")
Back to Journals » Infection and Drug Resistance » Volume 11
Methicillin-resistant Staphylococcus aureus: livestock-associated, antimicrobial, and heavy metal resistance
Authors Dweba CC , Zishiri OT , El Zowalaty M, El Zowalaty ME
Received 31 May 2018
Accepted for publication 1 August 2018
Published 28 November 2018 Volume 2018:11 Pages 2497—2509
DOI https://doi.org/10.2147/IDR.S175967
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Joachim Wink
Video abstract presented by Cwengile C Dweba, Oliver T Zishiri, and Mohamed E El Zowalaty
Views: 1804
Cwengile C Dweba,1 Oliver T Zishiri,1 Mohamed E El Zowalaty2
1Discipline of Genetics, School of Life Sciences, College of Agriculture Engineering and Sciences, University of KwaZulu-Natal, Durban, South Africa; 2Microbiology and Virology Research Group, School of Health Sciences, College of Health Sciences, University of KwaZulu-Natal, Durban, South Africa
Abstract: Staphylococcus aureus is an opportunistic pathogen of human and other mammals that is of increasing clinical and veterinary importance due to its ability to rapidly develop antimicrobial resistance. The injudicious use of antibiotics has given rise to the emergence of antibiotic resistant S. aureus strains, most importantly methicillin-resistant Staphylococcus aureus (MRSA). The emergence of livestock-associated MRSA (LA-MRSA) has highlighted the importance of directed research toward its prevention, as well as the need for the discovery and development of more efficient treatment than is currently available. Furthermore, the treatment of MRSA is complicated by the co-selection of heavy metal and antibiotic resistance genes by microorganisms. Livestock and livestock production systems are large reservoirs of heavy metals due to their use in feed as well as environmental contaminant, which has allowed for the selection of LA-MRSA isolates with heavy metal resistance. The World Health Organization reported that Africa has the largest gaps in data on the prevalence of antimicrobial resistance, with no reports on rates for LA-MRSA harboring heavy metal resistance in South Africa. This review aimed to report the emergence of LA-MRSA in South Africa, specifically the most frequent sequence type ST398, globally. Furthermore, we aimed to highlight the importance of LA-MRSA in clinical and food security, as well as this research gap in South Africa. This review sheds light on the prevalence of heavy metals in livestock farms and abattoirs, and focuses on the phenomenon of the co-selection of heavy metal and antibiotic resistance genes in MRSA, emphasizing the importance of a focused direction for research in humans, animals as well as environment using one-health approach.
Keywords: Staphylococcus aureus, methicillin resistance, livestock, MRSA, humans, antimicrobial resistance, antibiotics, resistance, virulence, heavy metals, LA-MRSA, one health
Staphylococcus aureus
Staphylococcus is a genus of the Gram-positive cocci family Staphylococcaceae and includes both coagulase-positive and -negative staphylococci which consists of over 30 species that cause various clinical manifestations.1 Staphylococcus aureus (S. aureus) is an opportunistic pathogen that forms part of the normal commensal flora of humans and livestock, colonizing ~30%–50% of the human population, and is considered to be the most clinically important species.2,3 S. aureus is associated with high infection and mortality rates and is one of the leading causes of minor and life-threatening diseases, most commonly including infections of the skin and respiratory tract, infective endocarditis, toxic shock syndrome, and osteomyelitis.4,5 S. aureus is considered to be one of the most clinically important multidrug-resistant threats globally, according to the recent global priority pathogens list (global PPL) of antibiotic-resistant bacteria by the WHO.6 Methicillin-resistant S. aureus (MRSA), vancomycin intermediate and resistant S. aureus strains are placed second on the list of bacteria of high priority for research and development of new antibiotics.7 The pathogenesis of S. aureus is attributed to the production of an arsenal of several toxins and virulence factors.8 S. aureus isolates are confirmed by the molecular detection of the nuc gene that encodes for a thermonuclease protein.9
Clinical emergence
Infections caused by S. aureus range from minor superficial skin to lethal deep-seated infections such as osteomyelitis and pneumonia which may spread to the blood stream, thereby causing septicaemia.10 S. aureus is a problematic bacterium with the capability to evade the immune system, secrete proteins that neutralize antimicrobial peptides and express several superantigens that disrupt humoral immune responses.10 In addition, MRSA causes significant morbidities and mortalities, with numerous staphylococcal disease outbreaks having been reported over several decades, raising major public health alarms worldwide.
S. aureus is isolated from different human anatomical sites, livestock and companion animals, foods, food production systems and the environment. Consumption of S. aureus-contaminated food caused outbreaks dating back to 1884 where food-borne diseases were found to be caused by the consumption of cheese contaminated with staphylococci.11 A decade later, a family was discovered to have illnesses caused by the consumption of meat from a cow that had died of fever caused by pyogenic staphylococci.11 Other food poisoning outbreaks were reported due to enterotoxin-producing S. aureus strains.12 These outbreaks highlight the health threats of consuming contaminated foods, highlighting the need to scrutinize food products and production systems for bacterial contamination, specifically S. aureus.
Apart from food-borne illnesses, major concerns have been raised in hospitals due to infections caused by S. aureus, and has been identified as the most prominent cause of infective endocarditis in the industrialized world.13 It was reported that a patient with atopic dermatitis developed endocarditis caused by S. aureus,14 this case highlighting that such colonization of these lesions represents an important source of invasive bacterial infections. This is supported by the fact that in native valve endocarditis, S. aureus accounts for 30%–35% of cases, whereas in a patient with atopic dermatitis, it is the exclusive cause of endocarditis .15
While humans are the main reservoir host of S. aureus, livestock have also been colonized with the bacterium, leading to the contamination of food products. This was shown when an outbreak of mastitis was observed in a closely packed dairy herd, and found to be caused by one strain of S. aureus.16 A survey was conducted on various livestock animals including cows, goats, sheep, rabbits, chickens, and cats, and identified S. aureus isolates that were harbouring various staphylococcal enterotoxin genes.17 The occurrence of outbreaks and the identification of novel genes predominating in S. aureus from animal hosts suggest that livestock are an important reservoir of staphylococcal infections. Strict hygiene at the time of milking, segregating any livestock with S. aureus infections, and intensive culling of those infected might be required to reduce the prevalence and incidence of highly transmissible strains of the bacterium.16 However, intensive culling of livestock could prove to be an economic loss to farmers and food production systems, making it important to explore various avenues to reduce the spread of the infection. Regardless of the host, S. aureus isolates produce a repertoire of toxins that allow the bacterium to be a highly threatening pathogen.
Toxins produced by S. aureus
Toxins are described as poisonous substances secreted by the microorganism that directly interacts with the host.4 Panton-Valentine Leukocidin (PVL) is one of the most important, potent, and prevalent toxins produced by S. aureus and is associated with a large percentage of S. aureus isolates that cause necrotizing skin lesions and severe necrotizing pneumonia as well as the destruction of white blood cells.18–20 Several studies were conducted to understand the determinant of PVL-positive S. aureus isolates. It was reported that PVL-producing S. aureus strains caused rapidly progressive, hemorrhagic, necrotizing pneumonia in healthy children and young adults with a high lethality rate.21 More recently, it was demonstrated that PVL-producing S. aureus isolates damaged neutrophils in humans, this being linked to the development of thrombosis in association with osteomyelitis.22 The virulence of PVL toxin lies in the formation of pores in the membrane of the host defense cells by the synergistic action of two proteins, that is, LukF-PV and LukS-PV which are encoded by two co-transcribed genes of a prophage integrated in the S. aureus chromosome.23 The PVL genes can be used as a marker for the presence and intensity of clinically infectious S. aureus isolates.
