")
Back to Journals » OncoTargets and Therapy » Volume 13
Low-Dose Radiation Promotes Invasion and Migration of A549 Cells by Activating the CXCL1/NF-κB Signaling Pathway
Authors Li J, Wu D, Han R, Yu Y, Deng S , Liu T , Zhang T, Xu Y
Received 27 December 2019
Accepted for publication 9 April 2020
Published 29 April 2020 Volume 2020:13 Pages 3619—3629
DOI https://doi.org/10.2147/OTT.S243914
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Nicola Silvestris
Jing Li,* Dong-ming Wu,* Rong Han,* Ye Yu, Shi-hua Deng, Teng Liu, Ting Zhang, Ying Xu
Clinical Laboratory, The First Affiliated Hospital, Collaborative Innovation Center of Sichuan for Elderly Care and Health of Chengdu Medical College, Chengdu, Sichuan 610041, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ying Xu
Clinical Laboratory, The First Affiliated Hospital of Chengdu Medical College, 278 Baoguang Road, Chengdu, Sichuan 610500, People’s Republic of China
Tel +86 2883016723
Email [email protected]
Purpose: Radiation has well-known and well-characterized direct toxic effects on cells and tissues. However, low-dose ionizing irradiation (LDIR) can also enhance the invasion and migration of tumor cells, and the mechanisms underlying these effects remain unclear. The present study aimed to investigate changes induced in the migration and invasion of A549 cells after LDIR and to explore the potential molecular mechanism.
Materials and Methods: A549 cells were irradiated with X-rays at different doses (0, 2, 4, and 6 Gy) and cultured for 24 or 48 h. Apoptosis and proliferation were evaluated by lactate dehydrogenase release, CCK8, colony formation, and flow cytometry assays. Wound-healing and transwell assays were performed to detect migration and invasion ability. CXCL1 or p65 were knocked down using lentivirus-mediated siRNA in A549 cell lines. Knockdown efficiency of CXCL1 and p65 was assessed by RT-qPCR. Western blotting and immunofluorescence were used to determine the changes in protein levels.
Results: In cells irradiated with a dose of 6 Gy, after 48 h, apoptosis was clearly induced while proliferation was inhibited. Irradiation with 4 Gy resulted in the upregulation of CXCL1 expression and activation of the NF-κB signaling pathway. Moreover, upon 4 Gy irradiation, migration, invasion, and epithelial–mesenchymal transition (EMT) were significantly enhanced in A549 cells. Importantly, CXCL1 or p65 knockdown inhibited radiation-induced migration, invasion, and EMT.
Conclusion: Low-dose radiation upregulates CXCL1 expression and activates the NF-κB signaling to regulate EMT in A549 cells, thereby promoting invasion and migration. These results provide new insights into the prevention of tumor invasion and metastasis induced by radiotherapy.
Keywords: ionizing radiation, non-small cell lung cancer, CXCL1, EMT
Introduction
Lung cancer is a common malignant tumor that threatens human health. It is characterized by rapid development, high malignancy, and poor prognosis.1 The death rate of lung cancer patients accounts for more than 1/6 of all cancer deaths.2,3 Currently, radiotherapy is still one of the main treatments for patients with lung cancer. Approximately 60% to 70% of patients with lung cancer need to receive radiation therapy, highlighting the importance of this therapeutic approach for patients with inoperable tumors or incompletely resected tumors and for those with recurrent disease.4
Radiotherapy has proven benefits for overall survival based on a number of clinical studies.5 However, some patients present with local recurrence or metastases during treatment.6 Therefore, exploring the related genes and mechanisms underlying the effects of ionizing radiation (IR) is key to clarifying the changes in biological behavior of tumor cells after radiotherapy. When tumor cells are exposed to low doses of X-rays, their proliferation ability is significantly suppressed, while their migration and invasion capabilities are significantly enhanced.7,8 In 1949, Kaplan and Murphy demonstrated this effect in a transplantable mouse model. They observed an increased rate of lung metastasis in tumor-bearing mice treated locally with low-dose radiation compared to that in sham-irradiated mice.9 Tumor invasion and metastasis is a complex, dynamic process involving the interactions among multiple genes and their products.9 Chemokines have been shown to not only play an important role in pathological processes such as inflammation, autoimmune diseases, and radiation damage, but also participate in tumor formation and metastasis.10 The chemokine CXCL1 (chemokine CXC motif ligand 1) belongs to the CXC subfamily.11 CXCL1 is related to the inflammatory response and directly affects the growth, invasion, and metastasis of certain tumors.12,13 In vitro studies have shown that CXCL1 can promote the invasion and metastasis of malignant tumors such as melanoma, bladder cancer, and gastric cancer.13,14 However, whether CXCL1 plays a role in the invasion and metastasis of IR-induced tumors is unclear. Herein, we demonstrated for the first time in vitro that radiation can regulate epithelial–mesenchymal transition (EMT) in tumor cells by upregulating CXCL1 expression and activating the NF-κB signaling, thereby enhancing tumor cell invasion and migration.
Materials and Methods
Antibodies
Antibodies against CXCL1 (12335-1-AP), GAPDH (66004-1-Ig), E-cadherin (20874-1-AP), vimentin (10366-1-AP), and N-cadherin (22018-1-AP) and secondary antibodies were obtained from Proteintech Group, Inc. (Wuhan, China); antibodies against p65 (D14E12) and phospho-p65 (Ser536) were purchased from Zenbio (Chengdu, China). All other kits and reagents were purchased from the Beyotime Institute of Biotechnology (Shanghai, China).
