")
Back to Journals » OncoTargets and Therapy » Volume 13
Metformin Inhibits Proliferation of Human Thyroid Cancer TPC-1 Cells by Decreasing LRP2 to Suppress the JNK Pathway
Authors He Y, Cao L, Wang L, Liu L, Huang Y, Gong X
Received 20 August 2019
Accepted for publication 13 December 2019
Published 6 January 2020 Volume 2020:13 Pages 45—50
DOI https://doi.org/10.2147/OTT.S227915
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjay Singh
Yang He,1,* Lingling Cao,2,* Li Wang,1,3 Lingping Liu,1 Ying Huang,1 Xuan Gong1
1Department of Endocrinology, Zhuhai People’s Hospital (Zhuhai Hospital Affiliated with Jinan University), Zhuhai, People’s Republic of China; 2Department of Endocrinology, Jiujiang No 1 People’s Hospital (Affiliated Jiujiang Hospital of Nanchang University), Jiujiang, People’s Republic of China; 3School of Medicine, Hangzhou Normal University, Hangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Li Wang
Department of Endocrinology, Zhuhai People’s Hospital (Zhuhai Hospital Affiliated with Jinan University), 79 Kangning Road, Xiangzhou District, Zhuhai, Guangzhou 519000, People’s Republic of China
Tel +86 13600365720
Email [email protected]
Objective: To uncover the potential effect of metformin on proliferation and apoptosis of thyroid cancer TPC-1 cell line, and the underlying mechanism.
Methods: Viability, apoptosis and LRP2 level in TPC-1 cells treated with different doses of metformin for different time points were determined. Besides, protein levels of p-JNK1 and c-Jun N-terminal kinases (JNK) in metformin-treated TPC-1 cells were detected by Western blot. Regulatory effects of LRP2 on the JNK pathway and cell viability in metformin-treated TPC-1 cells were assessed.
Results: Viability in TPC-1 cells gradually decreased with the treatment of increased doses of metformin either for 24 h or 48 h. The apoptotic rate was concentration-dependently elevated by metformin treatment. Relative levels of LRP2 and p-JNK1 were concentration-dependently downregulated by metformin treatment. In addition, overexpression of LRP2 partially abolished the inhibitory effect of metformin on the viability of TPC-1 cells.
Conclusion: Metformin treatment suppresses the proliferative ability and induces apoptosis of TPC-1 cells by downregulating LRP2 to block the JNK pathway.
Keywords: metformin, LRP2, JNK, proliferation
Introduction
Thyroid cancer is the most common malignancy in endocrine-related tumors. In recent years, thyroid cancer has been well concerned because of the rapid rise in its incidence.1 As a first-line drug applied for T2DM (type 2 diabetes mellitus), metformin contributes to stabilize blood glucose levels through inhibiting gluconeogenesis and reducing hepatic glycogenolysis.2 Interestingly, the application of metformin sharply decreases the incidences of tumor diseases, suppresses the growth of malignant cells and enhances chemotherapy-sensitivity.3 Therefore, metformin may exert a potential anti-tumor role. It is expected to be an adjuvant drug for the treatment of thyroid cancer.
LRP2 is a unique transmembrane receptor belonging to the family of low-density lipoprotein receptors (LDLR). It is highly similar to other members of the LDLR family in the structure.4 As a multi-ligand receptor, LRP2 interacts with lipoproteins, vitamin-binding proteins, hormones and enzymes based on its complement-like sequences, thus participating in transmembrane transport and re-absorption of multiple substances.5 Mutations in the LRP2 gene are closely related to plasma cholesterol and low-density lipoprotein levels.6 Therefore, LRP2 is able to regulate lipoprotein metabolism.
The c-Jun N-terminal kinases (JNK) signaling pathway is one of the critical members of the mitogen-activated protein kinase (MAPK) family.7 JNK is mainly expressed in the cytoplasm. Once it is activated, cytoplasmic JNK rapidly translocates into the nucleus and further activates transcription factors c-JUN and AP-1.8,9 As a result, abundant apoptosis-related genes are activated.10 Meanwhile, the inflammatory response is activated as well.11,12 In this paper, we mainly explored the regulatory effect of metformin on the proliferative and apoptotic changes in TPC-1 cells and the involvement of LRP2 and the JNK pathway.
