")
Back to Journals » Infection and Drug Resistance » Volume 13
Methicillin-Resistant Staphylococcus aureus (MRSA): One Health Perspective Approach to the Bacterium Epidemiology, Virulence Factors, Antibiotic-Resistance, and Zoonotic Impact
Authors Algammal A , Hetta HF , Elkelish A , Alkhalifah D , Hozzein W , Batiha G, El Nahhas N, Mabrok M
Received 23 July 2020
Accepted for publication 19 August 2020
Published 22 September 2020 Volume 2020:13 Pages 3255—3265
DOI https://doi.org/10.2147/IDR.S272733
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sahil Khanna
Abdelazeem M Algammal,1 Helal F Hetta,2,3 Amr Elkelish,4 Dalal Hussien H Alkhalifah,5 Wael N Hozzein,6,7 Gaber El-Saber Batiha,8 Nihal El Nahhas,9 Mahmoud A Mabrok10,11
1Department of Bacteriology, Immunology and Mycology, Faculty of Veterinary Medicine, Suez Canal University, Ismailia 41522, Egypt; 2Department of Medical Microbiology and Immunology, Faculty of Medicine, Assuit University, Assuit 71515, Egypt; 3Department of Internal Medicine, University of Cincinnati, College of Medicine, Cincinnati, OH 45267-0595, USA; 4Botany Department, Faculty of Science, Suez Canal University, Ismailia 41522, Egypt; 5Biology Department, College of Science, Princess Nourah Bint Abdulrahman University, Riyadh 11451, Saudi Arabia; 6Bioproducts Research Chair, Zoology Department, College of Science, King Saud University, Riyadh 11451, Saudi Arabia; 7Botany and Microbiology Department, Faculty of Science, Beni-Suef University, Beni-Suef 62511, Egypt; 8Department of Pharmacology and Therapeutics, Faculty of Veterinary Medicine, Damanhour University, Damanhour 22511, AlBeheira, Egypt; 9Department of Botany and Microbiology, Faculty of Science, Alexandria University, Alexandria 21515, Egypt; 10Fish Diseases and Management, Faculty of Veterinary Medicine, Suez Canal University, Ismailia 41522, Egypt; 11Fish Infectious Diseases Research Unit (FID RU), Faculty of Veterinary Science, Chulalongkorn University, Bangkok 10330, Thailand
Correspondence: Abdelazeem M Algammal
Department of Bacteriology, Immunology and Mycology, Faculty of Veterinary Medicine, Suez Canal University, Ismailia 41522, Egypt
Tel +201010667290
Email [email protected]
Abstract: Methicillin-resistant Staphylococcus aureus (MRSA) is a major human pathogen and a historically emergent zoonotic pathogen with public health and veterinary importance. In humans, MRSA commonly causes severe infectious diseases, including food poisoning, pyogenic endocarditis, suppurative pneumonia, otitis media, osteomyelitis, and pyogenic infections of the skin, soft tissues. In the horse, MRSA could cause a localized purulent infection and botryomycosis; in cattle and ewe, localized pyogenic infection and severe acute mastitis with marked toxemia; in sheep, abscess disease resembles caseous lymphadenitis caused by anaerobic strains; in dogs and cats, pustular dermatitis and food poisoning; in pig, exudative epidermatitis “greasy pig disease; in birds, MRSA causes bumble-foot. The methicillin resistance could be determined by PCR-based detection of the mecA gene as well as resistance to cefoxitin. In Egypt, MRSA is one of the important occasions of subclinical and clinical bovine mastitis, and the prevalence of MRSA varies by geographical region. In this review, we are trying to illustrate variable data about the host susceptibility, diseases, epidemiology, virulence factors, antibiotic resistance, treatment, and control of MRSA infection.
Keywords: MRSA, One Health Approach, pathogenicity, virulence factors, epidemiology, antimicrobial resistance, treatment
Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) is considered as a historic emergent zoonotic pathogen with public health and veterinary importance. S. aureus causes serious problems in both humans and animals and could resist adverse environmental conditions such as sunlight and desiccation.1,2 The bacterium could invade the skin, mucous membranes and internal organs causing serious illness in both animal and human, including suppurative infections of the skin, acne, osteomyelitis, endocarditis, respiratory tract infection, and septicemia.3–5 In addition, S. aureus is one of the principal occasions of cattle mastitis.6,7
Staphylococcus aureus is a Gram-positive, spherical shaped microorganism, non-motile, non-spore former, and some strains are capsulated. The first isolation is carried out by Alexander Ogston during the investigation of the septicemia and wound infection bacteria in 1880, the microscopical examination of 88 pus specimens revealed the presence of Gram-positive cocci (S. aureus).8,9 The majority of S. aureus strains (94%) are markedly reluctant to penicillin and its derivatives due to the release of penicillinase enzyme.10–12 Certain strains of S. aureus are resistant to methicillin, which has been identified as MRSA.13 The methicillin resistance could be determined clinically by PCR based detection of the mecA gene as well as resistance to cefoxitin. The mecA gene mainly encodes for the penicillin-binding protein (PBP-2A) which is responsible for this kind of antimicrobial resistance.14,15 MRSA are virulent zoonotic biovars of S. aureus, which exhibited specific criteria of being cefoxitin and methicillin-resistant. Various phenotypic and molecular patterns could distinguish between methicillin-susceptible S. aureus (MSSA) and MRSA. MRSA always shows a multidrug-resistant pattern, not only for penicillin but also for variable antimicrobial classes including; macrolides, fluoroquinolones, aminoglycosides, tetracyclines, and lincosamides.16–18 MRSA could induce severe infectious diseases in human including; pyogenic endocarditis, suppurative pneumonia, otitis media, osteomyelitis, pyogenic infections of the skin, soft tissues, and septic arthritis. The emergence of multidrug-resistant virulent MRSA strains is a remarkable public health problem.19
MRSA is known as the major cause of hospital-acquired infections (HA-MRSA) and community-acquired infections (CA-MRSA); therefore, MRSA is not only known as a nosocomial bacterium. The morbidity of CA-MRSA infections has elevated all over the world. CA-MRSA is genetically different from HA-MRSA by possessing a small type of SCCmec, and the frequent production of Panton-Valentine leukocidin, and cytotoxin. CA-MRSA strains are restricted people outside the health care practice and are usually cause mild infections such as skin and soft tissue infections. However, recent epidemic-molecular investigations reported that CA-MRSA could affect several patients within health care settings. Another type of MRSA is livestock-associated MRSA (LA-MRSA), which affects a wide variety of domestic animals. The widespread improper use of antibiotics in the veterinary sector results in the emergence of MRSA among livestock. This suggesting that the infected animal is a permanent reservoir of MRSA, which results in human infections.4,5,8 This review presents one health perspective approach to MRSA that emphasis on host susceptibility, diseases, the bacterium epidemiology, virulence factors, antibiotic resistance, treatment, and control of MRSA infection.
