")
Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 14
Microbial Ecosystem in Diabetes Mellitus: Consideration of the Gastrointestinal System
Authors Yehualashet AS , Yikna BB
Received 9 February 2021
Accepted for publication 31 March 2021
Published 28 April 2021 Volume 2021:14 Pages 1841—1854
DOI https://doi.org/10.2147/DMSO.S304497
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Antonio Brunetti
Awgichew Shewasinad Yehualashet, Berhan Begashaw Yikna
Pharmacology and Toxicology Unit, Department of Pharmacy, College of Health Sciences, Debre Berhan University, Debre Berhan, Ethiopia
Correspondence: Awgichew Shewasinad Yehualashet
Pharmacology and Toxicology Unit, Department of Pharmacy, College of Health Sciences, Debre Berhan University, PO Box 445, Debre Berhan, Ethiopia
Tel +251 935450290
Email [email protected]
Abstract: Intestinal microbiota is established to be a crucial element in the control of human health, and keeping the symbiotic relationship between the human body and intestinal microbes will have paramount importance. A number of investigations illustrated that many chronic diseases are associated with intestinal micro-ecological disorders implying intestinal floras as an important component among the environmental factors, and perturbations in their composition are correlated with metabolic disorders, including obesity and diabetes mellitus (DM). Increased evidence suggests that alterations in the gut microbial ecosystem have been involved in part in the pathogenesis of both type 1 and type 2 DM. Short chain fatty acids (SCFAs), derived from microbiota, have been studied for their potential action in modulating CNS, gut barrier axis, and the immune system as a promising mechanism for the observed protective effects on diabetes pathogenesis. Besides, the role of bile acid (BA) stimulated receptors to have a significant role in liver metabolism, and pathophysiology of liver-based metabolic diseases has also been investigated. In the current review, we will try to summarize the correlation between intestinal microbiota and diabetes considering the existing current evidence revealing the role of gut microbiota in onset and disease progression.
Keywords: gut microbiota, intestinal flora, microbial ecosystem, diabetes mellitus
Introduction
Diabetes mellitus (DM) is a chronic hyperglycemic condition as a result of pancreatic β cell dysfunction or insulin resistance, known to cause macrovascular and microvascular complications.1,2 The cause of defect in carbohydrate, fat, and protein metabolism in DM is the compromised action of insulin on its targets.3,4
Genetic predisposition and environmental factors are among the crucial elements in the pathogenesis of DM. Out of the environmental factors accounting for the disease; gut microbiota has been regarded as a potential contributory factor associated with the disease. Gastrointestinal microbiome is established to be a key element in controlling human health, and maintenance of the symbiotic relationship between the human body and gut microbes will have paramount importance. Numerous investigations have revealed that many chronic diseases are related to intestinal microecological disorders implying intestinal flora as an important component among the environmental factors, and its modifications could lead to some sort of metabolic problems like diabetes and obesity.5,6
Gut Microbiota and Diabetes
Homeostasis can be maintained as a result of a mutual interaction between the human host and gut microbes, therefore, any dysregulation to the normal composition of commensal communities, the so-called ‘dysbiosis’ could weaken the homeostasis and contribute in the emergence of many autoimmune disorders. With the help of high-throughput sequencing technologies, aggregate evidence has illustrated that there are visible variations in the profile of intestinal microbial between type 1 diabetic patient and the controls, implying a close interaction between gut microbiota and diabetes. Consequently, a number of studies have suggested that changes in gut microbiota composition have got a place in the pathogenesis of the disease. However, whether these microbial changes are causal, responsive, or both are becoming an active area of investigations requiring scrutiny in the scientific community. In this review, we will discuss the association between intestinal microbiota and diabetes considering the existing animal and human evidence revealing the influence of the gut microbial ecosystem in onset and disease progression.5–7
The microbiota is regarded as microbial ecosystems living together in a symbiotic interaction with their host, sheltered on skin, respiratory, and urogenital systems and the intestinal tract. Among the variety of microorganisms found in the gut, bacteria are relatively best studied.7,8 There are around a hundred trillion cells of microbes in the human gut, which is 10-times higher compared to the number of human cells. The majority of microbiota of the gut are categorized into the following phyla: Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria. The intestinal microbiota so-called “hidden organ“ is involved invarious physiological functions such as fighting against pathogens, energy production, intestinal epithelial integrity maintenance, and immune regulation.9
Functionally, the vital role of the gut is enhancing digestion of food, absorption of nutrients, helps in the removal of waste products, and protecting against invasion by pathogenic bacteria. The protective mechanism is by acting as an impermeable barrier on intestinal epithelial cells along with other cells.7,10 In addition, the gut gives protection against pathogen overgrowth, impeding their aggregation by inhibiting adherence, bacteriocin generation and nutrient competition. It participates in the metabolism of drugs, detoxification of exogenous toxins, synthesis of essential vitamins like B1, B2, B5, B6, B12, K, folic acid, and deconjugation of bile acids (BA). Moreover, the ability to ferment indigestible carbohydrates helps them to provide 5–10% of daily energy need. Besides, in the intact colon, it aids intestinal repair through enhancing cellular proliferation and differentiation which have a substantial role in the maintenance of gut integrity.7,10
Their role in maturation and continued education of the host’s immunity has been pointed out with recent investigations. As a result of continued education, the immune system learned to identify the commensal from pathogenic bacteria where commensal bacteria can trigger an anti-inflammatory response, while pathogenic bacteria provoke a pro-inflammatory condition. Hence, gut microbiota regulates migration and function of neutrophils and affects T-cell differentiation, favoring differentiation and expansion of regulatory T-cells (Tregs), which are crucial components in mediating immune tolerance. The ability of controlling the immune system is via generation of short-chain fatty acids (SCFAs), like butyrate, acetate, and propionate that are produced by bacterial fermentation of non-digestible carbohydrates. In T lymphocytes, SCFAs are employed in stimulating G protein-coupled receptors (GPR41/GPR43) signaling cascades, inhibiting histone deacetylases, and eliciting metabolic changes via promoting the action of a mammalian target of rapamycin (mTOR) complex. Consequently, inflammatory cascades will be inhibited; triggering the expansion of mucosal Tregs, and generation of inflammatory cytokines like interleukin-10 (IL-10) and interferon- (IFN-) will be reduced. Provided that they have a key role in regulating health of the host, it is not amazing that dysbiosis could be implicated in the pathogenesis of various extra intestinal diseases including diabetes.7
Changes of Intestinal Flora in Diabetic Populations
Family history and environmental factors are key elements in the pathogenesis of diabetes. Among environmental factors, gut microbiota has been regarded as a potential contributory factor associated with type 1 diabetes.11,12 Studies showed that the microbiota has a linkage with weight gain and adiposity through many other related processes including energy harvest and formation of microbial metabolites, although their effect on inflammatory reactions and the gut–brain axis. One major metabolic function of gut microbiota is generating non-gaseous and absorbable SCFAs which are vital in modulating gut health and the immune system, promoting intestinal hormone production and lipogenesis.12
The anti-inflammatory effects of SCFAs is as a result of produced immunoglobulin A and immunosuppressive cytokines. Therefore, loss of early-life exposure because of continued use of antibiotics and a reduced fiber intake could end up with dysbiosis, which in turn allows the incidences of inflammatory problems, including diabetes. Several investigations have reported that the number of bacteria employed in generation of SCFA was significantly lowered in people with type 2 diabetes. SCFAs bind to GPCRs and result in enhancing the release of glucagon-like peptide-1 (GLP-1); which has an incretin effect and hinders glucagon release, prevents gluconeogenesis in the liver, ameliorates insulin sensitivity, and builds up central satiety, thereby resulting in bodyweight loss. Moreover, SCFAs can directly hamper the low-grade inflammatory response produced because of the migration of bacteria from the intestines into the mesenteric adipose tissue and the blood.13
Dietary fats promote absorption and removal from lipopolysaccharides (LPS) through intestinal/epithelial cells by inducing chylomicrons. LPS has an affinity for chylomicrons and is co-transported along with other dietary fats. Insulin insensitivity is associated with obesity and low-grade inflammation which is distinguished in part to alterations in gut microbiota.14 Different animal and human studies were conducted to evaluate whether changes are causal or merely a consequence of obesity or metabolic syndrome. To date, different animal and human investigations have been undertaken to evaluate the role of gut microbiota in the pathogenesis of both types of DM. In the table below, we have been tried to summarize the existing evidence showing the causal relationship between the disease and the gut microbiota.
