")
Back to Journals » Drug Design, Development and Therapy » Volume 14
Metformin Inhibits Propofol-Induced Apoptosis of Mouse Hippocampal Neurons HT-22 Through Downregulating Cav-1
Authors Ge J, Huang Y, Zhang Y, Liu L, Gu T, Liu X, Yao L, Cai M, Sun J, Song J
Received 2 September 2019
Accepted for publication 13 December 2019
Published 21 April 2020 Volume 2020:14 Pages 1561—1569
DOI https://doi.org/10.2147/DDDT.S229520
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Qiongyu Guo
Jianyun Ge,1,* Yulin Huang,2,* Yi Zhang,3 Lin Liu,1 Tianyu Gu,1 Xu Liu,1 Lei Yao,1 Mengmeng Cai,1 Jiafeng Sun,1 Jie Song1
1Department of Anesthesiology, The Second Affiliated Hospital of Nantong University, Nantong, Jiangsu 226001, People’s Republic of China; 2Department of Anesthesiology, Affiliated Drum Tower Hospital of Medical School of Nanjing University, Nanjing, Jiangsu 210000, People’s Republic of China; 3Department of Neurosurgery, The Second Affiliated Hospital of Nantong University, Nantong, Jiangsu 226001, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jie Song
Department of Anesthesiology, The Second Affiliated Hospital of Nantong University, No. 6 North Haier Lane Road, Nantong 226001, Jiangsu, People’s Republic of China
Email [email protected]
Objective: To elucidate the neuroprotective function of metformin in suppressing propofol-induced apoptosis of HT-22 cells.
Methods: HT-22 cells were treated with 0, 10 or 100 μmol/L propofol, followed by determination of their proliferative ability. Subsequently, changes in proliferation and apoptosis of propofol-treated HT-22 cells induced with metformin were assessed. Apoptosis-associated genes in HT-22 cells were detected by Western blot. At last, regulatory effects of Cav-1 on propofol and metformin-treated HT-22 cells were examined.
Results: Propofol treatment dose-dependently decreased proliferative ability and increased apoptosis ability in HT-22 cells, which were partially blocked by metformin administration. Upregulated Bcl-2 and downregulated Bax were observed in propofol-treated HT-22 cells following metformin administration. In addition, Cav-1 level in HT-22 cells was regulated by metformin treatment. Notably, metformin reversed propofol-induced apoptosis stimulation and proliferation decline in HT-22 cells via downregulating Cav-1.
Conclusion: In our study, we found that propofol could induce apoptosis of HT-22 cells and metformin could rescue the apoptosis effect regulated by propofol. Then, we found that metformin protects propofol-induced neuronal apoptosis via downregulating Cav-1.
Keywords: metformin, propofol, Cav-1, apoptosis
Introduction
Propofol (2,6-diisopropylphenol) belongs to a type of short-acting intravenous anesthetics. Propofol is commonly used to induce and maintain anesthesia in adults owing to its rapid induction and recovery time.1,2 Since it has a good antiemetic effect, propofol is also widely applied in the induction and maintenance of pediatric and obstetric anesthesia.3 Nevertheless, a certain dose of propofol may induce neurotoxicity as the nervous system is extremely sensitive to changes in internal and external environment.4,5 Recent studies reported that propofol administration would induce neuronal apoptosis6,7 and may impair learning, memory and cognitive function in children during brain development.8 Therefore, it is necessary to clarify the mechanism underlying the neurotoxicity of propofol. So far, several mechanisms have been identified for explaining propofol-induced neurotoxicity, including mitochondrial dysfunction, translocation of apoptosis-inducing factors and the mTOR pathway.9,10 It is worth noting that propofol can regulate a variety of cellular pathways.10
Apoptosis results from changes in plasma membrane phospholipids, cell shrinkage and nuclear DNA condensation and cleavage.11 Apoptosis is regulated by abundant genes, especially the Bcl-2 family.12 Among them, Bax, Bad and Bak exert pro-apoptotic effects, while Bcl-2 and Bcl-xL are anti-apoptotic genes.13 The balance between expression levels of pro-apoptotic proteins (Bax, Bak, and Bad) and anti-apoptotic proteins (Bcl-2 and Bcl-xL) is critical for neuronal survival.14 Apoptosis may be a crucial target during the process of propofol-induced neurotoxicity. Suppression of neuronal apoptosis could be a novel strategy for the treatment of propofol-induced neurotoxicity.
Metformin is a first-line antidiabetic drug. Metformin exerts hypoglycemic outcomes through reducing hepatic gluconeogenesis, increasing glucose utilization and improving insulin sensitivity.15 Abundant evidence have suggested the potential function of metformin in anti-inflammation, anti-apoptosis and anti-oxidation in central nervous system diseases (ie, ischemic stroke, intracerebral hemorrhage and multiple sclerosis).16 As a result, metformin is believed to exert the function of neuronal protection.
