")
Back to Journals » Cancer Management and Research » Volume 14
Long Noncoding RNAs in Lung Cancer: From Disease Markers to Treatment Roles
Authors Chen L, Xie Y, Yu M, Gou Q
Received 10 March 2022
Accepted for publication 9 May 2022
Published 21 May 2022 Volume 2022:14 Pages 1771—1782
DOI https://doi.org/10.2147/CMAR.S365762
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Beicheng Sun
Lin Chen,1,2,* Yuxin Xie,1,3,* Min Yu,2 Qiheng Gou1
1Department of Head and Neck Oncology, Cancer Center, West China Hospital, Sichuan University, Chengdu, Sichuan Province, People’s Republic of China; 2Department of Thoracic Oncology, Cancer Center and State Key Laboratory of Biotherapy, West China Hospital, Sichuan University, Chengdu, Sichuan Province, People’s Republic of China; 3Laboratory of Molecular Diagnosis of Cancer, Clinical Research Center for Breast, West China Hospital, Sichuan University, Chengdu, Sichuan Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qiheng Gou, Department of Head and Neck Oncology, Cancer Center, West China Hospital, Sichuan University, 37 Guoxue Xiang, Wuhou District, Chengdu, 610041, People’s Republic of China, Tel +86-28-85423278, Email [email protected]
Abstract: There is an urgent need to identify reliable biomarkers that can be used in early diagnosis, prognostication prediction and as possible therapeutic targets for lung cancer due to its current poor prognosis. Long noncoding RNAs (lncRNAs) have recently attracted additional attention due to their potential role in carcinogenesis, invasion and metastasis. Issues involved in the biofunctions and regulatory mechanisms of oncogenic and tumor-suppressive lncRNAs in lung cancer are discussed. Some lncRNAs have shown good diagnostic value, especially in combination with conventional serum protein markers. The use of antisense oligonucleotides, small molecules and RNA interference techniques have shown promise as direct therapeutic tools for targeting lncRNAs in preclinical studies. The biomarker function of lncRNAs may also indirectly involved in tumor therapy as a reference to conventional therapy. Overall, the concept of using lncRNAs as biomarkers for prognostication and intervention in lung cancer is still in its infancy, and only with more in-depth studies could they have a significant impact.
Keywords: long noncoding RNAs, biomarker, lung cancer, diagnosis, treatment
Introduction
Lung cancer is a malignant tumor that originates in the mucosa or glands of the bronchi of the lung. Globally, lung cancer is the second most frequent cancer and the leading cause of cancer death in 2020, with an estimated 2.2 million new cases (11.4% of total cases) and 1.8 million deaths (18.0% of total deaths).1 Although the introduction of immune checkpoint inhibitors (ICIs) has fundamentally transformed the treatment of lung cancer, the 5-year overall survival rate remains low, rarely exceeding 30%.2
Noncoding ribonucleic acids, which comprise more than 97% of the human genome, have a variety of physiological and pathological functions and they play a major role in lung cancer tumorigenesis.3 Furthermore, noncoding ribonucleic acids can be divided into short and long noncoding RNAs (lncRNAs). LncRNAs are extensively involved in biological processes such as gene expression, cell differentiation, organogenesis and homeostasis in human tissues. In recent years, lncRNAs have drawn extra attention for their role in human hematological or solid malignancies. More than 95% of the articles retrieved from PubMed on lncRNAs and lung cancer were published after 2017.
Although current targeted therapies for lung cancer have relatively high response rates, the inevitability of treatment resistance and the rarity of driver mutations in squamous and small cell lung cancers limit the efficacy of targeted therapies and the prognostic value of driver mutation detection. To find new therapeutic and diagnostic targets, additional molecular pathways must be explored and investigated. LncRNAs may be potential candidates for this purpose.
In this article, we reviewed the role of lncRNAs in diagnosis, treatment response monitoring and prognostication of lung cancers.