Alpha toxin (α-toxin) is another toxin that is produced by S. aureus and plays a major role as a virulence factor in its pathogenesis, causing tissue invasion and necrosis while altering bacterial killing in macrophages.24,25 This toxin is encoded by the hla gene and is a pore-forming cytotoxin that is regulated by the global regulators agr, sarA, and sae.26 In a study of S. aureus food poisoning outbreaks in the UK between 1969 and 1970, 79% of the strains were found to produce α-toxin alone or together with another toxin.27 More recently, α-toxin was found to be an important virulence factor in opportunistic bacterial lung infections, where it radically increased bacterial growth and prevented acidification of bacteria-containing macrophages (phagosomes), thereby reducing the effective destruction of S. aureus.25
Apart from the major toxins, the pathogenesis of S. aureus also includes other toxins. The pyrogenic-toxin superantigens bind to major histocompatibility complex II protein which causes extensive T-cell proliferation and cytokine release, resulting in food poisoning and toxic shock.4 Furthermore, the pathogen also produces various enzymes including proteases, lipases, and hyaluronidases which destroy host tissue and may facilitate the spread of infection to adjoining tissues, with the enzyme β-lactamase inactivating penicillin (PBP2a) giving rise to antibiotic-resistant S. aureus.4,28 Staphylococci produce transpeptidase penicillin-binding proteins (PBPs) that reduces the activity of the β-lactam antibiotics;29,30 with second generation of semi-synthetic penicillins, namely methicillin and oxacillin, being developed in 1951 to overcome the β-lactamase activity.31 In 1981, an initial discovery of a reduced-affinity penicillin-binding protein 2a (PBP2a), encoded by the mecA gene was made.32 The low binding affinity of this PBP2a to β-lactam antibiotics enabled the continued synthesis of the peptidoglycan cell wall in MRSA despite the presence of lethal concentrations of methicillin.33 Interestingly, the pathogenesis of livestock-associated S. aureus (LA-SA) that is similar to human-associated S. aureus (HA-SA) is dependent on numerous virulence factors including toxins, cell-surface-associated adhesions, and secreted exo-proteins.34
Antimicrobial resistance
Antibiotics are types of antimicrobial agents that inhibit the growth or destroy bacteria. Previously, antibiotics were considered to be lethal compounds that were produced by living cells to kill or inhibit the growth of other competing microorganisms. However, antibiotics have more recently been described as antimicrobial agents that are also produced partly or wholly through synthetic means.35 Various antibiotics are currently licensed and available for use in animals and humans, and are commonly grouped into five classes based on their mode of action. The Five groups of antibiotics consist of the following: 1) cell wall synthesis inhibitors with a bactericidal effect such as beta-lactams (penicillins, cephalosproins, carbapenems, and monobactams) and vancomycin; 2) protein synthesis inhibitors where the antibiotics bind to bacterial ribosomal subunits either the 30S (such as tetracyclines and aminoglycosides) or the 50S (macrolides, clindamycin, linezolid, chloramphenicol, and streptogramins); 3) nucleic acid synthesis inhibitors that bind to DNA-gyrase or topoisomerase II and topoisomerase IV and thus interrupt the DNA breakage-reunion step (quinolones) or inhibit RNA polymerase activity (rifampicin); 4) folate pathway inhibitors that interfere with folic synthesis; and 5) inhibitors of cell membrane function (polyenes and polypeptides) that disrupts cell membrane permeability.36
The antibiotics most commonly prescribed to treat S. aureus infections include certain cephalosporins, nafcillin, and vancomycin among others. First-generation cephalosporins (such as cefazolin, cephalothin, and cephalexin), clindamycin, lincomycin, and erythromycin have been found to play important therapeutic roles in S. aureus infections such as infections of the skin and soft tissue.37
One of the challenging problems facing the healthcare system is the overprescription and imprudent use of antibiotics. They are injudiciously prescribed for self-limited viral infections with physicians trying to please patients who “feel” that they need antibiotics.38 It was reported that there is a lack of knowledge regarding the prudent and appropriate use of antibiotics and incorrect prescription practices.39–41 In South Africa, inappropriate use of antibiotics has been a common phenomenon of increasing concern with serious implications on the wide spread increase of antimicrobial resistance.39,40 The call for rational use of antimicrobial agents in humans and animals in South Africa is an urgent requirement, and the prudent use of last-resort agents should be a high priority in the country’s national antimicrobial stewardship programs. It was recently observed that antibiotics belonging to tetracyclines and quinolones (enrofloxacin) are misused on a non-prescription basis for backyard poultry and livestock animals in rural areas of Pietermaritzburg in KwaZulu-Natal Province, South Africa (personal communication, Animal Health, Department of Agriculture and Rural Development). This may explain why antimicrobial consumption including prescriptional and non-prescriptional uses is one of the main drivers of resistance in South Africa. It was previously reported that there is a direct correlation between veterinary antimicrobial agents use and the prevalence of resistance in commensal Escherichia coli isolated from various livestock including pigs, poultry, and cattle in Europe.42 Substantial educational efforts are required nationally and globally in order to alleviate the overuse and inappropriate prescription of antibiotics. Greater emphasis needs to be placed in the fact that infections such as “common flu” are caused by a virus and therefore cannot be treated with antibiotics. This alarming overuse of antibiotics has given rise to considerable antibiotic resistance by most bacteria, causing what used to be common infections to become difficult-to-treat, life-threatening infections with numerous complications.
Antibiotic resistance is however a natural phenomenon, which is best explained by the Darwinian rules of natural selection for all living beings. When antibiotics are introduced in a medium, they exert pressure on bacteria, and as only naturally resistant bacteria can survive the resistant bacteria multiply, and become predominant in the medium. The selection of antibiotic-resistant bacteria during antibiotic treatments is therefore involuntary and unavoidable. The phenomenon of persistence of antibiotic resistance in bacterial populations was detailed elsewhere.43 The more antibiotics are used, the more resistance is bound to occur, with three biological processes having been described to enhance the accumulation of bacterial drug resistance. These are new selection, gene epidemics, and strain epidemics with new resistance emerging by 1) advantaging entire species, 2) mutation, and 3) escaping resistance genes to mobile DNA.44 The steady rising of antibiotic resistance by various species showed that there is no single antimicrobial agent available for both human and animal use that has not demonstrated resistance by one or more microorganisms.45,46 In addition, the heritage of past decades of antibiotic use and misuse has resulted in clinically important bacteria being characterized by multiple antibiotic resistance.47
Globally, alarms of rising antimicrobial resistance have been ringing, with resistance to broad-spectrum antibiotics and multiple classes of antibiotics being reported in several European countries where it is increasingly reported as a serious public health threat.48
The world has seen a considerable rise (35%) in consumption of antibiotics between 2000 and 2010, with Brazil, Russia, India, China, and South Africa accounting for 76% of the increase.49 However, more worrisome is the 45% and 13% increased consumption of carbapenems and polymyxins, respectively, which are classified as last-resort antibiotic drugs, raising serious concerns for public health. The prevalence of antibiotic resistance in USA hospitals was also observed and clinically significant bacterial isolates resistant to essential antibiotics were evaluated between 2007 and 2010.50 Various resistant percentages were observed from different antibiotics such that 87.1% of Enterococcus faecium to vancomycin, 56.8% of S. aureus to oxacillin-methicillin, 39.7% of S. aureus to clindamycin, 32.6% Pseudomonas aeruginosa to fluoroquinolones, 31.3% E. coli to fluoroquinolones while 3.9% of E. faecium to daptomycin.50 While these resistance profiles highlight the urgency of establishing treatment protocols that bypass the development of antibiotic among bacterial isolates, more alarming is the prevalence of antibiotic-resistant isolates. Bacteria were found in both source and tap water collected from Michigan, Ohio, USA, with antibiotic-resistant bacterial isolates being found higher in tap water than in water that has just been introduced into the distribution system, which indicated that there was regrowth of the bacteria in the drinking water distribution systems, due to various water treatment strategies.51