Caveolins is a membrane protein family composed of three members with 21–24 kDa. Caveolin-1 (Cav-1) is abundantly expressed in endothelial cells, and caveolin-3 (CAV-3) is mainly expressed in muscle cells.17 Cav-1 is a structural protein responsible for maintaining vesicle morphology. It is involved in the disease progression of ischemia/reperfusion (I/R).18–20 A recent study demonstrated the extensive functions of Cav-1 in regulating multiple cellular behaviors.21 A recent study reported that Cav-1 knockdown increases the therapeutic sensitivity of lung cancer to cisplatin-induced apoptosis.22 Besides, β-Carotene induces apoptosis in esophageal squamous cell Carcinoma via the Cav-1/AKT/NF-κB signaling pathway.23 The previous study showed that Cav-1was strongly associated with apoptosis. However, the role of Cav-1 in propofol-induced apoptosis remains unclear.
In this paper, we mainly explored the potential regulatory effects of metformin and Cav-1 on propofol-induced neurotoxicity.
Methods
Cell Culture of HT-22
Immortalized mouse hippocampal neuron HT-22 (CL-0595) cells were provided by Cell Bank, Shanghai. Cells were cultured in DMEM containing 10% FBS, 100 μ/mL penicillin and 100 ng/mL streptomycin (Invitrogen, Carlsbad, CA, USA). They were maintained at 37°C, 5% CO2. Cells were inoculated in 6-well plates with 1×105 cells per well, and incubated with 0, 1, 10 or 100 μmol/L propofol for 24 hrs, respectively.
Transfection
HT-22 cells were treated with 100 μmol/L propofol and 10 μmol/L metformin for 24 h. Then, si-NC, si-Cav-1, pcDNA-NC and pcDNA-Cav-1 purchased from Invitrogen (Invitrogen, CA, USA) were transfected into HT-22 cells using Lipofectamine 3000. After transfection for 4–6 h, medium containing 10% FBS was replaced.
Cell Counting Kit-8 (CCK-8) Assay
Cells were seeded in the 96-well plate and cultured for 80% confluence. Twenty microliters of CCK-8 (Houston TX, USA) solution was added in each well and reacted at 37°C in dark for 2 h. Absorbance (A) at 450 nm was recorded at the appointed time points using the CCK-8 kit (Dojindo Laboratories, Kumamoto, Japan) for depicting the viability curves.
5-Ethynyl-2ʹ-Deoxyuridine (EdU) Assay
Cells were labeled with 100 μL of EdU (Ribobio, Guangzhou, China) reagent (50 μmol/L) per well for 2 h. After PBS washing, cells were fixed in 50 μL of fixation buffer, decolored with 2 mg/mL glycine and permeated with 100 μL of 0.5%Triton X-100 (Solarbio, Beijing, China). Cells were dyed with Apollo for 30 min, followed by Hoechst in dark for 10 min. EdU-positive cells, Hoechst-labeled cells and their merged images were determined under a fluorescent microscope.
Flow Cytometry
Cells were prepared into suspension with 1×106/mL. One hundred microliters of suspension was added in each tube, incubated with 10 μL of AnnexinV, 380 μL of buffer and 10 μL of PI (propidium iodide). After 15-min incubation in dark, apoptosis was analyzed by a BD FACSCalibur flow cytometer (BD Bioscience, USA).
TdT-Mediated dUTP Nick-End Labeling (TUNEL)
For TUNEL assay (Roche, Shanghai, China), cells were subjected to 30-min fixation in 4% paraformaldehyde, followed by 30-min incubation in H2O2 to inactivate the endogenous enzyme. Cells were immersed in 0.2% Triton X-100 solution for 5 min to enhance cell membrane permeability, and further incubated with deoxynucleotide terminal transferase (rTdT) at 37°C for 1 h. The cell nucleus was stained brown. Each section was randomly selected for 5 fields. Apoptotic rate (TUNEL-positive ratio) was finally calculated (magnification 400×).
Western Blot
Cellular protein was extracted and quantified by BCA method. After electrophoresis, proteins were transferred on a PVDF membrane. Membranes were immersed in 5% skim milk for blocking the non-specific sites. Two hours later, membranes were reacted with primary and secondary antibodies. Band exposure was achieved by ECL and analyzed by Image Software. The antibodies purchased from Proteintech used in our research were as follows: Bax (50599-2-Ig, 1:1000), Bcl-2 (26593-1-AP, 1:1000), Cav-1 (16447-1-AP, 1:1000), Gapdh (60004-1-Ig, 1:1000).
Statistical Processing
SPSS 22.0 was used for data analyses. Data were expressed as mean ± standard deviation. Differences were analyzed by one-way ANOVA. P < 0.05 was considered as statistically significant.
Results
Propofol Administration Inhibited Proliferation and Induced Apoptosis in HT-22 Cells
CCK-8 assay revealed a dose-dependent decline in the viability of HT-22 cells after propofol administration (Figure 1A). EdU assay further depicted the dose-dependently declined EdU-positive ratio in propofol-treated HT-22 cells (Figure 1B and C). After treatment of increased doses of propofol, the apoptotic rate gradually increased (Figure 1D and E). TUNEL-positive ratio was dose-dependently elevated by propofol treatment in HT-22 cells (Figure 1F and G). Apoptosis-associated genes were determined by Western blot. As data revealed, Bcl-2 was downregulated and Bax was upregulated in propofol-treated hippocampal neurons in a dose-dependent way (Figure 1H).