Cell Culture and Transfections
The non-small cell lung cancer cell line A549 (CVCL_0023) was obtained from Cell Bank of the Chinese Academy of Sciences (Shanghai, China). And the cell was maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum, 10 mM
X-Ray Radiation
A549 cells were grown as a monolayer and exposed to an X-RAD 160–225 instrument (Precision X-Ray, Inc., Branford, CT, USA; filter: 2 mm AI; 42 cm, 225 kV/s, 12.4 mA, 2.0 Gy/min) to attain the desired dose of 2, 4, or 6 Gy. Cell viability, apoptosis, and other assays were detected after irradiation for 24 or 48 h.
Cell Viability Assay
Cell viability was assessed by cell counting kit-8 (CCK-8) and colony formation assays. Cells were plated at 500 cells/well in a 6-well plate (Corning, Corning, NY, USA) after irradiation with the desired doses (0, 2, 4, or 6 Gy). Cells were cultured for 7 d with medium changes every 3 d, washed twice with PBS, fixed in methanol, and stained with 1% crystal violet. Ten thousand cells per well were seeded into 96-well plates incubated for 12 h as described previously herein. Then, the cells were exposed to an X-RAD 160–225 instrument to attain the desired dose of 2, 4, or 6 Gy and the cell viability was measured with a Cell Counting Kit-8 after 24 or 48 h.
Lactate Dehydrogenase (LDH) Release Assay
Cell viability was measured with the CytoTox 96 cytotoxicity assay. Briefly, the cells were exposed to an X-RAD 160–225 instrument to attain the desired dose of 2, 4, or 6 Gy and the culture supernatant was harvested after irradiation of 24 or 48 h. LDH levels were detected according to the manufacturer’s instructions.
Total RNA Extraction and qRT-PCR
Total RNA was extracted from A549 cell lines using a total RNA extraction kit (Solarbio, Beijing, China) following the manufacturer’s protocol. RNA samples were reversed transcribed using an iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA) to synthesize cDNA. qRT-PCR was performed using a CFX96 Real-time System (Bio-Rad) with SYBR Green Supermix (Bio-Rad) according to the manufacturer’s instructions. The 2−ΔΔCT method was used to calculate relative expression levels. The following sense and antisense primers were used: CXCL1: 5ʹ-TGCAGGGAATTCACCCCAAGAAC-3ʹ (sense), 5ʹ-AGTGTGGCTATGACTTCGGTTTGG-3ʹ (antisense); P65: 5ʹ-TGAACCGAAACTCTGGCAGC-3ʹ (sense), 5ʹ-CCACTTGTCGGTGCACATCA-3ʹ (antisense); β-actin: 5ʹ-CCTGGCACCCAGCACAAT-3ʹ(sense), 5ʹ-GGGCCGGACTCGTCATAC-3ʹ (antisense).
Cell Wound Healing, Migration, and Invasion Assays
Transwell assays were performed as previously described.15 For wound healing assays, briefly, irradiated cells were serum-starved for 24 h for cell cycle synchronization, and the confluent cell monolayer (seeded in a 6-well plate) was scraped with a 200-μL sterile pipette tip to create a wound artificially. The wound healing process was observed at the indicated time points and photographed at a magnification of 100×.
Immunofluorescence (IF) Staining
Cultured cells were fixed with 4% paraformaldehyde and washed twice with PBS and then blocked with PBS containing 10% normal goat serum. Then, the samples were incubated with E-cadherin, N-cadherin, and vimentin polyclonal antibodies overnight at 4 °C, washed twice with PBS, stained with Cy3 (red)-conjugated secondary antibody for 1 h at 37 °C, and washed twice with PBS before imaging. All IF images were scanned using a fluorescence microscope (Leica, DM4000B).
Western Blotting
Protein samples were resolved by SDS-PAGE on 12% gels and then transferred to nitrocellulose membranes, blocked for 1 h at room temperature in tris-buffered saline containing 0.1% tween-20 and 5% fat-free milk. Membranes were incubated with primary antibody solutions for 18 h at 4 °C. Subsequently, membranes were stained with secondary antibody solutions at room temperature for 1 h. Enhanced chemiluminescence (ECL) reagent (Millipore Corp. Billerica, MA, USA) or ECL Plus (Amersham Pharmacia Biotech, Buckinghamshire, UK) was used to detect immunoreactive signals according to the manufacturers’ instructions.
Flow Cytometry
Apoptosis was measured by flow cytometry with an Annexin V-FITC/PI apoptosis detection kit (KeyGEN, Jiangsu, China), following the manufacturer’s instructions. A549 cells treated with or without radiation were digested with trypsin without EDTA. The cells were harvested, washed with PBS, and resuspended in 500 μL binding buffer. Then, 5 μL Annexin V-FITC and 5 μL PI were added, incubating the reaction for 15 min at room temperature in the dark. The cells were analyzed with a flow cytometer (FACSCalibur, Becton-Dickinson, USA).
Statistical Analysis
Each in vitro experiment was independently performed at least three times. Data analysis was performed with a paired t-test or one-way ANOVA using GraphPad Prism 7.0 software (GraphPad, San Diego, CA, USA). Statistical significance was defined as P < 0.05 (*), P < 0.01 (**), or P < 0.001 (***).