Although antibiotic resistance is a global threat, developing countries would appear to shoulder the majority of the burden. These countries face numerous challenges such as limited studies and resources, a lack of surveillance systems, and poor education on proper use of antibiotics, thus antibiotic resistance is an emerging and serious public health threat. Moreover, the threat is aggravated by the limited availability and affordability of newly developed antibiotics. A study conducted in Rwanda revealed that antimicrobial resistance rates are high and pose a serious challenge to the clinical management of common infections.52 The conclusion was supported by the finding of resistance patterns in bacterial pathogens isolated from urine, blood, sputum, and wound swab specimens obtained at a hospital in Rwanda. Eighty percent of S. aureus isolates were found to be resistant to oxacillin (classified as MRSA); 31.4% and 58.7% of E. coli and Klebsiella, respectively, were found to be resistant to third-generation cephalosporins.52 In West Africa, it was reported that urinary tract infections caused by bacterial pathogens such as E. coli and Klebsiella spp. showed moderate to high antibiotic resistance to commonly used antibiotics such as cephalosporins.53 Common bloodstream Gram-negative pathogens were found to have moderate resistance rates to first-line antibiotics such as ampicillin, cotrimoxazole, gentamicin, and amoxicillin/clavulanate. In South Africa, the first example of multidrug-resistant Streptococcus pneumoniae was reported in pediatric patients treated prophylactically with antibiotics for serious viral infections.54 While various reports have shown a similar trend of consistently escalating antibiotic resistance rates in Africa, more alarming is the huge lack of data. The World Health Organisation reported that the African region has one of the largest data gaps on the prevalence of antimicrobial resistance.55 The lack of data, limited and lack of standardized treatment protocols, as well as the lack of adherence to prudent antibiotic use have led to the escalating increase in antimicrobial resistance in the region and globally. More in-depth investigation and research is required in order to consolidate the current treatment measures for a better global health outcome.56
Antimicrobial agents use in livestock
Apart from the inappropriate use of antibiotics by patients and physicians, another major cause of antibiotic resistance is the use of antimicrobial agents in livestock. These antimicrobial agents especially antibiotics are extensively used in livestock production systems to protect their health and enhance their growth performance. Several bacterial pathogens are responsible for devastating losses in livestock, which may account for the increased use and application of antibacterial agents in veterinary practice such as tetracyclines, penicillins, macrolides, fluoroquinolones, and sulfonamides.57 As the world population continues to grow, more food resources are required to sustain their nutritional demands and requirements. The inclusion of antimicrobial agents in animal feed has been found to contribute to lower costs of meat, milk, eggs, and other animal products for human consumption.58 However, the problems arise in that up to 78% of the antibiotics used in the USA are for food production livestock.59 The improper use of antibiotics as prophylactics and for nontherapeutic purposes in poultry, bovine, swine, and other livestock may be attributed to the nature of their production. For example, pigs and poultry production become very dangerous when only a few show symptoms but the entire livestock is treated to prevent contamination of other animals.60 It was reported that considerable quantities of antimicrobial agents were used for nontherapeutic purposes as growth promotors in factory farming globally,61–63 which may have escalated the resistance rates in humans. The prolonged exposure of healthy animals to antibiotics is a recipe for breeding antibiotic resistance among various bacterial populations. Antimicrobial resistance may not be completely avoidable and preventable. In the USA, a wide variety of these antibiotics used in animal treatment and feed are the same as those used in human medicine as shown in Table 1. However, the use of antibiotics as growth promoters has been phased out from 2014 to 2016.64,65
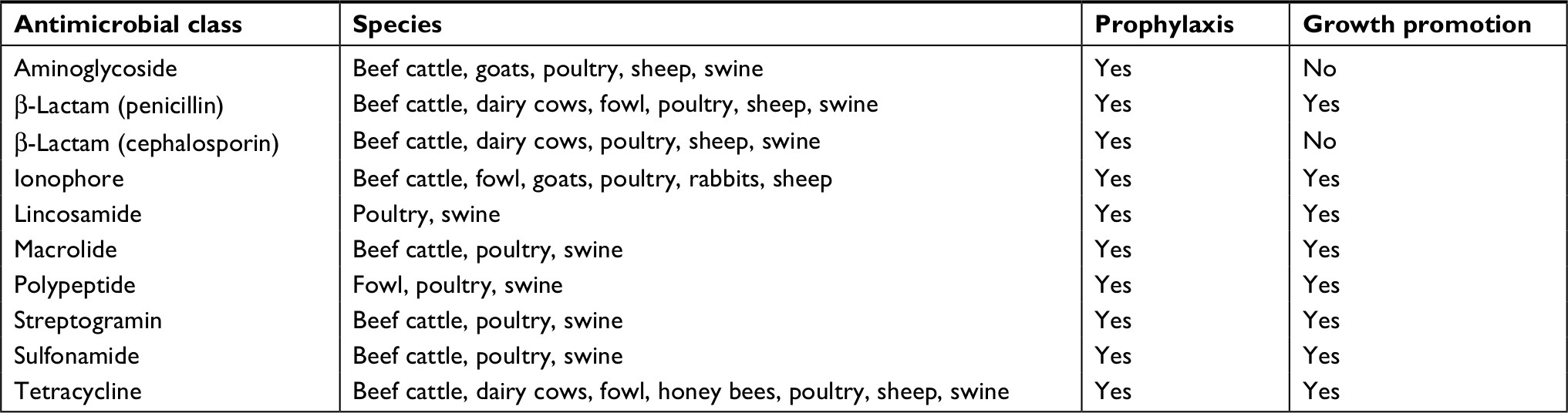 | Table 1 Major antimicrobial agent classes approved for nontherapeutic use in animals Notes: This table was reproduced from United States General Accounting Office. Food Safety. The agricultural use of antibiotics and its implications for human health; 1999. Available at: https://www.gao.gov/archive/1999/rc99074.pdf. Accessed July 12, 2018.142 |
Methicillin-resistant S. aureus
S. aureus is one of the most rapidly evolving bacteria, being able to develop resistance toward a wide variety of antibiotics. The first report of an important antibiotic was the development of penicillin in the early 1940s, with S. aureus becoming a clinical problem less than a decade later, as it had acquired resistance to penicillin by producing a β-lactamase enzyme that rendered penicillin inactive.66 A semi-synthetic penicillin called celbenin (currently referred to as methicillin) was developed in 1959,67,68 and by late 1960, there were already reports of S. aureus isolates resistant to it when the first MRSA strain was identified in the UK in 1961.69,70 S. aureus isolates have developed resistance against ceftaroline, which was a new cephalosporin approved in 2012 to treat MRSA infections.71,72 It is concerning that the rapid development of resistance in S. aureus is not fully understood. MRSA is one of the most important and problematic antibiotic-resistant pathogens globally. Since its initial identification, MRSA has become established as the most prevalent pathogen in hospitals worldwide, responsible for numerous hospital outbreaks globally.69,73–75 Various outbreaks of MRSA led to its classification into two types; healthcare-associated MRSA (HA-MRSA) and community-associated MRSA (CA-MRSA). HA-MRSA is defined as MRSA strains recovered from patients in healthcare facilities due to hospitalization, surgery, or hemodialysis and is typically resistant to clindamycin and other β-lactam antimicrobial agents.76 CA-MRSA is described as strains that have the capacity to infect otherwise healthy people outside of the hospital setting, combining methicillin resistance with enhanced virulence and fitness, with all these strains typically carrying a novel type of methicillin resistance locus that appears to cause less of a fitness burden.77 Apart from HA-MRSA and CA-MRSA, livestock-associated MRSA (LA-MRSA) has been identified and been isolated from numerous livestock. LA-MRSA was first isolated in 2005 from pigs and pig farmers in the Netherlands,78 and later from other animals including bovine, avian, equine, and from humans who are in close contact with animals.79–81 The frequency of LA-MRSA has been steadily increasing following the detection of LA-MRSA in healthy poultry.82 Numerous reports have focused on the identification and characterization of LA-MRSA from different livestock worldwide83,84 as well as in humans.80
With the current global food crisis, the health of food-producing livestock is essential, with LA-MRSA being a major hindrance. Since the emergence of MRSA in animals in 2005, LA-MRSA was reported in pigs and veal calves in particular.2 The first detection of LA-MRSA strain ST398 from poultry was reported, and a comparison of old and recent isolates (1970 and 2006, respectively) showed that the latter have a much more significant resistance rate to eight antibiotics, indicating that the overuse of antibiotics is essential for developing antibiotic resistant strains.82 MRSA was discovered in broiler flocks in the Netherlands,83 with more recent, S. aureus isolates with methicillin-resistant genes being isolated from commercial broiler chickens in South Africa.85 Apart from poultry, MRSA was isolated from a variety of meat products such as veal, beef, lamb, mutton, and game.86 Additional studies have shown that LA-MRSA was present in almost all types of food-producing livestock worldwide, including retail meat in Canada.87 In Korea, MRSA was isolated from imported and domestic meat,88 and from pigs and pig handlers, pig handler’s shower facilities, pork products and production processes.89 The global prevalence of MRSA in livestock has therefore necessitated extensive research towards characterizing LA-MRSA isolates. Previously, S. aureus phenotyping methods were used to identify strains and these included phage typing, biotyping, and typing of enzymes or toxins.90 However, genetic characterization of isolates has significantly gained momentum and has become the pinnacle of isolate identification and characterization.
