")
Back to Journals » OncoTargets and Therapy » Volume 12
Metabolomics: a promising diagnostic and therapeutic implement for breast cancer
Authors Chen Z, Li Z , Li H, Jiang Y
Received 14 May 2019
Accepted for publication 22 July 2019
Published 21 August 2019 Volume 2019:12 Pages 6797—6811
DOI https://doi.org/10.2147/OTT.S215628
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr William C. Cho
Zhanghan Chen,1 Zehuan Li,1 Haoran Li,2 Ying Jiang1
1Department of General Surgery, Zhongshan Hospital, Fudan University, Shanghai 200032, People’s Republic of China; 2Department of Thoracic Surgery, Zhongshan Hospital, Fudan University, Shanghai 200032, People’s Republic of China
Correspondence: Ying Jiang
Department of General Surgery, Zhongshan Hospital, Fudan University, 180 Fenglin Road, Xuhui District, Shanghai 200032, People’s Republic of China
Tel +86 216 404 1990
Fax +86 216 403 8472
Email [email protected]
Abstract: Breast cancer (BC) is the most commonly diagnosed cancer among women and the leading cause of cancer death. Despite the advent of numerous diagnosis and treatment methods in recent years, this heterogeneous disease still presents great challenges in early diagnosis, curative treatments and prognosis monitoring. Thus, finding promising early diagnostic biomarkers and therapeutic targets and approaches is meaningful. Metabolomics, which focuses on the analysis of metabolites that change during metabolism, can reveal even a subtle abnormal change in an individual. In recent decades, the exploration of cancer-related metabolomics has increased. Metabolites can be promising biomarkers for the screening, response evaluation and prognosis of BC. In this review, we summarized the workflow of metabolomics, described metabolite signatures based on molecular subtype as well as reclassification and then discussed the application of metabolomics in the early diagnosis, monitoring and prognosis of BC to offer new insights for clinicians in breast cancer diagnosis and treatment.
Keywords: diagnosis, biomarkers, treatment, metabolites
Background
Metabolomics (metabonomics), different from genomics and proteomics, focuses on metabolites with a molecular weight <1500 Da, such as glycolipids and polysaccharides, during metabolism at a precise time and condition to reflect the endogenous responses of metabolites to internal and external factors.1 Roger Williams described the concept of “metabolic profile” for the first time in the late 1940s and proposed that individuals had their own “metabolic profile” characterized by their body fluids.2 In the 1960s and 1970s, scientists started to utilize metabolites as biomarkers to illustrate the relationship between metabolites and diseases with advancing technology.3,4 Recently, metabolomics has provided a new avenue in cancer research because the chemical entity can reflect the cellular state and microenvironment of cancer.
Metabolomics is a more convenient, less damaged and easier measuring choice than traditional detection methods that may be invasive or radiant in detecting diseases.5 In addition, metabolomics samples can be obtained in a variety of ways, and the types of metabolomics are also extensive, such as blood, urine, saliva and tissue.6–9 Metabolites extracted from different clinical samples may reveal abnormal situations or diseases in the body.
Being the most commonly diagnosed cancer among women, breast cancer (BC) is the leading cause of cancer death, followed by colorectal cancer and lung cancer.10 Moreover, BC remains difficult to completely cure due to its heterogeneity. The diagnosis of BC with existing means of inspection is still not sufficiently timely; therefore, patients usually miss the optimal opportunity for treatment, leading to poor prognosis. The expression of many specific metabolites might be changed during the occurrence and development of cancers.11 Thus, the detection and monitoring of cancer would be possible with promising metabolic biomarkers.12,13 This review summarizes the current advances of BC in metabolomics and provides a reference for the diagnosis, therapy and monitoring of BC in clinical practices.
The workflow of metabolomics
Metabolomic analysis platforms
In general, the procedure for metabolomics can be concluded in sample preparation, component separation, detection and identification, data analysis and interpretation.5,14 According to the objective and purpose of the research, metabolomics strategies can be divided into four levels:15 (1) Metabolite target analysis focuses exclusively on one or several components. This strategy is mainly used for screening purposes and for analyses that need extreme sensitivity. (2) Metabolic profiling is a quantitative analysis focused on a smaller number of pre-defined metabolites. (3) Metabolic fingerprinting is a high-throughput and rapid qualitative strategy that analyses samples without isolation. Metabolic fingerprinting is suitable for diagnostic usage and clinical routines. and (4) Metabolomics is a qualitative and quantitative strategy that determines all metabolic components in a specific biological sample under defined conditions.15 Briefly, metabolomics strategies can be divided into targeted and untargeted approaches, and the main distinction is that the targeted approach is focused on specific metabolites or pathways.5
The representative detection methods of metabolomics include mass spectrometry (MS), which is used to identify and quantify metabolites following the optional separation by various methods, such as gas chromatography (GC), liquid chromatography (LC) or capillary electrophoresis (CE). Nuclear magnetic resonance (NMR) spectroscopy is a non-destructive and non-selective detection technique that can work independently.16,17 Additionally, there are many derivative technologies that perform better under a specific condition, such as ultra-performance liquid chromatography (UPLC)–MS, which is suitable for pharmaceutical research.18 Choosing a suitable platform is required for different purposes and sample types. MS is characterized by extremely high sensitivity that can detect low concentration metabolites. The liquid chromatography and mass spectrometry (LC/MS) combination has the advantage of analysing and determining non-volatile, polar and thermolabile compounds, such as proteins, peptides and polymers, while the gas chromatography and mass spectrometry (GC/MS) combination is mainly used for more volatile and less polar substances.19,20 The combination of capillary electrophoresis and mass spectrometry is good for analysing highly polar and charged metabolites in complex biological samples.19 NMR, as mentioned above, is a non-destructive and unbiased detection technique for dynamic detection in real-time.17 However, the low sensitivity of NMR makes it hard to detect low-abundance metabolites and the signal overlap hampers the identification and qualification of metabolites.21
Data processing and identification
Data processing and identification is the most difficult and complex step in metabolomics studies. Before analysis, the data acquired from instruments should undergo pre-processing, normalization and pre-treatment to eliminate unwanted variation and allow quantitative comparison.22–24 Simple univariate analyses cannot meet the requirements of the complex interplay between metabolites, thus multivariate statistical methods are necessary.25 The multivariate statistical methods can be broadly divided into unsupervised methods for which the analysis has no predefined values or groups and supervised methods for which the analysis of the data was based on pre-established groups or variables.20,26 Unsupervised methods include principal component analysis (PCA)27 and hierarchical cluster analysis (HCA).28 Supervised methods include partial least squares (PLS), soft independent modelling of class analogy (SIMCA), partial least squares discriminant analysis (PLS-DA), orthogonal PLS-DA (OPLS-DA) and random forest regression analysis.27 Generally, an unsupervised method is first used to classify unknown samples and find the different components. Then, based on the results, a supervised method is introduced to screen possible biomarkers and finally interpret the results to meaningful parameters.26
Factors influencing metabolomics raw data
Since the whole body participates in the metabolic process and each cell releases its own metabolites, many factors can influence the results of metabolomics raw data29 such as, gender,30 drug usage3 and dietary habits.31 African-American or Caucasian women have shown different metabolic profiles of triple-negative breast cancer (TNBC).32 These differences suggest that racial differences may alter the metabolites of BC. The screening of salivary volatiles of putative BC revealed that Portuguese samples and Indian samples possessed discrepancies.9 Moreover, the metabolite patterns of the pre-menopausal period differed from those of the postmenopausal period.33 Aside from race and age, the outcome of metabolite screening was also influenced by the sampling process. For example, the time in vacuum storage before freezing had a minimal effect on the exploration of histological, transcriptomic and proteomic profiles but markedly affected metabolite profiles. Additionally, the sample freezing time also influenced the metabolite outcome.34,35
Molecular signatures of BC based on metabolomics
The metabolic signatures of basic BC subtypes
BC can be classified into four subtypes, including Luminal A, Luminal B, HER-2-enriched and triple-negative breast cancer, based on the status of the hormone receptors, such as oestrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER-2), and the expression of ki-67.36,37 In addition, four basic molecular subtypes can be identified by metabolomics. Eight metabolites, including carnitine, lysophosphatidylcholine (20:4), proline, alanine, lysophosphatidylcholine (16:1), glycochenodeoxycholic acid, valine and 2-octenedioic acid, were selected as a panel to identify HER2-negative or HER2-positive patients and ER-positive or ER-negative patients. Compared to HER2-negative patients, HER2-positive patients showed elevated levels of carnitine, lysophosphatidylcholine (20:4), proline, valine, and 2-octenedioic acid. The ER-positive patients, compared with ER-negative patients, showed an elevated level of glycochenodeoxycholic acid and a decreased level of alanine, lysophosphatidylcholine (16:1), valine and 2-octenedioic acid.38 Further, ER-positive subtype revealed an elevated level of glutamine biosynthesis and glutamine secretion compared with those of the ER-negative subtype, and the ER-negative subtype showed remarkably higher serine metabolism and glutamine utilization than those of the ER-positive subtype.39 In addition, higher beta-alanine accumulation can be found in ER-negative BC, while 4-aminobutyrate aminotransferase expression was decreased in the ER-positive subtype.40 In addition, another group reported that the luminal B subgroup preferentially used fatty acids as an energy substrate, while HER-2 and basal-like subgroups revealed obvious alterations in glucose or glutamine metabolism.41 The epithelial mesenchymal transition (EMT) is an important phenotype that indicates a more aggressive behaviour of cancer cells.42 compared the less aggressive breast cancer cell line MCF-7 with the most aggressive breast cancer cell line MDA-MB-231 by using MALDI-TOF/TOF mass spectrometry and NMR and found differences in the lipidomic profiles between these cells. MDA-MB-231 presented lower de novo lipid synthesis and higher triacylglycerol (TAG) metabolism than did MCF7 cells, which could be a signature to differentiate basal-like breast cancer from luminal breast cancer.42 Determining the degree of malignancy based on metabolite patterns may provide clinicians with more accurate personalized treatments (Table 1).
