")
Back to Journals » Infection and Drug Resistance » Volume 13
Mechanisms of Heteroresistance and Resistance to Imipenem in Pseudomonas aeruginosa
Authors Xu Y, Zheng X, Zeng W, Chen T, Liao W, Qian J, Lin J, Zhou C, Tian X, Cao J, Zhou T
Received 12 February 2020
Accepted for publication 21 April 2020
Published 14 May 2020 Volume 2020:13 Pages 1419—1428
DOI https://doi.org/10.2147/IDR.S249475
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Ye Xu,1 Xiangkuo Zheng,2 Weiliang Zeng,2 Tao Chen,1 Wenli Liao,1 Jiao Qian,1 Jie Lin,1 Cui Zhou,1 Xuebin Tian,2 Jianming Cao,2 Tieli Zhou1
1Department of Clinical Laboratory, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang Province, People’s Republic of China; 2Department of Medical Laboratory Science, School of Laboratory Medicine and Life Science, Wenzhou Medical University, Wenzhou, Zhejiang Province, People’s Republic of China
Correspondence: Jianming Cao; Tieli Zhou Tel +86-577-88069595
; +86-577-86689885
Email [email protected]; [email protected]
Background: Heteroresistance is a phenomenon that occurs in all bacteria and can cause treatment failure. Yet, the exact mechanisms responsible for heteroresistance still remain unknown. The following study investigated the mechanisms of imipenem-heteroresistance and -resistance in Pseudomonas aeruginosa clinical isolates from Wenzhou, China.
Methods: Imipenem resistance was detected by the agar dilution method; heteroresistance was determined by population analysis profiles. Biofilm formation assay and modified carbapenem inactivation methods were also performed. Polymerase chain reaction (PCR) was conducted to detect oprD, and quantitative real-time PCR was used to determine expression levels of oprD, ampC and four efflux pump coding genes (mexB, mexD, mexE and mexY).
Results: Six imipenem-heteroresistant and -resistant P. aeruginosa isolates were selected respectively. Deficient oprD was detected in all resistant isolates and two heteroresistant isolates. No strains produced carbapenemases. Expression levels of oprD were down-regulated in heteroresistant isolates. Transcription levels of the mexE and mexY were significantly increased in all heterogeneous subpopulations compared with their respective native ones. Compared with the susceptible group, increased mean relative expression levels of mexE and mexY or the decreased mean relative expression levels of oprD were observed in the resistant group (P < 0.05), whereas transcription levels of the mexB and mexD remained unchanged.
Conclusion: Down-regulation of oprD contributed to the resistance and heteroresistance of imipenem in our P. aeruginosa clinical isolates. In addition, the marginal up-regulation of efflux systems may indirectly affect imipenem resistance. Contrarily, defective oprD was less common in our experimental heteroresistant strains than resistant strains.
Keywords: Pseudomonas aeruginosa, imipenem, resistance, heteroresistance, molecular mechanism, oprD
Introduction
Pseudomonas aeruginosa (P. aeruginosa) is a Gram-negative non-fermenting bacillus highly resistant to antibiotics and among the leading causes of nosocomial infections, such as pneumonia, bloodstream infections, surgical site infections, and urinary tract infections.1 Despite carbapenem-resistant P. aeruginosa, which has been frequently reported worldwide and is characterized by massive intrinsic resistance to multiple classes of antibiotics, carbapenems, primarily imipenem, is still considered the most potent agent of P. aeruginosa.2,3
Heteroresistance is a phenomenon that occurs in both Gram-positive and -negative bacteria, which was first time identified in Haemophilus influenzae in 1947.4 It is generally described as a phenotype where treated bacteria contain subpopulations with lower susceptibility to the antibiotic compared to the dominant population, which, in turn, may cause treatment failure.5,6 Heteroresistance may have certain similarities and differences when compared to homogenous resistance.7 Heteroresistance to carbapenems has been detected in clinical isolates of Acinetobacter baumannii, Enterobacter cloacae, Klebsiella pneumoniae, Escherichia coli and P. aeruginosa at high frequencies.8–14 Yet, unlike inherited resistance, the exact mechanisms responsible for heteroresistance still remain unknown.
Carbapenem homogenous resistance develops mainly due to the expression of the intrinsic AmpC β-lactamase, overexpression of Mex–Opr-type efflux pumps and production of metallo-β-lactamases (MBLs), as well as mutations or repression of OprD porin.15,16 Moreover, studies12,17,18 have suggested that low expression of the OprD porin, overexpression of efflux systems and intrinsic AmpC β-lactamase or metallo-β-lactamases (MBLs) contributed to imipenem-heteroresistant P. aeruginosa.
The aim of this study was to investigate the mechanisms for imipenem heteroresistance in clinical strains of P. aeruginosa isolated from a teaching hospital in Wenzhou, China. We hypostasized that mutations of oprD gene might regulate the imipenem-heteroresistant of P. aeruginosa.
Materials and Methods
Bacterial Strains
A total of 131 P. aeruginosa clinical isolates susceptible to imipenem were collected from the First Affiliated Hospital of Wenzhou Medical University, in 2017. All isolates were identified to species level using a VITEK® MS automated system (BioMérieux, Hazelwood, MO, USA). MICs of imipenem were additionally analyzed by the agar dilution method. The MIC breakpoint of imipenem for P. aeruginosa was interpreted following the latest CLSI guidelines (CLSI 2019):19 susceptible, ≤ 2 μg/mL; intermediate, 4 μg/mL; resistant, ≥ 8 μg/mL.
The disk diffusion method was performed to detect the presence of heterogeneous resistant subpopulations in 131 susceptible isolates. If a visible colony was found inside the clear zone of inhibition, the strains were suspected to be imipenem-heteroresistant. The experiment was carried out three times to see the reproducibility of emergence of colonies.