Host Susceptibility
MRSA could invade mammary glands, mucous membranes, serous membranes, skin, and internal organs of both man and different animals (cattle, chicken, horse, dog, pig, and cat) causing severe illness which is mainly associated with multiple antibiotic resistance.20 The first isolation of MRSA in farm animals all over the world was reported in the 1970, where MRSA strains were firstly isolated from mastitic dairy cows in Belgium.21
S. aureus is a popular human pathogen. In about 33% of seemingly healthy cases, S. aureus is colonized with the front nares, which are the main site. Moreover, the skin, throat, and digestive system are the most common extra-nasal sites of colonization. Several studies categorized the individuals into; non-carriers, intermittent and persistent carriers.22–24 The threat of emerging healthcare-associated S. aureus cases is five times elevated between the nasal carriers when compared to the non-carrier individuals. Also, the nasal colonizing strains are the most frequent causes of infection.25,26 The community-associated MRSA strains were found to be colonized at various body sites other than the front nares, as 23% of MRSA were colonized at the inguinal regions,27 in children-patients the rectum was the main organ of colonization,28 while in young cases, the throat was the main site S. aureus colonization (30%).29 Furthermore, in intensive care units (ICUs), the rectum and throat are found to be the main site of MRSA colonization.30
Virulence Factors of MRSA
Capsular Polysaccharides
Capsular polysaccharides are polysaccharide polymers that surround the cell wall of MRSA. It has been reported that 76–90% of clinical MRSA isolates produce capsular polysaccharides and 11 serologically distinct capsular polysaccharides types (CP 1-CP11) have been identified. Capsular polysaccharides enhance the virulence of S. aureus by impairing complement and antibody-mediated opsonization and inhibiting phagocytosis.31–33
Surface Associated Proteins
Staphylococcal Protein-A
Staphylococcal protein A is a cell wall structure that attaches to the circulating IgG, prevents the complement system opsonization and protects the microorganism from the phagocytosis process.34,35
Clumping Factors
Fibrinogen is a fundamental component of extracellular matrix protein in the host body. The S. aureus-fibrinogen adherence is initiated by clumping factor molecules that are present on the MRSA cell surface. Two distinct Clf proteins (Clf A and Clf B) characterized MRSA. Clf A present on the surface of MRSA in all phases of growth, while Clf B mainly detectable in the early exponential phase of growth.36–39
Extracellular Toxins
Staphylococcal Hemolysins
MRSA strains have been shown to express alpha, beta, gamma, and delta toxins, although different strains may vary in level in their production. Among these toxins, Alpha toxin is produced by most pathogenic MRSA strains and considered a major virulence factor.40 Alpha toxin could induce mammary gland necrosis and higher mortality rates among the infected animals.41,42 Enterotoxins produced by MRSA are mainly incriminated in food poisoning in both humans and animals.43–45
Staphylococcal Enterotoxins
Staphylococcal enterotoxins are pyrogenic exotoxins that belong to S. aureus superantigens. Enterotoxin types A, B, C, D, and E, are the main virulence factors incriminated in food poisoning in humans, especially SEA.46,47 The bacterial enterotoxins act as superantigens (SAgs) that stimulate the expression of IL-4 and IL-10 genes with subsequent activation of TH2 cells, resulting in suppress the clearance of invading pathogens. Panton-Valentine leucocidin, which is produced by MRSA, can pass through the blood-brain barrier causing serious damage to the cell membrane of human polymorphonuclear cells.48–50
Panton-Valentine Leukocidin
Panton-Valentine leukocidin is a powerful staphylococcal exotoxin and its activity is mediated by the action of two secretory proteins; F and S types. In human, PVL destroy the plasma-membrane of the polymorph-nuclear cells. In addition, PVL stimulates the release of the oxygen-metabolites from the polymorph-nuclear cells, the release of interleukin 8, the production of lysozymes, and the histamine-release from the human basophils. Moreover, the injection of PVL in rabbits induces complicated inflammatory reactions, necrotic lesions, degranulation of basophils, and leukocytic-infiltration. The lukS-PV and lukF-PV genes are found in the staphylococcal chromosome and are encoded for the PVL production. The bacteriophage meditated transfer of the lukS-PV and lukF-PV genes is carried out by the PVL-phages (such as; fSLT) that could infect the PVL-negative strains, resulting in the toxin-release.51
Toxic Shock Syndrome Toxin
Toxic shock syndrome toxin (TSST) is one of the most potent superantigens of S. aureus that incriminated in toxic shock syndrome (TSS) in humans. The disease occurred due to the release of TSST that activates the production of TNF-α, IL-1, and IL-2. Toxic shock syndrome is a fatal disease accompanied by high morbidity and mortalities. TSST commonly affects women during the menstruation, frequently in the 2nd and 3rd days. During this period, S. aureus grows well in the vagina and produce the toxin. The disease characterized by both systemic and gastrointestinal disorders includes fever, headache vomiting, abdominal pain, profuse diarrhea, generalized myalgias.24,34
Staphylococcal Exfoliative Toxins
Chemically, staphylococcal exfoliative toxins (ETs) are serine proteases in nature. ETs incriminated in staphylococcal scalded skin syndrome (SSSS) in humans that mainly involve the infants and neonates. Besides, Adults persons that suffer from immune-deficiency and or renal dysfunction could be affected. The disease is characterized by loss of superficial layers of skin, dryness, blistering of the skin with subsequent secondary bacterial infections. The toxins usually destroy the desmoglein-1 protein, which results in the sloughing of the epidermis.5,34
Extracellular Enzymes
Staphylococcal Coagulase
In 1903, S. aureus is reported to cause coagulation of human plasma due to the release of the coagulase (Coa) enzyme. Coagulase enzyme is chromosomally encoded and could coagulate both rabbit and human plasma. Up-to-date, about twelve various isoforms of coagulase are recognized, with remarkable sequence diversity (more than 50%), especially in the N-terminus coding domain among different MRSA isolates. Moreover, the Coagulase enzyme has a D1D2-domain that is specific for prothrombin-binding.40,41
Staphylokinase
Staphylokinase is an extracellular enzyme that stimulates the plasminogen for the lyses of fibrin clot with subsequent bacterial propagation. The enzyme released by lysogenic strains of MRSA. Besides, certain types of prophage encoding for staphylokinases such as serotype B phages and some serotype F phages. Staphylokinase is mainly species-specific and usually is active for dog, rabbit, sheep, goat, and human plasma, while it is inactive for buffaloes, pig, cow, and mouse plasma. Moreover, one of the favorable effects of staphylokinase is to minimize biofilm formation; therefore, staphylokinase-producing MRSA strains usually accompanied by less biofilm development in either in vitro or non-invasive infections of humans.3,5,41
Staphylococcal Nucleases
In 1956 Staphylococcal nuclease (DNase) was firstly recognized culture filtrates of S. aureus. Staphylococcal nuclease is heat-stable and Ca2+ dependent. Two types are found: endo- and exo-nuclease that destroy the DNA and RNA substrates. The total genome sequencing of MRSA revealed two various types of staphylococcal nuclease genes: nuc (SA0746) and nuc2 (SA1160) the main difference between Nuc and Nuc2 is the cell-localization: nuc is an extracellular enzyme with two various isoforms, NucB and NucA, while nuc2 is surface-bound.41–48
Staphylococcal Proteases
There are three types of staphylococcal proteases: 1-serine proteases, 2-metalloproteases, and 3-cysteine proteases. They play a significant role in host-defense evasiveness and bacterial dissemination. Up to date, about 12 various types of proteases are recognized including: one metalloprotease (aureolysin/Aur), 2 cysteine proteases (staphopain A, and staphopain B), and 9 various serine proteases.7–10,41
Staphylococcal Hyaluronidase
Hyaluronic acid is a polysaccharide molecule that constituted of repeating units of N-acetyl-glucosamine and glucuronic acid, linked by alternating β−1,3 and β−1,4 glycosidic bonds. It is a major constituent of extracellular matrices in mammals, responsible for the cell and tissue integrity as well as the host-immune regulation. Staphylococcal Hyaluronidase “spreading factor” is responsible for the breakdown of Hyaluronic acid into di-saccharides in both extracellular matrices and biofilms with subsequent bacterial dissemination.32,33,41