Materials and Methods
Cell Culture
Human thyroid cancer TPC-1 cells were cultured in Roswell Park Memorial Institute 1640 (RPMI 1640) (HyClone, South Logan, UT, USA) containing 10% fetal bovine serum (FBS) (Gibco, Rockville, MD, USA), 100 μg/mL penicillin and 0.1 mg/mL streptomycin, in a 37°C, 5% CO2 incubator. Cultured cells displayed monolayer growth. Cell passage was conducted by 0.25% trypsin.
Cell Counting Kit-8 (CCK-8)
One hundred microliters of cell suspension (5–8×104/mL) was applied in the 96-well plate and cultured overnight. On the other day, cells were treated with different doses of metformin for different time points. Absorbance (A) at 450 nm was recorded at the appointed time points using the CCK-8 kit (Dojindo Laboratories, Kumamoto, Japan) for depicting the viability curves.
Apoptosis Determination
Cells were washed with phosphate-buffered saline (PBS) twice, centrifuged at 3000 r/min for 5 min and prepared for suspension (5×104/mL). Cell suspension was diluted in 500 μL of binding buffer, incubated with 5 μL of Annexin V in dark for 15 min, and 5 μL of Propidium Iodide (PI) at 4°C, in the dark for another 15 min. After 5-min centrifugation at 3000 r/min, the precipitant was dissolved in 300 μL of binding buffer and subjected to flow cytometry.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Extraction of total RNA in cells was performed using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and RNA was subjected to reverse transcription. The extracted cDNA was applied for PCR using SYBR Green method (TaKaRa, Tokyo, Japan). Primer sequences were as follows: LRP2: F: 5′-GATCTGTGACCTTCATTCCTGGCCTGATC-3′, R: 5′-GCCATGACACCTGTAGATGTGGTGCTGAATAATTGGTTAA-3′; Glyceraldehyde 3-phosphate dehydrogenase (GAPDH): F: 5′-ACTGCCACCCAGAAGACT-3′, R: 5ʹ-GCTCAGTGTAGCCCAGGAT-3′.
Western Blot
Total protein was extracted from cells using radioimmunoprecipitation assay (RIPA) lysis butter and quantified by bicinchoninic acid (BCA) method (Beyotime, Shanghai, China). Protein sample was loaded for electrophoresis and transferred on polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA). After blocking non-specific antigens in 5% skim milk for 2 hrs, membranes were subjected to incubation with primary and secondary antibodies. Bands were exposed by chemiluminescence (ECL) reagents and analyzed by Image Software (NIH, Bethesda, MD, USA).
Transfection
The cells were detached in 0.25% trypsin and prepared for suspension (5×104/mL). Fifty pmol transfection vector and 2 μL of LipofectamineTM 3000 (Invitrogen, Carlsbad, CA, USA) were, respectively, diluted in 100 μL of Opti-MEM® I. After 5-min maintenance at room temperature, they were mixed and let stand for another 20 min. Subsequently, the mixture was applied to the suspension. Transfected cells for 48 h were harvested for functional experiments.
Statistical Analysis
Statistical Product and Service Solutions (SPSS) 20.0 (IBM, Armonk, NY, USA) was used for data analyses. Data were expressed as mean ± standard deviation. The differences among groups were statistically evaluated using Student’s t-test when comparing only two groups or one-way analysis of variance (ANOVA) when comparing more than two groups. P<0.05 was considered as statistically significant.
Results
Metformin Suppressed Proliferation and Induced Apoptosis in TPC-1 Cells
TPC-1 cells were treated with 0, 1.25, 2.5, 5.0, 10.0 or 20.0 mM metformin for 24 or 48 h, respectively. As CCK-8 assay results revealed, the viability concentration-dependently decreased either after 24 or 48 h treatment (Figure 1A). In addition, the apoptotic rate in TPC-1 cells treated with 0, 10.0 or 20.0 mM metformin for 48 h gradually increased (Figure 1B). The above data supported the abilities of metformin to attenuate viability and induce apoptosis in TPC-1 cells.