Population Analysis Profiles (PAPs)
Randomly selected six populations with suspected heteroresistance to imipenem were analyzed based on population analysis profiles (PAPs)1 with minor modifications. Briefly, 0.1 mL starting bacterial suspension with an optical density corresponding to 0.5 McFarland and serial dilutions were spread onto Muller-Hinton agar (OXOID, Basingstoke, United Kingdom) plates containing imipenem in serial two-fold dilutions for concentrations ranging from 0.0625 to 128 μg/mL. The agar plates were then incubated at 37 °C for 48 hours before the Colony-Forming Units (CFU) were calculated. The frequency of heteroresistant subpopulations at the highest drug concentration was counted by dividing the number of colonies grown on the imipenem-containing plate by the colony counts from the same bacterial inoculum grown on antibiotic-free plates. The imipenem-susceptible P. aeruginosa strains ATCC 27853 and PAO1 were used as the control strain. For each isolate, a single colony was selected from the highest antibiotic concentration, and the imipenem MIC was tested using the agar dilution method after serial passaging on antibiotic-free medium for one week to evaluate the stability of the heteroresistant phenotype. Interestingly, the imipenem-heteroresistant subpopulations showed at least fourfold higher MICs than their parental populations (unstable heteroresistant phenotype was excluded). Cultures with resistant or susceptible subpopulations were isolated from the highest imipenem concentration and drug-free medium for further studies, separately. Furthermore, six wild-type imipenem-susceptible (TL-2874, TL-2875, TL-2891, TL-2892, TL2907 and TL-2908, in which heteroresistance did not exist) and -resistant (TL-2854, TL-2858, TL-2859, TL-2878, TL-2958 and TL-2997) strains were selected for further experiments.
Pulsed-Field Gel Electrophoresis (PFGE)
Pulsed-field gel electrophoresis (PFGE) of SpeI-digested genomic DNA of P. aeruginosa isolates was performed with a CHEF-DRIII system (Bio-Rad, Hercules, CA); banding patterns were analyzed as previously described.20 Briefly, genomic DNA was extracted from the P. aeruginosa isolates, followed by restriction enzyme SpeI (Takara, Japan) digestion for 2.5 h. PFGE was performed using a CHEF-Mapper XA PFGE system (Bio-Rad, USA) for 18h with a switch time 5–25 s. Then DNA fingerprints were analyzed by GelRed staining. The banding patterns were visualized by GelDoc XR gel imaging system (Bio-Rad, USA), and cluster analysis of similarity values of the PFGE profiles was finally performed by Quantity One program (BioRad Laboratories, USA).
Antimicrobial Susceptibility Testing
The MICs of clinical routine antimicrobial agents including amikacin (AMK), ceftazidime (CAZ), ciprofloxacin (CIP), levofloxacin (LEV), tobramycin (TOB), cefepime (FEP), piperacillin/tazobactam (TZP) and meropenem (MEM) were determined by the agar dilution method, and carbapenemases were detected using mCIM in accordance with the latest guidelines of CLSI.
Polymerase Chain Reaction (PCR) and Sequencing
Genomic DNAs of imipenem-resistant, -susceptible and -heteroresistant isolates were extracted using the Biospin Bacteria Genomic DNA Extraction Kit (Bioflux, Tokyo, Japan) following the manufacturer’s instructions, were used as templates stored at −20°C for subsequent analysis. Primers for oprD gene amplification (oprD-F, 5ʹ-AATGAGGAAGACCTGGTGGC-3ʹ; oprD-R, 5ʹ-TCGGAACCTCAACTATCGCC-3ʹ) were designed from sequences in the GenBank nucleotide sequence database (https://www.ncbi.nlm.nih.gov/tools/primer-blast/). The amplified productions were sequenced by Shanghai BGI Technology Co., Ltd., and the deduced amino acid sequences of oprD were compared with that from wild-type imipenem-susceptible P. aeruginosa strain PAO1 by BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Quantitative Real-Time PCR (qRT-PCR)
Total RNAs of resistant isolates and heteroresistant isolates (including susceptible and resistant subpopulations) at the exponential phase of growth were extracted by the Bacterial RNA Miniprep Kit (Biomiga, Shanghai, China) according to the manufacturer’s protocol. RNA from each isolate was used as a template for cDNA synthesis using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher, Shanghai, China). The expression levels of ampC β-lactamase gene, the genes coding for porin (oprD) and efflux pumps (mexB, mexD, mexE and mexY) were analyzed by qRT-PCR. The rpsL gene was used as an internal reference. These primers were previously described.21,22 Gene expression levels of heteroresistant subpopulations were expressed as the fold increase relative to the respective parental populations that were used as controls.18 Correspondingly, increases or decreases in gene expression of ≥ 2-fold and ≤ 0.5-fold were considered to be significant. Student’s t-test was used to compare changes of expression levels between susceptible and resistant groups based on P. aeruginosa PAO1, which was assigned a value of 1.0.23
Biofilm Formation Assay
Biofilm formation assay was assayed by crystal violet staining of adherent cells as previously described.24 Briefly, eight parental strains and the respective mutants, as well as 12 susceptible and resistant isolates, adjusted to 1 × 106 CFU/mL were inoculated in LB broth in 96-well polystyrene plates. Following incubation at 37 °C for 18–24 h, the plates were washed two times with phosphate-buffered saline (PBS) and then stained with 1% crystal violet for 15 min. The absorbance of incorporated dye was determined at an optical density of 595 nm. Each isolate was assayed in three wells and the experiments were repeated three times.