Results
Radiation Induces Apoptosis and Inhibits the Proliferation of A549 Cells
To test radiation-induced toxicity using A549 cells, cell viability and LDH release were detected. As shown in Figure 1, radiation (4 or 6 Gy) treatment, after 48 h, significantly decreased cell viability in a dose-dependent manner and elevated LDH release in a time- and dose-dependent manner. Consistently, exposure to radiation (4, 6 Gy) induced dose-dependent apoptosis and cell counts/cell viability reached the lowest levels with 6 Gy (Figure 1B). Colony formation assays with the group exposed to radiation (4, 6 Gy) showed dose-dependent inhibition of cell proliferation but cell counts/cell viability reached the lowest levels with 6 Gy (Figure 1A). These results indicate that 6 Gy treatment induces significant cytotoxicity in A549 cells after 48 h. Accordingly, subsequent experiments were performed for 24 or 48 h after 2 or 4 Gy irradiation.
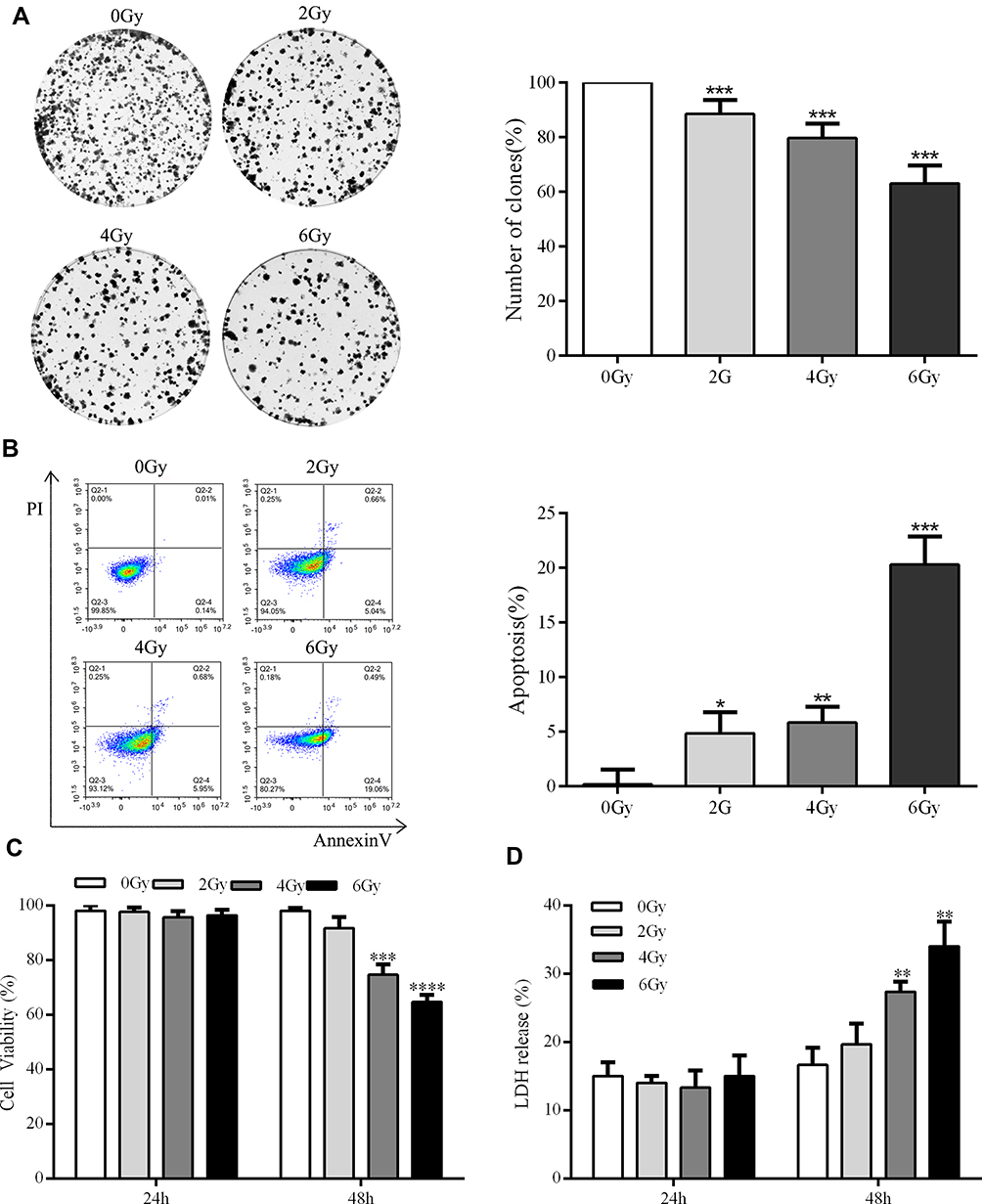 |
Figure 1 Radiation induces apoptosis in A549 cells and inhibits proliferation. (A) Representative images of sphere formation in the indicated groups of A549 cells. (B) Representative flow cytometry scatter plots. (C) Cells were treated with 0, 2, 4, or 6 Gy of radiation. After 24 or 48 h, cell viability was measured using a CCK-8 detection kit. (D) Cells were treated with 0, 2, 4, or 6 Gy of radiation for 24 or 48 h. LDH release was then measured using the CytoTox 96 kit. *P < 0.05, **P < 0.01, ***P <0.001, ****P <0.0001, two-tailed Student’s t-test. |
Low-Dose Radiation Promotes A549 Cell EMT, Migration, and Invasion
Next, we investigated the effect of low-dose radiation on the migration and invasion of A549 cells using an in vitro model. Cultured A549 cells were exposed to radiation with doses of 2 and 4 Gy. After 24 h, wound-healing, transwell, and Matrigel invasion assays were performed to evaluate the migration and invasion of cells. As shown in Figure 2, the results revealed that 4 Gy radiation significantly promoted the migration and invasion in A549 cells (Figure 2A and B). Therefore, we chose 4 Gy as the treatment dosage in our subsequent experiments. To further explore whether low-dose radiation can induce EMT in A549 cells, we analyzed the expression of EMT-related marker proteins after irradiation. Western blotting and IF showed that 4 Gy radiation markedly decreased the levels of the epithelial marker E-cadherin but increased mesenchymal markers N-cadherin and vimentin after 24 h (Figure 2C and D); this suggested that low-dose radiation could promote EMT in A549 cells. These data indicate that low-dose radiation promotes the migration and invasion of A549 cells and promotes EMT in A549 cells.