Evidence on the Role of Gut Microbiota in the Pathogenesis of Type 1 and Type 2 DM
Current Evidence on the Role of Gut Microbiota in the Pathogenesis of Type 1 DM
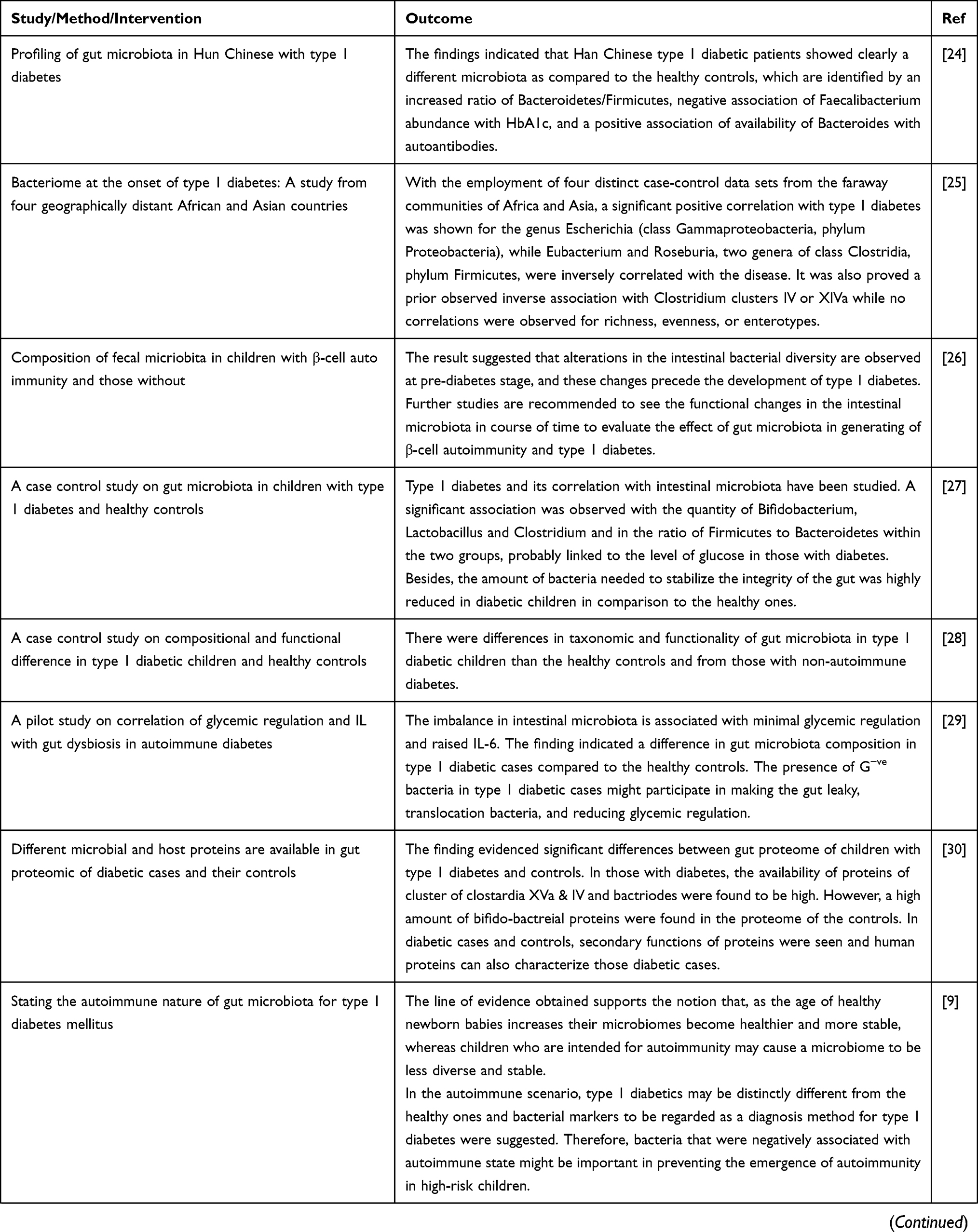
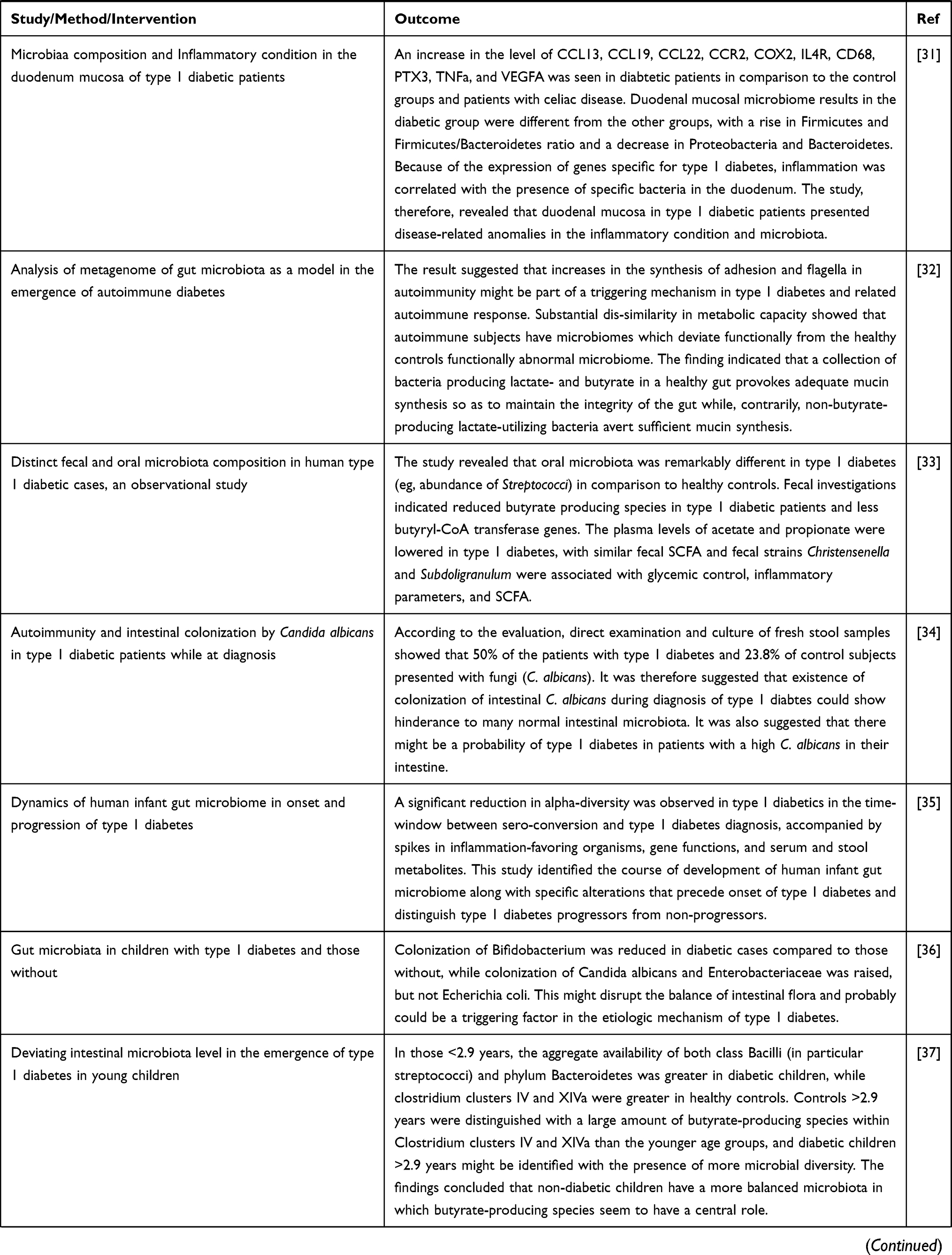
Until recently, the correlation between gut microbiota and pathogenesis of type 1 DM has been investigated in animal models, the condition may better be elaborated with various cohort studies to recognize the alteration of intestinal microbiota abundance prior to occurrence of the disease.