The Epidemiology of MRSA in Egypt
Unfortunately, in Egypt, limited epidemiological molecular surveys of MRSA-infections are carried out; only sporadic molecular studies are performed. In Egypt, the prevalence of MRSA bacteria varies by geographical region. A high incidence of MRSA was recorded in ICU-patients at Alexandria-University hospital (up to 75%). Besides, the prevalence of MRSA was 47.9% in hospitalized patients at two university hospitals in Cairo, and. In contrast, a lower prevalence (24.4%) was reported in hospitalized cases at AL-Minia-University hospital.51
MRSA strains are mostly recovered after surgery due to infection of the site of surgery and soft tissues. The prevalence of MRSA strains was estimated in 208 patients after surgeries at AL-Minia University Hospital, AL-Minia Province, Egypt. The bacteriological investigation proved that the prevalence of S. aureus was 61%, while the prevalence of MRSA was 15% in examined cases. Polymerase chain reaction revealed that the isolated MRSA strains are harbored the mecA gene. The isolated strains exhibit multiple drug resistance to cephalexin, cefotaxime, amoxicillin-clavulanic acid, ampicillin-sulbactam, erythromycin, tetracycline, and clindamycin.52
MRSA strains were retrieved from healthcare-associated as well as community-associated Staphylococcus aureus infected cases. The isolated strains were tested against 11 antimicrobial agents. MRSA was characterized in 343 (76.6%) and 21 (11.5%) of HA and CA S. aureus strains, respectively. Emerging of the multidrug-resistance was noticed in 85.8% of HA-MRSA and 48.6% of CA-MRSA. The elevated prevalence of HA-MRSA is frightening and impetus for proper infection control and continuous surveillance of the antibiotic resistance pattern.53
Community-acquired MRSA is an important threat to both public and veterinary health. A previous study was performed to investigate the prevalence of MRSA infection in cats, dogs, and their owners.54 In this study, the microbiological examination revealed that the prevalence of MRSA was 3.6% and 2.9% in the contact persons and the examined dogs, respectively. Also, two MRSA strains isolated from diseased dogs were found to be hospital-associated MRSA (HA-MRSA), while other isolates originated from apparently healthy dogs and human cases are defined as community-acquired MRSA (CA-MRSA). The authors concluded that the existence of MRSA in pet animals is considered a public health threat as it could spread in the community. The recurrent emergence of MRSA is regarded as a critical public health concern in hospitals. Moreover, limited data are available about the prevalence of MRSA in Egypt. The epidemiology of MRSA was investigated in the Patients of the ENT clinics (between April 2014 to April 2015), at Tanta University teaching hospital, Egypt. The prevalence of MRSA was 26% (n = 42) were identified from 160 S. aureus strains, which were retrieved from otitis media patients. The recovered MRSA strains are resistant to the β-lactam antibiotics, which is mainly attributed to the existence of the mecA gene. Most of the isolated MRSA strains (90.5%) are harbored the mecA gene. The continuous surveillance of MRSA is essential to control and prevent the occurrence of infection.55
S. aureus is one of the important occasions of subclinical and clinical bovine mastitis in Egypt. Algammal et al,56 reported that the prevalence of S. aureus in subclinical bovine mastitis was 35.9%; out of them 35.7% were MRSA strains. PCR revealed that the isolated strains are positive for enterotoxin A (30%) and enterotoxin C (10%) genes. Moreover, Enany et al,57 revealed that the prevalence of MRSA bovine clinical mastitis was 28.2%. The high prevalence of MRSA in raw milk of clinically healthy animals is of a great public health concern since the virulent MRSA strains could be transmitted to the human consumers causing serious illness and food poisoning.56
Fish is considered a major source of animal protein with high nutritive value and a better conversion rate.58–60 Staphylococci are not a normal fish microbiota; however, their presence could indicate post-harvest contamination due to bad personal hygiene, or fish disease. A previous study of Hammad et al61 reported that 174 out of 200 specimens of raw fish (sashimi) that gathered from the Japanese prefecture of Hiroshima were identified as methicillin-susceptible Staphylococcus aureus (MSSA), moreover, 10 strains of MRSA were retrieved from 10 examined specimens (5%, 10/200). The prevalence of S. aureus in catfish in Egypt was very high (46.6%), of which 21.4% were found to be MRSA strains. The isolated MRSA strains are multiple-drug resistant and pose 3 virulent (meca, coa and spa) genes. Virulent MRSA strains were isolated from freshwater catfish in Egypt as a first record, which gave serious concern to its public health importance.62 Foods that are widely handled during processing, and kept under erroneous refrigeration are predominantly accompanied with food poisoning with Staphylococcus. Saklani et al63 examined the survivability of MRSA in two seafood products under variable storage conditions and found that MRSA survival depends entirely on storage temperature and the inoculum level.
MRSA in Slaughterhouses and Food Products
MRSA could disseminate in the food processing plants and slaughterhouses. In the slaughterhouse, the source of contamination with MRSA was due to the movement of animals and the labors,64 while in the food processing plants, bad sanitary conditions of workers, machines, utensils and, environment are the main predisposing factors.65,66 The infected labors in the presence of bad hygienic conditions could transmit MRSA to processed food, including; beef meat products, chicken meat products, milk, and milk products. The access of MRSA to processed human food suggesting the occurrence of food-borne disease, which has public health concern.67,68
S. aureus found mainly in different retail meat products, 2217 specimens of variable types of meat-products from various stores were examined bacteriologically, where the total prevalence of MRSA was 11.9%. The intensity of MRSA in various meat products was listed as: turkey (35.3%); chicken (16.0%); veal (15.2%); pork (10.7%); beef (10.6%); lamb and mutton (6.2%). Most of the isolated MRSA strains are of animal origin (de Boer et al, 2009). Other investigations were performed in Japan and Switzerland, where the prevalence of MRSA in meat products was 65 and 23%, respectively.69,70 In the USA, a study was conducted on 120 retail meat specimens. The prevalence of S. aureus was 39.2%, out of them 5% were identified as MRSA strains (USA100-ST5, and USA300-ST8),71 while in Canada the prevalence of MRSA strains was7.7% in the examined retail meat specimens harbored MRSA; 30% of the isolated strains are of human origin.2
The existence of MRSA in bovine milk was predominant. After the first isolation of MRSA from dairy cows suffering from mastitis,21 several cases of MRSA clinical or subclinical mastitis were recorded in dairy animals. The prevalence of MRSA in dairy cattle in Korea was 0.18%,72 a higher prevalence of MRSA (15%) was noticed in Belgium in dairy cows.73 A previous study in Germany revealed that the prevalence of MRSA was 45% and 4.1% in nasal swabs (obtained from veal calves at slaughter) and bulk-milk tanks, respectively. The colonization of MRSA in the udder of dairy cows is frequent and resulting in subclinical mastitis with severe economic losses.74 The direct contact of the diseased workers with the lactating animals may transmit the infection to the animal and vice versa. A previous study was carried out in Hungary stated that the MRSA strains which were retrieved from mastitic cows and the worker are found to be analogous using both phenotypic and molecular characterization indicating the zoonotic transmission of infection between human and cows.75 Algammal et al76 reported that, MRSA is incriminated in various infections as well food-poisoning illness. The prevalence of MRSA in the examined processed poultry meat was 5.5% (n= 8). The S. aureus count was 102 CFU/g in breast, leg and gizzard specimens; moreover, it was 3.3 × 103 CFU/g in the examined frozen livers. All the recovered MRSA strains are cefoxitin-resistant, mecA gene-positive, and mecC gene-negative. The majority of the retrieved MRSA strains (75%) harbored seb gene. Using, RT-PCR, MRSA strains were found to initiate the production of SEB-toxin production in livers after 24 h of storage (>8 °C). The authors also concluded that MRSA strains isolated from chicken-meat in Egypt usually possess the seb gene. To prohibit the SEB-toxin production, the time of “out of refrigeration” should be limited and shorten.
