")
Back to Journals » Drug Design, Development and Therapy » Volume 12
Lycium barbarum polysaccharide prevents cisplatin-induced MLTC-1 cell apoptosis and autophagy via regulating endoplasmic reticulum stress pathway
Authors Yang F, Wei Y, Liao B, Wei G, Qin H, Pang X, Wang J
Received 5 June 2018
Accepted for publication 1 August 2018
Published 26 September 2018 Volume 2018:12 Pages 3211—3219
DOI https://doi.org/10.2147/DDDT.S176316
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Qiongyu Guo
This paper has been retracted.
Fenglian Yang,1 Yuxia Wei,2 Biyun Liao,2 Guijiang Wei,2 Haimei Qin,2 Xiaoxia Pang,2 Junli Wang2
1Department of Reproductive Medicine, Youjiang Medical College for Nationalities, Baise, Guangxi 533000, China; 2Department of Reproductive Medicine, Affiliated Hospital of Youjiang Medical College for Nationalities, Baise, Guangxi 533000, China
Background: Lycium barbarum polysaccharide (LBP) has been reported to contribute to the recovery of male hypogonadism and infertility.
Aim: The aim of current study was to investigate the underlying mechanisms of LBP on male infertility recovery.
Methods: Recently, it is reported that cell apoptosis mediated by endoplasmic reticulum stress (ERS) was distinguished from that mediated by death reporters and mitochondria pathway, which could induce cell apoptosis independently. The possible signaling mechanisms were investigated using diversified molecular biology techniques, such as flow cytometry, western blotting, and immunofluorescence.
Results: In this study, we found that LBP protected Leydig MLTC-1 cells against cisplatin (DDP) by regulating ERS-mediated signal pathway, which was evidenced by downregulation of phosphorylation PERK, phosphorylation of eukaryotic translation-initiation factor 2α and activating transcription factor 4. Meanwhile, LBP decreased DDP-induced MLTC-1 cell apoptosis via reducing ERS apoptosis-relative proteins caspase 3, caspase 7, and caspase 12. In addition, the result of monodansylcadaverine staining indicated that LBP significantly inhibited DDP-induced autophagosome formation in MLTC-1 cells. Moreover, immunofluorescences and Western blot assays demonstrated that LBP reversed DDP-induced LC3II and Atg5 upregulation in MLTC-1 cells. Finally, the data of enzyme-linked immunosorbent assay showed that LBP markedly recovered MLTC-1 cells testosterone level even in the presence of DDP.
Conclusion: Thus, we suggest that LBP protected MLTC-1 cells against DDP via regulation of ERS-mediated apoptosis and autophagy.
Keywords: L. barbarum polysaccharide, endoplasmic reticulum stress, hypogonadism, DDP, MLTC-1 cell, autophagy, apoptosis
Introduction
In the recent years, approximately 6% of males are infected with male hypogonadism with its main features being testosterone deficiency and relevant clinical features. This trend is becoming more general and the incidence is increased.1 Possibly, the main causes of male infertility are reproductive tract infection and inflammation.2 In addition, hypogonadism can lead to a variety of diseases, such as sexual dysfunction, testicular failure, sleep disturbance, abdominal adiposity, and so on.1,3 Along with increasing age, Leydig cell number and function will decline. Autocrine androgen action in Leydig cells is essential for all men’s life, which plays a vital role in spermatogenesis and lifelong health of Leydig cells.4 Testosterone is a main circulating androgen predominantly coming from Leydig cells in the gap space of the testes in mammals, which is vital for adjustment of different crucial processes such as sexual differentiation, sustained spermatogenesis, and reproductive function.3,5 Therefore, enhancing the males’ sperm energy and improving the hypogonadism are very important for male fertility, general health, and well-being.1,6
Lycium barbarum (L. barbarum) is a well-known, traditional Chinese medicinal herb, which has the homology of medicine and food.7 The fruit of L. barbarum has been used for nourishing liver and kidney and improving eyesight.8 The main active ingredient in L. barbarum is L. barbarum polysaccharide (LBP), which is composed of arabinose, glucose, galactose, mannose, xylose, and rhamnose.8 LBP has the function of reproductive system protection, which can be achieved by increasing antioxidant enzyme activities and suppressing cell death to mediated male spermatogenic dysfunction induced by diabetes.7 Moreover, it was found that LBP enormously increased the sperm count and vitality and enhanced the sexual function of male rats damaged by 60Co-γ irradiation.9 LBP also have a protective role on spermatogenesis of rats with reproductive system damages induced by cyclophosphamide.9
Endoplasmic reticulum (ER) is an important organelle that is related to various cellular processes including protein synthesis, secreting, folding, and assemble. However, calcium ion homeostasis destruction, protein unfoldable, misfolded, and accumulated in ER can induce endoplasmic reticulum stress (ERS).10,11 Thereafter, serious ERS could induce apoptosis. In order to alleviate the apoptosis of the cells, the subsequent response of this stimulation is called the unfold-protein reaction (UPR).12 UPR activates three transcription factors in mammalian cells: IRE1/ERN1 (inositol requiring 1), PERK/PEK (PEK like ER kinase), and ATF4 (activating transcription factor 4).13 PERK is an ER transmembrane protein kinase that inhibits protein translation via phosphorylating eukaryotic translation-initiation factor 2α subunit. This is a key factor for activating apoptosis in response to ERS, accompanied with enhancer-binding protein homologous protein (CHOP) upregulation.10,14,15
Autophagy is a lysosomal degradation pathway intracellular, and can eliminate organelles and proteins, which was regulated by mTOR pathway and autophagy-related (ATG) family members.16 Until now, several types of autophagy have been identified, such as mitophagy, microautophagy, macroautophagy, and chaperone-mediated autophagy.17,18 They all have autophagy-related protein (ATG) components and these components have almost identical set.19,20 All these different types of autophagy play vital roles in the different steps of autophagy.19,20
Although so many valuable studies about the biological activities and pharmacological functions of LBP have been published, there is scarcity of information concerning the protective effect of LBP on Leydig MLTC-1 cells. In this study, we aim to investigate the underlying mechanisms of how LBP prevents DDP-induced MLTC-1 cell apoptosis and autophagy.