Prospective studies can give information regarding progression of the disease in time, whereas etiological association could be obtained from interventional studies. Investigations in huge lines of children will be crucial to prove the involvement of microbiome disturbance prior to the development of type 1 DM.7,9 The aggregate evidence summarizing preclinical and clinical trials conducted to explore the correlation of microbiota and type 1DM have been presented in Table 1 .
 |
 |
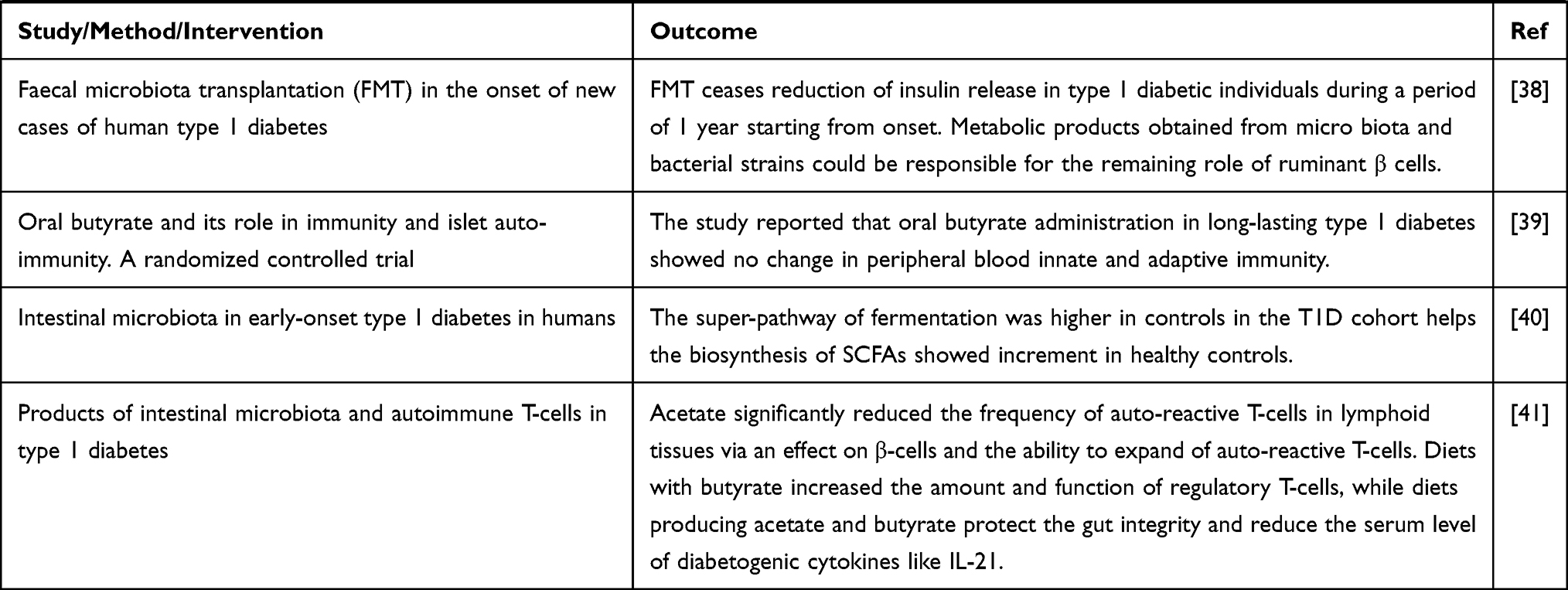 |
Table 1 Evidence from Animal and Human Studies Revealing the Causal Relation between Gut Microbiota and Type 1 Diabetes |