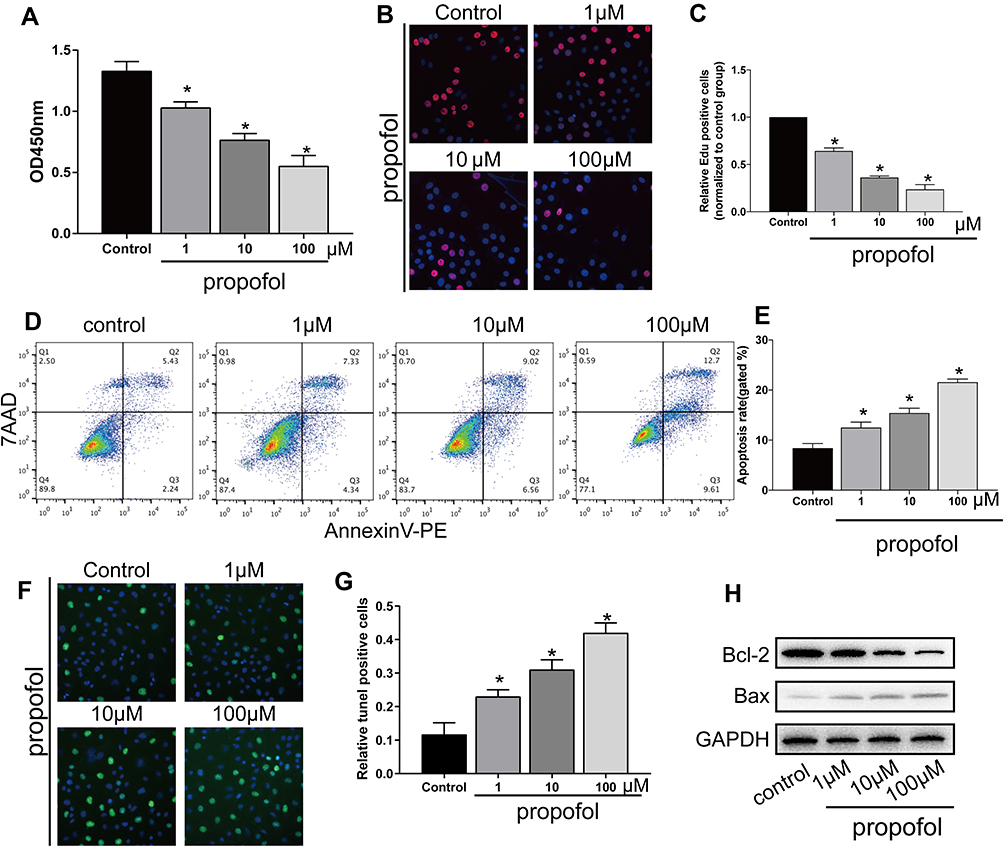 |
Figure 1 Propofol-induced apoptosis in HT-22 cells. (A) CCK-8 assay results showed viability in HT-22 cells treated with 0, 1, 10 and 100 μM propofol, respectively. (B and C) EdU assay results showed EdU-positive HT-22 cells treated with 0, 1, 10 and 100 μM propofol, respectively (B). Quantitative analysis of EdU-positive ratio (C). (D and E) Flow cytometry results showed distribution of apoptotic cells, necrotic cells and survival cells following the treatment of 0, 1, 10 and 100 μM propofol in HT-22 cells, respectively (D). Quantitative analysis of apoptosis rate (E). (F and G) TUNEL results showed TUNEL-positive cells following the treatment of 0, 1, 10 and 100 μM propofol in HT-22 cells, respectively (F). Quantitative analysis of TUNEL-positive rate (G). (H) Protein levels of Bcl-2 and Bax in HT-22 cells treated with 0, 1, 10 and 100 μM propofol, respectively (*p<0.05 compared to control group). |
Metformin Treatment Reversed Propofol-Induced Apoptosis in HT-22 Cells
To elucidate the influence of metformin on HT-22 cells, they were administrated with metformin and propofol. Interestingly, the declined viability owing to propofol treatment was reversed following metformin administration (Figure 2A). Similarly, decreased EdU-positive ratio in propofol-treated HT-22 cells was partially blocked by metformin (Figure 2B and C). Decreased apoptotic rate was observed after metformin administration in propofol-treated HT-22 cells (Figure 2D and E). Compared with those treated with propofol, TUNEL-positive ratio decreased in HT-22 cells treated with both propofol and metformin (Figure 2F and G). As data revealed, Bcl-2 was downregulated and Bax was upregulated in propofol-treated hippocampal neurons which were reversed by metformin (Figure 2H). As a result, metformin effectively reversed propofol-induced proliferation inhibition and apoptosis stimulation in hippocampal neurons.