Materials and methods
Cell culture
The mouse Leydig MLTC-1 cell line was acquired from Stem Cell Bank of Chinese Academy of Sciences (Shanghai, China) and were cultured in RPMI-1640 medium (Sigma, St Louis, MO, USA) with 10% fetal bovine serum (Thermo Fisher Scientific, Waltham, MA, USA), penicillin (100 U/mL, Sigma) and streptomycin (100 μg/mL, Sigma). Cells were grown at 37°C with 5% CO2 in a constant temperature cell culture incubator (Sanyo, Tokyo, Japan). LBP was a gift from Professor Lu.21
CCK-8 assay of cell viability
The CCK-8 assay (Dojindo, Kumamoto, Japan) was used to determine cell growth according to the protocol of manufacturer for indicated time points following treatment with LBP with or without DDP in MLTC-1 cells. Briefly, MLTC-1 cells were seeded into 96-well plate at the density of 5×103 cells/well overnight. Two hundred microliters of LBP (0, 25, 50, 100 μg/mL) with or without DDP (10 μM) were added into each well in triplicate when cells were totally adhered.
EdU detection and autophagosome formation
MLTC-1 cells were exposed to LBP (50 μg/mL) and/or DDP (10 μM) for 48 hours, and EdU detection was tested according to the method described in the EdU kit (Thermo Fisher, C10638). Immunofluorescence assay was performed according to the previously described method.22 Briefly, cells were incubated with primary antibodies for DAPI, EdU, or LC3 (Cell Signaling, Danvers, MA, USA; 1:200) at 4°C overnight and were incubated with Alexa 488-conjugated donkey anti-mouse secondary antibodies (Jackson Immuno Research, West Grove, PA, USA) at 37°C for 1 hour. EdU-positive cells and autophagosome formation were observed by fluorescence microscopy (Olympus Corporation, Tokyo, Japan).
Western blot analysis
Following treatment with LBP (50 μg/mL) and/or DDP (10 μM) for 48 hours, MLTC-1 cells were collected and lysed in RIPA buffer (Sigma). The extracted protein concentration was quantified using a BCA Protein Assay Kit (Beyotime, Shanghai, China) and separated on SDS-PAGE gels with 10%Tris-SDS gel. After the electrophoresis, the gel was transferred onto PVDF membranes (Thermo Fisher Scientific). The PVDF membrane was blocked in 5% defatted milk in TBST for 50 minutes and then probed with primary antibodies overnight at 4°C. The following primary antibodies were used: anti-PERK (Cell Signaling; 1:1,000), anti-p-PERK (Cell Signaling; 1:1,000), anti-p-elF2α (Cell Signaling; 1:1,000), anti-elF2α (Cell Signaling; 1:1,000), anti-ATF4 (Cell Signaling; 1:1,000), and anti-β-actin (Santa Cruz Biotechnology; 1:1,000), active caspase-3 (Cell Signaling; 1:1,000), active caspase-7 (Cell Signaling; 1:1,000), caspase-12 (Cell Signaling; 1:1,000), LC3 (Cell Signaling; 1:1,000), and Atg 5 (Cell Signaling; 1:1,000). Immunodetection was carried out using anti-rabbit (1:5,000) or anti-mouse (1:5,000) secondary antibodies and enhanced chemiluminescence detection kit (Thermo Fisher Scientific).23
Flow cytometric analysis of cell apoptosis
Apoptotic cells were analyzed according to the formerly described method.24 Briefly, MLTC-1 cells were exposed to LBP (50 μg/mL) and/or DDP (10 μM) for 48 hours. Subsequently, these three groups of MLTC-1 cells were collected and resuspended in PBS. Apoptotic cells were recognized with dual-staining of Annexin V-FITC and propidium iodide (PI; Thermo Fisher Scientific). For each group, the experiments were repeated in triplicate.