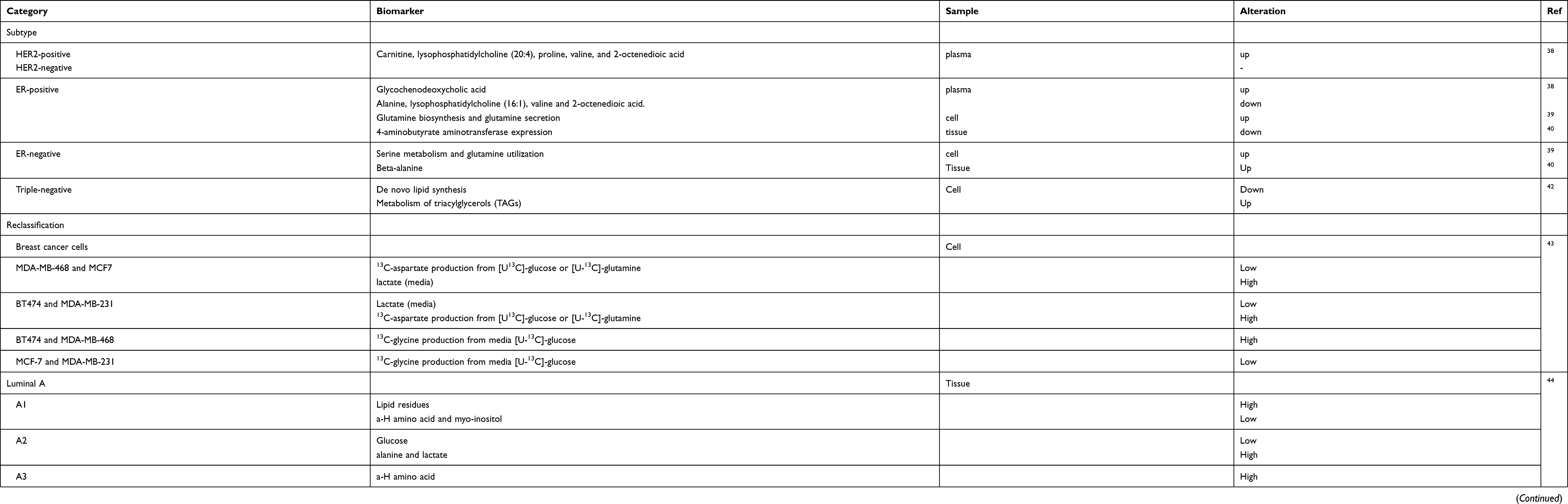 |
Table 1 Molecular subtypes of BC based on metabolomics |
Reclassification of BC based on metabolomics
The clinical response of treatment in BC can vary, even in the same subtype, owing to the cancer heterogeneity; thus, a more specific classification is needed.41 A study introduced stable isotopic 13C substrates to track the metabolism pathway of glucose, nucleotides and amino acids in BT474 and MCF-7 cell lines and MDA-MB-468 and MDA-MB-231 cell lines. The metabolites showed significant differences in the same subtype and similarity across the subtypes. Cell lines with the same hormone receptor status have different energy utilization patterns.43 Borgan et al analysed tissue samples combining gene expression microarrays and high-resolution magic angle spinning magnetic resonance spectroscopy (HR MAS MRS). These authors found that luminal A can be separated into three subgroups, A1-A3. The results also showed significant differences in glucose signals, a-H amino acid signals, alanine signals, lipid residue signals, and signals from myo-inositol among these three subtypes. Additionally, these subgroups showed great differences among biological processes and molecular functions by a GO terms enrichment analysis.44 BC molecular subtypes classified by metabolomics can help make personalized treatment decisions. In another study, the Luminal A subtype was classified into three distinct metabolic clusters (1–3) based on their metabolite patterns, which were mainly different in phospholipid metabolism, glycolytic activity and glutaminolysis. Different levels of metabolites in these sub-clusters were associated with different tumour biological behaviours and the prognosis of BC.45 Reclassifying the molecular subtypes based on metabolite patterns may facilitate clinicians to make better clinical decisions (Table 1).
The application of metabolomics in the clinical practice of BC
Metabolomics serves as a risk filter for BC
The metabolite levels may serve as an indicator for people who were likely candidates for cancer to take prevention measures.46 The results showed that women who have higher levels of valine, lysine, arginine, glutamine, creatine, creatinine and glucose or lower levels of lipoproteins, lipids, glycoproteins, acetone, glycerol-derived compounds and unsaturated lipids were at higher risk of developing BC.47 Body weight has previously been regarded as an independent risk factor for postmenopausal BC. A previous study reported that weight loss and energy restriction reduce BC risk.8 The relationship of body weight and BC was revealed by metabolites. The results showed that postmenopausal BMI is associated with increased circulating levels of parent oestrogens and the decreased methylation of catechol oestrogen metabolites, which were correlated with higher BC risk. This finding illustrated that endogenous oestrogens/oestrogen metabolites probably serve as indicators to evaluate the BMI-associated risk of BC.48 Another group found that 16a-hydroxy-DHEA-3-sulfate and 3-methylglutarylcarnitine (BMI-associated risk metabolites) were independently associated with invasive BC.49 Diet was another important factor that can influence the incidence of various cancers, including BC. Greek-Cypriot women have a decreased risk of BC because of their Mediterranean dietary habit.31 Playdon et al explored the circulating diet-related metabolites related to BC and found that the concentration of metabolites was associated with alcohol. Simultaneously, these authors found that animal fats associated with ER-positive BC and metabolites related to vitamin E can be protective factors.50 This finding offered a potential pre-diagnostic method of diet-related exposure that contributed to BC risk.
Metabolomics serves as a diagnostic detector of BC
The change in metabolites could be detected in an earlier stage, even if the abnormity was subtle. Even a subtle abnormity in the body can be increased through transcriptional, translational and molecular interactions, and metabolites are the terminal products of biochemical reactions throughout the body. Therefore, metabolomics can be a promising early biomarker of BC.51
Blood-derived sample
A dried blood spot-based direct infusion MS metabolomic analysis was used to find a quick way to diagnose BC, which targets 23 amino acids and 26 acylcarnitines. Twenty-one BC-related metabolites were differently changed in the blood, and a diagnostic panel was generated (piperamide, asparagine, proline, tetradecenoylcarnitine/palmitoylcarnitine, phenylalanine/tyrosine, and glycine/alanine). This panel showed a sensitivity of 92.2% and a specificity of 84.4% when used to test another set of BC and non-BC samples.52 The level of amino acid metabolites was also altered in the blood.53 In addition, the plasma samples of primary BC patients show lower levels of many amino acids.54 The serum amino acid and organic acid profiles of BC patients, benign patients and controls also showed significant differences. The most meaningful finding is the elevated levels of taurine and glutamic acid and the decreased levels of taurine and glutamic acid between BC patients and controls.55 Tryptophan exhibited a significantly higher level in in the serum samples of malignant tumours than in those of benign tumours.56 A differential analysis of the metabolomics profiles among invasive ductal carcinoma (IDC) patients in control and benign groups revealed that the combination of tryptophan, tyrosine, and creatine may serve as a convenient filter for both control and benign IDC subjects.57 Another group reported the 9-cisRA showed a prominent decrease from normal controls with localized BC to metastatic BC based on the analysis of 46 clinical serum and urine samples.58 Oestrogen was regarded as a dual effector of BC.59 Lysophosphatidylcholines and phosphatidylcholines were found to be significantly elevated in the oestrogen-induced group after comparing the serum metabolites profiles of an oestrogen-induced BC model with those of the control group through ultra-high performance liquid chromatography (UHPLC)-time-of-flight tandem mass spectrometer (QTOF-MS/MS).60 In summary, the metabolite patterns in blood might be a promising method to differentiate BC patients from putative healthy individuals.