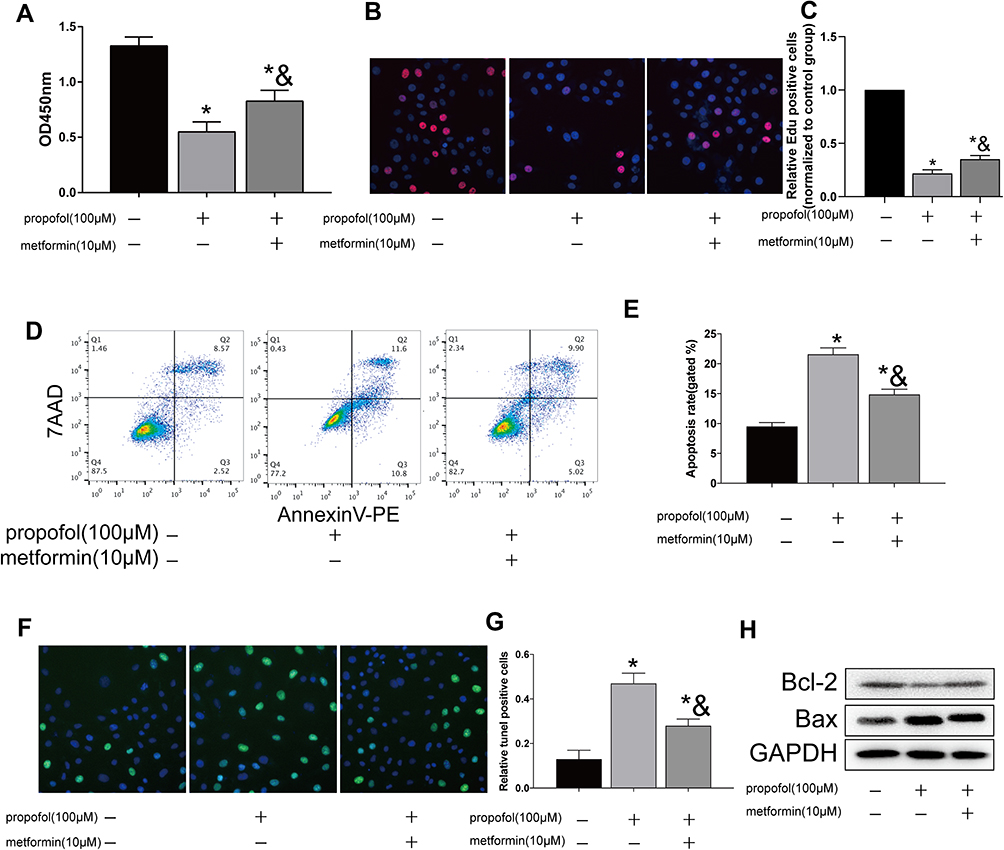 |
Figure 2 Metformin reversed propofol-induced apoptosis in HT-22 cells (A) CCK-8 assay results showed viability in propofol-induced HT-22 cells either treated with 10 μM metformin or not. (B and C) EdU assay results showed EdU-positive HT-22 cells with propofol induction, followed by 10 μM metformin treatment or not (B). Quantitative analysis of EdU-positive ratio (C). (D and E) Flow cytometry results showed distribution of apoptotic cells, necrotic cells and survival cells in propofol-induced HT-22 cells either treated with 10 μM metformin or not (D). Quantitative analysis of apoptosis rate (E). (F and G) TUNEL results showed TUNEL-positive cells in propofol-induced HT-22 cells either treated with 10 μM metformin or not (F). Quantitative analysis of TUNEL-positive rate (G). (H) Protein levels of Bcl-2 and Bax in propofol-induced HT-22 cells either treated with 10 μM metformin or not (*p<0.05 compared to control group; &p<0.05, compared to propofol (100μM) group). |
Metformin Regulated Cav-1 Level
Western blot analysis uncovered that the protein level of Cav-1 dose-dependently upregulated in propofol-treated HT-22 cells (Figure 3A and B). Furthermore, metformin treatment downregulated Cav-1 level in propofol-treated HT-22 cells (Figure 3C and D). Hence, metformin was able to regulate propofol-induced Cav-1 upregulation.
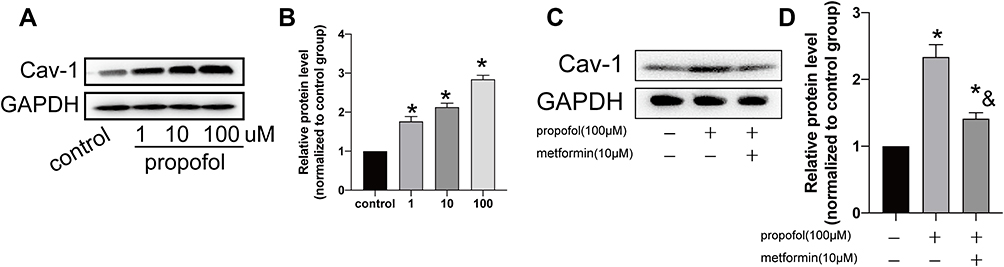 |
Figure 3 Metformin treatment regulated Cav-1 level. (A and B). Protein level of Cav-1 in HT-22 cells treated with 0, 1, 10 and 100 μM propofol, respectively (A). Grey value analysis of Cav-1 (B). (C and D). Protein level of Cav-1 in propofol-induced HT-22 cells either treated with 10 μM metformin or not (C). Grey value analysis of Cav-1 (D) (*p<0.05 compared to control group; &p<0.05, compared to propofol (100μM) group). |
Propofol-Induced Apoptosis in HT-22 Cells by Upregulating Cav-1
To further explore the function of propofol in inducing neuronal apoptosis, we constructed si-Cav-1 and pcDNA-Cav-1. Transfection efficacy of si-Cav-1 was determined in HT-22 cells by Western blot (Figure 4A). The protein level of Cav-1 was markedly upregulated after propofol treatment, which was then downregulated by transfection of si-Cav-1 (Figure 4B). CCK-8 assay showed that knockdown of Cav-1 partially reversed the declined viability in HT-22 cells treated with propofol (Figure 4C). EdU assay obtained a similar result (Figure 4D and E). The increased apoptotic rate in propofol-treated HT-22 cells was slightly reduced after transfection of si-Cav-1 (Figure 4F and G).