Diseases Caused by MRSA
MRSA strains could induce multiple human infections such as acne, wound suppuration, food poisoning, urinary tract infection, pyogenic pneumonia, endocarditis, nosocomial-infections, osteomyelitis, health-care-associated infections, otitis, mastitis, and septicemia.77 In horse, bacteria cause a localized purulent infection and botryomycosis “peculiar disease” (ie pyogenic inflammation of spermatic cord); in cattle and ewe, localized pyogenic infection and severe acute mastitis with marked toxemia; in sheep, abscess disease resemble caseous lymphadenitis caused by anaerobic strains; in dogs and cats, pustular dermatitis and food poisoning; in pig, exudative epidermatitis “greasy pig disease”; in birds, suppurative arthritis “Bumble-foot”.78,79
Antimicrobial Resistance of MRSA
The emergence of multidrug-resistance (MDR) in MRSA is an important threat that is resulting in the failure in treatment and control. MRSA developed new effective mechanisms to resist modern antibiotics, so it is considered a life-threatening microorganism for both humans and animals.17,80,81 Generally, the MDR phenomenon is mainly attributed to the frequent and haphazard use of antibiotics as well as the encoding of some antibiotic resistance genes.82–88 MRSA always exhibits resistance to multiple antimicrobial agents, including; penicillin, methicillin, oxacillin, cefoxitin, amoxicillin-clavulanic acid, amoxicillin-sulbactam, quinolones, macrolides, cephalosporins, tetracycline, and chloramphenicol.2,89 MDR-MRSA always not affected by the first line of antibiotic treatment in most cases, many studies illustrated MRSA resistance to the new generations of antibiotics such as vancomycin, linezolid and daptomycin.90,91 The release of β-lactamase enzyme by S. aureus is the main cause of penicillin and penicillin derivatives resistance, while the mecA gene (encodes for Penicillin-binding protein production) is responsible for methicillin resistance. The mecA gene is found on the MRSA chromosome (SCCmec); seven types of SCCmec were identified up to date. There are seven types of SCCmec (I–VII). The production of MRSA penicillin-binding protein is considered the most important cause of penicillin and methicillin resistance.92–94
Linezolid is a chemically synthetic oxazolidinone that acts as an antibacterial agent against S. aureus by inhibition of the bacterial protein synthesis through the binding with the 23S portion of the 50S subunit of ribosomes. The linezolid could exhibit potent antimicrobial activity against MRSA that was approved FDA for the treatment of MRSA-associated nosocomial pneumonia.95,96 Unlike vancomycin, linezolid found to be more effective against MRSA.97 The protracted use of linezolid (over 14 days) resulting in three manifestations; a- Remarkable thrombocytopenia,98 therefore weekly complete blood counts is essential in cases treated with linezolid,99 b- Mitochondrial dysfunction, which is incriminated in lactic acidosis and neuropathy,100 c- Serotonin toxicity, as linezolid is a non-potent monoamine-oxidase (MAO) inhibitor.101
The evaluation of the antimicrobial activity of Sidr honey against MRSA strains recovered from catfish in Egypt was investigated. The clinical findings revealed the ability of Sidr honey to inhibit or inactivate coa and spa virulence genes of the examined MRSA strains. In addition, the effect of Sidr honey on the morphological characteristics of MRSA strains was investigated using the scanning electron microscope. The morphology of Normal bacterial cells (MRSA) is illustrated in (Figure 1 A, and B). The bacterial cells undergo elongation after 24 h of exposure to sidr honey as shown in (Figure 1.C, and D), moreover, the bacterial cells exhibited irregular cell surfaces followed by the appearance of cell debris after the exposure to MRSA strains to Sidr honey for 48 as illustrated in (Figure 1.E and F). Sidr honey exerts an effective inhibition of the growth of isolated strains.62
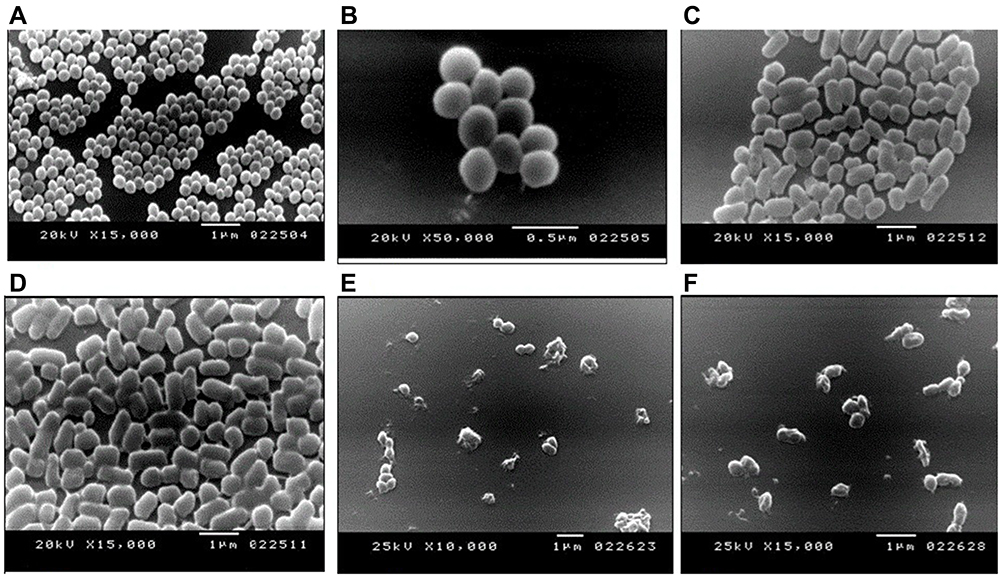 |
Figure 1 Scanning Electron Microscopy revealing the morphological changes of MRSA cells after the exposure to sidr honey. (A and B): Normal bacterial cells (MRSA). (C and D): The bacterial cells undergo elongation after 24 h of exposure to sidr honey. (E and F): The bacterial cells exhibited irregular cell surfaces followed by the appearance of cell debris after the exposure to MRSA strains to Sidr honey for 48 h. Reprinted with permission from Enany ME, Algammal AM, Shagar GI, Hanora AM, Elfeil WK, Elshaffy NM. Molecular typing and evaluation of Sidr honey inhibitory effect on virulence genes of MRSA strains isolated from catfish in Egypt. Pakistan J Pharm Sci. 2018;31.62 |
Treatment and Control of MRSA Infection
The frequent use of antibiotics in humans and animals is resulting in the emergence of MDR-MRSA. One Health approach of MDR-MRSA infections requires the collaboration between public health experts, microbiologists, epidemiologists, veterinary and medical clinicians. Nowadays, MRSA infections are resistant to most commercial antibiotics. To control the antibiotic resistance problem in both animals and humans, the investigation and screening of resistant strains are necessary using the antibiotic sensitivity test (Disc diffusion method). Furthermore, the application of alternative eco-friendly treatment regimens should be implemented including, herbal medications and symbiotic such as Austroeupatorium inulaefolium (H.B.K.) essential oil and leaves-extracted essential oil of Leoheo domatiophorus Chaowasku.102,103 Besides, therapies of natural origin should be implemented especially in dermal infections such as a combination of propolis, tea tree oil, and Aloe vera and the combination of Myrtus communisL., Origanum vulgare and tretinoin (MOTC).104,105 The spontaneous use of broad-spectrum antibiotics in the treatment of MRSA infection must be prohibited, and the ideal treatment should base on the eminent results of the antibiotic sensitivity test.106,107 New antibiotics are thought to be more effective against MRSA infections such as telavancin and tedizolid.108,109 Furthermore, an alternative approach would be to identify molecules that can interfere with the process of efflux.110 Hospitals and veterinary authorities must apply general precautions to reduce the widespread of MRSA in community, general hygiene practice, infection control routine, disinfection of the environment are necessary. The implementation of the HACCP guidelines is necessary during the handling and processing of meat products, milk, and milk products to minify the contamination of human food with MRSA. Infected animals with MRSA must be isolated and treated and infected wounds with MRSA should be disinfected and covered.111–114
Summary
The present study briefly illustrates variable data about the host susceptibility, diseases, epidemiology, virulence factors, antibiotic resistance, treatment and control of MRSA infection. Widespread of MRSA in the community is considered a public health threat. The development of multi-drug resistance is the main obstacle in the treatment of MRSA infections. The cooperation between public health consultants and veterinary authorities is essential to control the spread of MRSA infection. Continuous mentoring of the antibiotic sensitivity is the key for MRSA infection treatment. Public health awareness, general hygiene practice, and veterinary quarantine and biosecurity are necessary for the prevention of MRSA infection. In conclusion, MRSA is considered a substantial emergent zoonotic pathogen affecting both humans and animals; veterinarians are responsible for the protection of public health and food safety.