Genetic characterization of MRSA
Genetic characterization refers to the process by which bacterial isolates are identified or differentiated, and refers to any differences in the appearance or make-up of an accession.91 Therefore, genetic characterization of MRSA isolates is essential to identify old and new isolate types as well as their virulence factors, and facilitates the fight toward improved and effective treatment protocols. S. aureus isolates have been separated into various sequence types using genotyping methods, these being plasmid profile, analysis of chromosomal DNA, southern hybridization, pulse-field gel electrophoresis, PCR, Staphylococcal Cassette Chromosome mec (SCCmec) typing, sequence typing, incuding multilocus sequence typing and spa typing, and accessory gene regulator (Agr) typing.92–95 There have been numerous differentiation studies for S. aureus isolates from humans and animals, with 87% of human isolates being distributed among 11 clonal complexes,96 some rarer than others.97 The molecular epidemiology of S. aureus isolates from European wildlife was investigated using MLST (Multilocus sequence typing) and the isolates were assigned to numerous clonal complexes and sequence types, with those that share at least five MLST alleles being grouped into one clonal complex.98 The molecular characteristics of MRSA isolates among pigs in German farms were investigated using spa typing, MLST, and detection of toxin genes, with t011, t034, t108, t1451, and t2510, all being associated with sequence type ST398.99
Sequence type ST398 belongs to the clonal complex CC398 and is the most common sequence type of LA-MRSA found in livestock.99 Originally, sequence type ST398 was identified from S. aureus isolates isolated from a pig and other food animals,100,101 being later detected in chicken.82 Furthermore, a survey of fresh chicken and turkey meat and meat products in Germany revealed a 32% S. aureus prevalence with 87.5% of those belonging to the CC398, which is widespread among food-producing animals.102 The identified isolates were subsequently characterized using spa typing and exhibited t011, t034, t8999, t2346, and t6574.102 Although MRSA ST398 is associated with livestock, it is a major public health threat to humans in close contact with livestock due to high risk of its colonization. A very high prevalence of MRSA ST398 was reported among pig farmers and pigs in Spain, and the similar spa-types and phenotypes of resistance detected suggested animal-to-human transmission.103 The cross-transmission between animals and humans poses a high zoonotic threat and complicates treatment protocols. Of greater cause of alarm is the lack of research conducted in developing countries, especially in Africa. Although LA-MRSA has been frequently reported globally, the first detection of MRSA and (methicillin susceptible S. aureus) MSSA ST398 in food in an African country was only in 2015.79 MRSA ST398 isolate was detected from chicken samples, and was further characterized as ST398-CC398-t4358-agrLI-SCCnecIVa, resistant to β-lactams, tetracycline, and erythromycin while also harboring numerous virulence genes.104 The diversity and multiple drug resistance of this isolate highlights the importance of intense research in Africa, especially as many households depend on farming for their livelyhood. There is currently no study that reports on the prevalence of MRSA/MSSA ST398 in South African livestock and livestock production systems, proving a large gap in research that is required to be addressed. We are currently conducting research into the prevalence of MRSA/MSSA ST398 in livestock production systems in South Africa using samples from various livestock animal species, companion animals, and wild birds. We were able to isolate more than 100 isolates of livestock-associated S. aureus (unpublished data) among other important pathogens and are currently being characterized using molecular methods and whole genome sequencing. To our knowledge, this is the first study in literature investigating LA-SA from KwaZulu-Natal Province of South Africa.
Toxicity of heavy metals
The environment where bacteria are isolated and the environmental practices play an important role in the development of antibiotic resistance. Farms, abattoirs, and food production systems are home to numerous environmental contaminants from water, soil, animal excretions, and human contaminations, and sometimes runoff from paints used on the buildings. One of the most common contaminants in these environments is heavy metals, these being described as metals that have a relatively high density (>5 g/cm3) with most being toxic at low quantities.105 The most common heavy metals found in the environment include arsenic (As), lead (Pb), mercury (Hg), nickel (Ni), chromium (Cr), copper (Cu), zinc (Zn), and cadmium (Cd). As metals are naturally found in the earth’s crust and are required to maintain biochemical and physiological functions in various organisms, environmental factors and human interference can alter the natural concentrations of the heavy metals, leading to detrimental effects for animals, humans, and the environment.106 It was reported that heavy metals most commonly found in wastewater include As, Cd, Cr, Cu, Pb, Ni, and Zn.107,108 The toxic limits of the various heavy metals in animals and humans are shown in Table 2.109 Apart from the naturally occurring heavy metals on the earth’s crust, other sources of heavy metals include industrial effluents, urban runoff, sewage discharge, and chemical control agents applied to crops and livestock feeds.110,111 Of concern is that, heavy metals such as Hg, Cd, Pb, and As do not serve any essential biological function but have toxic properties, are readily transferred through food, and are not easily broken down.112
 | Table 2 Toxic limit and recommended/safe intake of heavy metal Notes: Adapted from Raikwar M, Kumar P, Singh M, Singh A. Toxic effect of heavy metals in livestock health. Vet World. 2009;1(1):28–30.109 |
One of the major sources of heavy metals is animal feeds; feed and manure samples from farms in China were investigated, and 14 metals were reported including Zn, Cu, Pb, Cd, As, and Hg.113 Copper, one of the most common heavy metals, was found to be at concentration levels as high as 1,726.3 mg/kg. Similarly, livestock feeds and manure samples from commercial farms in England and Wales were investigated to determine their heavy metal concentrations. Pig feeds showed a concentration of 150–2,920 mg Zn/kg and 18–217 mg Cu/kg while in poultry feeds, they ranged from 28 to 4,030 mg Zn/kg and 5 to 234 mg Cu/kg.114 Livestock manure was found to be responsible for an estimated 37%–40% of total Zn and Cu inputs.115 Analysis of animal feed and meat products from a metal processing region in east Kazakhstan showed high concentrations of Cd, Pb, and Zn.116 Higher levels of contamination were found in horse samples with their kidneys containing concentrations of 128 and 2.2 mg/kg for Cd and Pb, respectively, which exceeded the state maximal allowed concentrations and pose a risk to human and animal health.116
In developing countries, food safety is of great importance in both livestock abattoirs and vegetable gardens. A study conducted in South Africa showed heavy metal contamination in vegetables from residential gardens with concentration levels ranging from 0.01 to 1.12 mg/kg for Cd, 0.92 to 9.29 mg/kg for Cu, 0.04 to 373.38 mg/kg for Mg and 4.27 to 89.88 mg/kg for Zn, respectively.117 Heavy metal contamination in soil was also detected at slightly higher concentrations and although those for critical heavy metals such as Pb and Cd may not pose a grave threat, the Mg level was extremely high and Zn concentrations exceeded safe levels in various vegetables. This contamination by heavy metals showed the potential risk that underprivileged communities face as they almost exclusively depend on these vegetables. Studies show that toxic metal pollution in Africa has reached unprecedented levels and has attracted the attention of national and international environmentalists.118 In South Africa, mining is the major source of environmental pollution while Zambia holds Cu and Co deposits; with industrial, agricultural and domestic wastes having been highlighted as major contributors of heavy metal contamination. Pb, Cd, Cu, and Zn contamination in the Thohoyandou District rivers of South Africa has been attributed to the use of fertilizers, sewage treatment plants, and pesticides that contain Pb and Cd on surrounding farms.118 It was reported that the regulation of heavy metal content in meat products is lacking in many African countries despite that the general awareness of health risks associated with the consumption of heavy metal contaminated food has increased.119 Furthermore, the number of studies that have established a close association between exposure to heavy metals and cancer epidemiology in sub-Saharan Africa is increasing,120 which further demonstrates the need for research to be focused on their contamination.
Apart from contaminating the environment, heavy metals are also consumed in water and feeds that are eaten by food producing animals, which poses health threats to consumers.121 It was found that the average concentrations of Cu, Pb, and Cd in raw milk were significantly higher than the International Dairy Federation standards, showing that more active heavy metal surveillance in animal products is required.121 Similarly, a study from Nigeria showed chicken bone-muscles with concentrations of 28.23 mg/kg for Pb, 16.17 mg/kg for Zn and 4.27 mg/kg for Ni, while chicken brain had the highest levels of Cd and Mg (0.38 and 67.54 mg/kg, respectively).122 These results show that both Pb and Cd should be of great concern in Nigeria due to the contamination of meat and vegetables. In southern Nigeria, Cd (0.01-5.68 mg/kg), Ni (0.13-7.93 mg/kg), Cr (0.01-3.43 mg/kg) and Pb (0.01-4.60 mg/kg) levels were found to be above the permissible limits in chicken, chicken gizzards and turkey meat samples.123 Fish and shellfish from areas around Cochin in India showed the presence of heavy metals at varying levels, with all concentrations falling within the prescribed range for safe human consumption.124 This shows that with diligent surveillance, it is possible to achieve contamination levels that are safe for human consumption.
Heavy metal and antimicrobial resistance
Livestock farms, abattoirs, and food production systems are home to numerous heavy metals, and microorganisms are found in water, soil, livestock, food products, and livestock handlers. The cohabitation of microorganisms and heavy metals in the environment has led to the adaptation of microorganisms which develop resistance to heavy metals. Healthcare is currently challenged with a considerable burden of antimicrobial resistant microorganisms, and compounding of heavy metal resistance leads to a whole range of new alarms. Similar to how bacteria develop antibiotic resistance due to the injudicious use of antibiotics, the use of heavy metals in animal feed, environmental contamination, and therapeutic agents may enhance the development of plasmid-harboring resistant bacteria in these environments to toxic metals including Ag, Cd, Co, Cu, Hg, Ni, Pb, and Zn.125–130 Microorganisms have developed several resistance mechanisms to heavy metals including exclusion by permeability barrier, intracellular and extracellular, enzymatic detoxification, active transport efflux pumps, and reduction in the sensitivity of cellular targets to metal ions.131 It is important to understand the molecular background of microorganisms that are both antibiotic and heavy metal resistant, this aids in the continued fight for a better healthcare system for both humans and animals as well as the processes involved in producing safe food products.