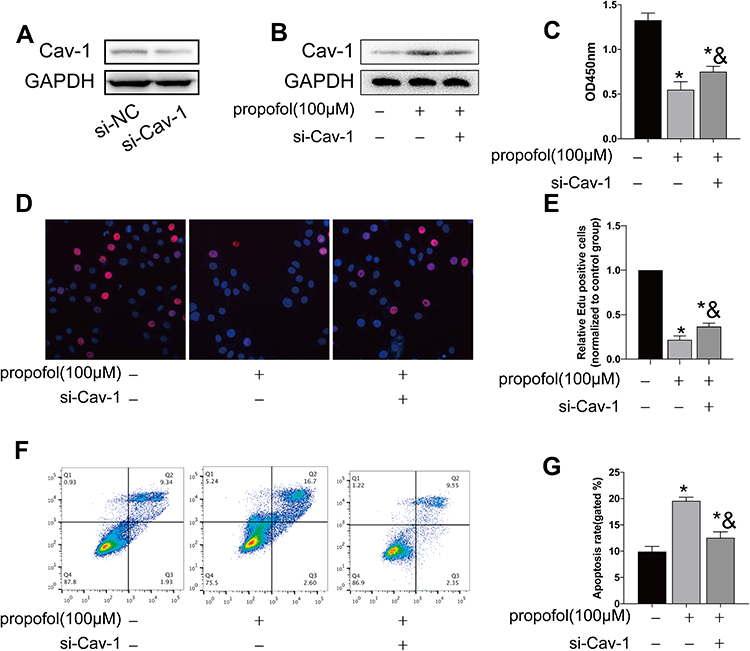 |
Figure 4 Propofol-induced apoptosis in HT-22 cells through upregulating Cav-1. (A) Transfection efficacy of si-Cav-1 in HT-22 cells. (B) Protein level of Cav-1 in propofol-induced HT-22 cells transfected with si-NC or si-Cav-1. (C) CCK-8 assay results showed viability in propofol-induced HT-22 cells transfected with si-NC or si-Cav-1. (D and E) EdU assay results showed EdU-positive cells in propofol-induced HT-22 cells transfected with si-NC or si-Cav-1 (D). Quantitative analysis of EdU-positive ratio (E). (F and G) Flow cytometry results showed distribution of apoptotic cells, necrotic cells and survival cells in propofol-induced HT-22 cells transfected with si-NC or si-Cav-1 (F). Quantitative analysis of apoptosis rate (G) (*p<0.05 compared to control group; &p<0.05, compared to propofol (100μM) group). |
Metformin Reversed Propofol-Induced Neuronal Apoptosis Through Downregulating Cav-1
Next, transfection efficacy of pcDNA-Cav-1 was verified in HT-22 cells (Figure 5A). Transfection of pcDNA-Cav-1 inhibited the upregulated protein level of Cav-1 in propofol-treated HT-22 cells (Figure 5B). Metformin reversed the declined viability and EdU-positive ratio in propofol-treated HT-22 cells, which were further reduced after overexpression of Cav-1 (Figure 5C–E). In the meantime, overexpression of Cav-1 enhanced apoptosis in propofol-treated neurons even after metformin treatment (Figure 5F and G). The above data proved that Cav-1 was responsible for protective effects of metformin on propofol-induced neurotoxicity.