Acknowledgments
The authors are grateful to the Deanship of Scientific Research at Princess Nourah bint Abdulrahman University through the Fast-track Research Funding Program.
Author Contributions
A.M.A.; conception, and study design. A.M.A, M.A.M., H.F.H.,N.E. A.A.E., G.E.B., W.N.H, and D.H.M.A; execution, acquisition of data, analysis and interpretation. A.M.A, and W.N.H.; drafting the manuscript. A.M.A, M.AM. W.N.H.,N.E., A.A.E., G.E.B., D.H.M.A and H.F.H.: critically reviewing the article. All authors have agreed on the journal to which the article has been submitted. All authors reviewed and agreed on all versions of the article before submission, during revision, the final version accepted for publication, and any significant changes introduced at the proofing stage, and agreed to take responsibility and be accountable for the contents of the article.
Disclosure
All authors declare no conflicts of interest for this work.
References
1. Atoum MF, Akel H, Battikhi MN. Comparison of PCR and disc diffusion methods in detecting methicillin resistance among Staphylococcus species from nosocomial infections. Saudi Med J. 2003;24:1410–1412.
2. Weese JS, van Duijkeren E. Methicillin-resistant Staphylococcus aureus and Staphylococcus pseudintermedius in veterinary gmedicine. Vet Microbiol. 2010;140:418–429. doi:10.1016/j.vetmic.2009.01.039
3. Aklilu E, Zunita Z, Hassan L, Chen H. Phenotypic and genotypic characterization of methicillin-resistant Staphylococcus aureus (MRSA) isolated from dogs and cats at University Veterinary Hospital, Universiti Putra Malaysia. Trop Biomed. 2010;27:483–492.
4. Azeez-Akande O. Global trend of methicillin-resistant Staphylococcus aureus and emerging challenges for control. African J Clin Exp Microbiol. 2010;11.
5. Alaklobi F, Aljobair F, Alrashod A, et al. The prevalence of community-associated methicillin-resistant Staphylococcus aureus among outpatient children in a tertiary hospital: a prospective observational study in Riyadh, Saudi Arabia. Intl J Pediatr Adolesc Med. 2015;2:136–140. doi:10.1016/j.ijpam.2015.09.001
6. Quinn P, Carter M, Markey B, Carter G. Mastitis. In: Clinical Veterinary Microbiology. London: Wolfe Publishing; 1994:327–344.
7. Elsayed MS, Aem E-B, Dawoud MA. Phenotypic and genotypic detection of virulence factors of Staphylococcus aureus isolated from clinical and subclinical mastitis in cattle and water buffaloes from different farms of Sadat City in Egypt. Vet World. 2015;8:1051. doi:10.14202/vetworld.2015.1051-1058
8. Lakhundi S, Zhang K. Methicillin-resistant Staphylococcus aureus: molecular characterization, evolution, and epidemiology. Clin Microbiol Rev. 2018;31(4).
9. Licitra G. Etymologia: Staphylococcus. Emerg Infect Dis. 2013;19:1553. doi:10.3201/eid1909.ET1909
10. Scherrer D, Corti S, Muehlherr J, Zweifel C, Stephan R. Phenotypic and genotypic characteristics of Staphylococcus aureus isolates from raw bulk-tank milk samples of goats and sheep. Vet Microbiol. 2004;101:101–107. doi:10.1016/j.vetmic.2004.03.016
11. Freeman-Cook L, Freeman-Cook KD, Alcamo IE. Staphylococcus Aureus Infections. Infobase Publishing; 2006.
12. Peacock SJ, Paterson GK. Mechanisms of methicillin resistance in Staphylococcus aureus. Annu Rev Biochem. 2015;84:577–601. doi:10.1146/annurev-biochem-060614-034516
13. Gajdács M. The continuing threat of methicillin-resistant Staphylococcus aureus. Antibiotics. 2019;8:52. doi:10.3390/antibiotics8020052
14. Kitti T, Boonyonying K, Sitthisak S. Prevalence of methicillin-resistant Staphylococcus aureus among university students in Thailand. Southeast Asian J Trop Med Public Health. 2011;42:1498.
15. Prenafeta A, Sitjà M, Holmes M, Paterson G. Biofilm production characterization of mecA and mecC methicillin-resistant Staphylococcus aureus isolated from bovine milk in Great Britain. J Dairy Sci. 2014;97:4838–4841. doi:10.3168/jds.2014-7986
16. Murakami K, Minamide W, Wada K, Nakamura E, Teraoka H, Watanabe S. Identification of methicillin-resistant strains of staphylococci by polymerase chain reaction. J Clin Microbiol. 1991;29:2240–2244. doi:10.1128/JCM.29.10.2240-2244.1991
17. Gajdács M. The concept of an ideal antibiotic: implications for drug design. Molecules. 2019;24:892. doi:10.3390/molecules24050892
18. Pantosti A, Venditti M. What is MRSA? Eur Respir J. 2009;34:1190–1196. doi:10.1183/09031936.00007709
19. Gajdács M, Zsoldiné Urbán E. Epidemiology and resistance trends of Staphylococcus aureus isolated from vaginal samples: a 10-year retrospective study in Hungary. Acta Dermatovenerol Alp Pannonica Adriat. 2019;28:143–147. doi:10.15570/actaapa.2019.35
20. Persoons D, Van Hoorebeke S, Hermans K, et al. Methicillin-resistant Staphylococcus aureus in poultry. Emerg Infect Dis. 2009;15:452. doi:10.3201/eid1503.080696
21. Devriese L, Van Damme L, Fameree L. Methicillin (cloxacillin)‐resistant Staphylococcus aureus strains isolated from bovine mastitis cases. Zentralblatt für Veterinärmedizin Reihe B. 1972;19:598–605. doi:10.1111/j.1439-0450.1972.tb00439.x
22. Eriksen NR, Espersen F, Rosdahl VT, Jensen K. Carriage of Staphylococcus aureus among 104 healthy persons during a 19-month period. Epidemiol infect. 1995;115:51–60. doi:10.1017/S0950268800058118
23. VandenBergh MF, Yzerman EP, van Belkum A, Boelens HA, Sijmons M, Verbrugh HA. Follow-up of Staphylococcus aureus nasal carriage after 8 years: redefining the persistent carrier state. J Clin Microbiol. 1999;37:3133–3140. doi:10.1128/JCM.37.10.3133-3140.1999
24. Diep BA, Chambers HF, Graber CJ, et al. Emergence of multidrug-resistant, community-associated, methicillin-resistant Staphylococcus aureus clone USA300 in men who have sex with men. Ann Int Med. 2008;148:249–257. doi:10.7326/0003-4819-148-4-200802190-00204
25. Von Eiff C, Becker K, Machka K, Stammer H, Peters G. Nasal carriage as a source of Staphylococcus aureus bacteremia. N Engl J Med. 2001;344:11–16. doi:10.1056/NEJM200101043440102
26. Wertheim HF, Vos MC, Ott A, et al. Risk and outcome of nosocomial Staphylococcus aureus bacteremia in nasal carriers versus non-carriers. Lancet. 2004;364:703–705. doi:10.1016/S0140-6736(04)16897-9
27. Yang E, Tan J, Eells S, Rieg G, Tagudar G, Miller L. Body site colonization in patients with community-associated methicillin-resistant Staphylococcus aureus and other types of S. aureus skin infections. Clin Microbiol Infect. 2010;16:425–431. doi:10.1111/j.1469-0691.2009.02836.x
28. Faden H, Lesse AJ, Trask J, et al. Importance of colonization site in the current epidemic of staphylococcal skin abscesses. Pediatrics. 2010;125:618–624. doi:10.1542/peds.2009-1523
29. Mertz D, Frei R, Periat N, et al. Exclusive Staphylococcus aureus throat carriage: at-risk populations. Arch Intern Med. 2009;169:172–178. doi:10.1001/archinternmed.2008.536
30. Batra R, Eziefula AC, Wyncoll D, Edgeworth J. Throat and rectal swabs may have an important role in MRSA screening of critically ill patients. Intensive Care Med. 2008;34(9):1703–1706. doi:10.1007/s00134-008-1153-1
31. Gillaspy AF, Lee CY, Sau S, Cheung AL, Smeltzer MS. Factors affecting the collagen binding capacity of Staphylococcus aureus. Infect Immun. 1998;66:3170–3178. doi:10.1128/IAI.66.7.3170-3178.1998
32. Ikawaty R, Brouwer E, Van Duijkeren E, Mevius D, Verhoef J, Fluit A. Virulence factors of genotyped bovine mastitis Staphylococcus aureus isolates in the Netherlands. Int J Dairy Sci. 2010;5:60–70. doi:10.3923/ijds.2010.60.70
33. Khandke L, Nonoyama A, Hodge TS, Nema S. Stable immunogenic compositions of Staphylococcus aureus antigens. Google Patents. 2013.