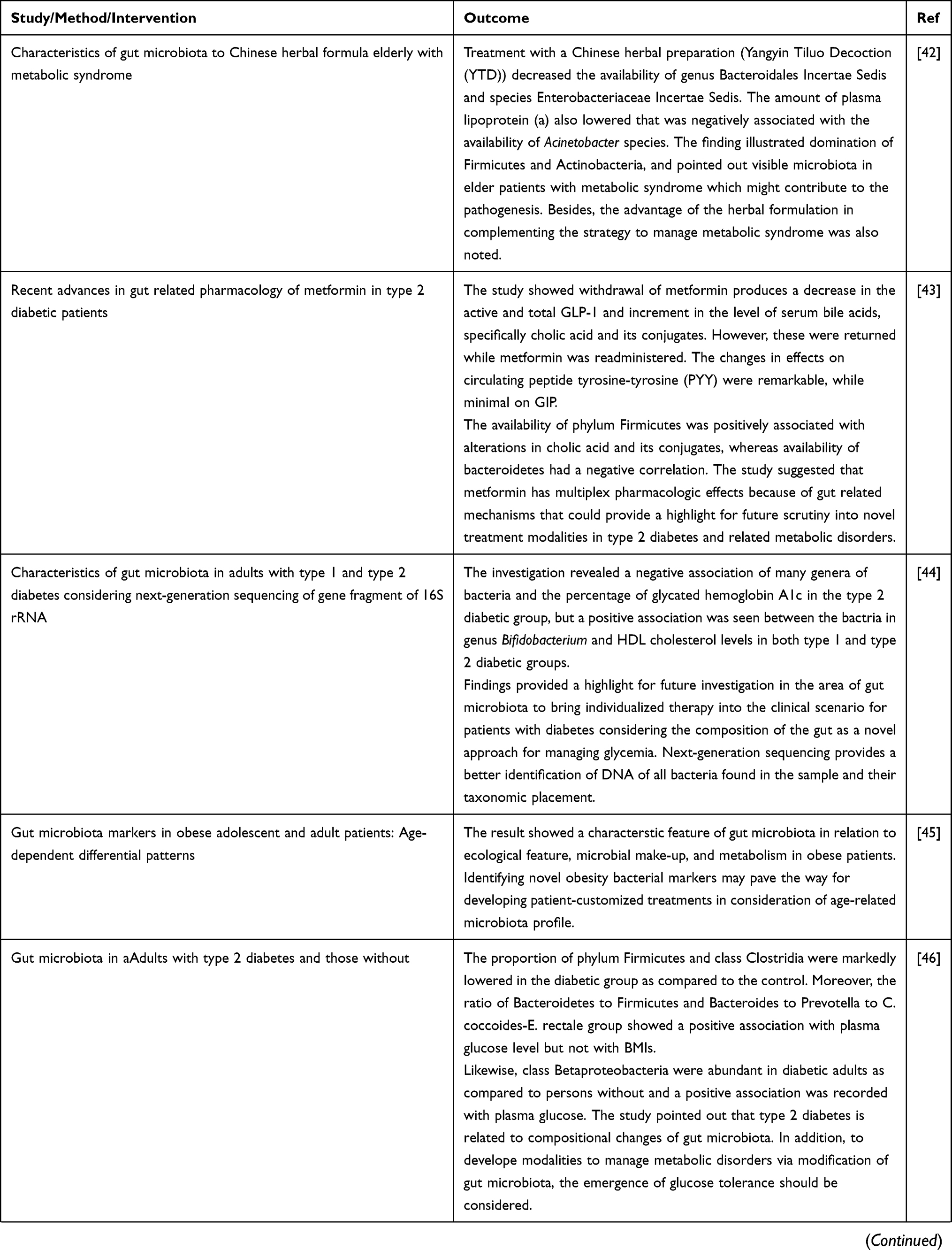
Current Evidence on the Role of Gut Microbiota in the Pathogenesis of Type 2 DM
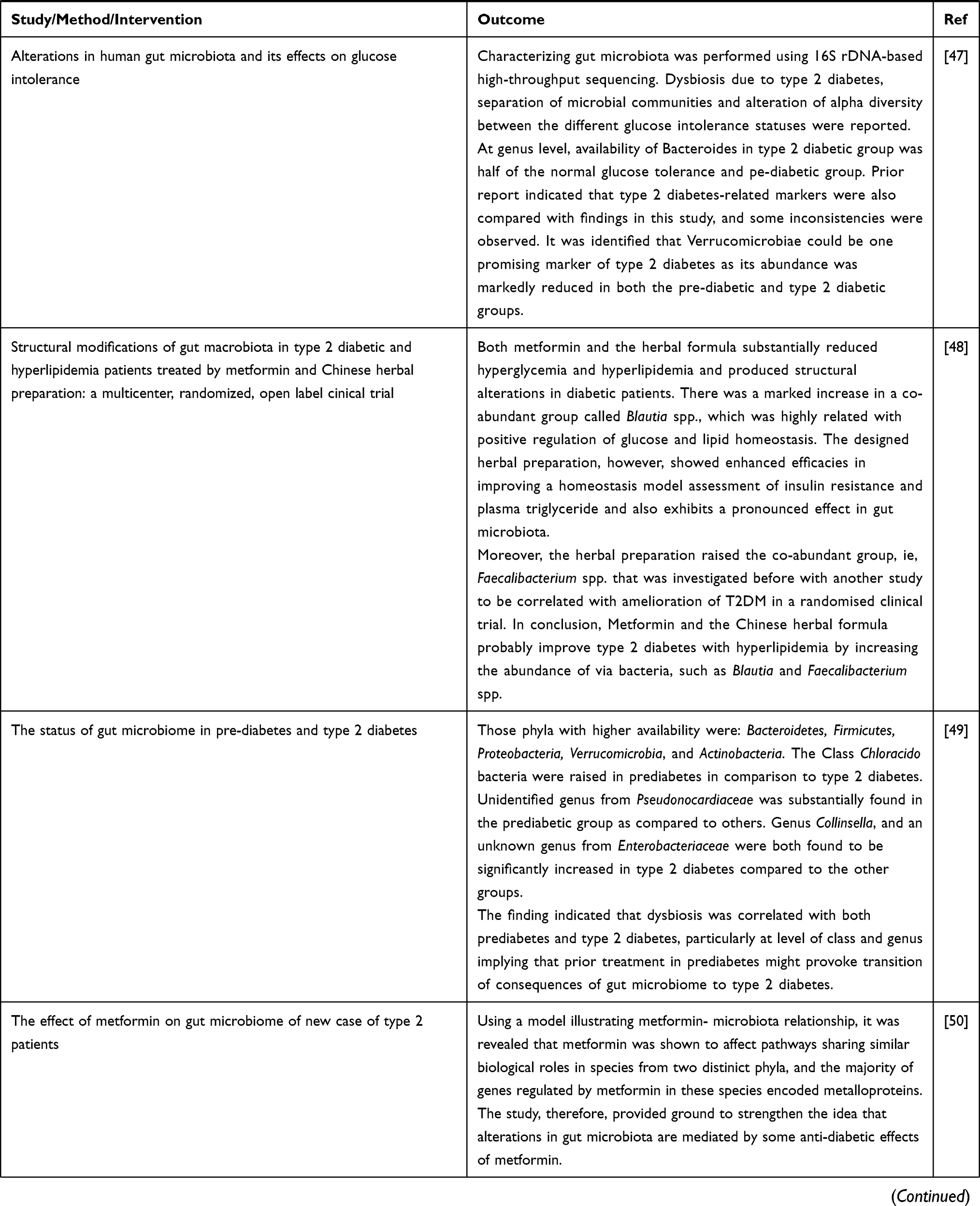
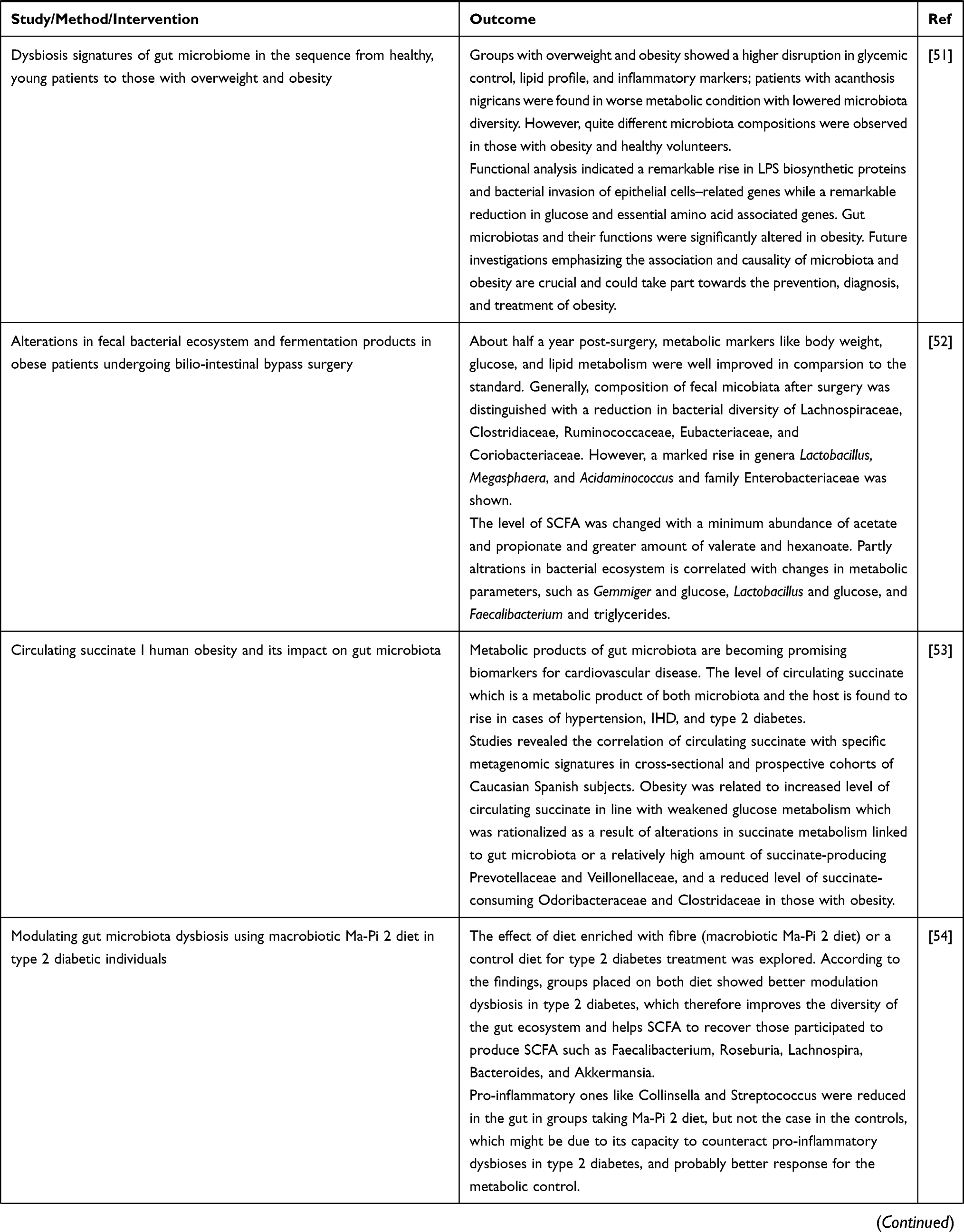
Alterations in the composition of intestinal microbiota is correlated with metabolic disease including type 2 DM. Different studies in animals showed various mechanisms contributing to the pathogenesis of the disease.12 Basically (as presented in Table 2), many of the investigations revealing the role of microbiota in glucose homeostasis are derived from studies in mice. But a change in the composition of microbiota has also been illustrated in a series of human cohort type 2 DM.