Monodansylcadaverine staining
MLTC-1 cells treated with LBP (50 μg/mL) and/or DDP (10 μM) at 37°C for 48 hours were stained with monodansylcadaverine (MDC; 50 μM) at 37°C for 30 minutes. After incubation, the cells were washed three times with PBS, and treated with 50 μM MDC in an incubator for 15 minutes. Autophagic vacuoles were immediately observed with a fluorescence microscope (Olympus Corporation) and quantified.25
Hormone assays
MLTC-1 cells were seeded in six-well plates at 2×105/well and treated with LBP (50 μg/mL) and/or DDP (10 μM) at 37°C for 12 hours. Subsequently, the culture medium was collected, and testosterone was analyzed by an enzyme-linked immunosorbent assay (ELISA) kit (R&D, Minneapolis, MN, USA).23
Statistical analysis
All experiments were performed at least three times and all data were expressed as the mean ± SD. One-way ANOVA followed by Dunnett’s test or least significance difference was used to analyze the difference between groups with P<0.05 being regarded as a statistically significant difference (*P<0.05, **P<0.01).
Results
LBP reversed DDP-induced MLTC-1 cell growth inhibition
The viability of MLTC-1 cells were detected at indicated time points with LBP dose escalation (0, 25, 50, 100 μg/mL) with or without DDP (10 μM) using the CCK-8 assay. As shown in Figure 1A, 10 μM DDP inhibited MLTC-1 cell proliferation significantly, while 50 μg/mL LBP promoted MLTC-1 cell proliferation. When the MLTC-1 cells were treated with LBP (50 μg/mL) plus DDP, the cell viability was significantly increased compared with DDP alone treatment (Figure 1A). Additionally, as indicated in Figure 1B, the quantity of MLTC-1 cells in LBP plus DDP group was much more than DDP alone group.
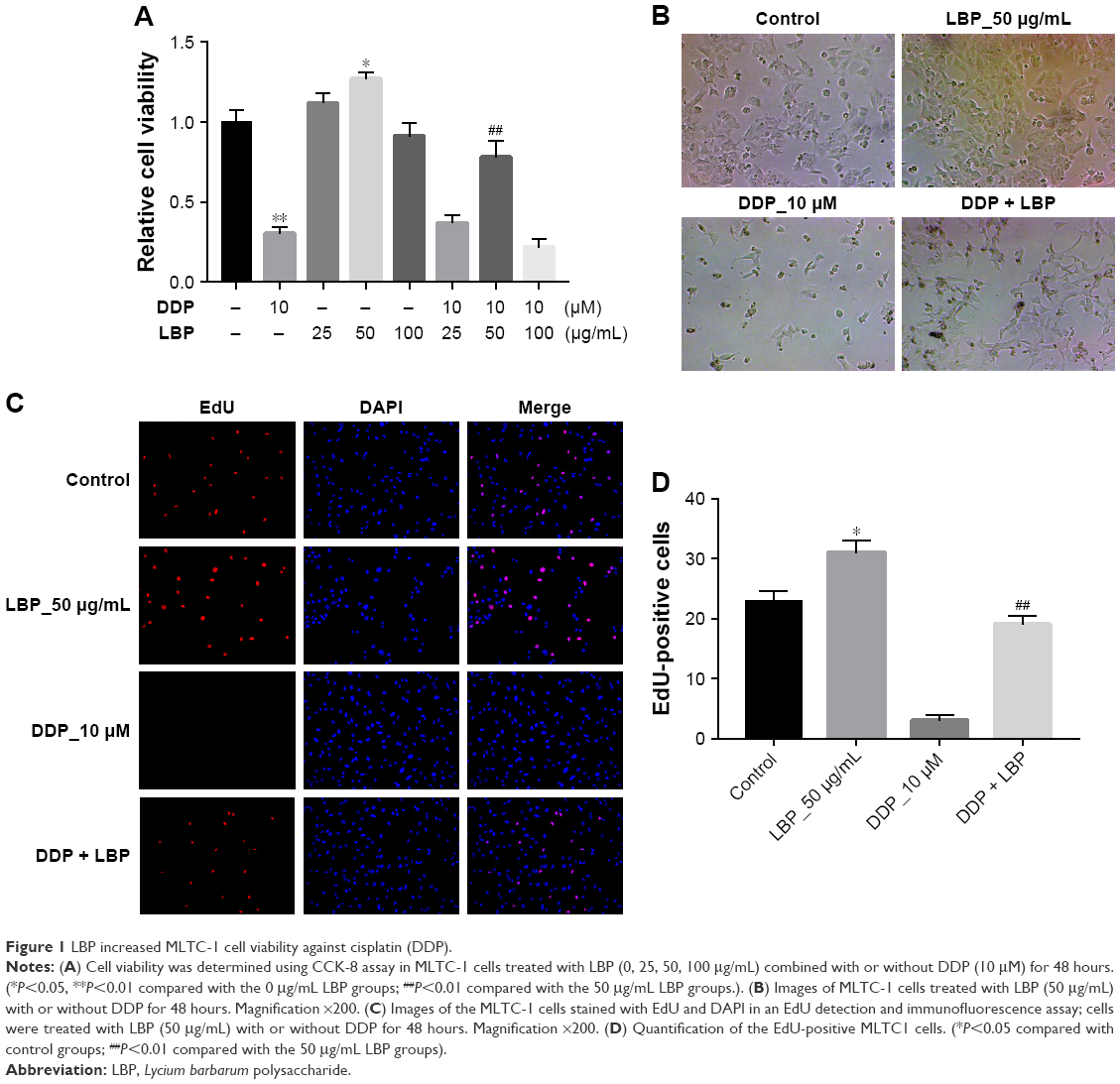 | Figure 1 LBP increased MLTC-1 cell viability against cisplatin (DDP). |
EdU is efficiently mixed with newly synthesized DNA and fluorescently labeled, which is an important index to evaluate cell activity. The EdU fluorescence assay revealed that the DDP could significantly decrease the EdU-positive cells, while LBP could dramatically increase the EdU-positive cells in DNA synthesis phase (Figure 1C and D). Meanwhile, LBP protected MLTC-1 cells against DDP, which was evidenced with the EdU-positive cells significantly increased in LBP plus DDP group compared with DDP alone group (Figure 1C and D). These results suggested that LBP could reverse DDP-induced cytotoxicity and increase MLTC-1 cell vitality.