Analysis of Outer Membrane Proteins
The extraction of outer membrane proteins was performed according to methods described elsewhere, with minor modifications.14 In brief, cells from an overnight culture in LB broth were harvested by centrifugation at 4000 ×g for 10 min at 4 °C, and the supernatant was removed. Samples were then mixed with 20 mL 50 mM PBS (PH 7.0), and washed in autoclaved ultrapure water. The cell pellets were suspended in suspension buffer (10 mM Tris-HCl [pH 8.0]) supplied with protease inhibitor phenylmethylsulfonyl fluoride (PMSF), followed by sonication for 10 min on ice to break the cells. Next, the unbroken cells were centrifugated at 1600 ×g for 10 min at 4 °C, and the membranes in the supernatant were further pelleted at 21,500 ×g for 30 min at 4 °C, suspended in 3 mL 10 mM PBS containing 2% sodium dodecyl sarcosine, and left on room temperature for 30 min before centrifugation at 20,500 ×g at 4 °C for an additional 40 min. The remaining precipitated outer membrane proteins were suspended in 10 mM PBS (PH 7.0) and quantified using Enhanced BCA Protein Assay Kit (Beyotime Biotechnology) before 1% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis.
Samples mixed with loading buffer (Takara 9173, Japan) were first denatured by heating at 100 °C for 3 min, then they were separated by SDS-PAGE using 12% of separation gel and 5% of concentration gel. Bands were visualized by dyeing the gels with 0.2% Coomassie brilliant blue (Solarbio C8430, China) in 10% acetic acid and 45% methanol. PAO1 served as a control strain for OMPs profiling.
Statistical Analysis
Data were analyzed using the GraphPad Prism analysis package 8.01. The Microsoft Visio Drawing 2019 was performed to the plot. Differences in the biofilm formation and expression of each gene of interest were tested using the Student’s t-test. P < 0.05 was considered statistically significant.
Results
Antibiotics Susceptibility and Homology Characteristics of Susceptibility, Resistance and Heteroresistance Among P. aeruginosa Isolates
After performing a disk diffusion method, 46 of 131 isolates were found within the inhibition zones around the imipenem disks (35.1%, 46/131), suggesting the possibility of inducible resistance to antibiotics in P. aeruginosa under antibiotic pressure. To confirm the heteroresistance in P. aeruginosa, six isolates were randomly selected to perform population analysis profiles (PAPs). Six isolates appeared in a heteroresistant subpopulation (TL-2856, TL-2862, TL-2870, TL-2872, TL-2873 and TL-2877). Minimal inhibitory concentrations (MICs) of the subpopulations were 4- to 8-fold higher than those of the parental cells and the frequency of heteroresistant subpopulations ranged from 2×10−7 to 5.3×10−6 (Table 1). The stability of the heteroresistance subpopulations was maintained after one week of subculturing in antibiotic-free medium. Six stable heteroresistant subpopulations were determined by 16s RNA sequence as P. aeruginosa, and the parental cells and their heteroresistant subpopulations exhibited identical PFGE profiles (Figure S1). The results of susceptibility tests showed four- to sixteen-fold higher MICs values of meropenem in the six isolates compared to susceptible populations (Table 2).