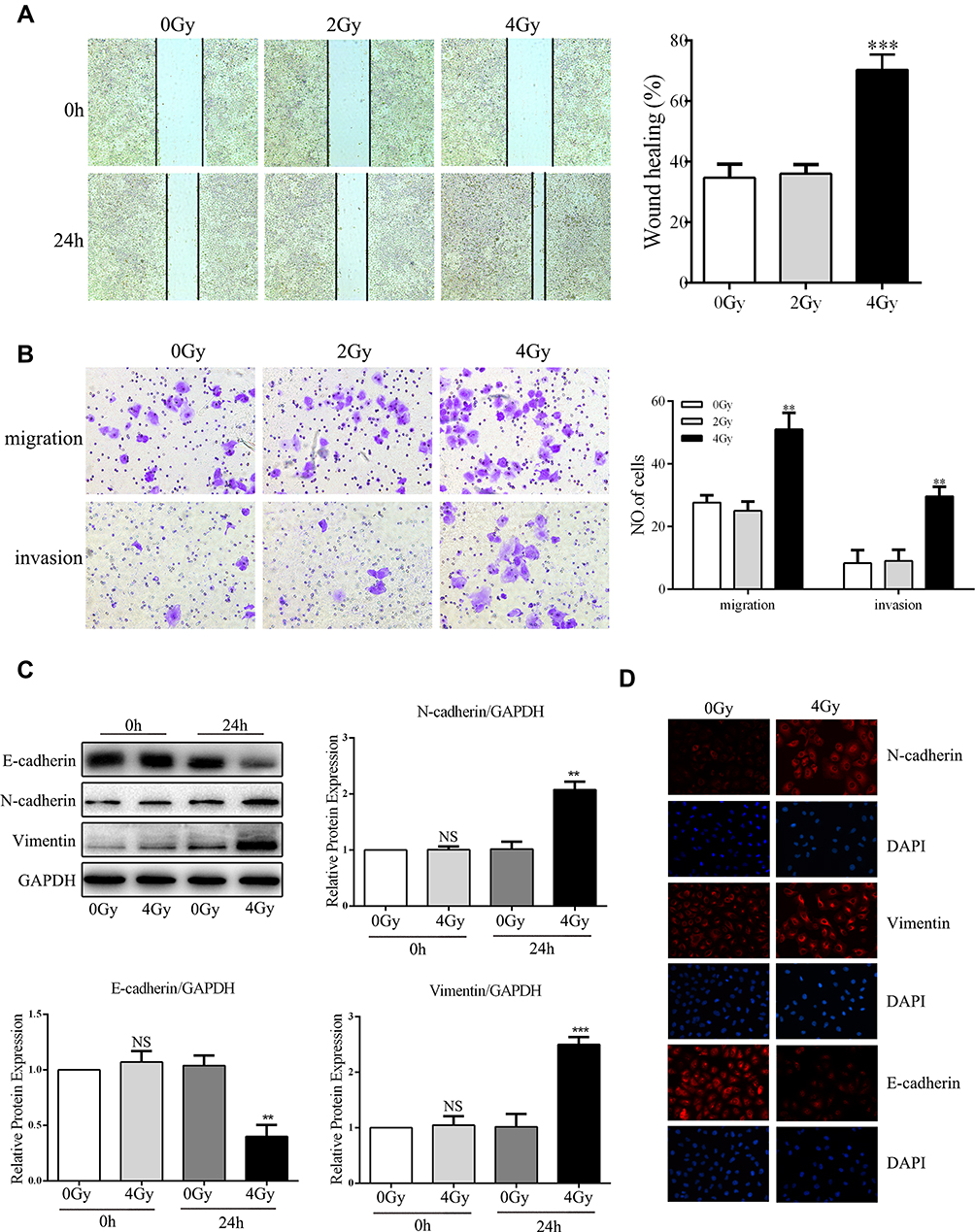 |
Figure 2 Low-dose radiation promotes A549 cells epithelial–mesenchymal transition (EMT), migration, and invasion. (A) Representative images and quantitative analysis of cell migration based on wound-healing assays (scale bar, 500 μm). (B) Representative images and quantitative analysis of cell migration and invasion based on transwell assays (scale bar, 100 μm). (C, D) Cells were irradiated with 0 and 4 Gy. After 24 or 48 h, analysis of EMT markers by Western blotting and immunofluorescence (IF) staining was performed (scale bar, 50 μm). **P < 0.01, ***P < 0.001. Two-tailed Student’s t-test. Abbreviation: NS, not significant. |
CXCL1 Is Essential for Low-Dose Radiation-Induced A549 Cell EMT
It has been widely reported that CXCL1 plays an important role during the progress and development of different cancers. To explore the mechanism of radiation-induced EMT in A549 cells, we investigated the role of CXCL1 in this process by analyzing the effects of radiation on its expression. Western blotting showed that, compared to that in the control group, the expression of CXCL1 in the radiation group was significantly upregulated (Figure 3A and B). Furthermore, stable CXCL1-knockdown cell lines were generated to explore the function of endogenous CXCL1 in radiation-induced A549 cell EMT. We utilized siRNA to induce CXCL1 knockdown in A549 cell lines. We confirmed that knockdown cells had lower mRNA levels of CXCL1 than control cells (Figure 3C). Moreover, as assessed by Western blotting and IF, CXCL1 knockdown markedly increased the epithelial marker E-cadherin, whereas it decreased expression of the mesenchymal markers N-cadherin and vimentin (Figure 3D and E). These data suggest that CXCL1 has a critical role in promoting radiation-induced A549 cells EMT.