LBP inhibited DDP-induced ERS-mediated proteins upregulation in MLTC-1 cells
To further demonstrate the effect of LBP on ERS-mediated cell apoptotic pathway, MLTC-1 cells were stimulated with LBP and/or DDP for 48 hours and harvested for western blotting analysis. The results showed that 50 μg/mL LBP alone could significantly decrease the proteins expressions of p-PERK, p-elF2α, and ATF4, while 10 μM DDP exerted the opposite effect on these proteins (Figure 2A–D). Meanwhile, DDP-induced p-PERK, p-elF2α, and ATF4 upregulation were dramatically reversed by LBP (Figure 2A–D). These results suggested that LBP inhibited DDP-induced ERS-mediated proteins upregulation in MLTC-1 cells.
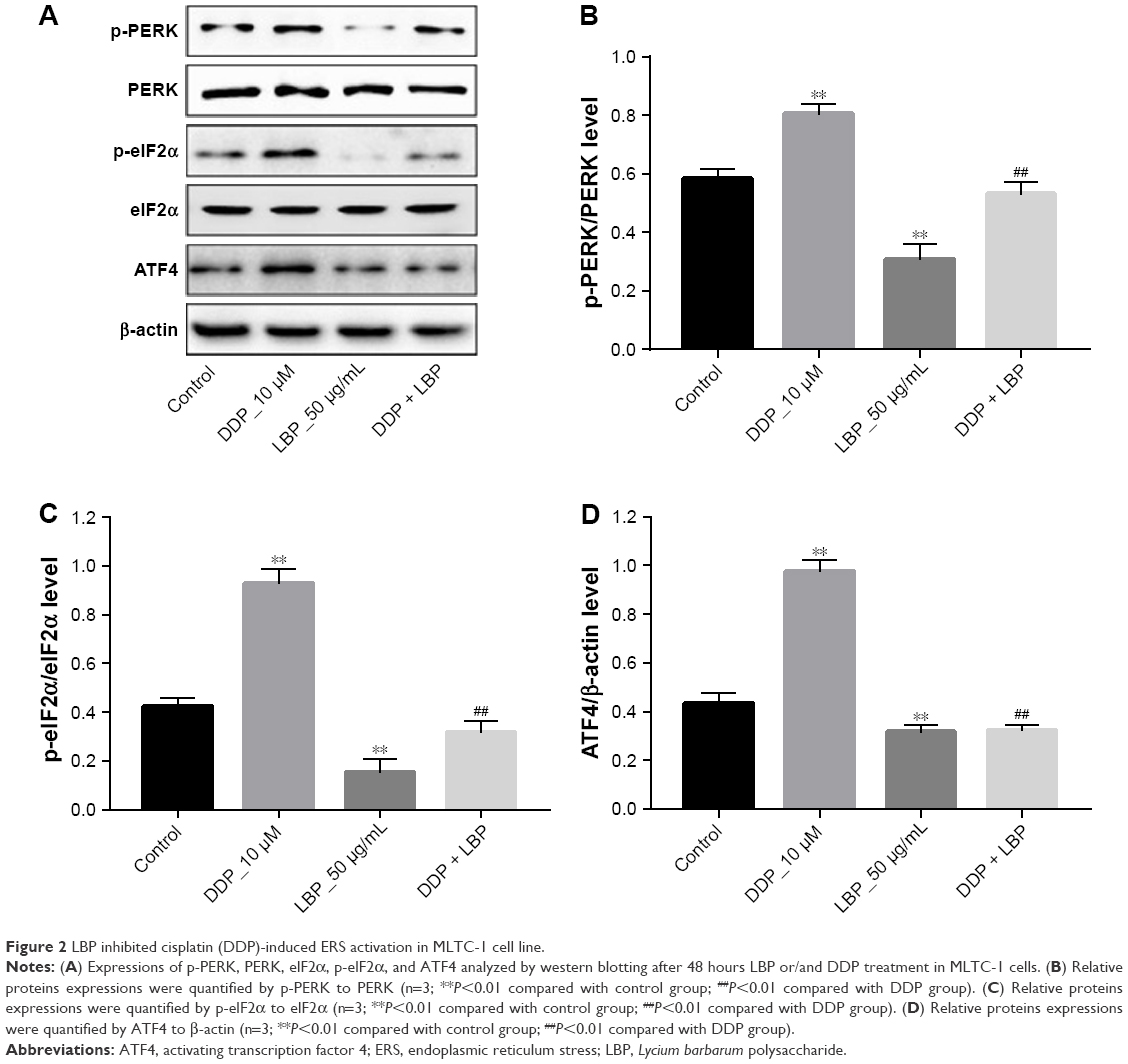 | Figure 2 LBP inhibited cisplatin (DDP)-induced ERS activation in MLTC-1 cell line. |
LBP inhibited DDP-induced ERS-mediated apoptosis in MLTC-1 cells
We next assessed if LBP could decrease ERS-mediated apoptosis induced by DDP in MLTC-1 cells. As shown in Figure 3A and B, 50 μg/mL LBP alone did not trigger any apoptosis in cells. In contrast, 10 μM DDP significantly induced cell apoptosis (31.89%) compared with control group. However, the cell apoptosis rate in LBP plus DDP group was reduced from 31.89% to 12.05% compared with DDP alone group (P<0.01). In addition, the results of Western blot revealed that DDP alone activated caspase 3, caspase 7, and caspase 12 proteins in cells. Similarly, the levels of caspase 3, caspase 7, and caspase 12 in cells were downregulated in the presence of LBP (Figure 3C–F). These data indicated that LBP could reverse DDP-induced ERS-mediated apoptosis in MLTC-1 cells.