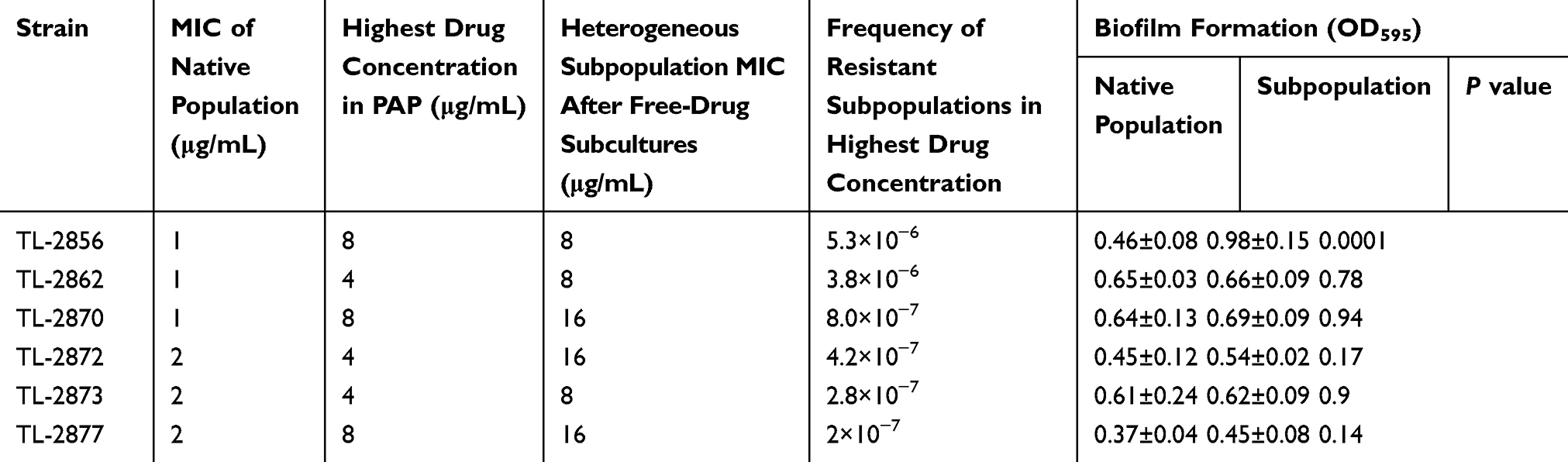 |
Table 1 Characteristics of Heteroresistant Isolates Determined Using PAP, MICs of Imipenem and Levels of Biofilm Formation |
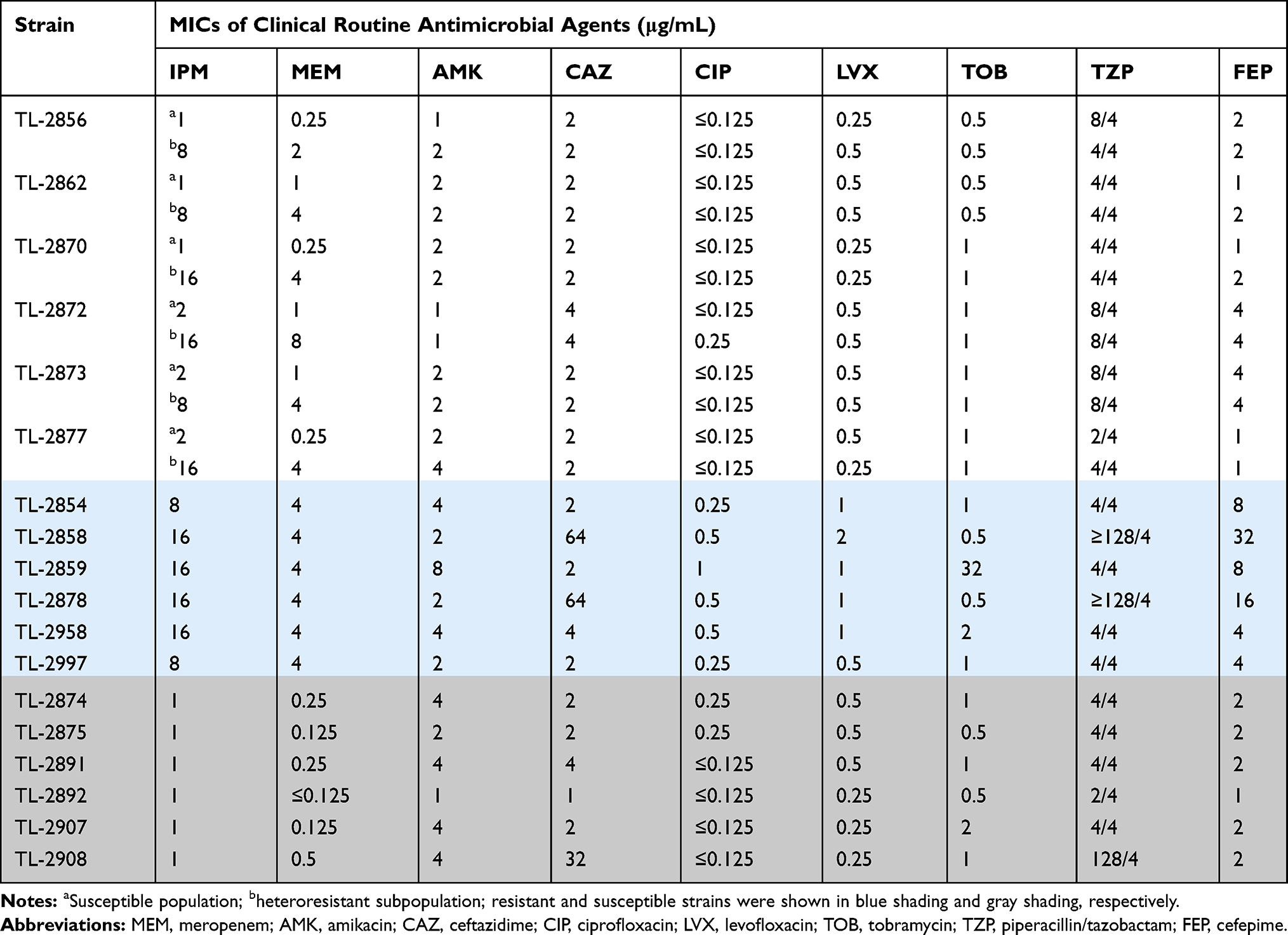 |
Table 2 The Profile of Routine Antibacterial Resistance in P. aeruginosa |
Detection of Carbapenemases and Analysis of Levels of Biofilm Formation
mCIM method indicated that all of the resistant and heteroresistant isolates were negative for carbapenemases. Moreover, the results showed that all heteroresistant subpopulations, except for TL-2856, had a stronger biofilm formation than their respectively native strains (P > 0.05) (Table 1). Moreover, compared with the susceptible group, the resistant group did not produce far-off equivalent biofilm (Figure 1).
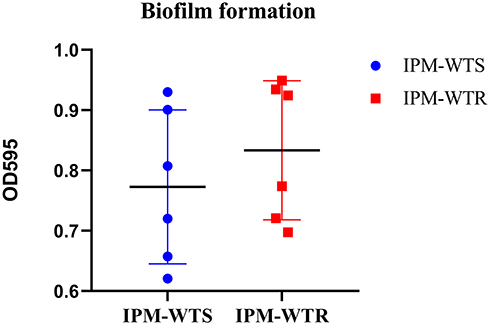 |
Figure 1 Levels of biofilm formation between wild-type susceptible and resistant isolates. IPM-WTS, wild-type imipenem susceptibility strains; IPM-WTR, wild-type imipenem resistance strains. |
Mutations in oprD Gene
In this study, oprD was sequenced in six resistant isolates, twelve susceptible isolates (including six wild-type susceptible strains and native susceptible populations of heteroresistance) and six heteroresistant isolates to investigate possible resistance mechanisms. Various types of amino acid substitutions in oprD gene were found in resistant, heteroresistant subpopulations and susceptible strains (including heteroresistant native susceptible populations and wild-type imipenem-susceptible strains). Furthermore, two 3 bp deletions at nt 1116–1118 and nt 1147–1149 occurred in three heteroresistant strains (TL-2856, TL-2872, and TL-2877), one resistant isolate TL-2997, and some susceptible strains. Interestingly, compared with their respective native strains, a subpopulation of the isolate TL-2870 deleted 1 bp at nt 348 of the oprD gene, causing amino acid 127 premature stop. Besides, a heteroresistant subpopulation of the isolate TL-2873 deleted 15 bp at nt 256–270, causing amino acid 439 premature stop. The sequence of isolate TL-2862 was similar to susceptible strains. In resistant isolates, TL-2854, TL-2859 and TL-2958 showed amino acid 417 premature stop translation. TL-2858 and TL-2878 deleted 379 bp in nt 410–788, causing amino acid 219 premature stop translation. TL-2997 showed 1 bp deletion in nt 917, causing amino acid 345 premature stop translation (Figure 2).