An Overview on lncRNAs
LncRNAs are nonprotein-coding RNAs of more than 200 nucleotides and can be loosely defined as transcripts that carry out their functions as RNA molecules and do not belong to any other class of small or structural RNA molecules.4 LncRNAs can be categorized into six groups based on the genomic localization of the nearest protein-coding gene, namely, (a) sense, (b) antisense, (c) bidirectional (d) intronic, (e) intergenic and (f) enhancer lncRNAs.4–6 Similar to messenger RNAs, lncRNA expression is also regulated transcriptionally and epigenetically.7,8 Unlike protein-coding genes, a higher density of DNA methylation around their transcription start sites has been observed, suggesting they play an important role in the epigenetic regulatory network.7 LncRNAs are usually transcribed by RNA polymerase II and their posttranscriptional processing is similar to that of protein-coding genes, including alternative splicing, 5′-capping, and polyadenylation.9
The diversity of lncRNAs helps explain the developmental complexity of higher evolved organisms and demonstrates their important functional role in biological processes by interacting with chromatin, interacting with RNA targets, and regulating proteins.4,9–12 LncRNAs are also involved in various posttranscriptional modifications, including phosphorylation and ubiquitination.13
LncRNAs in Lung Cancer
Although many new lncRNAs have been identified by current screening methods, their functional characteristics need to be experimentally demonstrated. Only a few validated and well-characterized lncRNAs have been reported to be deregulated and functional in lung cancer, as summarized in Table 1.
 |
Table 1 Summarization of Discussed lncRNAs |
Oncogenic lncRNAs
Oncogenes are genes whose encoded products can promote tumorigenesis and progression. Multiple oncogenic lncRNAs, such as metastasis-associated lung adenocarcinoma transcript 1 (MALAT1), HOX transcript antisense RNA (HOTAIR) and H19, promote the growth, proliferation and invasion of lung cancer.
MALAT1
MALAT1, also known as noncoding nuclear-enriched abundant transcript 2 (NEAT2), is a highly abundant and conserved nuclear lncRNA.14 MALAT1 is involved in many types of physiological processes, such as alternative splicing, nuclear organization, epigenetic regulation of gene expression, and pathological processes, ranging from diabetes complications to cancers.15,16 As one of the first identified cancer-associated lncRNAs, MALAT1 has been found to have prognostic value in different types of tumor models, including non-small-cell lung cancer (NSCLC), renal clear cell carcinoma, pancreatic cancer and hepatocellular carcinoma (HCC).17–21
In NSCLC, MALAT1 is highly expressed in primary tumors and tumors overexpressing it have a higher tendency to metastasize.18,22 Highly efficient knockdown of MALAT1 using zinc finger nuclease-based technology confirmed that MALAT1 promotes metastasis in vitro and in vivo without affecting cell proliferation.23–25 Conversely, the use of antisense oligonucleotides (ASOs) inhibits metastasis formation by reducing the expression level of MALAT1 in tumors and surrounding stromal cells.25 Furthermore, MALAT1 enhances the brain metastatic potential of lung cancer by regulating the epithelial-to-mesenchymal transition (EMT) process.26,27 Additionally, in a lung cancer homing model, MALAT1 and gene knockout reduced the possibility of lung cancer cells homing to the lung.25 The function of MALAT1 is regulated by multiple signaling pathways in tumor proliferation, invasion and metastasis.
The mode of action of MALAT1 is currently speculated to be multifaceted, including transcription and/or premRNA splicing of critical genes involved in migration and cell adhesion, as well as genes involved in critical cancer pathways. MALAT1 regulates the activity of serine/arginine (SR) splicing factors, thereby affecting gene expression through alternative splicing.28 In a normal human diploid fibroblast cell line, MALAT1 deprivation resulted in a proliferation defect, attributed in part to changes in alternative splicing of mitotic regulators such as transcription factor B-MYB.29 Moreover, MALAT1 is involved in the regulation of the cell cycle. The interaction of MALAT1 with nuclear heterogeneous nuclear ribonucleoprotein C (hnRNP C) is essential to promote hnRNP C translocation to the cytoplasm during G2/M phase. In addition, MALAT1 is also involved in the epigenetic control of gene expression by acting as a subnuclear molecular scaffold that specifically localizes unmethylated polycomb protein 2 to promote gene activation between chromatin bodies.30
Despite the diverse functions and high conservation of MALAT1, MALAT1 knockout mice are viable and fertile, with no defects in gene expression, SR splicing factor activity, or alternative splicing levels. Thus, MALAT1 emerges as a potential target for preventing metastasis in lung cancer.