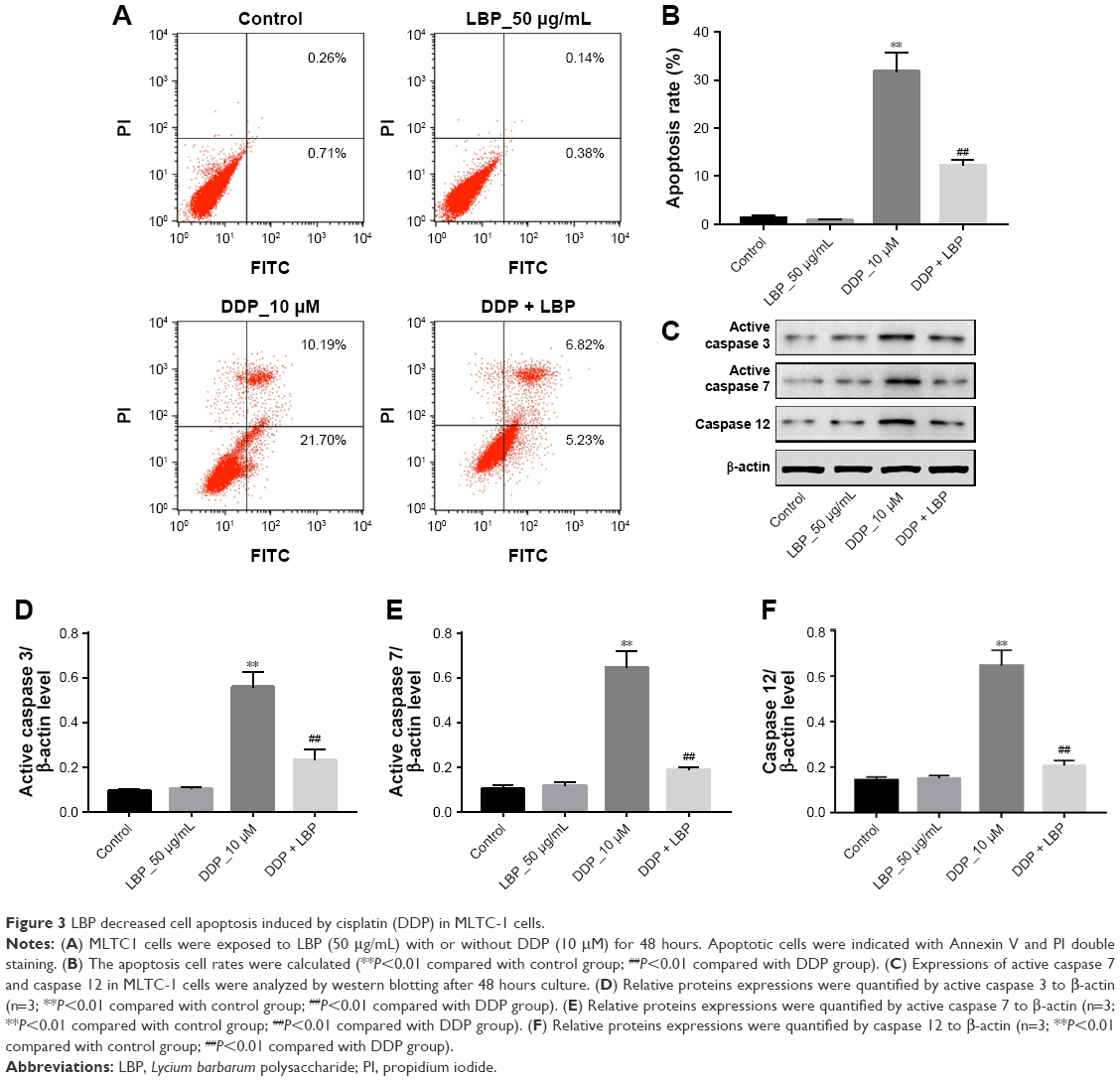 | Figure 3 LBP decreased cell apoptosis induced by cisplatin (DDP) in MLTC-1 cells. |
LBP reversed DDP-induced autophagy in MLTC-1 cells
ERS also activated autophagy-associated proteins LC3II and Atg5 expressions. Thereby, we performed MDC assay in order to further confirm the effect of LBP on autophagic vacuoles formation in cells. As shown in Figure 4A and B, 50 μg/mL LBP had no effect on MLTC-1 cells compared with control group, while 10 μM DDP significantly increased the autophagic vacuoles formation in cells. As expected, MDC-positive cells were significantly decreased by LBP treatment even in the presence of DDP. The immunofluorescence and Western blot data also confirmed that DDP-induced LC3II and Atg5 increases were inhibited by LBP (Figure 4C–F). All these results suggest that DDP induced ERS-mediated autophagy, which was partly reversed by LBP in MLTC-1 cells.