Urine-derived sample
A group analysed the urine samples of healthy controls and BC patients among Hispanic women based on LC-MS or GC-MS and found that dimethylheptanoylcarnitine and succinic acid were significantly changed. Then, the combination of these two metabolites was evaluated through receiver operating characteristic analysis with a sensitivity of 93% and a specificity of 86%.61 Nam et al combined tissue transcriptomics and urine metabolomics and selected nine metabolic pathways based on the breast cancer gene expression signatures. Finally, four metabolic biomarkers (homovanillate, 4-hydroxyphenylacetate, 5-hydroxyindoleacetate and urea) were found to be changed between normal and abnormal samples.62 These studies demonstrated that the analysis of the metabolites in urine was a promising biomarker at least for an early diagnosis of BC.
Salivary-derived sample
Saliva is a convenient and noninvasive sample. Salivary metabolite analysis and identification specific to BC showed cancer-specific changes in salivary metabolites but still need further confirmation.63 Another group, lysophosphatidylcholine (18:1), lysophosphatidylcholine (22:6) and monoacylglycerol (0:0/14:0/0:0), was upregulated by analysing saliva samples from BC patients and healthy controls, and their AUC values were 0.920, 0.920 and 0.929, respectively.64 Takayama et al established a diagnostic method to detect BC patients based on the ratios of polyamines and their acetylated forms in saliva by a UPLC-MS/MS analysis. In this research, eight polyamines or their acetylated forms (cadaverine, spermine, spermidine, N1-acetyl-spermine, N1-acetyl-spermidine and N8-acetyl-spermidine) were illustrated to be strongly correlated with cancer patients.65 In short, salivary samples can also distinguish BC patients from healthy individuals by metabolite analysis.
Other available samples
Nipple aspirate fluids (NAF) can reveal the total ductal/alveolar breast tree condition.66 A previous group introduced an untargeted metabolomics analysis of breast ductal fluid from both affected breasts and unaffected unilateral breast cancer based on ultra-performance liquid chromatography coupled with a quadrupole time-of-flight (UPLC-QTOF) mass spectrometer. The LASSO regression selected 21 metabolites after adjusting for race, menopausal status, smoking, grade and TNM stage, and this panel had an AUC of 0.956.67 Breath volatile organic compounds can also be a potential sample for the early diagnosis of breast cancer.68 The metabolites, including 2-pentanone, 2-heptanone, 3-methyl-3-buten-1-ol, ethyl acetate, ethyl propanoate and 2-methyl butanoate, can only be detected in BC cell lines by analysing volatile organic compounds from the headspace of BC cells and normal human mammary epithelial cells.69 Triple negative breast cancer (TNBC) has the same histological characteristics as germline BRCA1-associated tumours, while widely known mutations in BRCA-associated tumours. Metabolomics analysis performed on triple-negative hereditary breast cancer patients with BRCA1 mutations showed lower levels of adenine, N6-methyladenosine and 1-methyladenosine than did those without BRCA1 mutations. Therefore, these metabolites may be potential BC biomarkers associated with BRCA1 mutations.70
Metabolomics serves as a treatment evaluator and monitor of BC
The pharmacokinetics and pharmacodynamics revealed by metabolomics can act as guides to evaluate the efficiency and side effect of BC treatment.71 Ewens et al used a mouse model to examine the efficacy of doxorubicin plus interleukin-2 chemoimmunotherapy against BC and then analysed serum samples by NMR. The results showed that mice that responded to the treatment took on a unique NMR spectral profile with the high expression of chemicals.72 This finding indicated that metabolomics may predict the patients response to treatment to determine the best prescription for individuals. Another group reported that the polymer-conjugated form of doxorubicin showed a more prominent increase in apoptosis, less glycolysis and a decreased level of phospholipids via evaluating the biochemical alterations induced by doxorubicin and its N-(2-hydroxypropyl) methacrylamide copolymer-conjugated form.73 Another group analysed the metabolomics profile of trastuzumab-paclitaxel neo-adjuvant therapy and identified a good response in HER-2 positive BC and further revealed a higher level of circulating spermidine and a lower level of tryptophan in these patients.74 Another group identified 24 biomarkers and found the pathways of protein and cysteine synthesis, glutamine metabolism and the ammonia cycle were related to drug resistance by using MCF-7/ADR and its parental MCF-7/S cells