The co-contamination of heavy metals and antibiotics in the environment plays an important role in the rapidly growing concern of antibiotic resistance. Heavy metal resistance mechanisms have been described to indirectly select for antibiotic resistance as well via a phenomenon known as co-selection.132 Two classes of co-selection resistance mechanisms have been described, these being: 1) cross-resistance mechanisms that provide tolerance to more than one antimicrobial agent including antibiotics and heavy metals and 2) co-resistance of two or more resistance genes that are genetically linked.133 The effects of heavy metals on antibiotic resistance in a bacterium were investigated, with the presence of As, Cu, and Zn even at low levels in the environment being found to enhance resistance of the bacterium to tetracycline.134 Heavy metal induced antibiotic resistance is abundant among various microbial species, and it is suggested that it enhances the emergence and spread of antibiotic resistance in metal/antibiotic co-contaminated environments.134 More recently, antibiotic resistance of bacteria isolated from heavy metal-polluted soils in various land types was investigated, and with soil with the highest levels having the lowest number of bacteria, but interestingly, a much higher abundance of heavy metal-resistant bacteria and antibiotic-resistant bacteria with a high degree of resistance to ampicillin and amoxicillin.135 Other prevalent antibiotic-resistant bacteria were resistant to vancomycin, tetracycline, and streptomycin. Co-resistance toward Hg and antibiotics was observed among the Gram-negative isolates while Gram-positive isolates showed co-resistance toward Zn, Ni, Hg with β-lactam antibiotics.
MRSA and heavy metal resistance
Methicillin-resistant S. aureus, being one of the most clinically important classes of pathogens globally, has also been found to harbor resistance toward heavy metals as well, further complicating the treatment of MRSA infections. The recently reported co-resistance of Gram-positive isolates to β-lactam antibiotics and heavy metals including Zn, Ni, and Hg highlights the importance of combating MRSA.135 It was reported that 74% of MRSA CC398 isolates isolated from Danish swine had reduced susceptibility to ZnCl2, suggesting that there is a positive correlation between reduced heavy metal susceptibility and MRSA.136 This correlation was further supported in another study where it was reported that 75% of LA-MRSA isolates carried at least one metal-resistant gene.137 The study showed heavy metal resistance gene prevalences of 4.8% (As), 0.2% (Cd). 24.3% (Cu) and 71.5% (Zn) respectively.137 Heavy metal resistance plays a role in the co-selection of methicillin resistance in S. aureus as was previously reported where 74% of LA-MRSA CC398 and 48% of MRSA CC398 isolated from humans were resistant to zinc chloride and tested positive for the Zn-resistance gene czrC.138 The emergence of multidrug resistance coupled with heavy metal resistance is a major concern due to the co-localization of antimicrobial and heavy metal resistance genes. It was shown that three erm(T)-carrying multiple resistance plasmids isolated from porcine and human MRSA ST398, also harbored Cd and Cu resistance determinants.138 Furthermore, the plasmids were also found to carry numerous antibiotic resistance genes including tet(L), thereby conferring resistance to tetracycline.138 Most recently, it was shown that a novel copper-resistant locus that was horizontally transferred confers hyper-resistance.139 This locus is uniquely associated with the SCCmec elements of CA-MRSA USA300 strain that is highly virulent, epidemic, and with great implications for public health.139
Heavy metal resistance, including Zn and Cu in livestock associated S. aureus, was reported in Europe. Although co-selection of resistance was observed in both methicillin-resistant and susceptible S. aureus, MRSA isolates contained more czrC gene compared to the MSSA isolates.140 No reports from South Africa have shown correlations between MRSA/MSSA ST398 and heavy metal resistance. A significant positive correlation was reported between heavy metal resistance profiles for Cu and chromate (Cr) and erythromycin resistance in Gram-negative bacteria isolated from tilapia aquaculture systems confirming that heavy metal and antibiotic resistance is co-selected in fish-associated bacteria.141 Although the study is not related to LA-MRSA, the fundamental logic behind the study is sound and shows that similar studies can be conducted in South Africa to determine correlations between heavy metal resistance and MRSA ST398.
Conclusion
S. aureus is one of the most important clinical pathogens globally, with the development of antibiotic resistance, specifically MRSA, having imposed heavy burdens on the healthcare system. The characterization of S. aureus to identify virulence factors, clonal complexes, and sequence types plays an important role in the efforts toward understanding this pathogen in order to be better prepared to develop treatment protocols. The emergence of LA-MRSA further burdens the WHO as it highlights the crucial contamination of meat products, and with a rapidly growing global population, sufficient and safe food production is required. Heavy metals have been present in the environment since the beginning of time, with research showing that with rapidly evolving microorganisms as well as human interference, they are also a major environmental, health, and food safety concern. The proven positive correlation of heavy metal resistance and MRSA highlights the vulnerabilities of both humans and animals as compounded resistance is particularly difficult to treat effectively. Most alarmingly is the lack of sufficient research conducted in Africa on LA-MRSA, heavy metal resistance, and the stacked resistance in microorganisms. There is a gap in antibiotic and heavy metal resistance reporting in South Africa, with research efforts being required to combat the rapidly evolving resistant bacteria and their associated consequences. In addition, strict measures and policies to contain antimicrobial resistance through the rational use of antimicrobial agents for veterinary and agricultural purposes including awareness campaigns on their prudent use are urgently required to control the threat of antimicrobial resistance in South Africa.
Acknowledgments
We would like to acknowledge the four anonymous reviewers who provided valuable suggestions and comments that improved the manuscript. We would like to thank the South African National Research Foundation for supporting this research through the Thuthuka Funding Instrument (grant number TTK170411226583). We would also like to thank the College of Health Sciences and the University of KwaZulu-Natal for support through internal funding.
Ethical approval
This article does not contain any experimental studies involving human participants or animals performed by any of the authors. Parts of the manuscript involving data from the ongoing research projects where ethical approvals were obtained from the Animal Research Ethics Committee of the University of KwaZulu-Natal (Reference, AREC 071/017 and AREC 014/018 for Prof ME El Zowalaty). The field sampling protocols, samples collected from animals, and the research were conducted in full compliance with Section 20 of the Animal Diseases Act of 1984 (Act No 35 of 1984) and were approved by the South African Department of Agriculture, Forestry and Fisheries DAFF (Section 20 approval Reference, 12/11/1/5 granted to Prof ME El Zowalaty).
Disclosure
The authors report no conflicts of interest in this work.
References
Foster T. Staphylococcus. Medical Microbiology. 4th ed. Baron S, editor. Galveston, TX: University of Texas Medical Branch at Galveston; 1996. | ||
Graveland H, Duim B, van Duijkeren E, Heederik D, Wagenaar JA. Livestock-associated methicillin-resistant Staphylococcus aureus in animals and humans. Int J Med Microbiol. 2011;301(8):630–634. | ||
Wertheim HF, Melles DC, Vos MC, et al. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis. 2005;5(12):751–762. | ||
Otto M. Staphylococcus aureus toxins. Curr Opin Microbiol. 2014;17:32–37. | ||