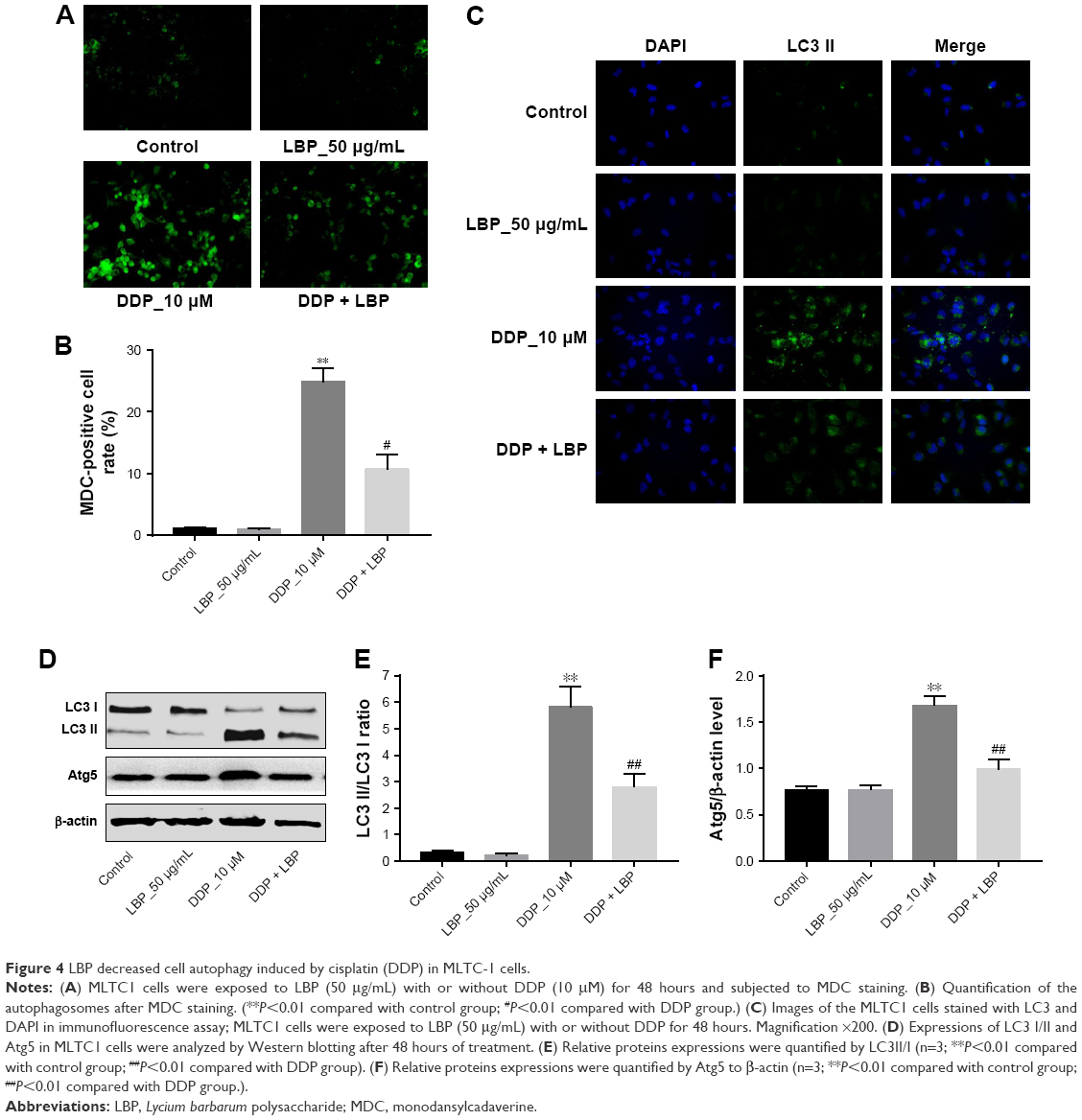 | Figure 4 LBP decreased cell autophagy induced by cisplatin (DDP) in MLTC-1 cells. |
LBP increased the level of testosterone in MLTC-1 cells
To assess the sperm motility effect of LBP on MLTC-1 cells in vitro, we detected the testosterone level using ELISA. As shown in Figure 5, 10 μM DDP significantly decreased the level of testosterone in culture medium, while that was dramatically reversed by LBP. These data demonstrated that LBP could protect the level of testosterone in MLTC-1 cells against DDP.