34. Kong C, Neoh HM, Nathan S. Targeting Staphylococcus aureus toxins: a potential form of anti-virulence therapy. Toxins. 2016;8:72. doi:10.3390/toxins8030072
35. van Kessel KP, Bestebroer J, van Strijp JA. Neutrophil-mediated phagocytosis of Staphylococcus aureus. Front Immunol. 2014;5:467. doi:10.3389/fimmu.2014.00467
36. McDevitt D, Francois P, Vaudaux P, Foster T. Molecular characterization of the clumping factor (fibrinogen receptor) of Staphylococcus aureus. Mol Microbiol. 1994;11:237–248. doi:10.1111/j.1365-2958.1994.tb00304.x
37. Normanno G, Firinu A, Virgilio S, et al. Bolzoni G. Coagulase-positive Staphylococci and Staphylococcus aureus in food products marketed in Italy. Int J Food Microbiol. 2005;98:73–79. doi:10.1016/j.ijfoodmicro.2004.05.008
38. Speziale P, Pietrocola G, Foster TJ, Geoghegan JA. Protein-based biofilm matrices in Staphylococci. Front Cell Infect Microbiol. 2014;4:171. doi:10.3389/fcimb.2014.00171
39. Eid HM, Algammal AM, Elfeil WK, Youssef FM, Harb SM, Abd-Allah EM. Prevalence, molecular typing, and antimicrobial resistance of bacterial pathogens isolated from ducks. Vet World. 2019;12:677. doi:10.14202/vetworld.2019.677-683
40. Otto M. Staphylococcus aureus toxins. Curr Opin Microbiol. 2014;17:32–37. doi:10.1016/j.mib.2013.11.004
41. Tam K, Torres VJ. Staphylococcus aureus secreted toxins and extracellular enzymes. Gram-Positive Pathog. 2019;1:640–668.
42. Otto M. Basis of virulence in community-associated methicillin-resistant Staphylococcus aureus. Ann Rev Microbiol. 2010;64:143–162. doi:10.1146/annurev.micro.112408.134309
43. Akineden Ö, Annemüller C, Hassan A, Lämmler C, Wolter W, Zschöck M. Toxin genes and other characteristics of Staphylococcus aureus isolates from milk of cows with mastitis. Clin Diagn Lab Immunol. 2001;8:959–964. doi:10.1128/CDLI.8.5.959-964.2001
44. Omoe K, Ishikawa M, Shimoda Y, Hu DL, Ueda S, Shinagawa K. Detection of seg, seh, and sei genes in Staphylococcus aureus isolates and determination of the enterotoxin productivities of S. aureus isolates harboring seg, seh, or sei genes. J Clin Microbiol. 2002;40:857–862. doi:10.1128/JCM.40.3.857-862.2002
45. Hata E, Katsuda K, Kobayashi H, Uchida I, Tanaka K, Eguchi M. Genetic variation among Staphylococcus aureus strains from bovine milk and their relevance to methicillin-resistant isolates from humans. J Clin Microbiol. 2010;48:2130–2139. doi:10.1128/JCM.01940-09
46. Chang BS, Bohach GA, Lee SU, et al. Immunosuppression by T regulatory cells in cows infected with Staphylococcal superantigen. J Vet Sci. 2005;6:247. doi:10.4142/jvs.2005.6.3.247
47. Rall V, Vieira F, Rall R, et al. PCR detection of staphylococcal enterotoxin genes in Staphylococcus aureus strains isolated from raw and pasteurized milk. Vet Microbiol. 2008;132:408–413. doi:10.1016/j.vetmic.2008.05.011
48. Burton JL, Erskine RJ. Immunity and mastitis some new ideas for an old disease. Vet Clin. 2003;19:1–45.
49. Barrio MB, Rainard P, Prévost G. LukM/LukF′ PV is the most active Staphylococcus aureus leukotoxin on bovine neutrophils. Microbes Infect. 2006;8:2068–2074. doi:10.1016/j.micinf.2006.03.004
50. Ono HK, Omoe K, Imanishi KI, et al. Identification and characterization of two novel staphylococcal enterotoxins, types S and T. Infect Immun. 2008;76:4999–5005. doi:10.1128/IAI.00045-08
51. Abouelfetouh A. The status of methicillin resistance among Egyptian Staphylococcus aureus isolates: an overview. Infect Disord Drug Targets. 2017;17:67–69.
52. Ahmed EF, Gad GF, Abdalla AM, Hasaneen AM, Abdelwahab SF. Prevalence of methicillin resistant Staphylococcus aureus among Egyptian patients after surgical interventions. Surg Infect. 2014;15:404–411. doi:10.1089/sur.2013.212
53. Abdel-Maksoud M, El-Shokry M, Ismail G, et al. Methicillin-resistant Staphylococcus aureus recovered from healthcare- and community-associated infections in Egypt. Int J Bacteriol. 2016;2016:1–5. doi:10.1155/2016/5751785
54. El-Jakee J, Nagwa AS, Bakry M, Zouelfakar SA, Elgabry E, El-Said WG. Characteristics of Staphylococcus aureus strains isolated from human and animal sources. Am-Eurasian J Agric Environ Sci. 2008;4:221–229.
55. El-Bouseary MM, El-Banna TE, Sonbol FI. Prevalence of MRSA among Staphylococcus aureus isolates recovered from patients with otitis media. Nat Sci. 2018;16(6):48–55.
56. Algammal AM, Enany ME, El-Tarabili RM, Ghobashy MO, Helmy YA. Prevalence, antimicrobial resistance profiles, virulence and enterotoxin-determinant genes of MRSA isolated from subclinical bovine mastitis samples in Egypt. Pathogens. 2020;9:362. doi:10.3390/pathogens9050362
57. Enany M, Younes S, AL gammal AL, Salem M, El Dieb H. Prevalence of coagulase (coa) gene and mecA gene of S. aureus isolated from bovine clinical mastitis. Suez Canal Vet Med J. 2013;18:149–157. doi:10.21608/scvmj.2013.78288
58. Aly S, Fathi M, Youssef E, Mabrok M. Trichodinids and monogeneans infestation among Nile tilapia hatcheries in Egypt: prevalence, therapeutic and prophylactic treatments. Aquac Int. 2020;28:1459–1471. doi:10.1007/s10499-020-00537-w
59. Abouelmaatti RR, Algammal AM, Elfeil WM, et al. Genetic characterization, cloning, and expression of Toll-like receptor 1 mRNA Nile tilapia (Oreochromis niloticus). Veterinarski Arhiv. 2020;90:185–196.