Tong SY, Davis JS, Eichenberger E, Holland TL, Fowler VG. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev. 2015;28(3):603–661. | ||
World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Geneva: World Health Organization; 2017. | ||
Tacconelli E, Carrara E, Savoldi A, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. | ||
Oliveira D, Borges A, Simões M. Staphylococcus aureus Toxins and Their Molecular Activity in Infectious Diseases. Toxins. 2018;10(6):252. | ||
Brakstad OG, Aasbakk K, Maeland JA. Detection of Staphylococcus aureus by polymerase chain reaction amplification of the nuc gene. J Clin Microbiol. 1992;30(7):1654–1660. | ||
Foster TJ. Immune evasion by staphylococci. Nat Rev Microbiol. 2005;3(12):948–958. | ||
Hennekinne JA, de Buyser ML, Dragacci S. Staphylococcus aureus and its food poisoning toxins: characterization and outbreak investigation. FEMS Microbiol Rev. 2012;36(4):815–836. | ||
Denayer S, Delbrassinne L, Nia Y, Botteldoorn N. Food-Borne Outbreak Investigation and Molecular Typing: High Diversity of Staphylococcus aureus Strains and Importance of Toxin Detection. Toxins. 2017;9(12):407. | ||
Fowler VG, Miro JM, Hoen B, et al. Staphylococcus aureus endocarditis: a consequence of medical progress. JAMA. 2005;293(24):3012–3021. | ||
Mohiyiddeen G, Brett I, Jude E. Infective endocarditis caused by Staphylococcus aureus in a patient with atopic dermatitis: a case report. J Med Case Rep. 2008;2(1):143–143. | ||
Mylonakis E, Calderwood SB, Eleftherios M, Stephen B. Infective endocarditis in adults. N Engl J Med. 2001;345(18):1318–1330. | ||
Smith TH, Fox LK, Middleton JR. Outbreak of mastitis caused by one strain of Staphylococcus aureus in a closed dairy herd. J Am Vet Med Assoc. 1998;212(4):553–556. | ||
Smyth DS, Hartigan PJ, Meaney WJ, et al. Superantigen genes encoded by the egc cluster and SaPIbov are predominant among Staphylococcus aureus isolates from cows, goats, sheep, rabbits and poultry. J Med Microbiol. 2005;54(Pt 4):401–411. | ||
Lina G, Piémont Y, Godail-Gamot F, et al. Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin Infect Dis. 1999;29(5):1128–1132. | ||
Adler A, Temper V, Block C, Abramson N, Moses A. Panton-Valentine Leukocidin–producing Staphylococcus aureus. Emerg Infect Dis. 2006;12(11):1789–1790. | ||
Ruffing U, Alabi A, Kazimoto T, et al. Community-Associated Staphylococcus aureus from Sub-Saharan Africa and Germany: A Cross-Sectional Geographic Correlation Study. Sci Rep. 2017;7(1):154. | ||
Gillet Y, Issartel B, Vanhems P, et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet. 2002;359(9308):753–759. | ||
Niemann S, Bertling A, Brodde MF, et al. Panton-Valentine Leukocidin associated with S. aureus osteomyelitis activates platelets via neutrophil secretion products. Sci Rep. 2018;8(1):2185. | ||
Melles DC, van Leeuwen WB, Boelens HA, Peeters JK, Verbrugh HA, van Belkum A. Panton-Valentine leukocidin genes in Staphylococcus aureus. Emerg Infect Dis. 2006;12(7):1174–1175. | ||
Bhakdi S, Tranum-Jensen J. Alpha-toxin of Staphylococcus aureus. Microbiol Rev. 1991;55(4):733–751. | ||
Cohen TS, Hilliard JJ, Jones-Nelson O, et al. Staphylococcus aureus α toxin potentiates opportunistic bacterial lung infections. Sci Transl Med. 2016;8(329):ra331. | ||
Xiong YQ, Willard J, Yeaman MR, Cheung AL, Bayer AS. Regulation of Staphylococcus aureus alpha-toxin gene (hla) expression by agr, sarA, and sae in vitro and in experimental infective endocarditis. J Infect Dis. 2006;194(9):1267–1275. | ||
Wieneke AA, Roberts D, Gilbert RJ. Staphylococcal food poisoning in the United Kingdom, 1969-90. Epidemiol Infect. 1993;110(3):519–531. | ||
Dinges MM, Orwin PM, Schlievert PM. Exotoxins of Staphylococcus aureus. Clin Microbiol Rev. 2000;13(1):16–34. | ||
Mckinney TK, Sharma VK, Craig WA, Archer GL. Transcription of the gene mediating methicillin resistance in Staphylococcus aureus (mecA) is corepressed but not coinduced by cognate mecA and beta-lactamase regulators. J Bacteriol. 2001;183(23):6862–6868. | ||
Peacock SJ, Paterson GK. Mechanisms of Methicillin Resistance in Staphylococcus aureus. Annu Rev Biochem. 2015;84:577–601. | ||
Fishovitz J, Hermoso JA, Chang M, Mobashery S. Penicillin-binding protein 2a of methicillin-resistant Staphylococcus aureus. IUBMB Life. 2014;66(8):572–577. | ||
Hartman BJ, Tomasz A. Expression of methicillin resistance in heterogeneous strains of Staphylococcus aureus. Antimicrob Agents Chemother. 1986;29(1):85–92. | ||
Santiago C, Pang EL, Lim KH, Loh HS, Ting KN. Inhibition of penicillin-binding protein 2a (PBP2a) in methicillin resistant Staphylococcus aureus (MRSA) by combination of ampicillin and a bioactive fraction from Duabanga grandiflora. BMC Complement Altern Med. 2015;15(1):178. | ||
Peacock SJ, Moore CE, Justice A, et al. Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect Immun. 2002;70(9):4987–4996. | ||
Etebu E, Arikekpar I. Antibiotics: classification and mechanisms of action with emphasis on molecular perspectives. Int J Appl Microbiol Biotechnol Res. 2016;4:90–101. | ||
Dowling A, O’Dwyer J, Adley C. Antibiotics: mode of action and mechanisms of resistance. In: Méndez-Vilas, A, editor. Antimicrobial Research: Novel Bioknowledge and Educational Programs; Volume 1. Badajoz: Formatex Research Centre; 2013:536–545. | ||
Rayner C, Munckhof WJ. Antibiotics currently used in the treatment of infections caused by Staphylococcus aureus. Intern Med J. 2005;35(s2):S3–S16. | ||
Shapiro E. Injudicious antibiotic use: an unforeseen consequence of the emphasis on patient satisfaction? Clin Ther. 2002;24(1):1 97–204. | ||
Morgan DJ, Okeke IN, Laxminarayan R, Perencevich EN, Weisenberg S. Non-prescription antimicrobial use worldwide: a systematic review. Lancet Infect Dis. 2011;11(9):692–701. | ||
Paruk F, Richards G, Scribante J, Bhagwanjee S, Mer M, Perrie H. Antibiotic prescription practices and their relationship to outcome in South Africa: findings of the prevalence of infection in South African intensive care units (PISA) study. S Afr Med J. 2012;102(7):613–616. | ||
Truter I. Antimicrobial prescribing in South Africa using a large pharmacy database: A drug utilisation study Ilse Truter. S Afr J Infect Dis. 2015;30(2):52–56. | ||
Chantziaras I, Boyen F, Callens B, Dewulf J. Correlation between veterinary antimicrobial use and antimicrobial resistance in food-producing animals: a report on seven countries. J Antimicrob Chemother. 2014;69(3):827–834. | ||
Andersson DI, Hughes D. Persistence of antibiotic resistance in bacterial populations. FEMS Microbiol Rev. 2011;35(5):901–911. | ||
Livermore DM. Epidemiology of antibiotic resistance. Intensive Care Med. 2000;26 Suppl 1:S014–S021. | ||
Raffi M, Mehrwan S, Bhatti TM, et al. Investigations into the antibacterial behavior of copper nanoparticles against Escherichia coli. Ann Microbiol. 2010;60(1):75–80. | ||
Usman MS, El Zowalaty ME, Shameli K, Zainuddin N, Salama M, Ibrahim NA. Synthesis, characterization, and antimicrobial properties of copper nanoparticles. Int J Nanomedicine. 2013;8:4467–4479. | ||
Levy SB, Marshall B. Antibacterial resistance worldwide: causes, challenges and responses. Nat Med. 2004;10(12 Suppl):S122–S129. | ||
European Centre for Disease Prevention and Control. Antimicrobial consumption. ECDC. Annual epidemiological report for 2014. Stockholm, Sweden; 2018. | ||
van Boeckel TP, Gandra S, Ashok A, et al. Global antibiotic consumption 2000 to 2010: an analysis of national pharmaceutical sales data. Lancet Infect Dis. 2014;14(8):742–750. | ||
Edelsberg J, Weycker D, Barron R, et al. Prevalence of antibiotic resistance in US hospitals. Diagn Microbiol Infect Dis. 2014;78(3):255–262. | ||
Xi C, Zhang Y, Marrs CF, et al. Prevalence of antibiotic resistance in drinking water treatment and distribution systems. Appl Environ Microbiol. 2009;75(17):5714–5718. | ||
Ntirenganya C, Manzi O, Muvunyi CM, Ogbuagu O. High prevalence of antimicrobial resistance among common bacterial isolates in a tertiary healthcare facility in Rwanda. Am J Trop Med Hyg. 2015;92(4):865–870. | ||
Bernabé KJ, Langendorf C, Ford N, Ronat JB, Murphy RA. Antimicrobial resistance in West Africa: a systematic review and meta-analysis. Int J Antimicrob Agents. 2017;50(5):629–639. | ||