with ultra-performance liquid chromatography-tandem mass spectrometry.75 Moreover, the ratios of GSH/GSSG, ATP/ADP and ATP/AMP were prominently higher in MCF-7/ADR cells compared to MCF-7/S cells.76 Statins are considered to have antitumour potential in recent years.77 The proteomics showed that the lovastatin regulates specific proteins related to cell proliferation, and lovastatin exposure referred to the induction of oxidative stress and the suppression of glycolytic, Krebs cycle or lipid biosynthesis.78 In addition, the possibilities of early recurrence may also be revealed by metabolite profiles.79 Baseline lactate, alanine, and body fat were all correlated with weight gain, which is a risk factor related to recurrence in patients receiving chemotherapy. In addition, the patients who respond to chemotherapy can also be revealed by the lactate level.80
Radiotherapy is also an indispensable part of BC treatment. The serum of 115 female patients who received loco-regional radiotherapy (RT) post-surgery showed a lower concentration of glycolysis, citric acid cycle and amino acid metabolism, especially serine, leucine and isoleucine, before radiotherapy and after surgery; however, this phenomenon was reversed completely or partially after radiotherapy. These changes in metabolites indicated that residual micro-metastatic diseases had been cleared by radiotherapy and thus be an evaluator of its efficacy. However, it is uncertain whether the delay in the post-surgery effect or the concomitant treatment with chemotherapy or hormone therapy can influence the reverse in the concentration of metabolites.81
As one of the most famous anti-neoplastic agents, cisplatin has been widely used for treating tumours.82 The cysteine-cysteine, 3-hydroxy-butyrate based on GC/MS analysis and 3-acylcarnitines and phosphatidylethanolamine with C18:2-C18:2 based on LC/MS analysis were significantly different in the early stage of cisplatin-induced nephrotoxicity, and these metabolites can be detected earlier than the traditional biomarkers of acute kidney injury, such as BUN and creatinine.83 Psychoneurologic (PN) symptom means that patients undergoing chemotherapy may experience pain, fatigue and depression. A previous group analysed changes in the global and targeted metabolomics of the tryptophan pathway before and after chemotherapy compared with the changes in PN symptoms pre-chemotherapy and post-chemotherapy in 19 early-stage BC samples. Global metabolites showed higher concentrations of acetyl-l-alanine and indoxyl sulfate and lower levels of 5-oxo-l-proline post-chemotherapy. Moreover, the targeted analysis revealed higher kynurenine levels and kynurenine/tryptophan ratios.84 Another group found that pre-treatment concentrations of histidine, phenylalanine, and threonine may be the candidate biomarkers of the severity of PN after chemotherapy.85 Metabolomics can be a complementary approach that offers information on the drug responses and side effects throughout the body.86
Metabolomics serves as a prognostic predictor of BC
The glycolysis process in malignant tumour cells is still active despite sufficient oxygen. The phenomenon that cancer cells incline to aerobic glycolysis rather than the more efficient oxidative phosphorylation pathway called the “Warburg effect”.87 An elevated level of ketones and lactates can accelerate cancer growth and metastasis, which may be promising biomarkers to predict poor clinical outcomes in BC patients.88 Neu5Ac (N-acetylneuraminic acid) was found to be the most obvious different glucose metabolite in TNBC cell lines— malignant (BPLER) and less malignant (HMLER). Further exploration showed that the overexpression of CMAS in HMLER promoted the invasive ability of the cells, and the knockdown of CMAS in BPLER prominently destroyed the invasiveness of the cells.89
Overweight is a risk factor for BC recurrence.90 Notably, 27-hydroxycholesterol, an oxysterol metabolite of cholesterol, was supposed to contribute to the initiation of distal metastasis.91 In highly invasive cell lines (MDA-MB-231; BT-549), membrane phospholipids, including phosphatidylglycerol (PG), phosphatidic acid (PA), phosphatidylethanolamine (PE), phosphatidylserine (PS), phosphatidylcholine (PC) and phosphatidylinositol (PI), were supposed to be the most increased metabolites, while sphingomyelin (SM), PE (O-18:0), fatty acids (FA), glycerophosphoglycerophosphoglycerols (CL) and three unknown lipids were identified as the most decreased metabolites. The high level of PG, which can affect the invasive ability, was supposed to correlate with mitochondrial dysfunction, and the low level of SM may be associated with decreased ceramide, which may affect the apoptosis process.92 The metabolic and lipidomic profiling of MCF-10A, MCF-7 and MDA-MB-231 BC cell lines showed a panel comprising xanthine, glucose-6-phosphate, mannose-6-phosphate, guanine, and adenine, which may be a prognostic marker for BC metastasis, and lipidomic profiling further revealed that the phospholipid levels of metastatic groups were higher than those in the normal cell.93 A pilot study showed that the serum samples from 20 invasive ductal carcinoma patients and healthy controls differentially expressed 15 metabolites, mainly concentrating on the sphingolipid metabolism pathway, phospholipid metabolism pathway and fatty acid β-oxidation pathway.94