60. El-Sayed M, Algammal A, Abouel-Atta M, Mabrok M, Emam A. Pathogenicity, genetic typing, and antibiotic sensitivity of Vibrio alginolyticus isolated from Oreochromis niloticus and Tilapia zillii. Revue de Médecine Vétérinaire. 2019;170:80–86.
61. Hammad AM, Watanabe W, Fujii T, Shimamoto T. Occurrence and characteristics of methicillin-resistant and-susceptible Staphylococcus aureus and methicillin-resistant coagulase-negative staphylococci from Japanese retail ready-to-eat raw fish. Intl J Food Microbiol. 2012;156:286–289. doi:10.1016/j.ijfoodmicro.2012.03.022
62. Enany ME, Algammal AM, Shagar GI, Hanora AM, Elfeil WK, Elshaffy NM. Molecular typing and evaluation of Sidr honey inhibitory effect on virulence genes of MRSA strains isolated from catfish in Egypt. Pakistan J Pharm Sci. 2018;31.
63. Saklani P, Lekshmi M, Nayak BB, Kumar SH. Survival of methicillin-resistant Staphylococcus aureus (MRSA) in fish and shrimp at different storage conditions. J Food Protect. 2020;83(5):844–848. doi:10.4315/JFP-19-546
64. Gilbert MJ, Bos ME, Duim B, et al. Livestock-associated MRSA ST398 carriage in pig slaughterhouse workers related to quantitative environmental exposure. Occup Environ Med. 2012;69:472–478. doi:10.1136/oemed-2011-100069
65. Soonthornchaikul N, Garelick H, Jones H, Jacobs J, Ball D, Choudhury M. Resistance to three antimicrobial agents of Campylobacter isolated from organically-and intensively-reared chickens purchased from retail outlets. Int J Antimicrob Agents. 2006;27:125–130. doi:10.1016/j.ijantimicag.2005.09.020
66. Kadariya J, Smith TC, Thapaliya D. Staphylococcus aureus and staphylococcal food-borne disease: an ongoing challenge in public health. BioMed Res Int. 2014;2014.
67. Mayrhofer S, Paulsen P, Smulders FJ, Hilbert F. Antimicrobial resistance profile of five major food-borne pathogens isolated from beef, pork and poultry. Int J Food Microbiol. 2004;97:23–29. doi:10.1016/j.ijfoodmicro.2004.04.006
68. Carey AJ, Saiman L, Polin RA. Hospital-acquired infections in the NICU: epidemiology for the new millennium. Clin Perinatol. 2008;35:223–249. doi:10.1016/j.clp.2007.11.014
69. Kitai S, Shimizu A, Kawano J, et al. Prevalence and characterization of Staphylococcus aureus and enterotoxigenic Staphylococcus aureus in retail raw chicken meat throughout Japan. J Vet Med Sci. 2005;67:269–274. doi:10.1292/jvms.67.269
70. Schraft H, Kleinlein N, Untermann F. Contamination of pig hindquarters with Staphylococcus aureus. Int J Food Microbiol 1992;15:191–194. doi:10.1016/0168-1605(92)90148-V
71. Pu S, Han F, Ge B. Isolation and characterization of methicillin-resistant Staphylococcus aureus strains from Louisiana retail meats. Appl Environ Microbiol. 2009;75:265–267. doi:10.1128/AEM.01110-08
72. Kwon NH, Park KT, Moon JS, et al. Staphylococcal cassette chromosome mec (SCC mec) characterization and molecular analysis for methicillin-resistant Staphylococcus aureus and novel SCC mec subtype IVg isolated from bovine milk in Korea. J Antimicrob Chemother. 2005;56:624–632. doi:10.1093/jac/dki306
73. Hendriksen RS, Mevius DJ, Schroeter A, et al. Prevalence of antimicrobial resistance among bacterial pathogens isolated from cattle in different European countries: 2002–2004. Acta Veterinaria Scandinavica. 2008;50:28. doi:10.1186/1751-0147-50-28
74. Tenhagen BA, Vossenkuhl B, Käsbohrer A, et al. Methicillin-resistant Staphylococcus aureus in cattle food chains–prevalence, diversity, and antimicrobial resistance in Germany. J Anim Sci 2014;92:2741–2751. doi:10.2527/jas.2014-7665
75. Juhász-Kaszanyitzky É, Jánosi S, Somogyi P, et al. MRSA transmission between cows and humans. Emerg Infect Dis. 2007;13:630. doi:10.3201/eid1304.060833
76. Algammal AM, Abolghait SK, Fathi AG, Youssef FM. Methicillin-resistant Staphylococcus aureus (MRSA) isolated from chicken meat and giblets often produces staphylococcal enterotoxin B (SEB) in non-refrigerated raw chicken livers. Int J Food Microbiol. 2020;328:108669.
77. Boucher HW, Corey GR. Epidemiology of methicillin-resistant Staphylococcus aureus. Clin Infect Dis. 2008;46:S344–S349. doi:10.1086/533590
78. Bergmann V, Köhler B, Vogel K. Staphylococcus aureus infection in chickens in industrialized poultry units. 1. Manifestations of Staphylococcus aureus infection in chickens. Archiv Fur Experimentelle Veterinarmedizin. 1980;34:891–903.
79. Vanderhaeghen W, Hermans K, Haesebrouck F, Butaye P. Methicillin-resistant Staphylococcus aureus (MRSA) in food production animals. Epidemiol Infect. 2010;138:606–625. doi:10.1017/S0950268809991567
80. Mubarack HM, Doss A, Vijayasanthi M, Venkataswamy R. Antimicrobial drug susceptibility of Staphylococcus aureus from subclinical bovine mastitis in Coimbatore, Tamilnadu, South India. Vet World. 2012;5:352. doi:10.5455/vetworld.2012.352-355
81. Tang SS, Apisarnthanarak A, Hsu LY. Mechanisms of β-lactam antimicrobial resistance and epidemiology of major community-and healthcare-associated multidrug-resistant bacteria. Adv Drug Deliv Rev. 2014;78:3–13. doi:10.1016/j.addr.2014.08.003
82. Whist AC, Østerås O, Sølverød L. Staphylococcus aureus and Streptococcus dysgalactiae in Norwegian herds after introduction of selective dry cow therapy and teat dipping. J Dairy Res. 2007;74:1–8. doi:10.1017/S0022029906002135
83. Eid HI, Algammal AM, Nasef SA, Elfeil WK, Mansour GH. Genetic variation among avian pathogenic E. coli strains isolated from broiler chickens. Asian J Anim Vet Adv. 2016;11:350–356. doi:10.3923/ajava.2016.350.356
84. Enany ME, Algammal AM, Nasef SA, et al. The occurrence of the multidrug resistance (MDR) and the prevalence of virulence genes and QACs resistance genes in E. coli isolated from environmental and avian sources. AMB Express. 2019;9:192. doi:10.1186/s13568-019-0920-4
85. Algammal AM, Wahdan A, Elhaig MM. Potential efficiency of conventional and advanced approaches used to detect Mycobacterium bovis in cattle. Microb Pathog. 2019;134:103574. doi:10.1016/j.micpath.2019.103574
86. Algammal AM, Mohamed MF, Tawfiek BA, Hozzein WN, El Kazzaz WM, Mabrok M. Molecular typing, antibiogram and PCR-RFLP based detection of aeromonas hydrophila complex isolated from oreochromis niloticus. Pathogens. 2020;9:238. doi:10.3390/pathogens9030238
87. Algammal AM, El-Kholy AW, Riad EM, et al. Genes encoding the virulence and the antimicrobial resistance in enterotoxigenic and shiga-toxigenic E. coli isolated from diarrheic calves. Toxins. 2020;12:383. doi:10.3390/toxins12060383
88. Algammal AM, El-Sayed ME, Youssef FM, et al. Prevalence, the antibiogram and the frequency of virulence genes of the most predominant bacterial pathogens incriminated in calf pneumonia. AMB Express. 2020;10:1–8. doi:10.1186/s13568-020-01037-z
89. Chopra I, Roberts M. Tetracycline antibiotics: mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol Rev. 2001;65:232–260.