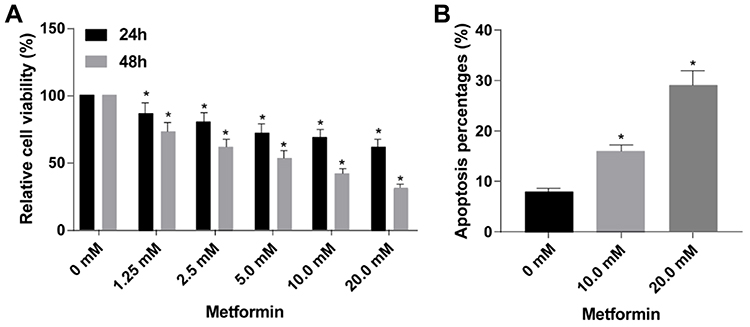 |
Figure 1 Metformin suppressed proliferation and induced apoptosis in TPC-1 cells. (A) Viability in TPC-1 cells treated with 0, 1.25, 2.5, 5.0, 10.0 or 20.0 mM metformin for 24 or 48 h, respectively. (B) Apoptotic rate in TPC-1 cells treated with 0, 10.0 or 20.0 mM metformin for 48 h (n=3, *p<0.05, significantly different from the 0 mM metformin group). |
Metformin Application Downregulated LRP2 in TPC-1 Cells
After treatment of 0, 10.0 or 20.0 mM metformin for 48 h, both protein and mRNA levels of LRP2 were downregulated, manifesting a dose–response relationship (Figure 2A and B).
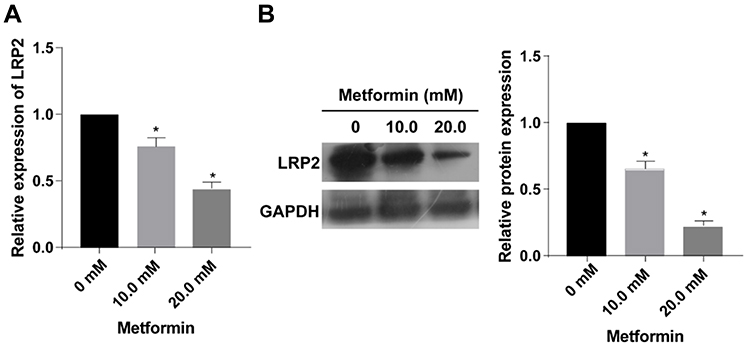 |
Figure 2 Metformin application downregulated LRP2 in TPC-1 cells. (A, B) The mRNA (A) and protein (B) levels of LRP2 in TPC-1 cells treated with 0, 10.0 or 20.0 mM metformin for 48 h (n=3, *p<0.05, significantly different from the 0 mM metformin group). |
Metformin Application Inhibited the Activation of JNK Pathway in TPC-1 Cells
To clarify the involvement of the JNK pathway in metformin-mediated progression of thyroid cancer, protein levels of p-JNK1 and JNK1 were determined by Western blot. Protein level of p-JNK1 in TPC-1 cells was concentration-dependently downregulated by metformin treatment (Figure 3). However, the protein level of JNK1 remained unchangeable.
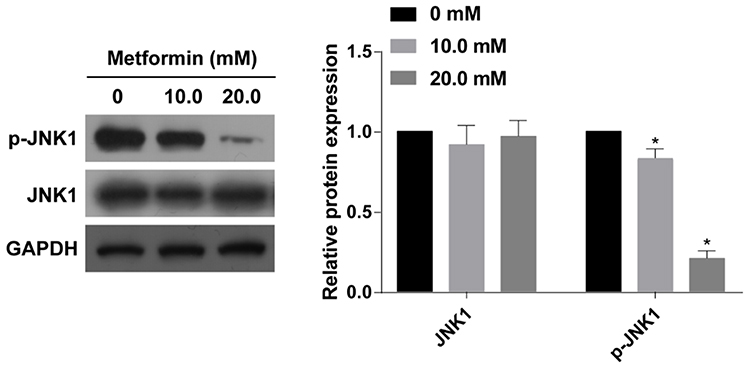 |
Figure 3 Metformin application inhibited the activation of JNK pathway in TPC-1 cells. Protein levels of p-JNK1 and JNK1 in TPC-1 cells treated with 0, 10.0 or 20.0 mM metformin for 48 h (n=3, *p<0.05, significantly different from the 0 mM metformin group). |
It is speculated that LRP2 was involved in the inactivation of JNK pathway. Subsequently, pcDNA-LRP2 was constructed. Transfection of pcDNA-LRP2 effectively downregulated both mRNA and protein levels of LRP2 in TPC-1 cells, showing a pronounced transfection efficacy (Figure 4A and B).