 |
Figure 2 oprD mutations in imipenem-(hetero) resistant P. aeruginosa clinical isolates. aIn imipenem-heteroresistant populations (IPM-HRP); bwild-type imipenem resistant isolates (IPM-WTR); ctwo 3 bp deletion at nucleotide 1116–1118 and nucleotide 1147–1149 in external loop7; d1 bp deletion at nucleotide 348; e379 bp deletion at nucleotide 410–788; f1 bp deletion at nucleotide 917; g1251G > A; h15 bp deletion at nucleotide 256–270; *premature stop codon; blue circle, amino acid substitutions were detected in wild-type imipenem susceptible isolates and native imipenem susceptible populations of heteroresistance and the amino acid substitutions were predicted neutral using http://provean.jcvi.org/seq_submit.php; orange circle, oprD mutation only was detected in heteroresistance and resistance. |
Analysis of Gene Expression Levels
Twenty-four non-carbapenemases-producing isolates (six heteroresistant and their respective native strains, six wild-type susceptible and resistant strains) were used to determine the expression levels of AmpC, efflux pumps and OprD porin by real-time RT-PCR tests. As shown in Figure 3A, all heteroresistant isolates, except for TL-2873 (0.966-fold), showed significantly lower expression levels in oprD compared to their native isolates (ranging from 0.121- to −0.33, P < 0.05). Nevertheless, compared with their native isolates, expression levels of ampC (ranging from 0.998- to 1.21-fold), efflux pump encoding genes mexB (ranging from 1.03- to 1.326-fold) and mexD (ranging from 1.056- to 1.824-fold) in heteroresistant isolates were not statistically significant (P > 0.05). Interestingly, increased expression levels of efflux pump-related genes mexE (ranging from 1.541- to 8.234-fold) and mexY (ranging from 1.561- to 3.398-fold) in all heteroresistant isolates were statistically significant (P < 0.05) relative to their native strains (Figure 3B–F).
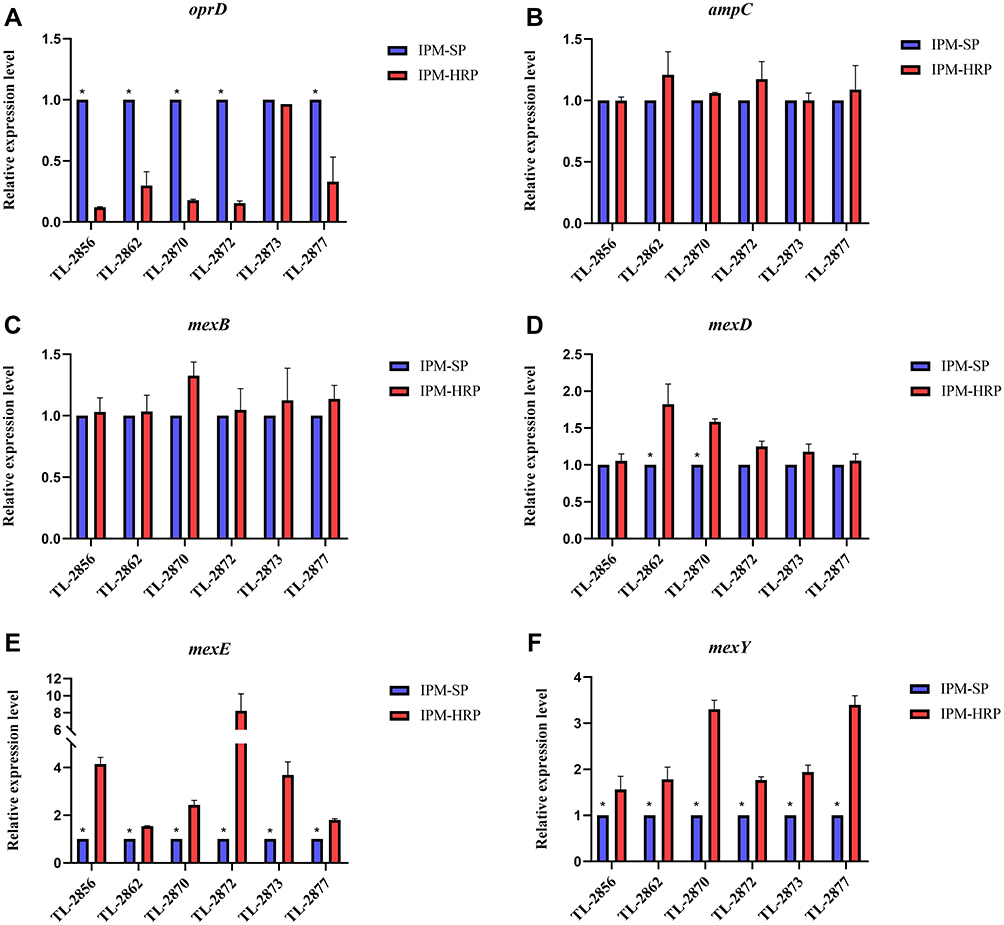 |
Figure 3 Expression levels of genes in heteroresistant subpopulations. (A) Expression levels of oprD. (B) Expression levels of ampC. (C) Expression levels of mexB. (D) Expression levels of mexD. (E) Expression levels of mexE. (F) Expression levels of mexY. IPM-SP, native imipenem-susceptible populations; IPM-HRP, imipenem-heteroresistant subpopulations; *P < 0.05. |
On the other hand, the expression levels of efflux pump genes and outer membrane protein-encoding gene oprD were analyzed in imipenem-resistant strains and an equal number of susceptible strains. The results of qRT-PCR showed that compared with PAO1 the expression levels of efflux pump-related genes mexE in wild-type susceptible group and the resistant group ranged from 0.443- to 1.173-fold and from 1.853- to 6.262-fold, respectively. The difference between a susceptible group and a resistant group was statistically significant (P < 0.05). Similarly, the relative expression level of oprD in resistant strains was lower than in susceptible strains (P < 0.05). Furthermore, ampC gene and efflux pumps encoding genes mexB and mexD were approximately equal to susceptible strains (P > 0.05). Expression of mexY in resistant isolates, except for strain TL-2997, was increased (ranging from 3.414- to 5.022-fold), and the difference was statistically significant (P < 0.05) (Figure 4).