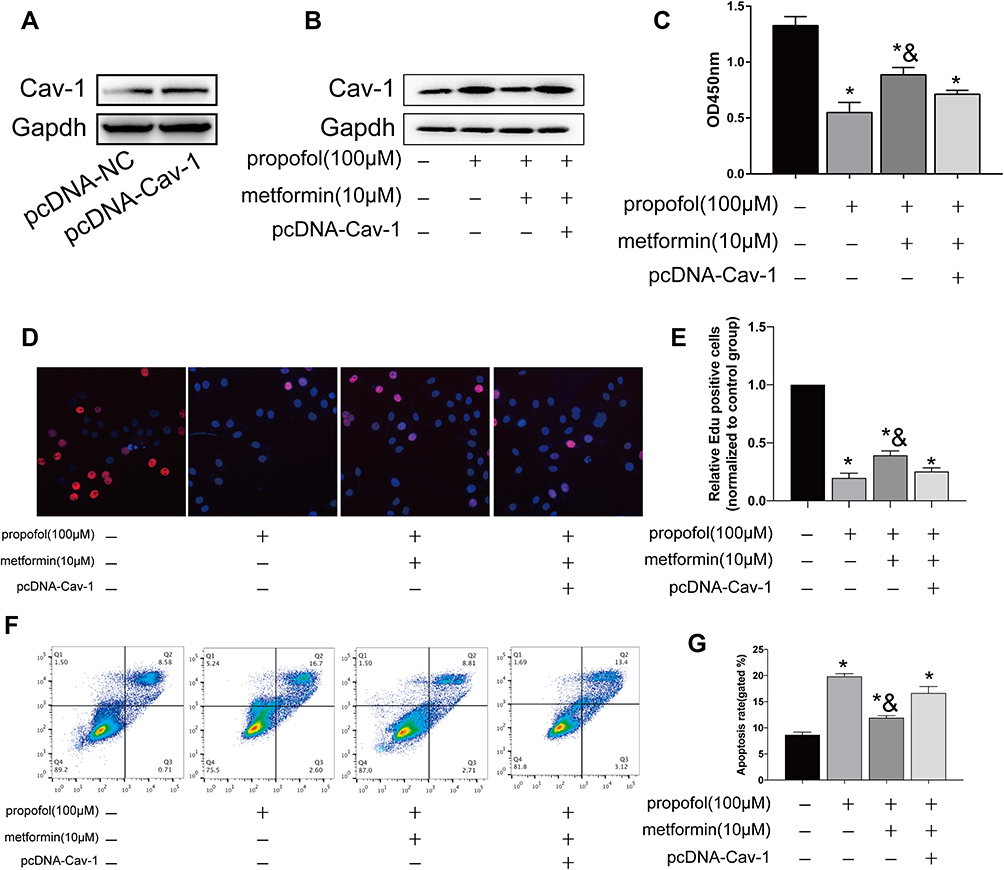 |
Figure 5 Metformin reversed propofol-induced apoptosis in HT-22 cells through downregulating Cav-1. (A) Transfection efficacy of pcDNA-Cav-1 in HT-22 cells. (B) Protein level of Cav-1 in propofol-induced HT-22 cells transfected with pcDNA-NC or pcDNA-Cav-1 with either metformin treatment or not. (C) CCK-8 assay results showed viability in propofol-induced HT-22 cells transfected with pcDNA-NC or pcDNA-Cav-1 with either metformin treatment or not. (D and E) EdU assay results showed EdU-positive cells in propofol-induced HT-22 cells transfected with pcDNA-NC or pcDNA-Cav-1 with either metformin treatment or not (D). Quantitative analysis of EdU-positive ratio (E). (F and G) Flow cytometry results showed distribution of apoptotic cells, necrotic cells and survival cells in propofol-induced HT-22 cells transfected with pcDNA-NC or pcDNA-Cav-1 with either metformin treatment or not (F). Quantitative analysis of apoptosis rate (G) (*p<0.05 compared to control group; &p<0.05, compared to propofol (100μM) group). |
Discussion
Increasing evidence have proved that propofol administration is able to induce developmental neurotoxicity, thereafter leading to long-term cognitive and learning abnormalities. Hence, the safety usage of propofol in pediatric anesthesia is well concerned.24 Fredriksson et al25 proposed that propofol administration dose-dependently aggravates the apoptotic degree in neurons of young rats, which could impair the long-term learning and memory ability of rats. In 7-day-old neonatal SD rats, propofol treatment markedly induces apoptosis in hippocampal neurons.26 Our results revealed that propofol treatment dose-dependently decreased proliferative ability and increased apoptosis in HT-22 cells.
Apoptosis is of significance in maintaining homeostasis and normal development. Nevertheless, excessive apoptosis leads to adverse biological consequences. For instance, uncontrolled apoptosis is closely linked to ischemic heart disease, AIDS and Alzheimer’s disease, Parkinson’s disease and other neurodegenerative diseases.27 Cell apoptosis has been well concerned in biology and medical researches.
Metformin has been verified as a safe and effective oral hypoglycemic agent extensively used in the treatment of diabetes.28,29 In nervous system, metformin stimulates the growth of newly formed hippocampal neurons and improves spatial learning and memory in mice.30,31 Through activating the S1P1-dependent ERK1/2 pathway, metformin protects rabbits from sevoflurane-induced neuronal apoptosis.32 Besides, metformin another AMPK activator AICAR activates AKT S473 which provides survival signal for cells by inhibiting apoptotic signaling pathways.33 In this analysis, metformin treatment protected propofol-induced proliferation inhibition and apoptosis stimulation in HT-22 cells. Therefore, we have proved the protective effect of metformin on propofol-induced neurotoxicity.
Cav-1 is a 22 kDa plasma membrane scaffold protein.34 A previous study reported that Cav-1 regulates Caspase 3-mediated apoptosis pathway by downregulating Survivin.35 Cav-1 deficiency has protective effects on both intracellular and extracellular apoptosis.36 In addition, through interacting with Ras, Cav-1 promotes the activation of the Ras/Raf/ERK pathway, an important pathway participating in cell proliferation and apoptosis.37–40 Here, Cav-1 level was upregulated in propofol-treated HT-22 cells, which was downregulated following metformin treatment. In particular, knockdown of Cav-1 suppressed propofol-induced apoptosis in HT-22 cells. Rescue experiments further confirmed that overexpression of Cav-1 abolished the anti-apoptosis function of metformin in propofol-treated HT-22 cells. Collectively, metformin protected propofol-induced neurotoxicity through Cav-1.