 |
 |
 |
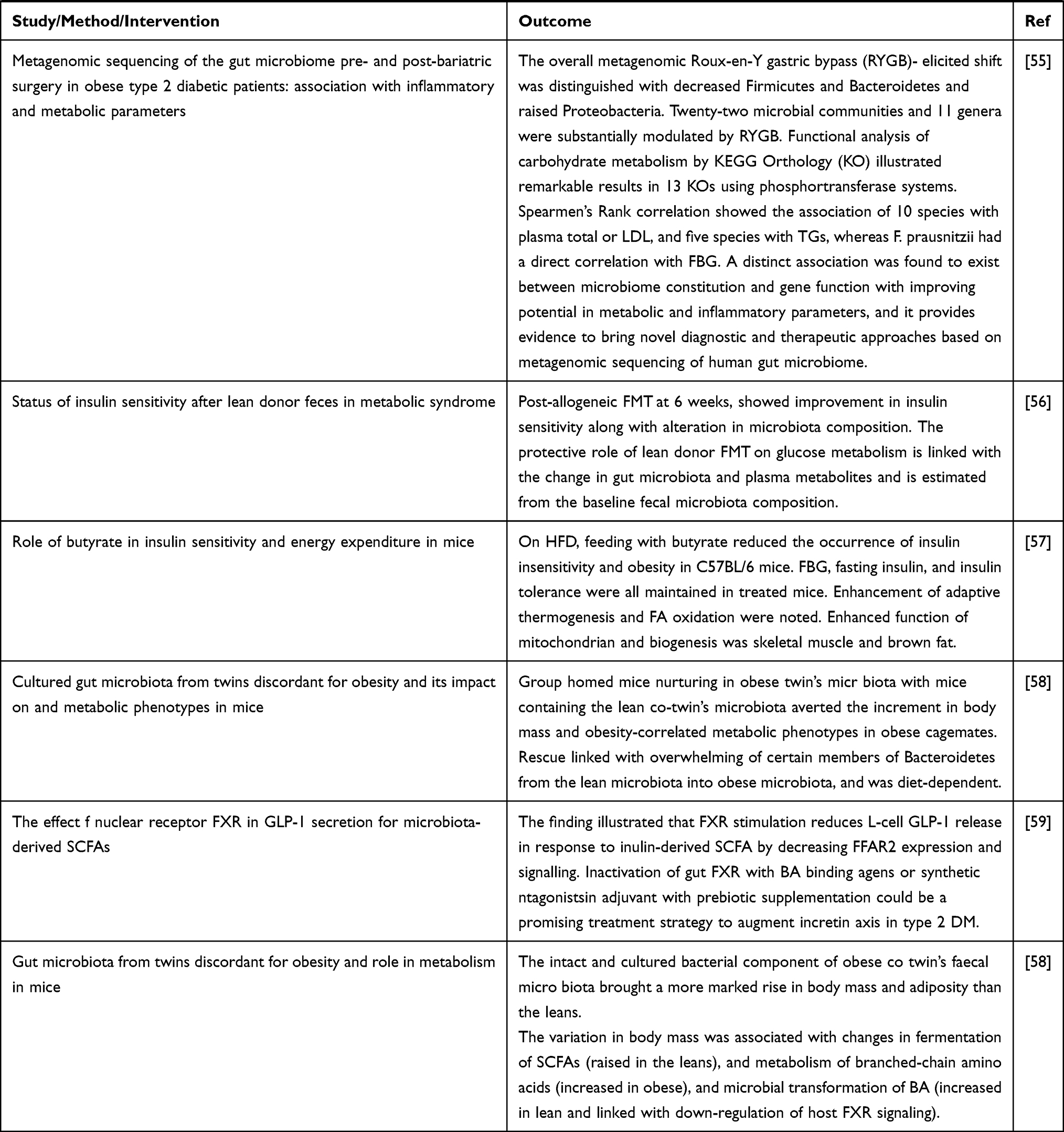 |
Table 2 Evidences from Animal and Human Studies Revealing the Causal Relation between Gut Microbiota and Type 2 Diabetes |
Possible Mechanisms of Gut Microbiota in Diabetes Pathogenesis
Mechanisms Involving SCFAs
According to a hypothesis by Trowell, dietary changes are blamed for diabetes, and increased use of processed carbohydrates with a low level of dietary fiber (DF) has been regarded as one potential risk factor for diabetes. Major DFs include soluble fibers, like pectin, inulin, arabinoxylan, and hemicellulose, and insoluble fibers such as cellulose. The soluble fibers can be fermented by microbiota to produce SCFAs acetate, propionate, and butyrate. The generation of SCFAs in distal gut is via bacterial fermentation of macrofibrous material that might get out of digestion by upper gastrointestinal tract and reach the colon (Figure 1). Current findings showed the role of SCFAs in modulating CNS, gut barrier axis, and the immune system which are assumed to be promising mechanisms for the observed protective effects of DF on diabetes pathogenesis. Recent findings indicated that microbiota is modulated by SCFAs for their protective activity on type 1 diabetes pathogenesis.15,16 Moreover, evidence suggested that protective potentials of SCFAs on β-cells and endogenous FFA2 expression probably accounted for this effect. Based on prior reports, it was concluded that sodium acetate and propionate reduced islet cell death elicited by cytokine or palmitate; FFA2 deficiency induced β-cell death and decreased β-cell mass, whereas SCFAs protected islet β-cells.17
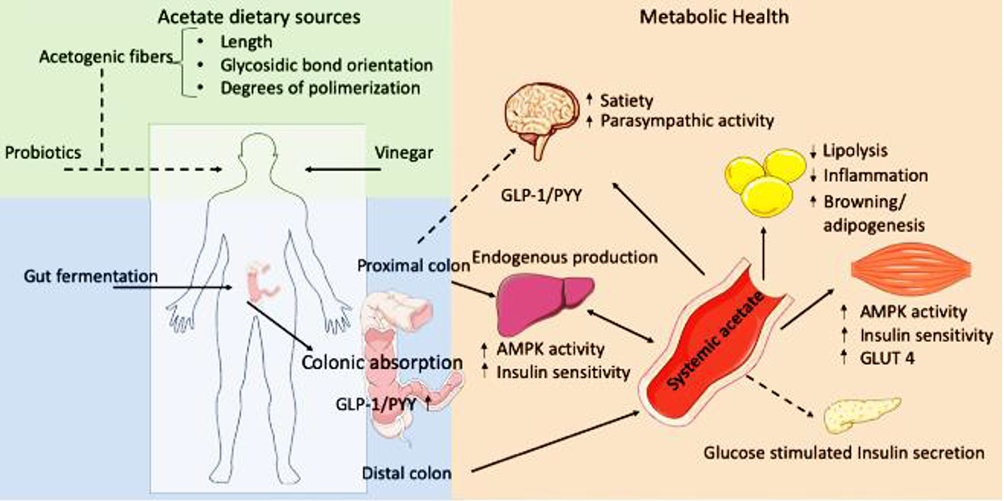 |
Figure 1 Acetate and effects in metabolic health. The solid lines show well studied effects of acetate, while broken lines imply more inconsistent findings. Reproduced from Hernández MAG, Canfora EE, Jocken JWE, Blaak EE. The Short Chain Fatty Acid Acetate in Body Weight Control and Insulin Sensitivity. Nutrients. 2019;11(1943).18 Copyright © 2019 by the authors. Licensee MDPI, Basel, Switzerland. Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/). |
Low grade inflammation and high level of cytokines like IL-6, IL-1, or TNF-α are characteristic features of Type 2 diabetes. These inflammatory markers are expressed in insulin target tissues, such as liver, adipose tissue, and muscles, hence leading to insulin insensitivity. It was investigated that changes in the gut microbial ecosystem provokes a pro-inflammatory condition in the adipose tissue that is related to obesity and resultant insulin resistance. The diversity of the gut microbiota, and its subsequent metabolic products is highly modified by diet changes; proper use of dietary fibers is usually correlated with a level of SCFA that could promote an anti-inflammatory effect, while a higher fat consumption was linked to reduced profile of SCFAs and raised LPS, implying aggregation of G−ve bacteria. LPS was found to elicit the generation of pro-inflammatory molecules that are taking part in enhancing permeability and inflammation in the gut epithelium, the so-called “metabolic endotoxemia“.16