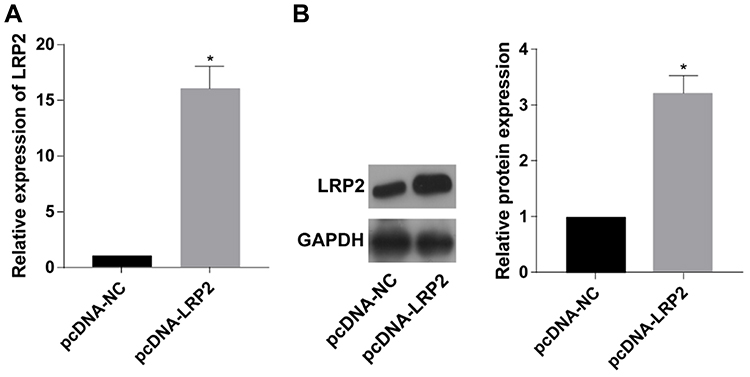 |
Figure 4 Transfection efficacy of pcDNA-LRP2. (A, B) The mRNA (A) and protein (B) levels of LRP2 in TPC-1 cells transfected with pcDNA-NC or pcDNA-LRP2 (n=3, *p<0.05, significantly different from the control group). |
Overexpression of LRP2 Abolished the Inhibitory Effects of Metformin on Cell Viability and Activation of the JNK Pathway
Transfection of pcDNA-LRP2 markedly reversed the inhibitory effect of metformin on protein levels of LRP2 and p-JNK1 in TPC-1 cells (Figure 5A). Moreover, decreased viability in metformin-treated TPC-1 cells was partially reversed by overexpression of LRP2 (Figure 5B). It is verified that LRP2 was responsible for metformin-induced viability suppression and JNK pathway inactivation.
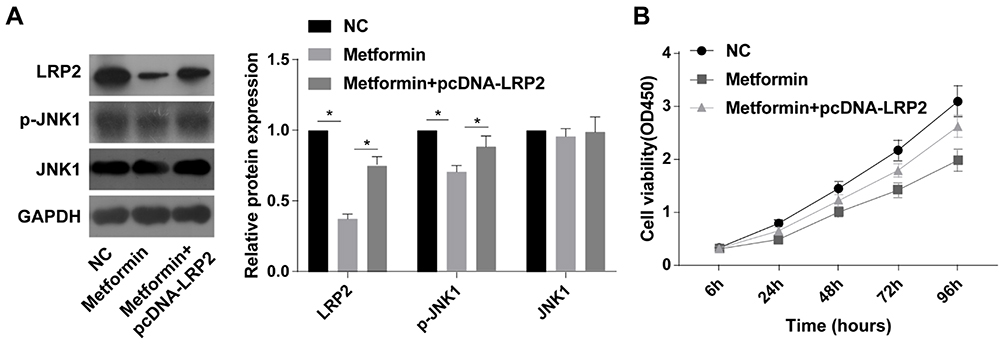 |
Figure 5 Overexpression of LRP2 abolished the inhibitory effects of metformin on cell viability and activation of the JNK pathway. TPC-1 cells were treated with NC, metformin or metformin+pcDNA-LRP2, respectively. (A) Protein levels of LRP2 and p-JNK1decreased significantly compared to the NC group, and overexpression of LRP2 reversed that. (B) Viability at 6, 24, 48, 72 and 96 h. Cell viability was decreased significantly compared to the NC group. Overexpression of LRP2 reversed this effect (n=3, *p<0.05; significantly different from the NC group or overexpression of LRP2 plus metformin group). |
Discussion
A retrospective epidemiological study conducted by Evans et al13 in 2005 pointed out that the application of metformin markedly reduces the incidences of malignant tumors in patients with diabetes. Later, the tumor-suppressor effect of metformin has been well concerned. A relevant study demonstrated that metformin treatment suppresses tumor growth, elevates the complete remission rate and prolongs the disease-free survival in patients with thyroid cancer.14 Cho et al15 suggested that the application of different doses of metformin reduces the tumor volume and suppresses tumor cell growth in tumor-bearing nude mice. In a recent study, metformin like drug AICAR that affects cancer cell proliferation which in turn effective to create cell death in cancer cells via activating AMPK pathway.16 In this paper, metformin treatment markedly inhibited proliferative ability and induced apoptosis in TPC-1 cells. Furthermore, LRP2 level was downregulated and the JNK pathway was activated by metformin application.