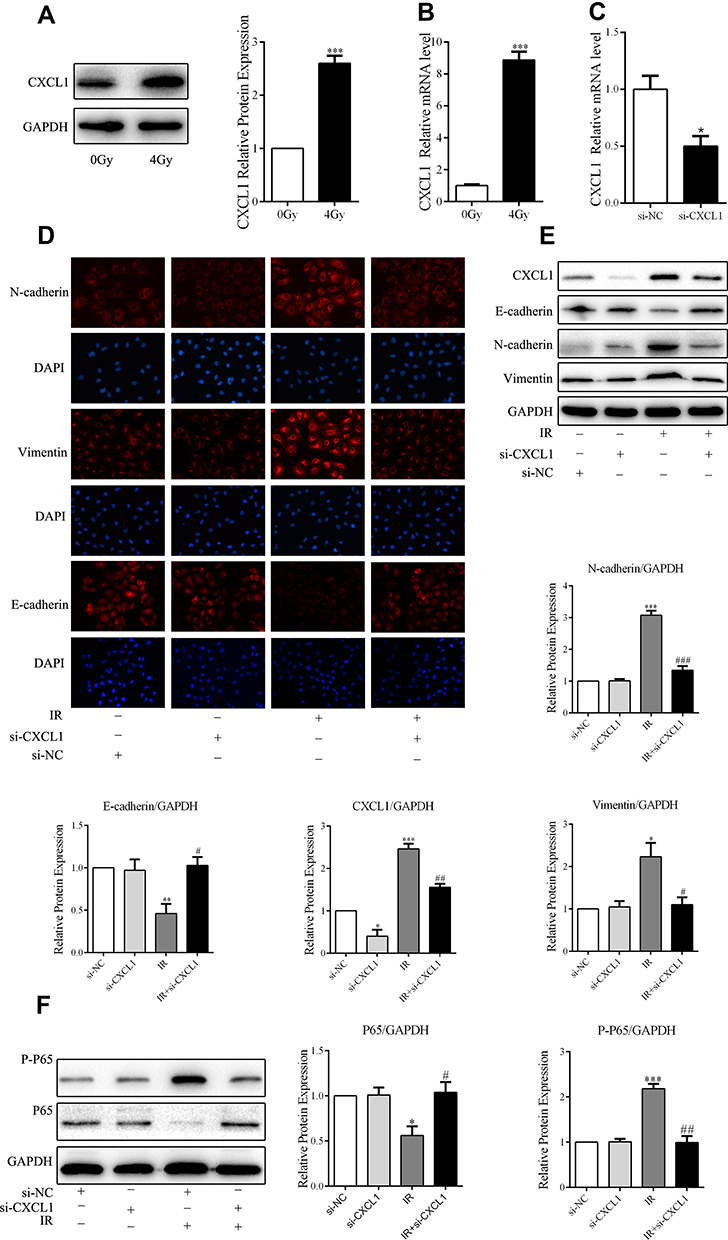 |
Figure 3 CXCL1 is essential for low-dose radiation-induced A549 cell epithelial–mesenchymal transition (EMT). (A) Immunoblot analysis of CXCL1-related proteins. (B) RT-qPCR assays of CXCL1-related mRNA. (C) Expression of CXCL1 in control (Ctl) and CXCL1-silenced (si-CXCL1) cells was detected by RT-qPCR. (D, E) Analysis of EMT markers by Western blotting and immunofluorescence staining (scale bar, 50 μm). (F) Immunoblot analysis of p65 and P-P65 proteins; *P < 0.05, **P < 0.01, ***P < 0.001; #P < 0.05, ##P <0.01, ###P <0.001, two-tailed Student’s t-test. |
The NF-κB Signaling Pathway Plays a Role in Low-Dose Radiation-Enhanced A549 Cell EMT, Migration, and Invasion
Previous studies have reported that CXCL1 strongly correlates with the induction of invasion via the NF-κB signaling pathway. To further explore the regulation and mechanism of radiation-enhanced EMT of A549 cells, we determined the effect of CXCL1 on the NF-κB signaling pathway by Western blot analysis in radiation-pretreated, CXCL1-silenced A549 cells. As expected, the levels of phosphorylated p65 in A549/IR cells were dramatically increased compared to those in control cells, whereas opposite effects were observed in A549/IR/si-CXCL1 cells (Figure 3F). Collectively, these results demonstrate that radiation can activate NF-κB signaling by upregulating CXCL1. Subsequently, we investigated the role of NF-κB signaling in radiation-induced EMT. First, we identified an effective RNAi oligonucleotide to silence p65 expression in A549 cells and confirmed p65 knockdown by qRT-PCR (Figure 4A). After irradiation, the mesenchymal markers N-cadherin and vimentin were significantly upregulated, and the epithelial marker E-cadherin was significantly downregulated in A549/si-NC cells. Accordingly, p65 knockdown in A549 cells mostly inhibited radiation-induced changes in EMT markers (Figure 4B and C). Combining these data, our results revealed that p65 knockdown inhibits radiation-induced EMT. Furthermore, wound-healing, transwell, and Matrigel invasion assays confirmed that p65 knockdown, but not scramble control, could inhibit radiation-induced migration and invasion in A549 cells (Figure 4D and E). In conclusion, our data suggest that p65 knockdown inhibits radiation-induced migration and invasion.