Jacobs MR, Koornhof HJ, Robins-Browne RM, et al. Emergence of multiply resistant pneumococci. N Engl J Med. 1978;299(14):735–740. | ||
World Health Organization. Antimicrobial Resistance: Global Report on Surveillance. World Health Organization; 2014. Available from: http://apps.who.int/iris/bitstream/handle/10665/112642/?sequence=1. Accessed May 5, 2018. | ||
El Zowalaty ME, Al Thani AA, Webster TJ, et al. Pseudomonas aeruginosa: arsenal of resistance mechanisms, decades of changing resistance profiles, and future antimicrobial therapies. Future Microbiol. 2015;10(10):1683–1706. | ||
Pagel SW, Gautier P. Use of antimicrobial agents in livestock. Rev Sci Tech. 2012;31(1):145–188. | ||
Dupont HL, Steele JH. Use of antimicrobial agents in animal feeds: implications for human health. Rev Infect Dis. 1987;9(3):447–460. | ||
Mellon M, Benbrook C, Benbrook KL. Hogging it: Estimates of Antimicrobial Abuse in Livestock. MA, USA: Union of Concerned Scientists Publications, Two Brattle Square Cambridge; 2001:7–9. Available from: www.ucsusa.org/publications. Accessed August 9, 2018. | ||
Shea KM, American Academy of Pediatrics Committee on Environmental Health, American Academy of Pediatrics Committee on Infectious Diseases. Nontherapeutic use of antimicrobial agents in animal agriculture: implications for pediatrics. Pediatrics. 2004;114(3):862–868. | ||
Goldman E. Antibiotic abuse in animal agriculture: Exacerbating drug resistance in human pathogens. Human Ecological Risk Assessment. 2004;10(1):121–134. | ||
Landers TF, Cohen B, Wittum TE, Larson EL. A review of antibiotic use in food animals: perspective, policy, and potential. Public Health Rep. 2012;127(1):4–22. | ||
van Boeckel TP, Brower C, Gilbert M, et al. Global trends in antimicrobial use in food animals. Proc Nat Acad Sci. 2015;112(18):5649–5654. | ||
Martin MJ, Thottathil SE, Newman TB. Antibiotics Overuse in Animal Agriculture: A Call to Action for Health Care Providers. American Public Health Association; 2015. | ||
Sneeringer S, MacDonald JM, Key N, McBride WD, Mathews K. Economics of Antibiotic Use in U.S. Livestock Production, ERR-200, U.S. Department of Agriculture, Economic Research Service. Vol. 2017. United States Department of Agriculture; 2015. Available from: https://ageconsearch.umn.edu/bitstream/229202/2/err200.pdf. Accessed October 9, 2018. | ||
Chambers HF, Deleo FR. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat Rev Microbiol. 2009;7(9):629–641. | ||
Jevons MP. “Celbenin” - resistant Staphylococci. BMJ. 1961;1(5219):124–125. | ||
Knox R. “Celbenin”-resistant Staphylococci. BMJ. 1960;2(5205):1085. | ||
Barber M. Methicillin-resistant staphylococci. J Clin Pathol. 1961;14(4):385–393. | ||
Çetin ET, Ang Ö. Staphylococci resistant to methicillin (“Celbenin”). Br Med J. 1962;2(5296):51–52. | ||
Alm RA, Mclaughlin RE, Kos VN, Sader HS, Iaconis JP, Lahiri SD. Analysis of Staphylococcus aureus clinical isolates with reduced susceptibility to ceftaroline: an epidemiological and structural perspective. J Antimicrob Chemother. 2014;69(8):2065–2075. | ||
Mendes RE, Tsakris A, Sader HS, et al. Characterization of methicillin-resistant Staphylococcus aureus displaying increased MICs of ceftaroline. J Antimicrob Chemother. 2012;67(6):1321–1324. | ||
Jevons MP, Parker MT. The evolution of new hospital strains of Staphylococcus aureus. J Clin Pathol. 1964;17(3):243–250. | ||
Stryjewski ME, Chambers HF. Skin and Soft-Tissue Infections Caused by Community-Acquired Methicillin-Resistant Staphylococcus aureus. Clinical Infectious Diseases. 2008;46(S5):S368–S377. | ||
Falagas ME, Karageorgopoulos DE, Leptidis J, Korbila IP. MRSA in Africa: filling the global map of antimicrobial resistance. PLoS One. 2013;8(7):e68024. | ||
David MZ, Glikman D, Crawford SE, et al. What Is Community-Associated Methicillin-Resistant Staphylococcus aureus? J Infect Dis. 2008;197(9):1235–1243. | ||
Otto M. Community-associated MRSA: what makes them special? Int J Med Microbiol. 2013;303(6-7):324–330. | ||
Voss A, Loeffen F, Bakker J, Klaassen C, Wulf M. Methicillin-resistant Staphylococcus aureus in pig farming. Emerg Infect Dis. 2005;11(12):1965-6. | ||
Crombé F, Willems G, Dispas M, et al. Prevalence and antimicrobial susceptibility of methicillin-resistant Staphylococcus aureus among pigs in Belgium. Microb Drug Resist. 2012;18(2):125–131. | ||
van Cleef BA, Monnet DL, Voss A, et al. Livestock-associated methicillin-resistant Staphylococcus aureus in humans, Europe. Emerg Infect Dis. 2011;17(3):502–505. | ||
Smith TC, Male MJ, Harper AL, et al. Methicillin-resistant Staphylococcus aureus (MRSA) strain ST398 is present in midwestern U.S. swine and swine workers. PLoS One. 2009;4(1):e4258. | ||
Nemati M, Hermans K, Lipinska U, et al. Antimicrobial resistance of old and recent Staphylococcus aureus isolates from poultry: first detection of livestock-associated methicillin-resistant strain ST398. Antimicrob Agents Chemother. 2008;52(10):3817–3819. | ||
Mulders MN, Haenen AP, Geenen PL, et al. Prevalence of livestock-associated MRSA in broiler flocks and risk factors for slaughterhouse personnel in The Netherlands. Epidemiol Infect. 2010;138(5):743–755. | ||
Paterson GK, Larsen J, Harrison EM, et al. First detection of livestock-associated meticillin-resistant Staphylococcus aureus CC398 in bulk tank milk in the United Kingdom, January to July 2012. Euro Surveill. 2012;17(50):20337. | ||
Mkize N, Zishiri OT, Mukaratirwa S. Genetic characterisation of antimicrobial resistance and virulence genes in Staphylococcus aureus isolated from commercial broiler chickens in the Durban metropolitan area, South Africa. J S Afr Vet Assoc. 2017;88(0):e1–e7. | ||
de Boer E, Zwartkruis-Nahuis J, Wit B, et al. Prevalence of methicillin-resistant Staphylococcus aureus in meat. Intern J Food Microbiol. 2009;134(1-2):52–56. | ||
Weese JS, Avery BP, Reid-Smith RJ. Detection and quantification of methicillin-resistant Staphylococcus aureus (MRSA) clones in retail meat products. Lett App Microbiol. 2010;51(3):338–342. | ||
Kim YJ, Dh O, Song BR, et al. Molecular Characterization, Antibiotic Resistance, and Virulence Factors of Methicillin-Resistant Staphylococcus aureus Strains Isolated from Imported and Domestic Meat in Korea. Foodborne Pathog Dis. 2015;12(5):390–398. | ||
Jackson CR, Davis JA, Barrett JB. Prevalence and characterization of methicillin-resistant Staphylococcus aureus isolates from retail meat and humans in Georgia. J Clin Microbiol. 2013;51(4):1199–1207. | ||
Pantosti A. Methicillin-Resistant Staphylococcus aureus Associated with Animals and Its Relevance to Human Health. Front Microbiol. 2012;3:127. | ||
Genetic characterization and its use in decision-making for the conservation of crop germplasm. The Role of Biotechnology in Exploring and Protecting Agricultural Genetic Resources. Available at: http://www.fao.org/biotech/docs/vicente.pdf. Accessed October 18, 2018. | ||
Schmitz FJ, Steiert M, Tichy HV, et al. Typing of methicillin-resistant Staphylococcus aureus isolates from Düsseldorf by six genotypic methods. J Med Microbiol. 1998;47(4):341–351. | ||
Strandén A, Frei R, Widmer AF. Molecular typing of methicillin-resistant Staphylococcus aureus: can PCR replace pulsed-field gel electrophoresis? J Clin Microbiol. 2003;41(7):3181–3186. | ||
Koreen L, Ramaswamy SV, Graviss EA, Naidich S, Musser JM, Kreiswirth BN. spa typing method for discriminating among Staphylococcus aureus isolates: implications for use of a single marker to detect genetic micro- and macrovariation. J Clin Microbiol. 2004;42(2):792–799. | ||
Makgotlho PE, Kock MM, Hoosen A, et al. Molecular identification and genotyping of MRSA isolates. FEMS Immunol Med Microbiol. 2009;57(2):104–115. | ||
Cuny C, Wieler LH, Witte W. Livestock-Associated MRSA: The Impact on Humans. Antibiotics. 2015;4(4):521–543. | ||
Monecke S, Gavier-Widén D, Hotzel H, et al. Diversity of Staphylococcus aureus Isolates in European Wildlife. PLoS One. 2016;11(12):e0168433. | ||
Feil EJ, Cooper JE, Grundmann H, et al. How clonal is Staphylococcus aureus? J Bacteriol. 2003;185(11):3307–3316. | ||
Köck R, Harlizius J, Bressan N, et al. Prevalence and molecular characteristics of methicillin-resistant Staphylococcus aureus (MRSA) among pigs on German farms and import of livestock-related MRSA into hospitals. Eur J Clin Microbiol Infect Dis. 2009;28(11):1375–1382. | ||