The previous study has shown that LRP2 is mainly involved in regulations on lipoprotein transporter activity, low-density lipoprotein receptor activity, lipid metabolic process, lipoprotein transport, coronary vasculature development, aorta development, cell proliferation, ventricular septum development, clathrin-coated vesicle membrane, lysosome, etc. It is of significance in the etiology of acute myocardial infarction.17,18 The JNK pathway is extensively involved in cellular behaviors, and its dysregulation is considered to be linked to disease progression.19–21 In MGC-803 cells (gastric cancer cells), JNK induces apoptosis through the mitochondrial pathway, triggering cytochrome release into the cytoplasm to initiate the apoptotic pathway.21,22 Besides, metformin is able to activate the AMPK, mTOR/6SK, p-ERK and p70S6K/pS6 pathways.14,23 It is capable of suppressing mitosis and proliferation of thyroid cancer stem cells and induces apoptosis.24 Our findings illustrated that metformin treatment inactivated the JNK pathway in TPC-1 cells, which could be abolished by overexpression of LRP2.
Collectively, metformin may be beneficial to inhibit the progression of thyroid cancer by downregulating LRP2 to inactivate the JNK pathway. Our findings provide new directions for the clinical treatment of thyroid cancer.
Conclusions
Metformin treatment suppresses the proliferative ability and induces apoptosis of TPC-1 cells by downregulating LRP2 to block the JNK pathway.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Xu Y, Zheng X, Qiu Y, Jia W, Wang J, Yin S. Distinct metabolomic profiles of papillary thyroid carcinoma and benign thyroid adenoma. J Proteome Res. 2015;14(8):3315–3321. doi:10.1021/acs.jproteome.5b00351
2. Zhen Z, Dong F, Shen H, Wang QG, Yang L, Hu J. MiR-524 inhibits cell proliferation and induces cell apoptosis in thyroid cancer via targeting SPAG9. Eur Rev Med Pharmacol Sci. 2018;22(12):3812–3818. doi:10.26355/eurrev_201806_15265
3. Chung HH, Moon JS, Yoon JS, Lee HW, Won KC. The relationship between metformin and cancer in patients with type 2 diabetes. Diabetes Metab J. 2013;37(2):125–131. doi:10.4093/dmj.2013.37.2.125
4. Bu G. Apolipoprotein E and its receptors in alzheimer’s disease: pathways, pathogenesis and therapy. Nat Rev Neurosci. 2009;10(5):333–344. doi:10.1038/nrn2620
5. Kozyraki R, Gofflot F. Multiligand endocytosis and congenital defects: roles of cubilin, megalin and amnionless. Curr Pharm Des. 2007;13(29):3038–3046. doi:10.2174/138161207782110507
6. Mii A, Nakajima T, Fujita Y, et al. Genetic association of low-density lipoprotein receptor-related protein 2 (LRP2) with plasma lipid levels. J Atheroscler Thromb. 2007;14(6):310–316. doi:10.5551/jat.e494
7. Liu H, Zhou M. Antitumor effect of quercetin on Y79 retinoblastoma cells via activation of JNK and p38 MAPK pathways. BMC Complement Altern Med. 2017;17(1):531. doi:10.1186/s12906-017-2023-6
8. Fung TS, Liu DX. Activation of the c-Jun NH2-terminal kinase pathway by coronavirus infectious bronchitis virus promotes apoptosis independently of c-Jun. Cell Death Dis. 2017;8(12):3215. doi:10.1038/s41419-017-0053-0
9. Jo AR, Han HS, Seo S, et al. Inhibitory effect of moschamine isolated from carthamus tinctorius on LPS-induced inflammatory mediators via AP-1 and STAT1/3 inactivation in RAW 264.7 macrophages. Bioorg Med Chem Lett. 2017;27(23):5245–5251. doi:10.1016/j.bmcl.2017.10.035