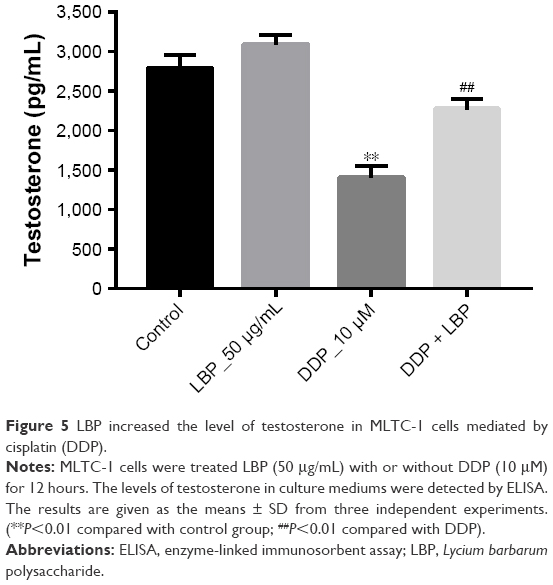 | Figure 5 LBP increased the level of testosterone in MLTC-1 cells mediated by cisplatin (DDP). |
Discussion
LBP has various biological characteristics including improving the male hypogonadism.7,9,26 However, the underlying mechanisms of how LBP regulated ERS-mediated apoptosis and autophagy were not fully illuminated. Our results suggested that 50 μg/mL LBP treatment stimulated cell growth, while 100 μg/mL LBP inhibited cell growth. This condition might be because of the high glucose in cell culture media. In the current study, we demonstrated that 50 μg/mL LBP treatment significantly inhibited DDP-induced MLTC-1 cell apoptosis by flow cytometry analysis with Annexin V-FITC/PI staining. Shi et al also revealed that LBP inhibited streptozotocin-induced apoptosis in diabetic male mice, which is consistent with our results.7 EdU is a kind of thymidine nucleoside, which can insert into DNA molecules during cell reproduction. In the current study, LBP increased MLTC-1 cell vitality via increasing DNA synthesize in S phase.
At the cellular level, we illustrated that pathological ERS activation could be the key signaling mechanism responsible for DDP-induced MLTC-1 cells apoptosis. The hallmarks of ERS, p-PERK, p-elF2α, and ATF4 were all downregulated in LBP-treated MLTC-1 cells even in the presence of DDP. Type I ERS induced cell death and apoptosis accompanied with a series steps of elF2α phosphorylation mediated by PERK and ATF4. In the current study, LBP inhibited DDP-induced MLTC-1 cell apoptosis via downregulation of p-PERK/p-elF2α/ATF4 pathway, which was further validated by cytometry analysis. Shi et al reported that LBP protected male sexual dysfunction and fertility impairments by activating hypothalamic pituitary gonadal axis in streptozotocin-induced type 1 diabetic male mice.23 Consistent with this work, LBP also protected MLTC-1 cells against DDP.
Caspase 7 was major mainly in spermatogenic cells and have apoptotic function in spermatogenic dysfunction.27 In the current study, we found that LBP could reverse DDP-induced caspase 3, caspase 7, and caspase 12 upregulation. In addition, the formation of autophagic vacuoles was detected using immunohistochemistry and MDC staining. We illustrated that the numbers of MDC and autolysosome were increased by DDP alone treated compared with control cells, while LBP could reverse this kind of increases in the cells. Liu et al reported that LBP could protect mTOR-mediated autophagy in diabetic rats; LC3 and p62 were involved; the expression of LC3II was decreased and p62 was increased, indicating a decreased activation of autophagy.28 Similarly, the expressions of LC3II and Atg5 were both downregulated by LBP in MLTC-1 cells even in the presence of DDP in the present study.
It is reported that LBP exerted protective effects on the male spermatogenic dysfunction, and reproductive system of male rats might have a relationship with testosterone level.7,9,26 Here, our data confirmed that LBP also preserve testosterone level against DDP in MLTC-1 cell medium. This finding was consistent with Shi et al.7
In conclusion, we found that the LBP could reverse DDP-induced ERS activation-mediated autophagy and apoptosis in MLTC-1 cells. In addition, LBP effectively protected testosterone production against DDP in MLTC-1 cell medium. Our findings provide a new insight into the protective effect mechanisms of LBP on spermatogenic dysfunction and propose that LBP may be an alternative medicine for the treatment of male infertility and hypogonadism diseases.
Acknowledgment
We would like to thank Dr Ke Wu for careful proofreading. This study was supported by the 2018 Natural Science Foundation of Baise in China (No. 20182511); the Foundation of the Health Department of Guangxi Province, China (No. Z20170234; Z20170236); the “139” Medical High-Level Talent Training Plan and the Thousands of Young and Middle-aged Backbone Teachers Cultivation Plan of Guangxi Province, China; and the 2018–2020 Professional and Experimental Practice Teaching Base Construction Projects of Guangxi Province, China.
Disclosure
The authors report no conflicts of interest in this work.