HOTAIR
HOTAIR is transcribed in the antisense direction from chromosome 12 between HOXC11 and HOXC12. Sequence analysis of HOTAIR showed that it is present in mammals with poorly conserved sequences but a reasonably conserved structure.31 HOTAIR conserved sequences predate whole genome duplication events in vertebrate roots, with evidence that HOTAIR has 32 nucleotide-long conserved noncoding elements (CNEs) similar to those in the HOXD cluster region.32
HOTAIR is thought to function as a modular scaffold due to its 5′ and 3′ structural domains interacting with polycomb repressive complex 2 (PRC2) and lysine-specific demethylase 1 (LSD1), respectively.33,34 By interacting with PRC2, HOTAIR triggers histone 3 lysine 27 (H3K27) trimethylation of the HOXD locus on human chromosome 2, which modulates the chromatin state.33,35 Additionally, since LSD1 is part of the RE1-silencing transcription factor CoREST-REST repressor complex, the interaction of the 3′ structural domain of HOTAIR with LSD1 will lead to the demethylation of the H3K4me2 histone mark near the transcription start site of the HOXD gene, resulting in epigenetic regulation of HOXD gene transcription.34
HOTAIR is highly expressed in most tumors, including breast cancer, HCC, colorectal cancer, gastric cancer and NSCLC. High HOTAIR levels are associated with advanced or metastatic disease and a poor prognosis.35–39 In NSCLC, 22.1% of patients show at least a 2-fold increase in HOTAIR expression, and the more advanced the stage, the higher the fold increase.39 Reduced invasive and metastatic potential of cancer cells was observed in vivo and in vitro assays through siRNA-mediated downregulation of HOTAIR.39 Studies further confirm that HOTAIR mediates its prometastatic and proinvasive functions by regulating the levels of matrix metalloproteinase 2 (MMP2), MMP9 and homeobox protein A5 (HOXA5) proteins but not EMT-promoting proteins. In addition to its prometastatic function, high expression of HOTAIR leads to resistance to cisplatin by downregulating CDKN1A in a lung adenocarcinoma model.40,41 In terms of targeted therapy, downregulation of HOTAIR increases the sensitivity of NSCLC cell lines to crizotinib by inhibiting the phosphorylation of ULK1.42 Similar to NSCLC, in small cell lung cancer (SCLC), downregulation of HOTAIR can improve the sensitivity of SCLC to cisplatin, adriamycin and etoposide by reducing the expression of DNA methyltransferase 1 and 3B.43
In summary, HOTAIR is involved in gene regulation and tumor metastasis processes and is significantly associated with a poor prognosis and treatment resistance in cancer. Therefore, it has a high biomarker potential. In the future, with more detailed studies about the regulation and mode of action of HOTAIR, it may become a therapeutic target for tumors.