The common metastatic sites of BC include the lungs, brain, bone and liver.95 A previous group analysed plasma along with pulmonary metastasis progression by inoculating 4T1 metastatic BC cells in mice by LC-MS/MS-based targeted metabolomics and lipidomics and found that alterations in the arginase (ARG) pathway could imply early stage cancer metastasis and be a possible target for therapy in the future.96 Alterations in glycolysis and the signalling and structure of lipids could be alternative biomarkers for aggressive cancer and late phase metastasis.96 A research group found that creatinine and allantoin decreased significantly and trimethylamine, trimethylamine-N-oxide, creatine+phosphocreatine and taurine were all more abundant in the early phase when analysing urine metabolites of BALB/c mice by (1)H NMR spectroscopy.97 In addition, an elevation in pro-metastatic arachidonic acid and a reduction in antimetastatic lysophosphatidylcholines can be identified during bone metastasis.98
The survival outcome can be evaluated by metabolomics. Metabolites, including glucose, glycine, serine and acetate, are significantly associated with patient survival.99 Low concentrations of Z-4OHtam and Z-endoxifen refer to the poor survival outcome of patients.100 Thus, the outcomes of the curative effects of patients may be evaluated directly through metabolomics.
Conclusions and future expectations
Metabolism alteration in tumours are mainly focused on glucose, lipid, amino acids, glutamine,41 purine and pyrimidine.70 Glucose metabolism in BC is usually refers to the “Warburg effect”, which illustrates that cancer glucose utilization, even with adequate oxygen, would incline to the glycolysis.87 Lipid metabolism as an energy source is connected to membrane fluidity and the adhesion of tumour cells, which is related to the metastasis ability of tumour cells.91,98 Elevations in amino acid, purine and pyrimidine metabolism are mainly caused by the high requirement of substrates for the biosynthesis of BC cells, indicating the active proliferation of cancer cells.93 Glutamine metabolism alterations in several cancers has garnered attention because it provided carbon and nitrogen sources for cancer cell and ATP biosynthesis. In short, glutamine not only acts as a biosynthesis substrate promoting cancer growth but also offers energy resources.101–103 Recent metabolomics research is summarized in Table 2. We found that studies that refer to the same metabolism pathway do not exactly show the same results, which may be attributed to the different choices of technologies, samples and data processing methods. The requirement of analysing metabolomics is relatively high due to short times and unbiased small metabolite analyses, and data processing and pattern recognition are still immature. Another marked challenge of metabolomics is the heterogeneity of the tumour. Despite these challenges, metabolic pathways may be potential targets for drug therapy, and metabolomics in cancer have provided new insights to diagnose and treat patients. Metabolomics makes the further classification of breast cancer possible so that patients can receive more specific personal treatment options. The different metabolites in cancer have been revealed as biomarkers for the diagnosis, treatment and monitoring of BC.104 Therefore, the application of metabolomics in BC is promising, despite the temporary limitations.
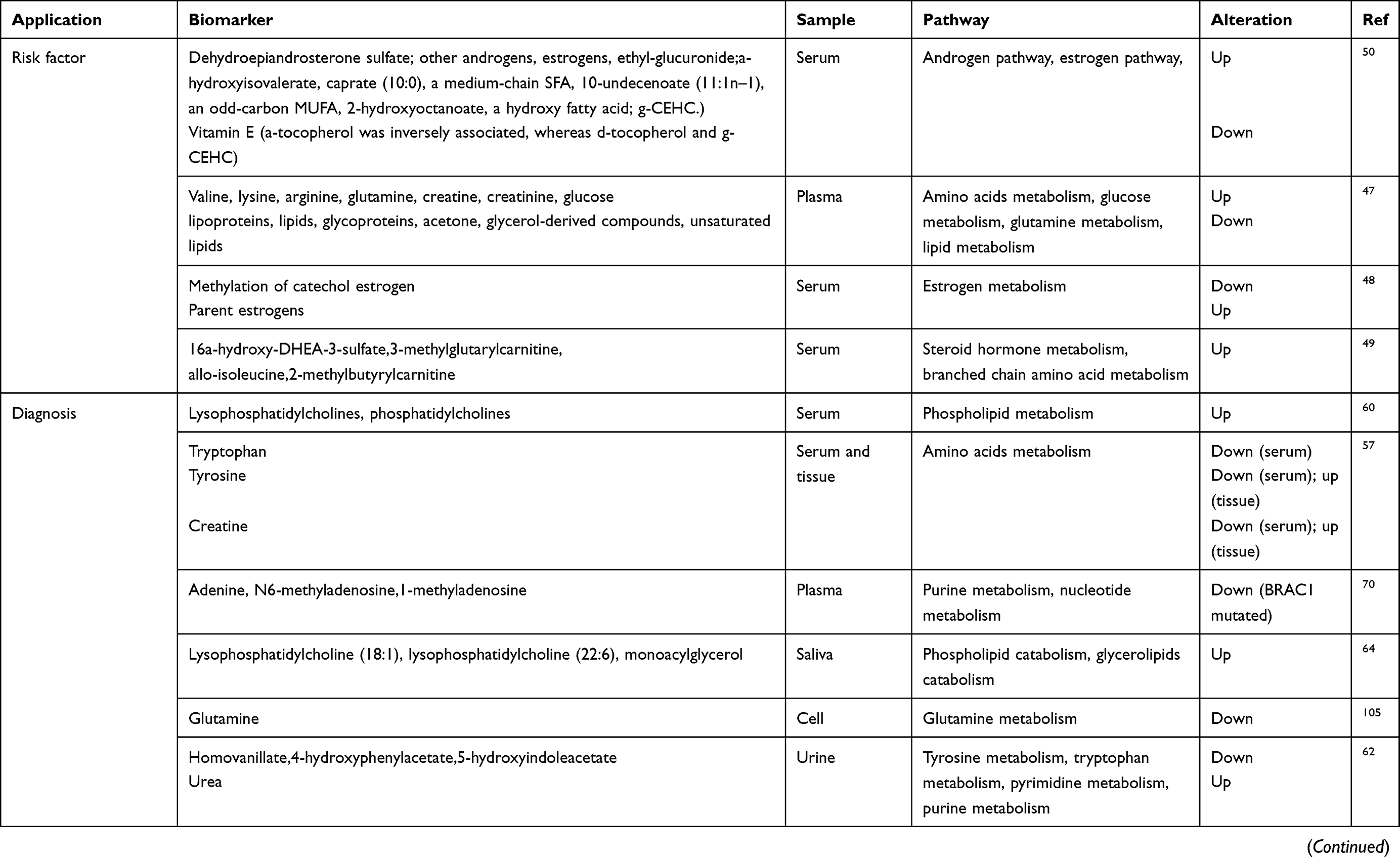 |
Table 2 The application of metabolomics in the risk factors, diagnosis, treatment and prognosis of breast cancer |
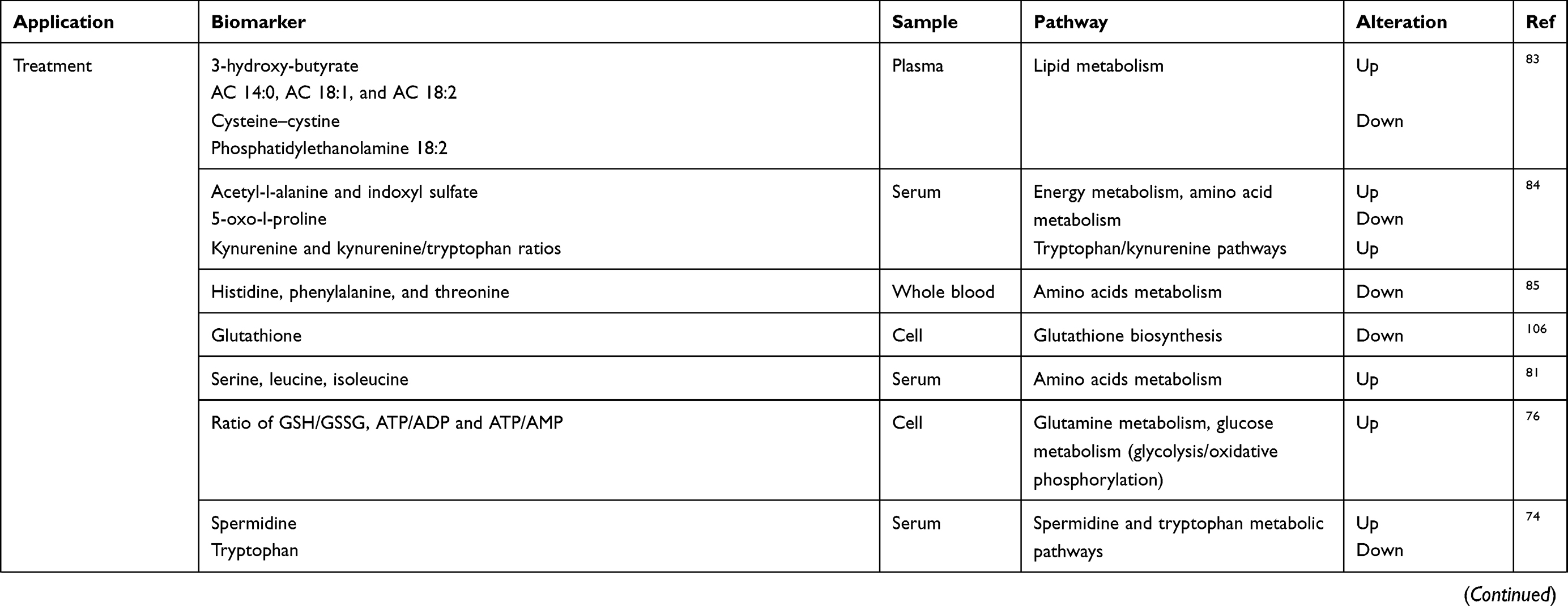 |
Table 2 (Continued). |
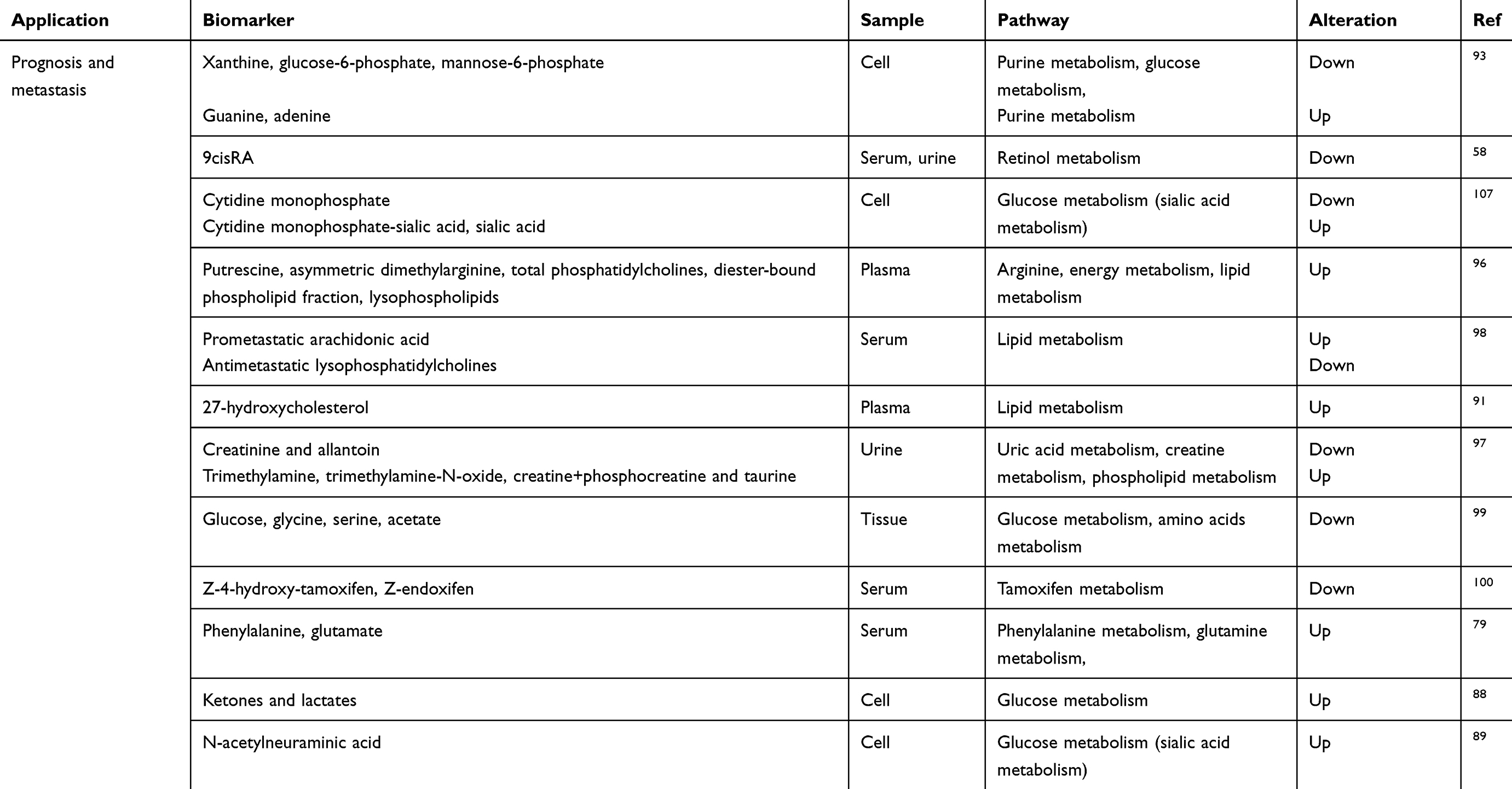 |
Table 2 (Continued). |
Acknowledgments
This work was supported by an individualized support project for original scientific research of Fudan University (XM03180313), a grant (14‑E‑34) from the Funded Project of Baoshan District Science and Technology Commission, Shanghai, and a grant (SHDC12014207) from the Shen Kang Hospital Development Center Foundation.