References
Tremellen K, Mcphee N, Pearce K, Benson S, Schedlowski M, Engler H. Endotoxin-initiated inflammation reduces testosterone production in men of reproductive age. Am J Physiol Endocrinol Metab. 2018;314(3):E206–E213. | ||
Leisegang K, Henkel R. The in vitro modulation of steroidogenesis by inflammatory cytokines and insulin in TM3 Leydig cells. Reprod Biol Endocrinol. 2018;16(1):26. | ||
Dohle GR, Smit M, Weber RF. Androgens and male fertility. World J Urol. 2003;21(5):341–345. | ||
O’Hara L, Mcinnes K, Simitsidellis I, et al. Autocrine androgen action is essential for Leydig cell maturation and function, and protects against late-onset Leydig cell apoptosis in both mice and men. FASEB J. 2015;29(3):894–910. | ||
Basaria S, Hypogonadism M. Male hypogonadism. The Lancet. 2014;383(9924):1250–1263. | ||
Walker WH. Testosterone signaling and the regulation of spermatogenesis. Spermatogenesis. 2011;1(2):116–120. | ||
Shi GJ, Zheng J, Wu J, et al. Beneficial effects of Lycium barbarum polysaccharide on spermatogenesis by improving antioxidant activity and inhibiting apoptosis in streptozotocin-induced diabetic male mice. Food Funct. 2017;8(3):1215–1226. | ||
Cheng J, Zhou ZW, Sheng HP, et al. An evidence-based update on the pharmacological activities and possible molecular targets of Lycium barbarum polysaccharides. Drug Des Devel Ther. 2015;9:33–78. | ||
Luo Q, Cui X, Yan J, et al. Antagonistic effects of Lycium barbarum polysaccharides on the impaired reproductive system of male rats induced by local subchronic exposure to 60Co-γ irradiation. Phytother Res. 2011;25(5):694–701. | ||
Tao YK, Pl Y, Bai YP, et al. Role of PERK/eIF2α/CHOP endoplasmic reticulum stress pathway in oxidized low-density lipoprotein mediated induction of endothelial apoptosis. Biomed Environ Sci. 2016;29(12):868–876. | ||
Hauck AK, Bernlohr DA. Oxidative stress and lipotoxicity. J Lipid Res. 2016;57(11):1976–1986. | ||
Wang X, Eno CO, Altman BJ, et al. ER stress modulates cellular metabolism. Biochem J. 2011;435(1):285–296. | ||
Hetz C, Martinon F, Rodriguez D, Glimcher LH. The unfolded protein response: integrating stress signals through the stress sensor IRE1α. Physiol Rev. 2011;91(4):1219–1243. | ||
Werner N, Nickenig G, Laufs U. Pleiotropic effects of HMG-CoA reductase inhibitors. Basic Res Cardiol. 2002;97(2):105–116. | ||
Porter KE, Turner NA. Statins and myocardial remodelling: cell and molecular pathways. Expert Rev Mol Med. 2011;13:e22. | ||
Ma Y, Zhou Y, Zhu YC, Wang SQ, Ping P, Chen XF. Lipophagy contributes to testosterone biosynthesis in male rat leydig cells. Endocrinology. 2018;159(2):1119–1129. | ||
Kim I, Rodriguez-Enriquez S, Lemasters JJ. Selective degradation of mitochondria by mitophagy. Arch Biochem Biophys. 2007;462(2):245–253. | ||
Singh R, Kaushik S, Wang Y, et al. Autophagy regulates lipid metabolism. Nature. 2009;458(7242):1131–1135. | ||
Ravikumar B, Sarkar S, Davies JE, et al. Regulation of mammalian autophagy in physiology and pathophysiology. Physiol Rev. 2010;90(4):1383–1435. | ||
Mizushima N, Yoshimori T, Ohsumi Y. The role of Atg proteins in autophagosome formation. Annu Rev Cell Dev Biol. 2011;27:107–132. | ||
Lu SP, Zhao PT. Chemical characterization of Lycium barbarum polysaccharides and their reducing myocardial injury in ischemia/reperfusion of rat heart. Int J Biol Macromol. 2010;47(5):681–684. | ||
Wang B, Hendricks DT, Wamunyokoli F, Parker MI. A growth-related oncogene/CXC chemokine receptor 2 autocrine loop contributes to cellular proliferation in esophageal cancer. Cancer Res. 2006;66(6):3071–3077. | ||
Sun D, Cui Y, Jin B, Zhang X, Yang X, Gao C. Effects of the yanjing capsule extract on steroidogenesis and apoptosis in mouse leydig cells. Evid Based Complement Alternat Med. 2012;2012(3):985457:1–10. | ||
Bai F, Ni B, Liu M, Feng Z, Xiong Q, Shao G. Mycoplasma hyopneumoniae-derived lipid-associated membrane proteins induce inflammation and apoptosis in porcine peripheral blood mononuclear cells in vitro. Vet Microbiol. 2015;175(1):58–67. | ||
Zheng Q, Li Z, Zhou S, et al. Heparin-binding hemagglutinin of Mycobacterium tuberculosis is an inhibitor of autophagy. Front Cell Infect Microbiol. 2017;7:33. | ||
Qian L, Yu S. Protective effect of polysaccharides from Lycium barbarum on spermatogenesis of mice with impaired reproduction system induced by cyclophosphamide. Am J Reprod Immunol. 2016;76(5):383–385. | ||
Lei B, Zhou X, Lv D, et al. Apoptotic and nonapoptotic function of caspase 7 in spermatogenesis. Asian J Androl. 2017;19(1):47–51. | ||
Liu SY, Chen L, Li XC, Hu QK, He LJ. Lycium barbarum polysaccharide protects diabetic peripheral neuropathy by enhancing autophagy via mTOR/p70S6K inhibition in Streptozotocin-induced diabetic rats. J Chem Neuroanat. 2018;89:37–42. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.