Conclusions
In our study, we found that propofol treatment dose-dependently decreased proliferative ability and increased apoptosis in HT-22 cells. Besides, metformin could rescue the apoptosis effect of propofol induced in HT-22. In addition, we found that propofol induced the apoptosis of HT-22 by up-regulating Cav-1 and metformin protects propofol-induced neuronal apoptosis via downregulating Cav-1.
Funding
This project was supported by the Scientific Research Project of Nantong Health and Family Planning Commission (MA2019004) and the Top Six Types of Talents’ Financial Assistance of Jiangsu Province Grant (2019-WSW-199), Nantong Science and Technology Project (JCZ18004).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Trapani G, Altomare C, Liso G, et al. Propofol in anesthesia. Mechanism of action, structure-activity relationships, and drug delivery. Curr Med Chem. 2000;7:249–271. doi:10.2174/0929867003375335
2. Kotani Y, Shimazawa M, Yoshimura S, et al. The experimental and clinical pharmacology of propofol, an anesthetic agent with neuroprotective properties. CNS Neurosci Ther. 2008;14(2):95–106. doi:10.1111/j.1527-3458.2008.00043.x
3. Bercker S, Bert B, Bittigau P, et al. Neurodegeneration in newborn rats following propofol and sevoflurane anesthesia. Neurotox Res. 2009;16(2):140–147. doi:10.1007/s12640-009-9063-8
4. Olney JW, Farber NB, Wozniak DF, et al. Environmental agents that have the potential to trigger massive apoptotic neurodegeneration in the developing brain. Environ Health Perspect. 2000;108(Suppl 3):383–388. doi:10.1289/ehp.00108s3383
5. Kahraman S, Zup SL, McCarthy MM, et al. Gabaergic mechanism of propofol toxicity in immature neurons. J Neurosurg Anesthesiol. 2008;20(4):233–240. doi:10.1097/ANA.0b013e31817ec34d
6. Cattano D, Young C, Straiko MM, et al. Subanesthetic doses of propofol induce neuroapoptosis in the infant mouse brain. Anesth Analg. 2008;106(6):1712–1714. doi:10.1213/ane.0b013e318172ba0a
7. Creeley C, Dikranian K, Dissen G, et al. Propofol-induced apoptosis of neurones and oligodendrocytes in fetal and neonatal rhesus macaque brain. Br J Anaesth. 2013;110(Suppl 1):i29–i38. doi:10.1093/bja/aet173
8. Mellon RD, Simone AF, Rappaport BA. Use of anesthetic agents in neonates and young children. Anesth Analg. 2007;104:509–520. doi:10.1097/sa.0b013e31815c1022
9. Tao T, Li CL, Yang WC, et al. Protective effects of propofol against whole cerebral ischemia/reperfusion injury in rats through the inhibition of the apoptosis-inducing factor pathway. Brain Res. 2016;1644:9–14. doi:10.1016/j.brainres.2016.05.006
10. Noh HS, Shin IW, Ha JH, et al. Propofol protects the autophagic cell death induced by the ischemia/reperfusion injury in rats. Mol Cells. 2010;30(5):455–460. doi:10.1007/s10059-010-0130-z
11. Bredesen DE. Neural apoptosis. Ann Neurol. 1995;38(6):839–851. doi:10.1002/ana.410380604
12. White E. Life, death, and the pursuit of apoptosis. Genes Dev. 1996;10(1):1–15. doi:10.1101/gad.10.1.1
13. Hacker G, Vaux DL. Apoptosis. A sticky business. Curr Biol. 1995;5(6):622–624. doi:10.1016/s0960-9822(95)00126-6
14. Smale G, Nichols NR, Brady DR, et al. Evidence for apoptotic cell death in Alzheimer’s disease. Exp Neurol. 1995;133(2):225–230. doi:10.1006/exnr.1995.1025
15. Schulte JM, Rothaus CS, Adler JN. Clinical decisions. Management of type 2 diabetes–polling results. N Engl J Med. 2014;370(1):e2. doi:10.1056/NEJMclde0801078
16. Nath N, Khan M, Paintlia MK, et al. Metformin attenuated the autoimmune disease of the central nervous system in animal models of multiple sclerosis. J Immunol. 2009;182(12):8005–8014. doi:10.4049/jimmunol.0990060
17. Patel HH, Murray F, Insel PA. Caveolae as organizers of pharmacologically relevant signal transduction molecules. Annu Rev Pharmacol Toxicol. 2008;48(1):359–391. doi:10.1146/annurev.pharmtox.48.121506.124841
18. Fridolfsson HN, Patel HH. Caveolin and caveolae in age associated cardiovascular disease. J Geriatr Cardiol. 2013;10:66–74. doi:10.3969/j.issn.1671-5411.2013.01.011
19. Kang JW, Lee SM. Impaired expression of caveolin-1 contributes to hepatic ischemia and reperfusion injury. Biochem Biophys Res Commun. 2014;450(4):1351–1357. doi:10.1016/j.bbrc.2014.06.131
20. Das M, Gherghiceanu M, Lekli I, et al. Essential role of lipid raft in ischemic preconditioning. Cell Physiol Biochem. 2008;21(4):325–334. doi:10.1159/000129391
21. Rothberg KG, Heuser JE, Donzell WC, et al. Caveolin, a protein component of caveolae membrane coats. Cell. 1992;68(4):673–682. doi:10.1016/0092-8674(92)90143-z
22. Liu Y, Fu Y, Hu X, et al. Caveolin-1 knockdown increases the therapeutic sensitivity of lung cancer to cisplatin-induced apoptosis by repressing Parkin-related mitophagy and activating the ROCK1 pathway. J Cell Physiol. 2019. doi:10.1002/jcp.29033
23. Mukhopadhyay S, Chatterjee A, Kogan D, et al. 5-Aminoimidazole-4-carboxamide-1-beta-4-ribofuranoside (AICAR) enhances the efficacy of rapamycin in human cancer cells. Cell Cycle. 2015;14(20):3331–3339. doi:10.1080/15384101.2015.1087623
24. Bosnjak ZJ, Logan S, Liu Y, et al. Recent insights into molecular mechanisms of propofol-induced developmental neurotoxicity: implications for the protective strategies. Anesth Analg. 2016;123(5):1286–1296. doi:10.1213/ANE.0000000000001544
25. Fredriksson A, Ponten E, Gordh T, et al. Neonatal exposure to a combination of n-methyl-d-aspartate and gamma-aminobutyric acid type a receptor anesthetic agents potentiates apoptotic neurodegeneration and persistent behavioral deficits. Anesthesiology. 2007;107(3):427–436. doi:10.1097/01.anes.0000278892.62305.9c
26. Han D, Jin J, Fang H, et al. Long-term action of propofol on cognitive function and hippocampal neuroapoptosis in neonatal rats. Int J Clin Exp Med. 2015;8(7):10696–10704.