10. Dhanasekaran DN, Reddy EP. JNK-signaling: a multiplexing hub in programmed cell death. Genes Cancer. 2017;8(9–10):682–694. doi:10.18632/genesandcancer.155
11. Lin CY, Wang WH, Chen SH, et al. Lipopolysaccharide-induced nitric oxide, prostaglandin E2, and cytokine production of mouse and human macrophages are suppressed by pheophytin-b. Int J Mol Sci. 2017;18:12. doi:10.3390/ijms18122637
12. Han Q, Bing W, Di Y, et al. Kinsenoside screening with a microfluidic chip attenuates gouty arthritis through inactivating NF-kappaB signaling in macrophages and protecting endothelial cells. Cell Death Dis. 2016;7(9):e2350. doi:10.1038/cddis.2016.255
13. Evans JM, Donnelly LA, Emslie-Smith AM, Alessi DR, Morris AD. Metformin and reduced risk of cancer in diabetic patients. BMJ. 2005;330(7503):1304–1305. doi:10.1136/bmj.38415.708634.F7
14. Klubo-Gwiezdzinska J, Costello JJ, Patel A, et al. Treatment with metformin is associated with higher remission rate in diabetic patients with thyroid cancer. J Clin Endocrinol Metab. 2013;98(8):3269–3279. doi:10.1210/jc.2012-3799
15. Cho SW, Yi KH, Han SK, et al. Therapeutic potential of metformin in papillary thyroid cancer in vitro and in vivo. Mol Cell Endocrinol. 2014;393(1–2):24–29. doi:10.1016/j.mce.2014.05.021
16. Mukhopadhyay S, Chatterjee A, Kogan D, et al. 5-aminoimidazole-4-carboxamide-1-β-4-ribofuranoside (AICAR) enhances the efficacy of rapamycin in human cancer cells. Cell Cycle. 2015;14(20):3331–3339. doi:10.1080/15384101.2015.1087623
17. Cases O, Obry A, Ben-Yacoub S, et al. Impaired vitreous composition and retinal pigment epithelium function in the FoxG1:: LRP2myopic mice. Biochim Biophys Acta Mol Basis Dis. 2017;1863(6):1242–1254. doi:10.1016/j.bbadis.2017.03.022
18. Zhang B, Wang LL, Ren RJ, et al. MicroRNA-146a represses LRP2 translation and leads to cell apoptosis in alzheimer’s disease. FEBS Lett. 2016;590(14):2190–2200. doi:10.1002/1873-3468.12229
19. Chowdhury A, Lu J, Zhang R, et al. Mangiferin ameliorates acetaminophen-induced hepatotoxicity through APAP-Cys and JNK modulation. Biomed Pharmacother. 2019;117:109097. doi:10.1016/j.biopha.2019.109097
20. Valvassori SS, Gava FF, Dal-Pont GC, et al. Effects of lithium and valproate on ERK/JNK signaling pathway in an animal model of mania induced by amphetamine. Heliyon. 2019;5(5):e1541. doi:10.1016/j.heliyon.2019.e01541
21. Shen HM, Liu ZG. JNK signaling pathway is a key modulator in cell death mediated by reactive oxygen and nitrogen species. Free Radic Biol Med. 2006;40(6):928–939. doi:10.1016/j.freeradbiomed.2005.10.056
22. Takagi Y, Nozaki K, Sugino T, Hattori I, Hashimoto N. Phosphorylation of c-Jun NH(2)-terminal kinase and p38 mitogen-activated protein kinase after transient forebrain ischemia in mice. Neurosci Lett. 2000;294(2):117–120. doi:10.1016/s0304-3940(00)01552-4
23. Klubo-Gwiezdzinska J, Jensen K, Costello J, et al. Metformin inhibits growth and decreases resistance to anoikis in medullary thyroid cancer cells. Endocr Relat Cancer. 2012;19(3):447–456. doi:10.1530/ERC-12-0046
24. Chen G, Xu S, Renko K, Derwahl M. Metformin inhibits growth of thyroid carcinoma cells, suppresses self-renewal of derived cancer stem cells, and potentiates the effect of chemotherapeutic agents. J Clin Endocrinol Metab. 2012;97(4):E510–E520. doi:10.1210/jc.2011-1754
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.