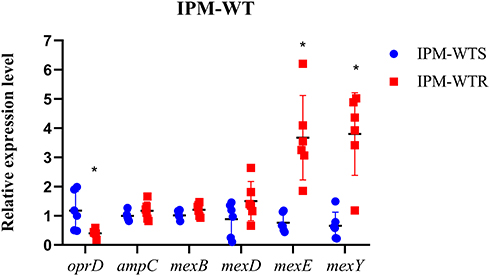 |
Figure 4 Expression levels of genes in wild-type susceptible and resistant isolates. IPM-WTS, wild-type imipenem-susceptible strains; IPM-WTR, wild-type imipenem-resistant strains; PAO1 was served as standardized strain *P < 0.05. |
Analysis of Outer Membrane Proteins
Further analysis of outer membrane proteins by SDS-PAGE was performed. The results of expression of an approximately 46 kDa OprD protein are shown in supplementary materials Figure S2. It could be seen that heteroresistant isolate TL-2856 and TL-2870 did not show an OprD band; the OprD porin band in other heteroresistant and resistant isolates were weaker than PAO1 and susceptible strains TL-2892.
Discussion
Heteroresistance is not detected generally by automated susceptibility assays or the official MIC breakpoint-based criterion.25 Currently, PAP test is the most commonly used method for assessing heteroresistance.26 According to previous reports, heteroresistance is in an intermediate phase, ie, a transition from susceptibility to full resistance under certain conditions.27,28 So far, many studies have analyzed the levels of heteroresistance and its association with patients’ treatment and clinical outcomes.5,11,17 Still, the exact underlining mechanism of heteroresistance remains poorly understood.
In this study, the imipenem-heteroresistant resistance rate was 35.1% in P. aeruginosa. Although the rate was lower than previously reported in Chongqing, China (54.3%),17 the imipenem-heteroresistant P. aeruginosa should be carefully considered at our hospital.
Mutations of oprD have a crucial role in imipenem-resistant P. aeruginosa.21,29,30 According to previous studies, mutational inactivation of OprD was primarily responsible for the resistance to imipenem, and was reported to be the most common mechanism in China and Korea.31,32 Recently, a prospective observational study reported that mortality rate caused by OprD-defective P. aeruginosa was higher than that caused by OprD-intact P. aeruginosa in bloodstream infections.33 These results further disclosed that OprD is essential in the process of resistance to imipenem in P. aeruginosa. Through oprD sequence analysis of 24 isolates, we found two 3 bp deletions at nt 1116–1118 and nt 1147–1149 in three heteroresistant strains (TL-2856, TL-2872, and TL-2877), one resistant isolate TL-2997, and some susceptible strains. Previous studies on mutated oprD located in the external loop7 (oprD-Group 1A Allele with deletion of short stretches of amino acids in loop7) reported no effect on the OprD assembly into outer membranes,34,35 thus suggesting that these substitutions may not be related to imipenem resistance. Nevertheless, a more recent study reported that a specific shortening in loop 7 might likely induce a formation of carbapenem-resistant P. aeruginosa.30 We believe that the deletion of two 3 bp at nt 1116–1118 and 1147–1149 has an important role in the development of heteroresistance and resistance in P. aeruginosa, thus, should carefully considered when assessing drug resistance. Interestingly, in the present study, a deficient oprD was identified in our heteroresistant strains. Amino acid substitutions (eg T103S and V189T) were detected in our (hetero) resistant and susceptible strains, which were also reported in other studies, thus suggesting that these substitutions were not directly involved in imipenem resistance. Certainly, these mutations were located in the external loops of OprD that were responsible for the binding of carbapenems;21,30,34,36,37 nonetheless, the effect of these amino-acid substitutions on the development of resistance to imipenem needs to be further elucidated. Simultaneously, we found that OprD premature stop translation occurred in all resistant isolates. However, this diversification only appeared in two heteroresistant isolates (TL-2870 and TL-2873), implying that the premature stop translation of OprD may be more commonly observed in resistant isolates than heteroresistant isolates.
Inconsistent with He et al study,17 the levels of biofilm formation were not increased in both (hetero) resistant and susceptible strains, except for resistant subpopulation of TL-2856, which indicated that biofilm formation was unrelated to resistance or heteroresistance in our clinical tested isolates. In addition, no strains produced carbapenemases, which suggested that carbapenemases were not contributing to resistance in P. aeruginosa. Consequently, the detection of carbapenemases was essential in heteroresistance and resistance in our clinical setting. The susceptibility results indicated that MICs of heteroresistance to imipenem were slightly changed in other classes of antibiotics. Therefore, the combination of imipenem and other antibiotics may be more optimal against the heteroresistant strains to imipenem compared to imipenem alone.