Author contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wishart DS. Current progress in computational metabolomics. Brief Bioinform. 2007;8(5):279–293. doi:10.1093/bib/bbm030
2. Gates SC, Sweeley CC. Quantitative metabolic profiling based on gas chromatography. Clin Chem. 1978;24(10):1663–1673.
3. Yan M, Qi H, Xia T, et al. Metabolomics profiling of metformin-mediated metabolic reprogramming bypassing AMPKalpha. Metabolism. 2019;91:18–29. doi:10.1016/j.metabol.2018.11.010
4. Preti G. Metabolomics comes of age? The Scientist. 2005;19(11):8.
5. Patti GJ, Yanes O, Siuzdak G. Metabolomics: the apogee of the omics trilogy. Nat Rev Mol Cell Biol. 2012;13(4):263–269. doi:10.1038/nrm3314
6. Jagannathan NR, Sharma U. Breast tissue metabolism by magnetic resonance spectroscopy. Metabolites. 2017;7:2. doi:10.3390/metabo7020025
7. Hadi NI, Jamal Q, Iqbal A, Shaikh F, Somroo S, Musharraf SG. Serum metabolomic profiles for breast cancer diagnosis, grading and staging by gas chromatography-mass spectrometry. Sci Rep. 2017;7(1):1715. doi:10.1038/s41598-017-01924-9
8. Harvie MN, Sims AH, Pegington M, et al. Intermittent energy restriction induces changes in breast gene expression and systemic metabolism. Breast Cancer Res. 2016;18(1):57. doi:10.1186/s13058-016-0714-4
9. Cavaco C, Pereira JAM, Taunk K, et al. Screening of salivary volatiles for putative breast cancer discrimination: an exploratory study involving geographically distant populations. Anal Bioanal Chem. 2018;410(18):4459–4468. doi:10.1007/s00216-018-1103-x
10. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
11. Zaimenko I, Lisec J, Stein U, Brenner W. Approaches and techniques to characterize cancer metabolism in vitro and in vivo. Biochimica Et Biophysica Acta Rev Cancer. 2017;1868(2):412–419. doi:10.1016/j.bbcan.2017.08.004
12. Cardoso MR, Santos JC, Ribeiro ML, Talarico MCR, Viana LR, Derchain SFM. A metabolomic approach to predict breast cancer behavior and chemotherapy response. Int J Mol Sci. 2018;19:2. doi:10.3390/ijms19020617
13. Haukaas TH, Euceda LR, Giskeodegard GF, Bathen TF. Metabolic portraits of breast cancer by HR MAS MR spectroscopy of intact tissue samples. Metabolites. 2017;7:2. doi:10.3390/metabo7020018
14. Denkert C, Bucher E, Hilvo M, et al. Metabolomics of human breast cancer: new approaches for tumor typing and biomarker discovery. Genome Med. 2012;4(4):37.
15. Fiehn O. Combining genomics, metabolome analysis, and biochemical modelling to understand metabolic networks. Comp Funct Genomics. 2001;2(3):155–168. doi:10.1002/cfg.82
16. Zhang T, Zhang A, Qiu S, Yang S, Wang X. Current trends and innovations in bioanalytical techniques of metabolomics. Crit Rev Anal Chem. 2016;46(4):342–351. doi:10.1080/10408347.2015.1079475
17. Markley JL, Bruschweiler R, Edison AS, et al. The future of NMR-based metabolomics. Curr Opin Biotechnol. 2017;43:34–40. doi:10.1016/j.copbio.2016.08.001
18. Nassar AF, Wu T, Nassar SF, Wisnewski AV. UPLC-MS for metabolomics: a giant step forward in support of pharmaceutical research. Drug Discov Today. 2017;22(2):463–470. doi:10.1016/j.drudis.2016.11.020
19. Ramautar R. Capillary electrophoresis-mass spectrometry for clinical metabolomics. Adv Clin Chem. 2016;74:1–34. doi:10.1016/bs.acc.2015.12.002
20. Goodacre R, Vaidyanathan S, Dunn WB, Harrigan GG, Kell DB. Metabolomics by numbers: acquiring and understanding global metabolite data. Trends Biotechnol. 2004;22(5):245–252. doi:10.1016/j.tibtech.2004.03.007
21. Kim HK, Choi Yh Fau - Verpoorte R, Verpoorte R. NMR-based Plant Metabolomics: Where Do We Stand, Where Do We Go? Trends in biotechnology. 2011;29(6):267–275. doi:10.1016/j.tibtech.2011.02.001
22. Karaman I. Preprocessing and pretreatment of metabolomics data for statistical analysis. Adv Exp Med Biol. 2017;965:145–161. doi:10.1007/978-3-319-47656-8_6
23. Wu Y, Li L. Sample normalization methods in quantitative metabolomics. J Chromatogr A. 2016;1430:80–95. doi:10.1016/j.chroma.2015.12.007
24. van Den Berg RA, Hoefsloot HC, Westerhuis JA, Smilde AK, van der Werf MJ. Centering, scaling, and transformations: improving the biological information content of metabolomics data. BMC Genomics. 2006;7:142. doi:10.1186/1471-2164-7-142
25. Krumsiek J, Bartel J, Theis FJ. Computational approaches for systems metabolomics. Curr Opin Biotechnol. 2016;39:198–206. doi:10.1016/j.copbio.2016.04.009
26. Lind MV, Savolainen OI, Ross AB. The use of mass spectrometry for analysing metabolite biomarkers in epidemiology: methodological and statistical considerations for application to large numbers of biological samples. Eur J Epidemiol. 2016;31(8):717–733. doi:10.1007/s10654-016-0166-2
27. Considine EC, Thomas G, Boulesteix AL, Khashan AS, Kenny LC. Critical review of reporting of the data analysis step in metabolomics. Metabolomics. 2017;14(1):7. doi:10.1007/s11306-017-1299-3
28. Caesar LK, Kvalheim OM, Cech NB. Hierarchical cluster analysis of technical replicates to identify interferents in untargeted mass spectrometry metabolomics. Anal Chim Acta. 2018;1021:69–77. doi:10.1016/j.aca.2018.03.013
29. Shen J, Yan L, Liu S, Ambrosone CB, Zhao H. Plasma metabolomic profiles in breast cancer patients and healthy controls: by race and tumor receptor subtypes. Transl Oncol. 2013;6(6):757–765. doi:10.1593/tlo.13619
30. Audano M, Maldini M, De Fabiani E, Mitro N, Caruso D. Gender-related metabolomics and lipidomics: from experimental animal models to clinical evidence. J Proteomics. 2018;178:82–91. doi:10.1016/j.jprot.2017.11.001
31. Kakkoura MG, Sokratous K, Demetriou CA, et al. Mediterranean diet-gene interactions: a targeted metabolomics study in Greek-Cypriot women. Mol Nutr Food Res. 2017;61:4. doi:10.1002/mnfr.201600558
32. Tayyari F, Gowda GAN, Olopade OF, et al. Metabolic profiles of triple-negative and luminal A breast cancer subtypes in African-American identify key metabolic differences. Oncotarget. 2018;9(14):11677–11690. doi:10.18632/oncotarget.24433
33. Tredwell GD, Miller JA, Chow HH, Thompson PA, Keun HC. Metabolomic characterization of nipple aspirate fluid by (1)H NMR spectroscopy and GC-MS. J Proteome Res. 2014;13(2):883–889. doi:10.1021/pr400924k
34. Veneroni S, Dugo M, Daidone MG, et al. Applicability of under vacuum fresh tissue sealing and cooling to omics analysis of tumor tissues. Biopreserv Biobank. 2016;14(6):480–490. doi:10.1089/bio.2015.0093
35. Haukaas TH, Moestue SA, Vettukattil R, et al. Impact of freezing delay time on tissue samples for metabolomic studies. Front Oncol. 2016;6:17. doi:10.3389/fonc.2016.00017
36. Eroles P, Bosch A, Perez-Fidalgo JA, Lluch A. Molecular biology in breast cancer: intrinsic subtypes and signaling pathways. Cancer Treat Rev. 2012;38(6):698–707. doi:10.1016/j.ctrv.2011.11.005
37. Kalimutho M, Parsons K, Mittal D, Lopez JA, Srihari S, Khanna KK. Targeted therapies for triple-negative breast cancer: combating a stubborn disease. Trends Pharmacol Sci. 2015;36(12):822–846. doi:10.1016/j.tips.2015.08.009
38. Fan Y, Zhou X, Xia TS, et al. Human plasma metabolomics for identifying differential metabolites and predicting molecular subtypes of breast cancer. Oncotarget. 2016;7(9):9925–9938. doi:10.18632/oncotarget.7155
39. Jerby L, Wolf L, Denkert C, et al. Metabolic associations of reduced proliferation and oxidative stress in advanced breast cancer. Cancer Res. 2012;72(22):5712–5720. doi:10.1158/0008-5472.CAN-12-2215