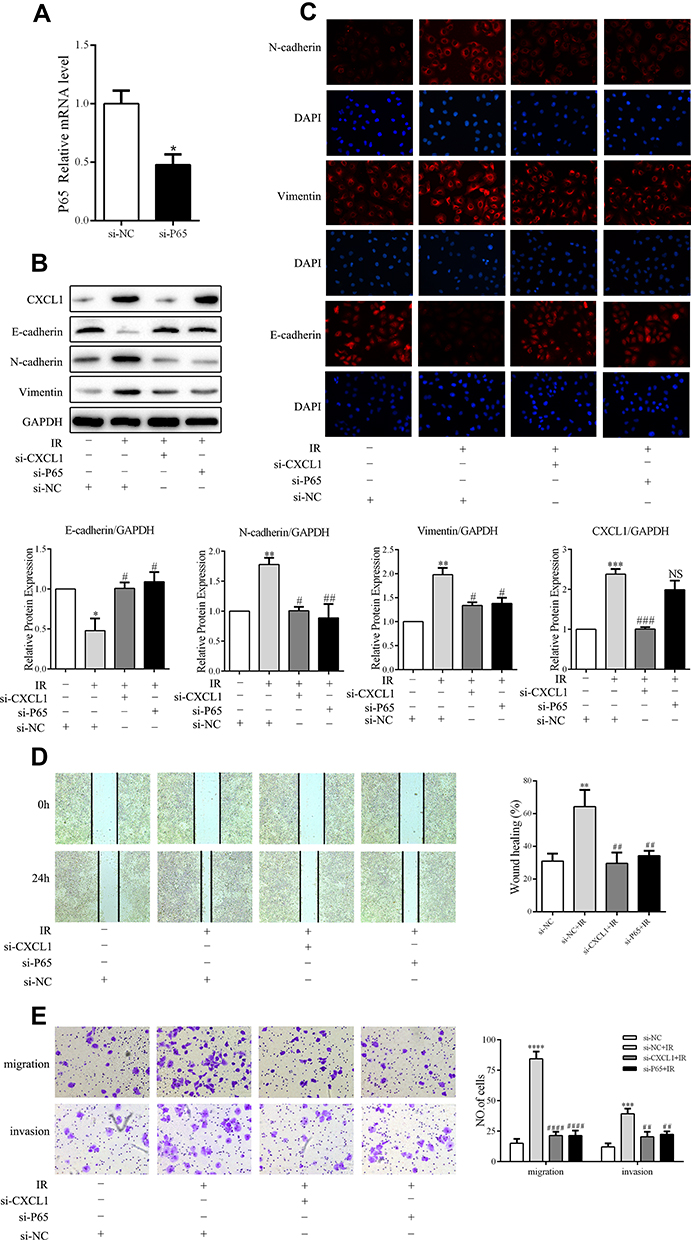 |
Figure 4 The NF-κB signaling pathway plays a role in low-dose radiation-enhanced A549 cell epithelial–mesenchymal transition (EMT), migration, and invasion. (A) Expression of p65 in control (si-NC) and p65-silenced (si-P65) cells was detected by RT-qPCR. (B, C) Analysis of EMT markers by Western blotting and immunofluorescence staining (scale bar, 50 μm). (D) Representative images and quantitative analysis of cell migration based on wound-healing assays (scale bar, 500 μm). (E) Representative images and quantitative analysis of cell migration and invasion based on transwell assays (scale bar, 100 μm). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; #P < 0.05, ##P < 0.01, ###P < 0.001, ####P < 0.0001. Two-tailed Student’s t-test. Abbreviation: NS, not significant. |
Discussion
According to the latest statistics of global cancer in 2018, lung cancer is one of the most malignant tumors with the highest morbidity and mortality worldwide.16 Currently, radiotherapy is still one of the main methods for treating this cancer.4 Whereas the overall survival benefits of localized radiation have been demonstrated in several clinical studies, local recurrence or distant metastases that develop after local treatment in some patients remain a major therapeutic challenge.5 Thus, investigating the molecular mechanisms that regulate radiation-induced tumor cell invasion and migration is the key to effectively responding to radiotherapy-induced tumor recurrence and metastasis.
IR activates the intracellular cycle and apoptotic pathway through direct or indirect effects and plays a role in inhibiting tumor cell proliferation or promoting its death.8 Our results show that apoptosis was not significantly induced in cells treated with 2 and 4 Gy but was significantly induced when the radiation dose was increased to 6 Gy. These results are consistent with the observations of other researchers.8 Moreover, recent studies have found that low-dose radiation can lead tumor cells to acquire high migration and invasion capabilities.17–19 Accordingly, we found that upon 4-Gy irradiation, cell proliferation was inhibited, whereas invasion and migration were significantly enhanced, and these results suggested that that low-dose ionizing radiation can promote tumor invasion and migration.