H19
H19 is a paternally imprinted gene localized on human chromosome 11p15 that is only transcribed from the maternally inherited allele. H19 was originally named ASM because it is expressed in adult skeletal muscle (“ASM”) of rats.44 Mineral dust and tobacco smoke are well-known carcinogens, both of which can increase the expression level of H19 through epigenetic changes, further contributing to the poor prognosis of lung cancer.45,46 H19 is known to have oncogenic effects in several cancers, such as hepatocellular carcinoma, bladder cancer, breast cancer, gastric cancer, colorectal cancer and lung cancer.47 In NSCLC, H19 induces cell proliferation, migration, survival, invasion and EMT while reducing apoptosis.48–51
In NSCLC, the expression level of H19 is significantly increased, which impairs the inhibitory effect of miR-138 on PDK1 protein expression, resulting in decreased miR-138 and enhanced cell proliferation ability.49 Similarly, overexpression of H19 inhibits the function of miR-200a, thereby upregulating the miR-200a targeted genes ZEB1 and ZEB2, which enhanced the proliferation and metastasis of lung cancer.50 LncRNA H19/miR-29b-3p/STAT3 signaling is involved in the EMT process in lung adenocarcinoma.51
Studies have confirmed that hypoxia is a major inducer of H19.52,53 Thus, H19 functions as a stress modulator and survival factor and is involved in several essential processes triggered by hypoxia. In lung cancer cells, H19 is induced in a p53-dependent manner.53 Reconstitution of wild-type p53 in lung cancer cells with null p53 mutations eliminated the induction of H19 by hypoxia. For lung cancer cells possessing wild-type p53, both inhibition of p53 and overexpression of HIF1-α are required to significantly upregulate H19 in response to hypoxia.53
In the clinical setting, by using quantitative reverse transcription PCR (qRT–PCR) to detect H19 expression in tumor tissues and corresponding nontumor NSCLC tissues of 70 patients, it was found that high H19 expression was positively correlated with advanced stage and tumor size. Multivariate analysis revealed that H19 expression could be an independent prognostic factor for overall survival in NSCLC.54 In therapeutic applications, approaches have targeted different cancer-specific transcriptional products of H19 or used their transcriptional regulatory elements to drive the expression of cytotoxic genes, specifically targeting tumor cells. These would have a key advantage over current conventional therapeutic agents, as they could spare normal cells.
In summary, H19 is considered to be a biomarker of cancer and a potential therapeutic target for these human diseases, but further investigations are necessary required.
Tumor-Suppressive lncRNAs
Multiple tumor suppressor lncRNAs, such as maternally expressed gene 3 (MEG3), growth arrest-specific transcript 5 (GAS5), and sprouty 4 intronic transcript 1 (SPRY4-IT1), are downregulated in NCSLC, thereby promoting the development and progression of NSCLC.
MEG3
MEG3 is a maternally expressed, imprinted lncRNA located on chromosome 14q32.3. MEG3 acts as a growth suppressor in tumor cells and activates p53.55,56
A reduction or loss of MEG3 expression has been observed in various cancers, including lung cancer, and its lower expression level is associated with advanced stage and poor outcome.57 MEG3 was found to be decreased in lung cancer stem cells, reducing its suppression of migration and invasion.58 Moreover, overexpression of MEG3 contributes to restoring the sensitivity of tumor cells to chemotherapy by inhibiting cell differentiation, inducing apoptosis and reducing autophagy, both in vivo and in vitro.59,60 MEG3 overexpression also leads to reduced expression of N-cadherin, vimentin, snail-1, and -catenin, thereby decreasing EMT.61
In most cases, MEG3 dysregulation is due to copy number loss of 14q32 or CpG hypermethylation in functionally critical genomic regions upstream of the MEG3 gene, which further inhibits the p53 pathway.62–64 MEG3 interacts with the transcription factor c-Jun, which inhibits PHLPP1 transcription and leads to activation of the Akt/p70S6K/S6 axis and translation of the HIF-1α protein, thus exerting a tumor suppressive effect.65
Due to its prognostic properties and tumor suppressor function, MEG3 could theoretically serve as a marker of prognosis or therapeutic response or even be a target for tumor therapy. In gliomas, the DNA methylation inhibitor 5-aza-2’-deoxycytidine (5-azadC) reduced the abnormal hypermethylation of the MEG3 promoter and prevented the hypoexpression of MEG3.64 However, more studies on the role of MEG3 in lung cancer are necessary.