40. Budczies J, Brockmoller SF, Muller BM, et al. Comparative metabolomics of estrogen receptor positive and estrogen receptor negative breast cancer: alterations in glutamine and beta-alanine metabolism. J Proteomics. 2013;94:279–288. doi:10.1016/j.jprot.2013.10.002
41. Cappelletti V, Iorio E, Miodini P, Silvestri M, Dugo M, Daidone MG. Metabolic footprints and molecular subtypes in breast cancer. Dis Markers. 2017;2017:7687851. doi:10.1155/2017/7687851
42. Giudetti AM, De Domenico S, Ragusa A, et al. A specific lipid metabolic profile is associated with the epithelial mesenchymal transition program. Biochimica Et Biophysica Acta Mol Cell Biol Lipids. 2019;1864(3):344–357. doi:10.1016/j.bbalip.2018.12.011
43. Winnike JH, Stewart DA, Pathmasiri WW, McRitchie SL, Sumner SJ. Stable isotope-resolved metabolomic differences between hormone-responsive and triple-negative breast cancer cell lines. Int J Breast Cancer. 2018;2018:2063540. doi:10.1155/2018/2063540
44. Borgan E, Sitter B, Lingjaerde OC, et al. Merging transcriptomics and metabolomics–advances in breast cancer profiling. BMC Cancer. 2010;10:628. doi:10.1186/1471-2407-10-663
45. Aure MR, Vitelli V, Jernstrom S, et al. Integrative clustering reveals a novel split in the luminal A subtype of breast cancer with impact on outcome. Breast Cancer Res. 2017;19(1):44. doi:10.1186/s13058-017-0812-y
46. Sun YS, Zhao Z, Yang ZN, et al. Risk factors and preventions of breast cancer. Int J Biol Sci. 2017;13(11):1387–1397. doi:10.7150/ijbs.21635
47. Lecuyer L, Victor Bala A, Deschasaux M, et al. NMR metabolomic signatures reveal predictive plasma metabolites associated with long-term risk of developing breast cancer. Int J Epidemiol. 2018;47(2):484–494. doi:10.1093/ije/dyx271
48. Oh H, Coburn SB, Matthews CE, et al. Anthropometric measures and serum estrogen metabolism in postmenopausal women: the Women’s Health Initiative Observational Study. Breast Cancer Res. 2017;19(1):28. doi:10.1186/s13058-017-0810-0
49. Moore SC, Playdon MC, Sampson JN, et al. A metabolomics analysis of body mass index and postmenopausal breast cancer risk. J Natl Cancer Inst. 2018;110(6):588–597. doi:10.1093/jnci/djx244
50. Playdon MC, Ziegler RG, Sampson JN, et al. Nutritional metabolomics and breast cancer risk in a prospective study. Am J Clin Nutr. 2017;106(2):637–649. doi:10.3945/ajcn.116.150912
51. Beger RD, Dunn W, Schmidt MA, et al. Metabolomics enables precision medicine: “A white paper, community perspective”. Metabolomics. 2016;12(9):149. doi:10.1007/s11306-016-1094-6
52. Wang Q, Sun T, Cao Y, et al. A dried blood spot mass spectrometry metabolomic approach for rapid breast cancer detection. Onco Targets Ther. 2016;9:1389–1398. doi:10.2147/OTT.S95862
53. Siminska E, Koba M. Amino acid profiling as a method of discovering biomarkers for early diagnosis of cancer. Amino Acids. 2016;48(6):1339–1345. doi:10.1007/s00726-016-2215-2
54. Yuan B, Schafferer S, Tang Q, et al. A plasma metabolite panel as biomarkers for early primary breast cancer detection. Int J Cancer. 2019;144(11):2833–2842. doi:10.1002/ijc.31996
55. Wang X, Zhao X, Chou J, et al. Taurine, glutamic acid and ethylmalonic acid as important metabolites for detecting human breast cancer based on the targeted metabolomics. Cancer Biomarkers. 2018. doi:10.3233/CBM-181500
56. Liao HW, Chen GY, Wu MS, Liao WC, Lin CH, Kuo CH. Development of a postcolumn infused-internal standard liquid chromatography mass spectrometry method for quantitative metabolomics studies. J Proteome Res. 2017;16(2):1097–1104. doi:10.1021/acs.jproteome.6b01011
57. More TH, RoyChoudhury S, Christie J, et al. Metabolomic alterations in invasive ductal carcinoma of breast: a comprehensive metabolomic study using tissue and serum samples. Oncotarget. 2018;9(2):2678–2696. doi:10.18632/oncotarget.23626
58. Wu J, Yang R, Zhang L, et al. Metabolomics research on potential role for 9-cis-retinoic acid in breast cancer progression. Cancer Sci. 2018;109(7):2315–2326. doi:10.1111/cas.13629
59. Lipovka Y, Konhilas JP. The complex nature of oestrogen signalling in breast cancer: enemy or ally? Biosci Rep. 2016;36:3. doi:10.1042/BSR20160046
60. Okamoto Y, Aoki A, Ueda K, Jinno H. Metabolomic analysis uncovered an association of serum phospholipid levels with estrogen-induced mammary tumors in female ACI/Seg rats. Toxicol Lett. 2018;288:65–70. doi:10.1016/j.toxlet.2018.02.017
61. Cala M, Aldana J, Sanchez J, Guio J, Meesters RJW. Urinary metabolite and lipid alterations in Colombian Hispanic women with breast cancer: a pilot study. J Pharm Biomed Anal. 2018;152:234–241. doi:10.1016/j.jpba.2018.02.009
62. Nam H, Chung BC, Kim Y, Lee K, Lee D. Combining tissue transcriptomics and urine metabolomics for breast cancer biomarker identification. Bioinformatics. 2009;25(23):3151–3157. doi:10.1093/bioinformatics/btp558
63. Sugimoto M, Wong DT, Hirayama A, Soga T, Tomita M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics. 2010;6(1):78–95. doi:10.1007/s11306-009-0178-y
64. Zhong L, Cheng F, Lu X, Duan Y, Wang X. Untargeted saliva metabonomics study of breast cancer based on ultra performance liquid chromatography coupled to mass spectrometry with HILIC and RPLC separations. Talanta. 2016;158:351–360. doi:10.1016/j.talanta.2016.04.049
65. Takayama T, Tsutsui H, Shimizu I, et al. Diagnostic approach to breast cancer patients based on target metabolomics in saliva by liquid chromatography with tandem mass spectrometry. Clin Chim Acta. 2016;452:18–26. doi:10.1016/j.cca.2015.10.032
66. Mannello F. New horizon for breast cancer biomarker discoveries: what might the liquid biopsy of nipple aspirate fluid hold? Proteomics Clin appli. 2017;11:9–10. doi:10.1002/prca.v11.9-10
67. Matos Do Canto L, Marian C, Varghese RS, et al. Metabolomic profiling of breast tumors using ductal fluid. Int J Oncol. 2016;49(6):2245–2254. doi:10.3892/ijo.2016.3732
68. Phillips M, Cataneo R, Lebauer C, Mundada M, Saunders C. Breath mass ion biomarkers of breast cancer. J Breath Res. 2017;11(1):5. doi:10.1088/1752-7163/aa549b
69. Silva CL, Perestrelo R, Silva P, Tomas H, Camara JS. Volatile metabolomic signature of human breast cancer cell lines. Sci Rep. 2017;7:43969. doi:10.1038/srep43969
70. Roig B, Rodríguez-Balada M, Samino S, et al. Metabolomics reveals novel blood plasma biomarkers associated to the BRCA1-mutated phenotype of human breast cancer. Sci Rep. 2017;7. doi:10.1038/s41598-017-17897-8
71. Kaddurah-Daouk R, Kristal BS, Weinshilboum RM. Metabolomics: a global biochemical approach to drug response and disease. Annu Rev Pharmacol Toxicol. 2008;48:653–683. doi:10.1146/annurev.pharmtox.48.113006.094715
72. Ewens A, Luo L, Berleth E, et al. Doxorubicin plus interleukin-2 chemoimmunotherapy against breast cancer in mice. Cancer Res. 2006;66(10):5419–5426. doi:10.1158/0008-5472.CAN-05-3963
73. Arminan A, Palomino-Schatzlein M, Deladriere C, et al. Metabolomics facilitates the discrimination of the specific anti-cancer effects of free- and polymer-conjugated doxorubicin in breast cancer models. Biomaterials. 2018;162:144–153. doi:10.1016/j.biomaterials.2018.02.015