The correlation between lung cancer metastasis and EMT, one of the pivotal cytological foundations of tumor invasion and migration, has recently attracted attention. EMT can lead polar cells to undergo morphological changes and transform into interstitial cells, providing prerequisites for tumor cell invasion and metastasis.19,20 Studies have shown that LDIR promotes the invasiveness of breast cancer cells through EMT.21 However, whether LDIR can induce EMT in A549 cells remained elusive. We demonstrated here that after administering 4 Gy irradiation to tumor cells, the expression of EMT marker proteins vimentin and N-cadherin increased significantly, whereas the expression of E-cadherin decreased significantly. This therefore suggests that low-dose radiation can increase the invasion and metastasis ability of A549 cells and simultaneously induce EMT.
Recent studies have reported that CXCL1 is a key molecule leading to EMT and plays an important role in the development of various malignancies and metastasis.22,23 Moreover, its high expression is associated with poor prognosis.23 In this study, we showed for the first time that low-dose radiation can significantly upregulate the expression of CXCL1. Furthermore, the expression of EMT marker proteins in irradiated CXCL1-silenced cells was significantly reduced, compared to that in irradiated non-silenced cells. Taken together, low-dose ionizing radiation might induce CXCL1 expression and thereby promote tumor cell EMT.
The NF-κB signaling pathway is closely related to tumorigenesis, invasion, and metastasis.24–26 Yang et al found that p65-regulated EMT induced by the low-dose irradiation of cervical cancer cell lines promotes invasiveness and metastasis.27 Interestingly, studies have reported that CXCL1 promotes breast cancer metastasis via NF-κB.28,29 Therefore, we speculated that the possible mechanism whereby CXCL1 regulates radiation-induced tumor cell EMT is the activation of NF-κB signaling. We confirmed that IR can activate the NF-κB pathway. Furthermore, we found that compared to that in cells only irradiated, the expression of vimentin and N-cadherin in the p65-silenced and irradiated cells was significantly lower, whereas the expression of E-cadherin was significantly higher. Importantly, the invasion and metastasis ability were significantly reduced in the p65-silenced and irradiated cells. Collectively, these data suggest that CXCL1 might regulate low-dose radiation-induced tumor cell EMT by activating the NF-κB signaling pathway and affecting invasion and metastasis.
Conclusion
In summary, we have revealed for the first time that low-dose ionizing radiation might enhance tumor cell invasion and migration by inducing CXCL1 expression, thereby activating the NF-κB signaling pathway and regulating tumor cell EMT. This study sheds light on radiation-induced tumor cell invasion and migration, paving the way for novel therapeutic avenues in the treatment of clinical recurrence and metastasis after radiotherapy. Further, detecting other EMT markers and performing these experiments in animal models will further strengthen our conclusions. Additionally, the specific mechanism whereby NF-κB affects tumor cell EMT needs to be further investigated.
Key Points
- High-dose radiation induces apoptosis and inhibits the proliferation of A549 cells. Instead, low-dose radiation promotes tumor cell invasion, migration, and epithelial-mesenchymal transition via upregulation of CXCL1 expression and activation of the NF-κB signaling pathway.
- We demonstrated low dose radiation can regulate the epithelial-mesenchymal transition of tumor cells. These findings provide new insights into the prevention of tumor invasion and metastasis induced by radiotherapy.
Acknowledgments
Jing Li, Dong-ming Wu and Rong Han are first co-authors. This study was funded by the National Natural Science Foundation of China (81802955, 81972977), the Foundation of Sichuan Science and Technology Agency (2018JY0648, 2019YJ0589), the Foundation of The First Affiliated Hospital of Chengdu Medical College (CYFY2017ZD03, CYFY2018ZD02), the Foundation of Collaborative Innovation Center of Sichuan for Elderly Care and Health, Chengdu Medical College (19Z01), and the Foundation of Chengdu Medical College (CYCG16-04). The authors thank Editage for English language editing.
Disclosure
The authors have no competing financial interests to disclose.
References
1. Barta JA, Powell CA, Wisnivesky JP. Global epidemiology of lung cancer. Ann Glob Health. 2019;85:8. doi:10.5334/aogh.2419
2. Tsoi KK, Hirai HW, Chan FC, Griffiths S, Sung JJ. Cancer burden with ageing population in urban regions in China: projection on cancer registry data from World Health Organization. Br Med Bull. 2017;121:83–94. doi:10.1093/bmb/ldw050
3. Wu C, Li M, Meng H, et al. Analysis of status and countermeasures of cancer incidence and mortality in China. Sci China Life Sci. 2019;62:640–647. doi:10.1007/s11427-018-9461-5
4. Jumeau R, Vilotte R, Durham AD, Ozsahin EM. Current landscape of palliative radiotherapy for non-small-cell lung cancer. Transl Lung Cancer Res. 2019;8:S192–S201. doi:10.21037/tlcr.2019.08.10
5. Roy SF, Louie AV, Liberman M, Wong P, Bahig H. Pathologic response after modern radiotherapy for non-small cell lung cancer. Transl Lung Cancer Res. 2019;8:S124–S134. doi:10.21037/tlcr.2019.09.05
6. Nesbit EG, Leal TA, Kruser TJ. What is the role of radiotherapy for extensive-stage small cell lung cancer in the immunotherapy era? Transl Lung Cancer Res. 2019;8:S153–S162. doi:10.21037/tlcr.2019.05.01
7. Vilalta M, Rafat M, Graves EE. Effects of radiation on metastasis and tumor cell migration. Cell Mol Life Sci. 2016;73:2999–3007. doi:10.1007/s00018-016-2210-5
8. Blyth BJ, Cole AJ, MacManus MP, Martin OA. Radiation therapy-induced metastasis: radiobiology and clinical implications. Clin Exp Metastasis. 2018;35:223–236. doi:10.1007/s10585-017-9867-5
9. Kaplan HS, Murphy ED. The effect of local roentgen irradiation on the biological behavior of a transplantable mouse carcinoma; increased frequency of pulmonary metastasis. J Natl Cancer Inst. 1949;9:407–413.