Regulation of BGLs, insulin insensitivity, and GLP-1 release in improving metabolic function is as a result of expressed SCFA receptors in various tissues, hence are able to induce a protective effect provoked by them. The blood glucose lowering potential of SCFAs might be because of their beneficial effects such as: reduced inflammatory condition, hence decreases insulin insensitivity enhance decreased inflammatory state that reduces insulin resistance, simultaneously enhances the release of GLP-1 that promotes insulin release, and ameliorate β-cell function, thereby helping to maintain glucose homeostasis. Different preclinical and clinical trials showed that the acetate has a role in host energy and substrate metabolism through its positive effect on the release of GLP-1 and peptide YY, therefore affecting appetite by reducing lipolysis, pro-inflammatory mediators, and increasing utilization of energy and fat oxidation.16,18
Mechanisms Involving Bile Acids
BAs are metabolites of cholesterol catabolism in the liver and their synthesis accounts for about half of the daily cholesterol output, whereas 40% of the output is because of biliary secretion, and the remaining 10% is employed for membrane and steroid hormone synthesis. As a result, BA metabolism has a central role in regulating cholesterol homeostasis. Extensive research regarding BAs has created a ground for viewing them as signaling molecules that activate several nuclear receptors: farnesoid X receptor (FXR), vitamin D receptor (VDR), pregnane X receptor (PXR); and the membrane GPCRs: Takeda GPCR 5 (TGR5), sphingosine-1 phosphate receptor 2 (S1PR2), and muscarinic M2 receptor. These BA stimulated receptors have a significant role in liver metabolism, and the roles of FXR and TGR5 in regulating metabolism and pathophysiology of liver based metabolic diseases has been investigated.19,20 The human BA pool is determined by the enterohepatic cycle and microbial metabolism in the gut (Figure 2). BAs can undergo decongugations induced by a broad spectrum of gut bacteria. Therefore, regarding the level of BAs excreted in feces, the majority contain secondary BAs, and highly rely on gut microbiota metabolism.21
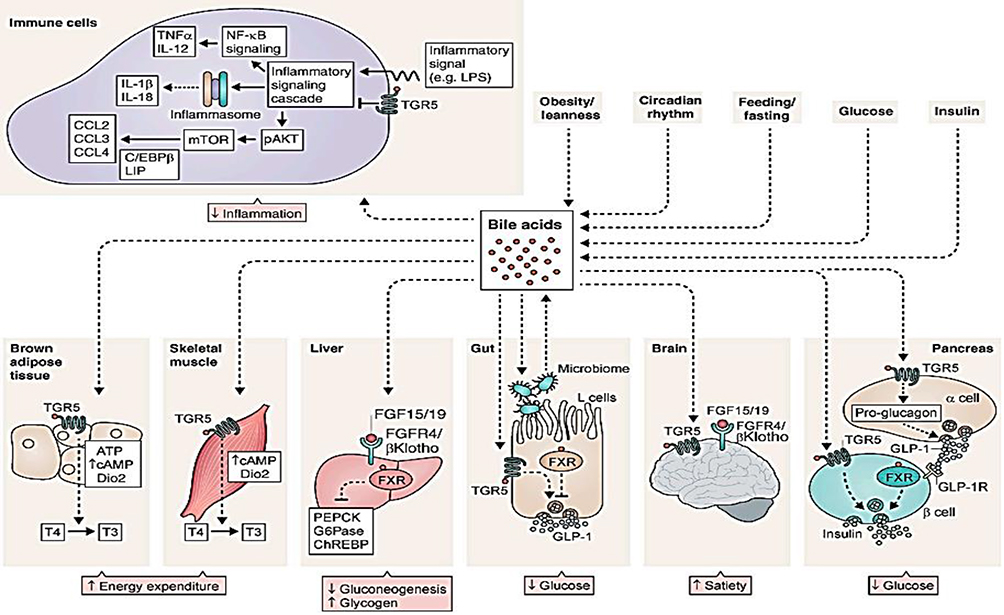 |
Figure 2 BA signaling controls the systemic glycemic response. Reproduced from Shapiro H, Kolodziejczyk AA, Halstuch D, Elinav E. Bile acids in glucose metabolism in health and disease. J ExpMed. 2018;215(2):383–396.20 Copyright © 2018 Shapiro et al. Creative Commons License (Attribution–Noncommercial–Share Alike 4.0 International license, as described athttps://creativecommons.org/licenses/by-nc-sa/4.0/). |
Recent data suggests that dysregulation in luminal enteral communication because of BA sequestrants could reveal the attractive effects of these drugs. Various gastrointestinal cells and hormones work together to contribute in neuroendocrine regulation of digestion and metabolic function. Detecting a way to augment endogenous GLP1 secretion, without increasing overall energy intake and deposition, is an attractive concept in the future treatment of type 2 diabetes – simultaneously improving our understanding of endocrine gut physiology.22 To date, the use of BA chelates are the only treatment modalities for diabetes involving BA, however, it was revealed that part of the advantages of bariatric surgery on controlling glucose level in diabetic patients is something associated with BA metabolism.23
In the brain, BA-TGR5 cascading regulates satiety. In skeletal muscles and brown adipose tissue, BA-TGR5 sensing enhances the conversion of T4 to T3, provoking a rise in energy expenditure. In the pancreas, both BA-TGR5 and BA-FXR signaling in β-cells stimulates insulin production. Glucose-induced insulin release is also enhanced by BA-TGR5 signaling in α-cells, that promotes proglucagon to be changed to GLP-1 and GLP-1 release. TGR5-BA in immune cells inhibits NLRP3-inflammasome and reduced inflammation.