74. Miolo G, Muraro E, Caruso D, et al. Pharmacometabolomics study identifies circulating spermidine and tryptophan as potential biomarkers associated with the complete pathological response to trastuzumab-paclitaxel neoadjuvant therapy in HER-2 positive breast cancer. Oncotarget. 2016;7(26):39809–39822. doi:10.18632/oncotarget.9489
75. Zhang R, Zhuang X, Zong L, Liu S, Liu Z, Song F. Investigations on the cell metabolomics basis of multidrug resistance from tumor cells by ultra-performance liquid chromatography-mass spectrometry. Anal Bioanal Chem. 2016;408(21):5843–5854. doi:10.1007/s00216-016-9696-4
76. Zong L, Pi Z, Liu S, Xing J, Liu Z, Song F. Liquid extraction surface analysis nanospray electrospray ionization based lipidomics for in situ analysis of tumor cells with multidrug resistance. Rapid Commun Mass Spectrom. 2018;32(19):1683–1692. doi:10.1002/rcm.8229
77. Borgquist S, Bjarnadottir O, Kimbung S, Ahern TP. Statins: a role in breast cancer therapy? J Intern Med. 2018;284(4):346–357. doi:10.1111/joim.12806
78. Klawitter J, Shokati T, Moll V, Christians U, Klawitter J. Effects of lovastatin on breast cancer cells: a proteo-metabonomic study. Breast Cancer Res. 2010;12(2):R16. doi:10.1186/bcr2722
79. Hart CD, Vignoli A, Tenori L, et al. Serum metabolomic profiles identify ER-positive early breast cancer patients at increased risk of disease recurrence in a multicenter population. Clin Cancer Res. 2017;23(6):1422–1431. doi:10.1158/1078-0432.CCR-16-1153
80. Keun HC, Sidhu J, Pchejetski D, et al. Serum molecular signatures of weight change during early breast cancer chemotherapy. Clin Cancer Res. 2009;15(21):6716–6723. doi:10.1158/1078-0432.CCR-09-1452
81. Arenas M, Rodriguez E, Garcia-Heredia A, et al. Metabolite normalization with local radiotherapy following breast tumor resection. PLoS One. 2018;13(11):e0207474. doi:10.1371/journal.pone.0207474
82. Dasari S, Tchounwou PB. Cisplatin in cancer therapy: molecular mechanisms of action. Eur J Pharmacol. 2014;740:364–378. doi:10.1016/j.ejphar.2014.07.025
83. Ezaki T, Nishiumi S, Azuma T, Yoshida M. Metabolomics for the early detection of cisplatin-induced nephrotoxicity. Toxicol Res (Camb). 2017;6(6):843–853. doi:10.1039/c7tx00171a
84. Lyon DE, Starkweather A, Yao Y, et al. Pilot study of metabolomics and psychoneurological symptoms in women with early stage breast cancer. Biol Res Nurs. 2018;20(2):227–236. doi:10.1177/1099800417747411
85. Sun Y, Kim JH, Vangipuram K, et al. Pharmacometabolomics reveals a role for histidine, phenylalanine, and threonine in the development of paclitaxel-induced peripheral neuropathy. Breast Cancer Res Treat. 2018;171(3):657–666. doi:10.1007/s10549-018-4862-3
86. Lindon JC, Holmes E, Bollard ME, Stanley EG, Nicholson JK. Metabonomics technologies and their applications in physiological monitoring, drug safety assessment and disease diagnosis. Biomarkers. 2004;9(1):1–31. doi:10.1080/13547500410001668379
87. Warburg O, Wind F, Negelein E. THE METABOLISM OF TUMORS IN THE BODY. J Gen Physiol. 1927;8(6):519–530. doi:10.1085/jgp.8.6.519
88. Martinez-Outschoorn UE, Prisco M, Ertel A, et al. Ketones and lactate increase cancer cell “stemness,” driving recurrence, metastasis and poor clinical outcome in breast cancer: achieving personalized medicine via metabolo-genomics. Cell Cycle. 2011;10(8):1271–1286. doi:10.4161/cc.10.8.15330
89. O’Day EM, Idos GE, Hill C, Chen JW, Wagner G. Cytidine monophosphate N-acetylneuraminic acid synthetase enhances invasion of human triple-negative breast cancer cells. Onco Targets Ther. 2018;11:6827–6838. doi:10.2147/OTT.S177639
90. Kyrgiou M, Kalliala I, Markozannes G, et al. Adiposity and cancer at major anatomical sites: umbrella review of the literature. BMJ. 2017;356:j477. doi:10.1136/bmj.j477
91. Baek AE, Yu YA, He S, et al. The cholesterol metabolite 27 hydroxycholesterol facilitates breast cancer metastasis through its actions on immune cells. Nat Commun. 2017;8(1):864. doi:10.1038/s41467-017-00910-z
92. Wang S, Chen X, Luan H, et al. Matrix-assisted laser desorption/ionization mass spectrometry imaging of cell cultures for the lipidomic analysis of potential lipid markers in human breast cancer invasion. Rapid Commun Mass Spectrom. 2016;30(4):533–542. doi:10.1002/rcm.7466
93. Kim HY, Lee KM, Kim SH, Kwon YJ, Chun YJ, Choi HK. Comparative metabolic and lipidomic profiling of human breast cancer cells with different metastatic potentials. Oncotarget. 2016;7(41):67111–67128. doi:10.18632/oncotarget.11560
94. Cui M, Wang Q, Chen G. Serum metabolomics analysis reveals changes in signaling lipids in breast cancer patients. Biomed Chromatogr. 2016;30(1):42–47. doi:10.1002/bmc.3556
95. Xie H-Y, Shao Z-M, Li D-Q. Tumor microenvironment: driving forces and potential therapeutic targets for breast cancer metastasis. Chin J Cancer. 2017;36(1):36. doi:10.1186/s40880-017-0202-y
96. Kus K, Kij A, Zakrzewska A, et al. Alterations in arginine and energy metabolism, structural and signalling lipids in metastatic breast cancer in mice detected in plasma by targeted metabolomics and lipidomics. Breast Cancer Res. 2018;20(1):148. doi:10.1186/s13058-018-1075-y
97. Larkin JR, Dickens AM, Claridge TD, et al. Early diagnosis of brain metastases using a biofluids-metabolomics approach in mice. Theranostics. 2016;6(12):2161–2169. doi:10.7150/thno.16538
98. Krzeszinski JY, Schwaid AG, Cheng WY, et al. Lipid osteoclastokines regulate breast cancer bone metastasis. Endocrinology. 2017;158(3):477–489. doi:10.1210/en.2016-1570
99. Auslander N, Yizhak K, Weinstock A, et al. A joint analysis of transcriptomic and metabolomic data uncovers enhanced enzyme-metabolite coupling in breast cancer. Sci Rep. 2016;6:29662. doi:10.1038/srep29662
100. Helland T, Henne N, Bifulco E, et al. Serum concentrations of active tamoxifen metabolites predict long-term survival in adjuvantly treated breast cancer patients. Breast Cancer Res. 2017;19(1):125. doi:10.1186/s13058-017-0916-4
101. El Ansari R, McIntyre A, Craze ML, Ellis IO, Rakha EA, Green AR. Altered glutamine metabolism in breast cancer; subtype dependencies and alternative adaptations. Histopathology. 2018;72(2):183–190. doi:10.1111/his.13334
102. Dong J, Xiao D, Zhao Z, et al. Epigenetic silencing of microRNA-137 enhances ASCT2 expression and tumor glutamine metabolism. Oncogenesis. 2017;6(7):e356. doi:10.1038/oncsis.2017.59
103. Cha YJ, Kim ES, Koo JS. Amino acid transporters and glutamine metabolism in breast cancer. Int J Mol Sci. 2018;19:3. doi:10.3390/ijms19030907
104. McCartney A, Vignoli A, Biganzoli L, et al. Metabolomics in breast cancer: a decade in review. Cancer Treat Rev. 2018;67:88–96. doi:10.1016/j.ctrv.2018.04.012
105. Coloff JL, Murphy JP, Braun CR, et al. Differential glutamate metabolism in proliferating and quiescent mammary epithelial cells. Cell Metab. 2016;23(5):867–880. doi:10.1016/j.cmet.2016.03.016
106. Beatty A, Fink LS, Singh T, et al. Metabolite profiling reveals the glutathione biosynthetic pathway as a therapeutic target in triple-negative breast cancer. Mol Cancer Ther. 2018;17(1):264–275. doi:10.1158/1535-7163.MCT-17-0407
107. Teoh ST, Ogrodzinski MP, Ross C, Hunter KW, Lunt SY. Sialic acid metabolism: a key player in breast cancer metastasis revealed by metabolomics. Front Oncol. 2018;8:174. doi:10.3389/fonc.2018.00174
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.