokine. 2017;92:93–102. doi:10.1016/j.cyto.2016.12.013
18. Hao Y, Zhang S, Sun S, Zhu J, Xiao Y. MiR-595 targeting regulation of SOX7 expression promoted cell proliferation of human glioblastoma. Biomed Pharmacother. 2016;80:121–126. doi:10.1016/j.biopha.2016.03.008
19. Yin J, Weng C, Ma J, Chen F, Huang Y, Feng M. MicroRNA1288 promotes cell proliferation of human glioblastoma cells by repressing ubiquitin carboxylterminal hydrolase CYLD expression. Mol Med Rep. 2017;16(5):6764–6770. doi:10.3892/mmr.2017.7481
20. Hede K. Studies define role of microRNA in cancer. J Natl Cancer Inst. 2005;97(15):1114–1115. doi:10.1093/jnci/dji260
21. Zhou YJ, Yang HQ, Xia W, et al. Down-regulation of miR-605 promotes the proliferation and invasion of prostate cancer cells by up-regulating EN2. Life Sci. 2017;190:7–14. doi:10.1016/j.lfs.2017.09.028
22. Chen L, Cao Y, Rong D, Wang Y, Cao Y. MicroRNA-605 functions as a tumor suppressor by targeting INPP4B in melanoma. Oncol Rep. 2017;38(2):1276–1286. doi:10.3892/or.2017.5740
23. Li J, Tian F, Li D, et al. MiR-605 represses PSMD10/Gankyrin and inhibits intrahepatic cholangiocarcinoma cell progression. FEBS Lett. 2014;588(18):3491–3500. doi:10.1016/j.febslet.2014.08.008
24. Wang L, He S, Yuan J, et al. Oncogenic role of SOX9 expression in human malignant glioma. Med Oncol. 2012;29(5):3484–3490. doi:10.1007/s12032-012-0267-z
25. Sang Q, Liu X, Sun D. Role of miR-613 as a tumor suppressor in glioma cells by targeting SOX9. Onco Targets Ther. 2018;11:2429–2438. doi:10.2147/OTT.S156608
26. Xu X, Wang Z, Liu N, et al. Association between SOX9 and CA9 in glioma, and its effects on chemosensitivity to TMZ. Int J Oncol. 2018;53(1):189–202. doi:10.3892/ijo.2018.4382
27. Liu N, Zhang L, Wang Z, et al. MicroRNA-101 inhibits proliferation, migration and invasion of human glioblastoma by targeting SOX9. Oncotarget. 2017;8(12):19244–19254. doi:10.18632/oncotarget.13706
28. Liu X, Wang H, Zhu Z, Ye Y, Mao H, Zhang S. MicroRNA-105 targets SOX9 and inhibits human glioma cell progression. FEBS Lett. 2016;590(23):4329–4342. doi:10.1002/1873-3468.12458
29. Gao J, Zhang JY, Li YH, Ren F. Decreased expression of SOX9 indicates a better prognosis and inhibits the growth of glioma cells by inducing cell cycle arrest. Int J Clin Exp Pathol. 2015;8(9):10130–10138.
30. Liu H, Liu Z, Jiang B, Peng R, Ma Z, Lu J. SOX9 overexpression promotes glioma metastasis via Wnt/beta-catenin signaling. Cell Biochem Biophys. 2015;73(1):205–212. doi:10.1007/s12013-015-0647-z
31. Zhao Y, Pang W, Yang N, Hao L, Wang L. MicroRNA-511 inhibits malignant behaviors of breast cancer by directly targeting SOX9 and regulating the PI3K/Akt pathway. Int J Oncol. 2018;53(6):2715–2726. doi:10.3892/ijo.2018.4576
32. Xia S, Feng Z, Qi X, et al. Clinical implication of Sox9 and activated Akt expression in pancreatic ductal adenocarcinoma. Med Oncol. 2015;32(1):358. doi:10.1007/s12032-014-0358-0
33. Luo JW, Wang X, Yang Y, Mao Q. Role of micro-RNA (miRNA) in pathogenesis of glioblastoma. Eur Rev Med Pharmacol Sci. 2015;19(9):1630–1639.
34. Costa PM, Cardoso AL, Mano M, de Lima MC. MicroRNAs in glioblastoma: role in pathogenesis and opportunities for targeted therapies. CNS Neurol Disord Drug Targets. 2015;14(2):222–238. doi:10.2174/1871527314666150116123610
35. Wang H, Xu T, Jiang Y, Yan Y, Qin R, Chen J. MicroRNAs in human glioblastoma: from bench to beside. Front Biosci. 2015;20:105–118. doi:10.2741/4300
36. Guo X, Xiong L, Sun T, et al. Expression features of SOX9 associate with tumor progression and poor prognosis of hepatocellular carcinoma. Diagn Pathol. 2012;7:44. doi:10.1186/1746-1596-7-44
37. Huang J, Guo L. Knockdown of SOX9 inhibits the proliferation, invasion, and EMT in thyroid cancer cells. Oncol Res. 2017;25(2):167–176. doi:10.3727/096504016X14732772150307
38. Sun M, Uozaki H, Hino R, et al. SOX9 expression and its methylation status in gastric cancer. Virchows Archiv. 2012;460(3):271–279. doi:10.1007/s00428-012-1201-7
39. Higo N, Okumura H, Uchikado Y, et al. Expression of SOX9 is related to prognosis in patients with oesophageal squamous cell carcinoma. In Vivo. 2018;32(4):835–838. doi:10.21873/invivo.11316
40. Chen W, Zhao W, Chen S, et al. Expression and correlation of MALAT1 and SOX9 in non-small cell lung cancer. Clin Respir J. 2018;12(7):2284–2291. doi:10.1111/crj.12906
41. Rani SB, Rathod SS, Karthik S, Kaur N, Muzumdar D, Shiras AS. MiR-145 functions as a tumor-suppressive RNA by targeting Sox9 and adducin 3 in human glioma cells. Neuro-oncology. 2013;15(10):1302–1316. doi:10.1093/neuonc/not090
42. Liu S, Li X, Zhuang S. miR-30c impedes glioblastoma cell proliferation and migration by targeting SOX9. Oncol Res. 2019;27(2):165–171. doi:10.3727/096504018X15193506006164
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.