Conclusion
The current review of aggregate evidence from animal and human investigations has shown that dysbiosis in gut microbial ecosystem and reduction of their diversity was associated with the onset and progression of both types of diabetes. The possible mechanisms include the generation of SCFAs in distal gut is via bacterial fermentation of macrofibrous material that might get out of digestion by upper gastrointestinal tract participates in modulating CNS, gut barrier axis, and immune system which are assumed to be promising mechanisms for the observed protective effects of SCFAs on diabetes pathogenesis. Furthermore, extensive research regarding BAs has created a ground for viewing them as signaling molecules that activate several nuclear receptors in regulating metabolism and pathophysiology of liver based metabolic diseases. Considering these findings, the research community should give emphasis on studies illustrating the significance of maintaining healthy microbial composition because related findings could be useful for developing strategies to control diabetes by modifying the gut microbiota. In fact, bringing these understandings into reasonable therapeutic measures is challenging. Therefore, in the future, ongoing randomized controlled trials in huge human cohorts are needed to uncover unresolved questions and prove the efficacy of microbiota-based therapeutic approaches.
Author Contributions
Both authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest for this work.
References
1. Punthakee Z, Goldenberg R, Katz P, et al. Definition, classification and diagnosis of diabetes, prediabetes and metabolic syndrome. Can J Diabetes. 2018;42:S10–S15. doi:10.1016/j.jcjd.2017.10.003
2. Ubaid M, Arif M, Usmani A, Anayatullah KN, Dash PP, Mishra A. Advances in clinical endocrinology and metabolism: an insight to diabetes mellitus and its complications. Adv Clin Endo Met. 2019;2(1):37–46.
3. Olokobam AB, Obateru OA, Olokoba LB. Type 2 diabetes mellitus: a review of current trends. Oman Med J. 2012;27(4):269–273. doi:10.5001/omj.2012.68
4. Baynes HW. Classification, pathophysiology, diagnosis and management of diabetes mellitus. J Diabetes Metab. 2015;6:541. doi:10.4172/2155-6156.1000541
5. Ma Q, Li Y, Li P, et al. Research progress in the relationship between type 2 diabetes mellitus and intestinal flora. Biomed Pharmacother. 2019;117:109138.
6. Xu H, Liu M, Cao J, et al. The dynamic interplay between the gut microbiota and autoimmune diseases. J Immunol Res. 2019.
7. Durazzo M, Ferro A, Gruden G. Gastrointestinal microbiota and type 1 diabetes mellitus: the state of art. J Clin Med. 2019;8:1843. doi:10.3390/jcm8111843
8. Wang H, Wei CX, Min L, Zhu LY. Good or bad: gut bacteria in human health and diseases. Biotechnol Biotechnol Equip. 2018;32(5):1075–1080. doi:10.1080/13102818.2018.1481350
9. Zhou H, Sun L, Zhang S, Zhao X, Gang X, Wang G. Evaluating the causal role of gut microbiota in type 1 diabetes and its possible pathogenic mechanisms. Front Endocrinol (Lausanne). 2020;11:125. doi:10.3389/fendo.2020.00125
10. Hsiao WW, Metz C, Singh DP, Roth J. The microbes of the intestine: an introduction to their metabolic and signalling capabilities. Endocrinol Metab Clin North Am. 2008;37(4):857–871. doi:10.1016/j.ecl.2008.08.006
11. Wen L, Duffy A. Factors Influencing the gut microbiota, inflammation, and type 2 diabetes. J Nutr. 2017;147:1468S–1475S. doi:10.3945/jn.116.240754
12. Lazar V, Ditu L-M, Pircalabioru GG, et al. Gut microbiota, host organism, and diet trialogue in diabetes and obesity. Front Nutr. 2019;6:21. doi:10.3389/fnut.2019.00021
13. Aw W, Shinji Fukuda S. Understanding the role of the gut ecosystem in diabetes mellitus. J Diabetes Investig. 2018;9:5–12. doi:10.1111/jdi.12673
14. Olden CV, Groen AK, Nieuwdorp M. Role of intestinal microbiome in lipid and glucose metabolism in diabetes mellitus. Clin Ther. 2015;37(6).
15. Kim CH. Microbiota or short-chain fatty acids: which regulates diabetes? Cell Mol Immunol. 2018;15:88–91. doi:10.1038/cmi.2017.57
16. Puddu A, Sanguineti R, Montecucco F, Viviani GL. Evidence for the gut microbiota short-chain fatty acids as key pathophysiological molecules improving diabetes. Mediators Inflamm. 2014;1–9. doi:10.1155/2014/162021
17. Liu JL, Segovia I, Yuan XL, Gao ZH. Controversial roles of gut microbiota-derived short-chain fatty acids (SCFAs) on pancreatic β-cell growth and insulin secretion. Int J Mol Sci. 2020;21:910. doi:10.3390/ijms21030910
18. Hernández MAG, Canfora EE, Jocken JWE, Blaak EE. The short chain fatty acid acetate in body weight control and insulin sensitivity. Nutrients. 2019;11:1943. doi:10.3390/nu11081943
19. Ferrell JM, Chiang JYL. Understanding Bile Acid Signaling in Diabetes: from Pathophysiology to Therapeutic Targets. Diabetes Metab J. 2019;43:257–272. doi:10.4093/dmj.2019.0043
20. Shapiro H, Kolodziejczyk AA, Halstuch D, Elinav E. Bile acids in glucose metabolism in health and disease. J Exp Med. 2018;215(2):383–396. doi:10.1084/jem.20171965
21. Molinero N, Ruiz L, Sánchez B, et al. Intestinal bacteria interplay with bile and cholesterol metabolism: implications on host physiology. Front Physiol. 2019;10:185. doi:10.3389/fphys.2019.00185
22. Sonne DP, Hansen M, Knop FK. Bile acid sequestrants in type 2 diabetes: potentialeffects on GLP1 secretion. European Journal of Endocrinology. 2014;171(2):R47–R65. doi:10.1530/EJE-14-0154
23. González-Regueiro JA, Moreno-Castañeda L, Uribe M, et al. The role of bile acids in glucose metabolism and their relation with diabetes. Ann Hematol. 2018;16(1):s15–s20. doi:10.5604/01.3001.0010.5672
24. Huang Y, Li S-C, Hu J, et al. Gut microbiota profiling in Han Chinese with type 1 diabetes. Diabetes Res Clin Pract. 2018;141:256–263. doi:10.1016/j.diabres.2018.04.032
25. Cinek O, Kramna L, Mazankova K, et al. The bacteriome at the onset of type 1 diabetes: a study from four geographically distant African and Asian countries. Diabetes Res Clin Pract. 2018;144:51–62. doi:10.1016/j.diabres.2018.08.010
26. De Goffau MC, Luopajarvi K, Knip M, et al. Fecal microbiota composition differs between children with b-cell autoimmunity and those without. Diabetes. 2013;62:1238–1244. doi:10.2337/db12-0526
27. Murri M, Leiva I, Gomez-Zumaquero JM, et al. Gut microbiota in children with type 1 diabetes differs from that in healthy children: a case-control study. +n. 2013;11:46.