Previous studies have suggested that oprD, ampC and efflux pump encoding genes (mexB, mexD, mexE and mexY) may regulate the mechanisms of imipenem heteroresistance and resistance in P. aeruginosa.12,17,18 Consistent with data from previous reports, heteroresistant subpopulations expressed lower levels of oprD compared with the respective parental isolates in this study. A correlation between mexE and mexY was observed in imipenem heteroresistant and meropenem non-resistant isolates, and was consistent with previous reports.17,18 Conversely, the current results revealed that ampC, mexB and mexD might not be associated with imipenem heteroresistance and resistance. Similar results were seen in the susceptible and resistant groups. In the present study, there was no significant difference in the mechanisms between imipenem resistance and heteroresistant P. aeruginosa. This phenomenon may be due to different resistance patterns and external stimuli such as imipenem or other antibacterial drug pressure.
Notably, as the results of SDS-PAGE, we found that heteroresistant subpopulation of TL-2873 was a mutant with down-regulated or deficient OprD protein. However, the transcription level of oprD was not down-regulated in this isolate. It is possible that the discrepancy among OprD protein and oprD transcription levels occurred due to the existence of post-transcriptional regulatory pathways.38
Interestingly, a fragment of mutL of heteroresistant strain TL-2856 was sequenced and had a mutation (S515G) which may be related to hypermutator phenotype.
Conclusion
Our data suggested that oprD mutations may regulate heteroresistance of P. aeruginosa to imipenem. Decreased oprD expression and mutations of oprD contributed to imipenem heteroresistance and resistance mechanisms of P. aeruginosa isolates from Wenzhou, China. In addition, marginal up-regulation of efflux pump encoding genes mexE and mexY might play a role in imipenem heteroresistance; yet, future studies are necessary to further verify these findings. Comparative results consistently revealed that resistance and heteroresistance might be mainly triggered by decreased and mutant oprD in P. aeruginosa. Indeed, the difference in resistance mechanisms was prevalent between them in which resistance may also be mainly caused by premature stop translation of oprD. The high rate of heteroresistance should be worth our attention.
Ethical Statement
No samples were specifically collected for this research; anonymized clinical residual samples collected during routine hospital procedures were used for this study.
Acknowledgments
The Health Department of Zhejiang Province of the People’s Republic of China (no. 2011KYA106) supported this study.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Oikonomou O, Panopoulou M, Ikonomidis A. Investigation of carbapenem heteroresistance among different sequence types of Pseudomonas aeruginosa clinical isolates reveals further diversity. J Med Microbiol. 2011;60:1556–1558. doi:10.1099/jmm.0.032276-0
2. Quale J, Bratu S, Gupta J, et al. Interplay of efflux system, ampC, and oprD expression in carbapenem resistance of Pseudomonas aeruginosa clinical isolates. Antimicrob Agents Chemother. 2006;50:1633–1641. doi:10.1128/aac.50.5.1633-1641.2006
3. El-Domany RA, Emara M, El-Magd MA, et al. Emergence of imipenem-resistant Pseudomonas aeruginosa clinical isolates from egypt coharboring VIM and IMP carbapenemases. Microbial Drug Resist. 2017;23:682–686. doi:10.1089/mdr.2016.0234
4. Alexander HE. Mode of action of streptomycin on type B Hemophilus influenzae. Am J Dis Child. 1948;75:428–430.
5. Nicoloff H, Hjort K, Levin BR, et al. The high prevalence of antibiotic heteroresistance in pathogenic bacteria is mainly caused by gene amplification. Nat Microbiol. 2019;4:504–514. doi:10.1038/s41564-018-0342-0
6. Band VI, Weiss DS. Heteroresistance: a cause of unexplained antibiotic treatment failure? PLoS Pathog. 2019;15:e1007726. doi:10.1371/journal.ppat.1007726
7. El-Halfawy OM, Valvano MA. Antimicrobial heteroresistance: an emerging field in need of clarity. Clin Microbiol Rev. 2015;28:191–207. doi:10.1128/cmr.00058-14
8. Fernandez Cuenca F, Sanchez Mdel C, Caballero-Moyano FJ, et al. Prevalence and analysis of microbiological factors associated with phenotypic heterogeneous resistance to carbapenems in Acinetobacter baumannii. Int J Antimicrob Agents. 2012;39:472–477. doi:10.1016/j.ijantimicag.2012.01.015
9. da Silva AEB, Martins AF, Nodari CS, et al. Carbapenem-heteroresistance among isolates of the Enterobacter cloacae complex: is it a real concern? Eur J Clin Microbiol Infect Dis. 2018;37:185–186. doi:10.1007/s10096-017-3138-x
10. Tato M, Morosini M, Garcia L, et al. Carbapenem heteroresistance in VIM-1-producing Klebsiella pneumoniae isolates belonging to the same clone: consequences for routine susceptibility testing. J Clin Microbiol. 2010;48:4089–4093. doi:10.1128/jcm.01130-10
11. Sun JD, Huang SF, Yang SS, et al. Impact of carbapenem heteroresistance among clinical isolates of invasive Escherichia coli in Chongqing, southwestern China. Clin Microbiol Infect. 2015;21:
12. Mei S, Gao Y, Zhu C, et al. Research of the heteroresistance of Pseudomonas aeruginosa to imipenem. Int J Clin Exp Med. 2015;8:6129–6132.