90. Bonnstetter KK, Wolter DJ, Tenover FC, McDougal LK, Goering RV. Rapid multiplex PCR assay for identification of USA300 community-associated methicillin-resistant Staphylococcus aureus isolates. J Clin Microbiol. 2007;45:141–146. doi:10.1128/JCM.01228-06
91. Traczewski MM, Katz BD, Steenbergen JN, Brown SD. Inhibitory and bactericidal activities of daptomycin, vancomycin, and teicoplanin against methicillin-resistant Staphylococcus aureus isolates collected from 1985 to 2007. Antimicrob Agents Chemother 2009;53:1735–1738. doi:10.1128/AAC.01022-08
92. Ubukata K, Nonoguchi R, Matsuhashi M, Konno M. Expression and inducibility in Staphylococcus aureus of the mecA gene, which encodes a methicillin-resistant S. aureus-specific penicillin-binding protein. J Bacteriol. 1989;171:2882–2885. doi:10.1128/JB.171.5.2882-2885.1989
93. Galdiero E, Liguori G, D’Isanto M, Damiano N, Sommese L. Distribution of mecA among methicillin-resistant clinical staphylococcal strains isolated at hospitals in Naples, Italy. Eur J Epidemiol. 2003;18:139–145. doi:10.1023/A:1023067930211
94. Pournaras S, J Sabat A, Grundmann H, Hendrix R, Tsakris A, W Friedrich A. Driving forces of mechanisms regulating oxacillin-resistance phenotypes of MRSA: truly oxacillin-susceptible mecA-positive Staphylococcus aureus clinical isolates also exist. Curr Pharm Des. 2015;21:2048–2053. doi:10.2174/1381612821666150310103754
95. Rybak MJ, Cappelletty DM, Moldovan T, Aeschlimann JR, Kaatz GW. Comparative in vitro activities and postantibiotic effects of the oxazolidinone compounds eperezolid (PNU-100592) and linezolid (PNU-100766) versus vancomycin against Staphylococcus aureus, coagulase-negative staphylococci, Enterococcus faecalis, and Enterococcus faecium. Antimicrob Agents Chemother. 1998;42:721–724. doi:10.1128/AAC.42.3.721
96. Weigelt J, Itani K, Stevens D, Lau W, Dryden M, Knirsch C. Linezolid versus vancomycin in treatment of complicated skin and soft tissue infections. Antimicrob Agents Chemother. 2005;49:2260–2266. doi:10.1128/AAC.49.6.2260-2266.2005
97. Wunderink RG, Rello J, Cammarata SK, Croos-Dabrera RV, Kollef MH. Linezolid vs vancomycin: analysis of two double-blind studies of patients with methicillin-resistant Staphylococcus aureus nosocomial pneumonia. Chest. 2003;124:1789–1797. doi:10.1016/S0012-3692(15)33412-7
98. Walkey AJ, O’Donnell MR, Wiener RS. Linezolid vs glycopeptide antibiotics for the treatment of suspected methicillin-resistant Staphylococcus aureus nosocomial pneumonia: a meta-analysis of randomized controlled trials. Chest. 2011;139:1148–1155. doi:10.1378/chest.10-1556
99. Gerson SL, Kaplan SL, Bruss JB, et al. Hematologic effects of linezolid: summary of clinical experience. Antimicrob Agents Chemother. 2002;46:2723–2726. doi:10.1128/AAC.46.8.2723-2726.2002
100. Legout L, Senneville E, Gomel J, Yazdanpanah Y, Mouton Y. Linezolid-induced neuropathy. Clin Infect Dis. 2004;38:767–768. doi:10.1086/381762
101. Lawrence KR, Adra M, Gillman PK. Serotonin toxicity associated with the use of linezolid: a review of postmarketing data. Clin Infect Dis 2006;42:1578–1583. doi:10.1086/503839
102. Bua A, Usai D, Donadu MG, et al. Antimicrobial activity of Austroeupatorium inulaefolium (HBK) against intracellular and extracellular organisms. Nat Prod Res. 2018;2(23):2869–2871. doi:10.1080/14786419.2017.1385014
103. Trong Le N, Viet Ho D, Quoc Doan T, et al. In vitro antimicrobial activity of essential oil extracted from leaves of Leoheo domatiophorus Chaowasku, DT Ngo and HT Le in Vietnam. Plants. 2020;9(4):453. doi:10.3390/plants9040453
104. Mazzarello V, Donadu MG, Ferrari M, et al. Treatment of acne with a combination of propolis, tea tree oil, and Aloe vera compared to erythromycin cream: two double-blind investigations. Clin Pharm. 2018;10:175.
105. Mazzarello V, Gavini E, Rassu G, et al. Clinical assessment of new topical cream containing two essential oils combined with tretinoin in the treatment of acne. Clin Cosmetic Investig Dermatol. 2020;13:233. doi:10.2147/CCID.S236956
106. Pitkälä A, Haveri M, Pyörälä S, Myllys V, Honkanen-Buzalski T. Bovine mastitis in Finland 2001—prevalence, distribution of bacteria, and antimicrobial resistance. J Dairy Sci. 2004;87:2433–2441. doi:10.3168/jds.S0022-0302(04)73366-4
107. Leonard F, Markey B. Meticillin-resistant Staphylococcus aureus in animals: a review. Vet J. 2008;175:27–36. doi:10.1016/j.tvjl.2006.11.008
108. Gajdács M, Albericio F. Antibiotic resistance: from the bench to patients. Antibiotics. 2019;8:129. doi:10.3390/antibiotics8030129
109. Ito T, Okuma K, Ma XX, Yuzawa H, Hiramatsu K. Insights on antibiotic resistance of Staphylococcus aureus from its whole genome: genomic island SCC. Drug Resist Updates. 2003;6:41–52. doi:10.1016/S1368-7646(03)00003-7
110. Usai D, Donadu M, Bua A, et al. Enhancement of antimicrobial activity of pump inhibitors associating drugs. J Infect Dev Countr. 2019;13(2):162–164. doi:10.3855/jidc.11102
111. Burke SL, Rose WE. New pharmacological treatments for methicillin-resistant Staphylococcus aureus infections. Expert Opin Pharmacother. 2014;15:483–491. doi:10.1517/14656566.2014.876991
112. Roberts JC. Community-associated methicillin-resistant Staphylococcus aureus epidemic clone USA100; more than a nosocomial pathogen. Springerplus. 2013;2:133. doi:10.1186/2193-1801-2-133
113. Sergelidis D, Angelidis A. Methicillin‐resistant Staphylococcus aureus: a controversial food‐borne pathogen. Lett Appl Microbiol. 2017;64(6):409–418. doi:10.1111/lam.12735
114. Abd El-Baky RM, Sandle T, John J, Abuo-Rahma GE, Hetta HF. A novel mechanism of action of ketoconazole: inhibition of the NorA efflux pump system and biofilm formation in multidrug-resistant Staphylococcus aureus. Infect Drug Res. 2019;12:1703. doi:10.2147/IDR.S201124
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.