10. Sundahl N, Duprez F, Ost P, De Neve W, Mareel M. Effects of radiation on the metastatic process. Mol Med. 2018;24:16. doi:10.1186/s10020-018-0015-8
11. Fujita M, Yamada S, Imai T. Irradiation induces diverse changes in invasive potential in cancer cell lines. Semin Cancer Biol. 2015;35:45–52. doi:10.1016/j.semcancer.2015.09.003
12. Wang L, Zhang C, Xu J, et al. CXCL1 gene silencing inhibits HGC803 cell migration and invasion and acts as an independent prognostic factor for poor survival in gastric cancer. Mol Med Rep. 2016;14:4673–4679. doi:10.3892/mmr.2016.5843
13. Wang Y, Liu J, Jiang Q, et al. Human adipose-derived mesenchymal stem cell-secreted CXCL1 and CXCL8 facilitate breast tumor growth by promoting angiogenesis. Stem Cells. 2017;35:2060–2070. doi:10.1002/stem.2643
14. Zhuo C, Wu X, Li J, et al. Chemokine (C-X-C motif) ligand 1 is associated with tumor progression and poor prognosis in patients with colorectal cancer. Biosci Rep. 2018;38:BSR20180580. doi:10.1042/BSR20180580
15. Ma L, Teruya-Feldstein J, Weinberg RA. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature. 2007;449:682–688. doi:10.1038/nature06174
16. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. doi:10.3322/caac.21492
17. Young AGH, Bennewith KL. Ionizing radiation enhances breast tumor cell migration in vitro. Radiat Res. 2017;188:381–391. doi:10.1667/RR14738.1
18. Moncharmont C, Levy A, Guy JB, et al. Radiation-enhanced cell migration/invasion process: a review. Crit Rev Oncol Hematol. 2014;92:133–142. doi:10.1016/j.critrevonc.2014.05.006
19. Nagaraja SS, Krishnamoorthy V, Raviraj R, Paramasivam A, Nagarajan D. Effect of Trichostatin A on radiation induced epithelial-mesenchymal transition in A549 cells. Biochem Biophys Res Commun. 2017;493:1534–1541. doi:10.1016/j.bbrc.2017.10.031
20. Jolly MK, Celià-Terrassa T. Dynamics of phenotypic heterogeneity associated with EMT and stemness during cancer progression. J Clin Med. 2019;8:E1542. doi:10.3390/jcm8101542
21. Zhang X, Li X, Zhang N, et al. Low doses ionizing radiation enhances the invasiveness of breast cancer cells by inducing epithelial-mesenchymal transition. Biochem Biophys Res Commun. 2011;412:88–92. doi:10.1016/j.bbrc.2011.07.074
22. Lu Y, Dong B, Xu F, et al. CXCL1-LCN2 paracrine axis promotes progression of prostate cancer via the Src activation and epithelial-mesenchymal transition. Cell Commun Signal. 2019;17:118. doi:10.1186/s12964-019-0434-3
23. Kasashima H, Yashiro M, Nakamae H, et al. Clinicopathologic significance of the CXCL1-CXCR2 axis in the tumor microenvironment of gastric carcinoma. PLoS One. 2017;12:e0178635. doi:10.1371/journal.pone.0178635
24. Mali AV, Joshi AA, Hegde MV, Kadam SS. Enterolactone modulates the ERK/NF-κB/Snail signaling pathway in triple-negative breast cancer cell line MDA-MB-231 to revert the TGF-β-induced epithelial-mesenchymal transition. Cancer Biol Med. 2018;15:137–156. doi:10.20892/j.issn.2095-3941.2018.0012
25. Zhang T, Liu L, Lai W, et al. Interaction with tumor‑associated macrophages promotes PRL‑3‑induced invasion of colorectal cancer cells via MAPK pathway‑induced EMT and NF-κB signaling‑induced angiogenesis. Oncol Rep. 2019;41:2790–2802. doi:10.3892/or.2019.7049
26. Li W, Li S, Yang J, Cui C, Yu M, Zhang Y. ITGBL1 promotes EMT, invasion and migration by activating NF-κB signaling pathway in prostate cancer. Onco Targets Ther. 2019;12:3753–3763. doi:10.2147/OTT.S200082
27. Yan S, Wang Y, Yang Q, et al. Low-dose radiation-induced epithelial-mesenchymal transition through NF-κB in cervical cancer cells. Int J Oncol. 2013;42:1801–1806. doi:10.3892/ijo.2013.1852
28. Wu Z, Neufeld H, Torlakovic E, Xiao W. Uev1A-Ubc13 promotes colorectal cancer metastasis through regulating CXCL1 expression via NF-κB activation. Oncotarget. 2018;9:15952–15967. doi:10.18632/oncotarget.24640
29. Wang N, Liu W, Zheng Y, et al. CXCL1 derived from tumor-associated macrophages promotes breast cancer metastasis via activating NF-κB/SOX4 signaling. Cell Death Dis. 2018;9:880. doi:10.1038/s41419-018-0876-3
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.