28. Leiva-Gea I, Sánchez-Alcoholado L, Martín-Tejedor B, et al. Gut microbiota differs in composition and functionality between children with type 1 diabetes and MODY2 and healthy control subjects: a case-control study. Diabetes Care. 2018;41:2385–2395. doi:10.2337/dc18-0253
29. Higuchi BS, Rodrigues N, Gonzaga MI, et al. Intestinal dysbiosis in autoimmune diabetes is correlated with poor glycemic control and increased interleukin-6: a pilot study. Front Immunol. 2018;9:1689. doi:10.3389/fimmu.2018.01689
30. Pinto E, Anselmo M, Calha M, et al. The intestinal proteome of diabetic and control children is enriched with different microbial and host proteins. Microbiology. 2017;163:161–174. doi:10.1099/mic.0.000412
31. Brown CT, Davis-Richardson AG, Giongo A, et al. Gut microbiome metagenomics analysis suggests a functional model for the development of autoimmunity for type 1 diabetes. PLoS One. 2011;6(10):e25792. doi:10.1371/journal.pone.0025792
32. De Groot PF, Belzer C, Aydin Ö, et al. Distinct fecal and oral microbiota composition in human type 1 diabetes, an observational study. PLoS One. 2017;12(12):e0188475. doi:10.1371/journal.pone.0188475
33. Gürsoy S, Koçkar T, Atik SU, et al. Autoimmunity and intestinal colonization by Candida albicans in patients with type 1 diabetes at the time of the diagnosis. Korean J Pediatr. 2018;61(7):217–220. doi:10.3345/kjp.2018.61.7.217
34. Kostic AD, Gevers D, Siljander H, et al. The dynamics of the human infant gut microbiome in development and in progression towards type 1 diabetes. Cell Host Microbe. 2015;17(2):260–273. doi:10.1016/j.chom.2015.01.001
35. Soyucen E, Gulcan A, Aktuglu-Zeybek AC, et al. Differences in the gut microbiota of healthy children and those with type 1 diabetes. Pediatr Int. 2014;56:336–343. doi:10.1111/ped.12243
36. De Goffau MC, Fuentes S, van den Bogert B, et al. Aberrant gut microbiota composition at the onset of type 1 diabetes in young children. Diabetologia. 2014;57(8):1569–1577. doi:10.1007/s00125-014-3274-0
37. Giongo A, Gano KA, Crabb DB, et al. Toward defining the autoimmune microbiome for type 1 diabetes. Int Soc Microb Ecol. 2011;5:82–91.
38. de Groot P, Nikolic T, Pellegrini S, et al. Faecal microbiota transplantation halts progression of human new-onset type 1 diabetes in a randomized controlled trial. Gut. 2021;70:92–105. doi:10.1136/gutjnl-2020-322630
39. de Groot P, Nikolic T, Imangaliyev S, et al. Oral butyrate does not affect innate immunity and islet autoimmunity in individuals with longstanding type 1 diabetes: a randomised controlled trial. Diabetologia. 2020;63(3):597–610. doi:10.1007/s00125-019-05073-8
40. Vatanen T, Franzosa EA, Schwager R, et al. The human gut microbiome in early-onset type 1 diabetes from the TEDDY study. Nature. 2018;562:589–594. doi:10.1038/s41586-018-0620-2
41. Mariño E, Richards JL, McLeod KH, et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat Immunol. 2017;18(5):552–562. doi:10.1038/ni.3713
42. Ni Y, Mu C, He X, et al. Characteristics of gut microbiota and its response to a Chinese Herbal Formula in elder patients with metabolic syndrome. Drug Discov Ther. 2018;12(3):161–169. doi:10.5582/ddt.2018.01036
43. Napolitano A, Miller S, Nicholls AW, et al. Novel gut-based pharmacology of metformin in patients with type 2 diabetes mellitus. PLoS One. 2014;9(7):e100778. doi:10.1371/journal.pone.0100778
44. Salamon D, Sroka-Oleksiak A, Kapusta P, et al. Characteristics of gut microbiota in adult patients with type 1 and type 2 diabetes based on next-generation sequencing of the 16S rRNA gene fragment. Pol Arch Intern Med. 2018;128(6):336343. doi:10.20452/pamw.4246
45. Del Chierico F, Abbatini F, Russo A, et al. Gut microbiota markers in obese adolescent and adult patients: age-dependent differential patterns. Front Microbiol. 2018;9:1210. doi:10.3389/fmicb.2018.01210
46. Larsen N, Vogensen FK, van den Berg FWJ, et al. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS One. 2010;5(2):e9085. doi:10.1371/journal.pone.0009085
47. Zhang X, Shen D, Fang Z, et al. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS One. 2013;8(8):e71108. doi:10.1371/journal.pone.0071108
48. Tong X, Xu J, Lian F, et al. Structural alteration of gut microbiota during the amelioration of human type 2 diabetes with hyperlipidemia by metformin and a traditional Chinese herbal formula: a multicenter, randomized, open label clinical trial. mBio. 2018;9(3):e02392–17. doi:10.1128/mBio.02392-17
49. Lambeth SM, Carson T, Lowe J, et al. Composition, diversity and abundance of gut microbiome in prediabetes and type 2 diabetes. J Diabetes Obes. 2015;2(3):1–7. doi:10.15436/2376-0949.15.031
50. Wu H, Esteve E, Tremaroli V, et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat Med. 2017;23(7):850–858. doi:10.1038/nm.4345
51. Renyuan Gao R, Zhu C, Li H, et al. Dysbiosis signatures of gut microbiota along the sequence from healthy, young patients to those with overweight and obesity. Obesity. 2018;26(2):351–361. doi:10.1002/oby.22088
52. Patrone V, Vajana E, Minuti A, et al. Postoperative changes in fecal bacterial communities and fermentation products in obese patients undergoing bilio-intestinal bypass. Front Microbiol. 2016;7:200. doi:10.3389/fmicb.2016.00200
53. Serena C, Ceperuelo-Mallafré V, Keiran N, et al. Elevated circulating levels of succinate in human obesity are linked to specific gut microbiota. ISME J. 2018;12:1642–1657. doi:10.1038/s41396-018-0068-2
54. Candela M, Biagi E, Soverini M, et al. Modulation of gut microbiota dysbioses in type 2 diabetic patients by macrobiotic Ma-Pi 2 diet. Br J Nutr. 2016;116:80–93. doi:10.1017/S0007114516001045
55. Graessler J, Qin Y, Zhong H, et al. Metagenomic sequencing of the human gut microbiome before and after bariatric surgery in obese patients with type 2 diabetes: correlation with inflammatory and metabolic parameters. Pharmacogenomics J. 2013;13(6):514–522. doi:10.1038/tpj.2012.43
56. Kootte RS, Levin E, Salojärvi J, et al. Improvement of insulin sensitivity after lean donor feces in metabolic syndrome is driven by baseline intestinal microbiota composition. Cell Metab. 2017;26:611–619. doi:10.1016/j.cmet.2017.09.008
57. Gao Z, Yin J, Zhang J, et al. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes. 2009;58:1509. doi:10.2337/db08-1637
58. Ridaura VK, Faith JJ, Rey FE, et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science. 2013;341(6150). doi:10.1126/science.1241214
59. Ducaste S, Touche V, Trabelsi M-S, et al. The nuclear receptor fXR inhibits glucagon-like peptide-1 secretion in response to microbiota-derived short-chain fatty acids. Sci Rep. 2020;10(1):174. doi:10.1038/s41598-019-56743-x
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.