Voss A, Loeffen F, Bakker J, Klaassen C, Wulf M. Methicillin-resistant Staphylococcus aureus in pig farming. Emerg Infect Dis. 2005;11(12):1965-6–1966. | ||
Witte W, Strommenger B, Stanek C, Cuny C. Methicillin-resistant Staphylococcus aureus ST398 in humans and animals, Central Europe. Emerg Infect Dis. 2007;13(2):255–258. | ||
Fessler AT, Kadlec K, Hassel M, et al. Characterization of methicillin-resistant Staphylococcus aureus isolates from food and food products of poultry origin in Germany. Appl Environ Microbiol. 2011;77(20):7151–7157. | ||
Reynaga E, Navarro M, Vilamala A, et al. Prevalence of colonization by methicillin-resistant Staphylococcus aureus ST398 in pigs and pig farm workers in an area of Catalonia, Spain. BMC Infect Dis. 2016;16(1):716. | ||
Chairat S, Gharsa H, Lozano C, et al. Characterization of Staphylococcus aureus from Raw Meat Samples in Tunisia: Detection of Clonal Lineage ST398 from the African Continent. Foodborne Pathog Dis. 2015;12(8):686–692. | ||
Pandey G, Madhuri S. Heavy metals causing toxicity in animals and fishes. Research Journal of Animal, Veterinary and Fishery Sciences. 2014;2(2):17–23. | ||
Khlifi R, Hamza-Chaffai A. Head and neck cancer due to heavy metal exposure via tobacco smoking and professional exposure: a review. Toxicol Appl Pharmacol. 2010;248(2):71–88. | ||
Lambert M, Leven B, Green R. New methods of cleaning up heavy metal in soils and water. Environmental Science and Technology Briefs for Citizens. Manhattan, KS: Kansas State University; 2000. Available from: http://www.engg.ksu.edu/HSRC/Tosc/metals.pdf. September 15, 2018. | ||
Smith CJ, Hopmans P, Cook FJ. Accumulation of Cr, Pb, Cu, Ni, Zn and Cd in soil following irrigation with treated urban effluent in Australia. Environ Pollut. 1996;94(3):317–323. | ||
Raikwar M, Kumar P, Singh M, Singh A. Toxic effect of heavy metals in livestock health. Vet World. 2009;1(1):28–30. | ||
Alloway BJ. Sources of Heavy Metals and Metalloids in Soils. In: Alloway B, editor. Heavy Metals in Soils. Environmental Pollution; volume 22. Dordrecht: Springer; 2013:11-50. | ||
Morais S, e Costa FG, de Lourdes Pereira M. Heavy metals and human health. Environmental Health-Emerging Issues and Practice Oosthuizen Jacques, editor; 2012. Available from: https://www.intechopen.com/download/pdf/27687. Accessed October 17, 2018. | ||
López-Alonso M. Animal feed contamination by toxic metals. In: Fink-Gremmels J, editor. Animal Feed Contamination. Effects on livestock and food safety. Sawston: Woodhead Publishing; 2012;183–204. | ||
Cang L, Wang YJ, Zhou DM, Dong YH. Heavy metals pollution in poultry and livestock feeds and manures under intensive farming in Jiangsu Province, China. J Environ Sci. 2004;16(3):371–374. | ||
Nicholson FA, Chambers BJ, Williams JR, Unwin RJ. Heavy metal contents of livestock feeds and animal manures in England and Wales. Bioresour Technol. 1999;70(1):23–31. | ||
Nicholson FA, Smith SR, Alloway BJ, Carlton-Smith C, Chambers BJ. An inventory of heavy metals inputs to agricultural soils in England and Wales. Sci Total Environ. 2003;311(1-3):205–219. | ||
Farmer AA, Farmer AM. Concentrations of cadmium, lead and zinc in livestock feed and organs around a metal production centre in eastern Kazakhstan. Sci Total Environ. 2000;257(1):53–60. | ||
Bvenura C, Afolayan AJ. Heavy metal contamination of vegetables cultivated in home gardens in the Eastern Cape. S Afr J Sci. 2012;108(9/10):1–6. | ||
Yabe J, Ishizuka M, Umemura T. Current levels of heavy metal pollution in Africa. J Vet Med Sci. 2010;72(10):1257–1263. | ||
Sedki A, Lekouch N, Gamon S, Pineau A. Toxic and essential trace metals in muscle, liver and kidney of bovines from a polluted area of Morocco. Sci Total Environ. 2003;317(1-3):201–205. | ||
Fasinu P, Orisakwe OE. Heavy metal pollution in sub-Saharan Africa and possible implications in cancer epidemiology. Asian Pac J Cancer Prev. 2013;14(6):3393–3402. | ||
Younus M, Abbas T, Zafar M, et al. Assessment of heavy metal contamination in raw milk for human consumption. S Afr J Anim Sci. 2016;46(2):166–169. | ||
Orisakwe OE, Oladipo OO, Ajaezi GC, Udowelle NA. Horizontal and Vertical Distribution of Heavy Metals in Farm Produce and Livestock around Lead-Contaminated Goldmine in Dareta and Abare, Zamfara State, Northern Nigeria. J Environ Public Health. 2017;2017:3506949–12. | ||
Iwegbue CMA, Nwajei GE, Iyoha EH. Heavy metal residues of chicken meat and gizzard and turkey meat consumed in southern Nigeria. Bulgarian J Vet Med. 2008;11(4):275–280. | ||
Sivaperumal P, Sankar T, Viswanathannair P. Heavy metal concentrations in fish, shellfish and fish products from internal markets of India vis-a-vis international standards. Food Chem. 2007;102(3):612–620. | ||
Baquero F, Martínez JL, Cantón R. Antibiotics and antibiotic resistance in water environments. Curr Opin Biotechnol. 2008;19(3):260–265. | ||
Ji X, Shen Q, Liu F, et al. Antibiotic resistance gene abundances associated with antibiotics and heavy metals in animal manures and agricultural soils adjacent to feedlots in Shanghai; China. J Hazard Mater. 2012;235-236:178–185. | ||
Martinez JL. Environmental pollution by antibiotics and by antibiotic resistance determinants. Environ Pollut. 2009;157(11):2893–2902. | ||
Zhu YG, Johnson TA, Su JQ, et al. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc Natl Acad Sci U S A. 2013;110(9):3435–3440. | ||
Zhao Z, Wang J, Han Y, et al. Nutrients, heavy metals and microbial communities co-driven distribution of antibiotic resistance genes in adjacent environment of mariculture. Environ Pollut. 2017;220(Pt B):909–918. | ||
Silver S. Bacterial resistances to toxic metal ions--a review. Gene. 1996;179(1):9–19. | ||
Bruins MR, Kapil S, Oehme FW. Microbial resistance to metals in the environment. Ecotoxicol Environ Saf. 2000;45(3):198–207. | ||
Koditschek LK, Guyre P. Resistance transfer fecal coliforms isolated from the whippany river. Water Res. 1974;8(10):747–752. | ||
Chapman JS. Disinfectant resistance mechanisms, cross-resistance, and co-resistance. Int Biodeterior Biodegradation. 2003;51(4):271–276. | ||
Chen S, Li X, Sun G, Zhang Y, Su J, Ye J. Heavy metal induced antibiotic resistance in bacterium LSJC7. Int J Mol Sci. 2015;16(10):23390–23404. | ||
Safari Sinegani AA, Younessi N. Antibiotic resistance of bacteria isolated from heavy metal-polluted soils with different land uses. J Glob Antimicrob Resist. 2017;10:247–255. | ||
Aarestrup FM, Cavaco L, Hasman H. Decreased susceptibility to zinc chloride is associated with methicillin resistant Staphylococcus aureus CC398 in Danish swine. Vet Microbiol. 2010;142(3-4):455–457. | ||
Argudín MA, Lauzat B, Kraushaar B, et al. Heavy metal and disinfectant resistance genes among livestock-associated methicillin-resistant Staphylococcus aureus isolates. Vet Microbiol. 2016;191:88–95. | ||
Gómez-Sanz E, Kadlec K, Feßler AT, Zarazaga M, Torres C, Schwarz S. Novel erm(T)-carrying multiresistance plasmids from porcine and human isolates of methicillin-resistant Staphylococcus aureus ST398 that also harbor cadmium and copper resistance determinants. Antimicrob Agents Chemother. 2013;57(7):3275–3282. | ||
Purves J, Thomas J, Riboldi GP, et al. A horizontally gene transferred copper resistance locus confers hyper-resistance to antibacterial copper toxicity and enables survival of community acquired methicillin resistant Staphylococcus aureus USA300 in macrophages. Environ Microbiol. 2018;20(4):1576–1589. | ||
Nair R, Thapaliya D, Su Y, Smith TC. Resistance to zinc and cadmium in Staphylococcus aureus of human and animal origin. Infect Control Hosp Epidemiol. 2014;35 Suppl 3(S3):S32–S39. | ||
Chenia HY, Jacobs A. Antimicrobial resistance, heavy metal resistance and integron content in bacteria isolated from a South African tilapia aquaculture system. Dis Aquat Organ. 2017;126(3):199–209. | ||
United States General Accounting Office. Food Safety. The agricultural use of antibiotics and its implications for human health; 1999. Available at: https://www.gao.gov/archive/1999/rc99074.pdf. Accessed July 12, 2018. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.