13. Band VI, Crispell EK, Napier BA, et al. Antibiotic failure mediated by a resistant subpopulation in Enterobacter cloacae. Nat Microbiol. 2016;1:16053. doi:10.1038/nmicrobiol.2016.53
14. Lee HY, Chen CL, Wang SB, et al. Imipenem heteroresistance induced by imipenem in multidrug-resistant Acinetobacter baumannii: mechanism and clinical implications. Int J Antimicrob Agents. 2011;37:302–308. doi:10.1016/j.ijantimicag.2010.12.015
15. Lister PD, Wolter DJ, Hanson ND. Antibacterial-resistant Pseudomonas aeruginosa: clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin Microbiol Rev. 2009;22:582–610. doi:10.1128/cmr.00040-09
16. Kohler T, Michea-Hamzehpour M, Epp SF, et al. Carbapenem activities against Pseudomonas aeruginosa: respective contributions of OprD and efflux systems. Antimicrob Agents Chemother. 1999;43:424–427. doi:10.1128/AAC.43.2.424
17. He J, Jia X, Yang S, et al. Heteroresistance to carbapenems in invasive Pseudomonas aeruginosa infections. Int J Antimicrob Agents. 2018;51:413–421. doi:10.1016/j.ijantimicag.2017.10.014
18. Ikonomidis A, Tsakris A, Kantzanou M, et al. Efflux system overexpression and decreased OprD contribute to the carbapenem heterogeneity in Pseudomonas aeruginosa. FEMS Microbiol Lett. 2008;279:36–39. doi:10.1111/j.1574-6968.2007.00997.x
19. CLSI. Performance Standards for Antimicrobial Susceptibility Testing.
20. Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33:2233–2239. doi:10.1128/JCM.33.9.2233-2239.1995
21. Yin S, Chen P, You B, et al. Molecular typing and carbapenem resistance mechanisms of Pseudomonas aeruginosa Isolated from a Chinese burn center from 2011 to 2016. Front Microbiol. 2018;9:1135. doi:10.3389/fmicb.2018.01135
22. Singh M, Yau YCW, Wang S, et al. MexXY efflux pump overexpression and aminoglycoside resistance in cystic fibrosis isolates of Pseudomonas aeruginosa from chronic infections. Can J Microbiol. 2017;63:929–938. doi:10.1139/cjm-2017-0380
23. Tomas M, Doumith M, Warner M, et al. Efflux pumps, OprD porin, AmpC beta-lactamase, and multiresistance in Pseudomonas aeruginosa isolates from cystic fibrosis patients. Antimicrob Agents Chemother. 2010;54:2219–2224. doi:10.1128/aac.00816-09
24. Rossi Goncalves I, Dantas RCC, Ferreira ML, et al. Carbapenem-resistant Pseudomonas aeruginosa: association with virulence genes and biofilm formation. Braz J Microbiol. 2017;48:211–217. doi:10.1016/j.bjm.2016.11.004
25. Nielsen EI, Friberg LE. Pharmacokinetic-pharmacodynamic modeling of antibacterial drugs. Pharmacol Rev. 2013;65:1053–1090. doi:10.1124/pr.111.005769
26. Andersson DI, Nicoloff H, Hjort K. Mechanisms and clinical relevance of bacterial heteroresistance. Nat Rev Microbiol. 2019;17:479–496. doi:10.1038/s41579-019-0218-1
27. Lin J, Xu C, Fang R, et al. Resistance and heteroresistance to colistin in Pseudomonas aeruginosa isolates from Wenzhou, China. Antimicrob Agents Chemother. 2019;63. doi:10.1128/aac.00556-19
28. Superti SV, Martins Dde S, Caierao J, et al. Indications of carbapenem resistance evolution through heteroresistance as an intermediate stage in Acinetobacter baumannii after carbapenem administration. Rev Inst Med Trop Sao Paulo. 2009;51:111–113. doi:10.1590/s0036-46652009000200010
29. Rodriguez-Martinez JM, Poirel L, Nordmann P. Molecular epidemiology and mechanisms of carbapenem resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2009;53:4783–4788. doi:10.1128/aac.00574-09
30. Shu JC, Kuo AJ, Su LH, et al. Development of carbapenem resistance in Pseudomonas aeruginosa is associated with OprD polymorphisms, particularly the amino acid substitution at codon 170. J Antimicrob Chemother. 2017;72:2489–2495. doi:10.1093/jac/dkx158
31. Fang ZL, Zhang LY, Huang YM, et al. OprD mutations and inactivation in imipenem-resistant Pseudomonas aeruginosa isolates from China. Infect Genet Evol. 2014;21:124–128. doi:10.1016/j.meegid.2013.10.027
32. Kim CH, Kang HY, Kim BR, et al. Mutational inactivation of OprD in carbapenem-resistant Pseudomonas aeruginosa isolates from Korean hospitals. J Microbiol. 2016;54:44–49. doi:10.1007/s12275-016-5562-5
33. Yoon EJ, Kim D, Lee H, et al. Mortality dynamics of Pseudomonas aeruginosa bloodstream infections and the influence of defective OprD on mortality: prospective observational study. J Antimicrob Chemother. 2019;74:2774–2783. doi:10.1093/jac/dkz245
34. Shu JC, Su LH, Chiu CH, et al. Reduced production of OprM may promote oprD mutations and lead to imipenem resistance in Pseudomonas aeruginosa carrying an oprD-group 1A allele. Microbial Drug Resist. 2015;21:149–157. doi:10.1089/mdr.2014.0116
35. Huang H, Jeanteur D, Pattus F, et al. Membrane topology and site-specific mutagenesis of Pseudomonas aeruginosa porin OprD. Mol Microbiol. 1995;16:931–941. doi:10.1111/j.1365-2958.1995.tb02319.x
36. Li H, Luo YF, Williams BJ, et al. Structure and function of OprD protein in Pseudomonas aeruginosa: from antibiotic resistance to novel therapies. Int J Med Microbiol. 2012;302:63–68. doi:10.1016/j.ijmm.2011.10.001
37. Rojo-Bezares B, Estepa V, Cebollada R, et al. Carbapenem-resistant Pseudomonas aeruginosa strains from a Spanish hospital: characterization of metallo-beta-lactamases, porin OprD and integrons. Int J Med Microbiol. 2014;304:405–414. doi:10.1016/j.ijmm.2014.01.001
38. Kohler T, Michea-Hamzehpour M, Henze U, et al. Characterization of MexE-MexF-OprN, a positively regulated multidrug efflux system of Pseudomonas aeruginosa. Mol Microbiol. 1997;23:345–354. doi:10.1046/j.1365-2958.1997.2281594.x
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.