27. Kannan K, Jain SK. Oxidative stress and apoptosis. Pathophysiology. 2000;7(3):153–163. doi:10.1016/s0928-4680(00)00053-5
28. Yuan X, Wei W, Bao Q, et al. Metformin inhibits glioma cells stemness and epithelial-mesenchymal transition via regulating yap activity. Biomed Pharmacother. 2018;102:263–270. doi:10.1016/j.biopha.2018.03.031
29. Fu X, Pan Y, Cao Q, et al. Metformin restores electrophysiology of small conductance calcium-activated potassium channels in the atrium of gk diabetic rats. BMC Cardiovasc Disord. 2018;18(1):63. doi:10.1186/s12872-018-0805-5
30. Ge X-H, Zhu G-J, Geng D-Q, et al. Metformin protects the brain against ischemia/reperfusion injury through pi3k/akt1/jnk3 signaling pathways in rats. Physiol Behav. 2017;170:115–123. doi:10.1016/j.physbeh.2016.12.021
31. Li J, Deng J, Sheng W, et al. Metformin attenuates Alzheimer’s disease-like neuropathology in obese, leptin-resistant mice. Pharmacol Biochem Behav. 2012;101(4):564–574. doi:10.1016/j.pbb.2012.03.002
32. Yue H, Hu B, Luo Z, et al. Metformin protects against sevoflurane-induced neuronal apoptosis through the s1p1 and erk signaling pathways. Exp Ther Med. 2019;17(2):1463–1469. doi:10.3892/etm.2018.7098
33. Zhu X, Zhang Y, Li Q, et al. beta-carotene induces apoptosis in human esophageal squamous cell carcinoma cell lines via the Cav-1/AKT/NF-kappaB signaling pathway. J Biochem Mol Toxicol. 2016;30(3):148–157. doi:10.1002/jbt.21773
34. Fujimoto T, Kogo H, Nomura R, et al. Isoforms of caveolin-1 and caveolar structure. J Cell Sci. 2000;113(Pt 19):3509–3517. doi:10.1023/A:1005629427630
35. Jin Y, Kim HP, Chi M, et al. Deletion of Caveolin-1 protects against oxidative lung injury via up-regulation of heme oxygenase-1. Am J Respir Cell Mol Biol. 2008;39(2):171–179. doi:10.1165/rcmb.2007-0323OC
36. Wang X, Wang Y, Kim HP, et al. Carbon monoxide protects against hyperoxia-induced endothelial cell apoptosis by inhibiting reactive oxygen species formation. J Biol Chem. 2007;282(3):1718–1726. doi:10.1074/jbc.M607610200
37. Kortum RL, Fernandez MR, Costanzo-Garvey DL, et al. Caveolin-1 is required for kinase suppressor of ras 1 (ksr1)-mediated extracellular signal-regulated kinase 1/2 activation, h-rasv12-induced senescence, and transformation. Mol Cell Biol. 2014;34(18):3461–3472. doi:10.1128/MCB.01633-13
38. Liu P, Ying Y, Anderson RG. Platelet-derived growth factor activates mitogen-activated protein kinase in isolated caveolae. Proc Natl Acad Sci U S A. 1997;94(25):13666–13670. doi:10.2307/43790
39. Li W, Huang R, Shetty RA, et al. Transient focal cerebral ischemia induces long-term cognitive function deficit in an experimental ischemic stroke model. Neurobiol Dis. 2013;59:18–25. doi:10.1016/j.nbd.2013.06.014
40. Lin L, Su Z, Lebedeva IV, et al. Activation of Ras/Raf protects cells from melanoma differentiation-associated gene-5-induced apoptosis. Cell Death Differ. 2006;13(11):1982–1993. doi:10.1038/sj.cdd.4401899
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.