GAS5
GAS5 was originally identified by Schneider from a cDNA library enriched for RNA sequences in growth arrested mouse fibroblasts and was found to be located on human chromosome 1q25.66,67 The structure and biological function of GAS5 is conserved in mice and humans.68 GAS5 expression is increased during growth arrest, making the cells sensitive to apoptosis inducers.69 Further study revealed that GAS5 could bind competitively to glucocorticoid response elements on DNA, thus preventing the activation of glucocorticoid response genes and reducing cellular metabolism, growth arrest, and apoptosis sensitization.70 Due to its physiological role in growth arrest and apoptosis, GAS5 also has a tumor suppressive role and its expression is downregulated in a variety of tumors, including lung cancer.69,71,72 Downregulation of GAS5 is correlated with advanced stage, increased tumor size and a worse poorer prognosis. Further exploration of the GAS5 mechanism shows that ectopic overexpression of GAS5 increases the protein levels of P53 and P21 and decreases the expression of E2F1.72
From a clinical point of view, since GAS5 levels are correlated with patient survival, they can be used as a prognostic marker and efficacy predictor for a variety of tumors. The regulatory mechanisms of GAS5 in inhibiting cell proliferation and promoting apoptosis offer the possibility of pharmacological intervention. GAS5 antitumor effects could theoretically be exerted either by drug-induced expression of endogenous GAS5 or by expression of RNA containing GAS5ʹs functional domain.
SPRY4-IT1
SPRY4-IT1 is located within the second intron of the SPRY4 gene on chromosome 5q31. SPRY4 acts as an inhibitor of the MAPK signaling pathway, which acts in the upstream pathway of RAS activation and inhibits the formation of GTP-RAS.73 SPRY4-IT1 acts as a tumor suppressor in lung cancer by inhibiting tumor cell growth, migration, invasion and EMT. SPRY4 mRNA and protein levels are decreased in lung cancer cell lines.74 Of clinical importance, patients with low SPRY4-IT1 expression have a worse prognosis than those with relatively high expression.
It has been demonstrated that SPRY4-IT1 is negatively correlated with the expression of EZH2, a core subunit of PRC2. In lung adenocarcinoma cell lines, EZH2 promotes H3K27 trimethylation by binding to the promoter of SPRY4-IT1, which in turn downregulates SPRY4-IT1 expression.75 It has also been found that knockdown of SPRY4-IT1 results in downregulation of EMT-related e-cadherin expression and upregulation of vimentin expression.76 Thus, EZH2 overexpression promotes invasion and migration of lung adenocarcinoma cells by suppressing SPRY4-IT1 expression.
Interestingly, unlike in lung cancer, SPRY4-IT1 is upregulated and exerts tumor-promoting effects in a variety of tumors, including melanoma, ovarian, bladder, and colorectal cancers.77–80 Thus, SPRY4-IT1 downregulation may be a specific biomarker for lung cancer patients.
Overall although some lncRNAs have shown potential as markers for predicting disease staging and metastasis and assessing patient prognosis, there are still many obstacles to the application of lncRNAs in the clinical setting. On the one hand, preclinical experiments have shown some results, but many lncRNAs are primate/human-specific and cannot be studied in mice. Additionally, even though altered expression of lncRNAs has been determined to be associated with various tumors, it is not yet clear whether the alterations are a cause or a consequence of the disease; thus, a comprehensive understanding of the structure, mechanisms, intermolecular interactions of lncRNAs and the development of new quantitative analytical screening drugs is needed.
LncRNA-Based Diagnostics and Therapies in Lung Cancer
Lung cancer is a malignancy with a very poor prognosis and a low 5-year overall survival rate, mainly due to late diagnosis and rapid development of treatment resistance. There is an immediate need for new highly sensitive, specific, stable, easily and rapidly detectable biomarkers. LncRNA detection is facilitated due to its stability and ease of obtaining it from plasma or other body fluids. Second, since the expression of multiple lncRNAs is altered during tumor formation, lncRNAs can be used in combination approaches to establish more reliable diagnostic and prognostic models.81 Taken together, lncRNAs hold promise as ideal biomarkers.
LncRNAs in Lung Cancer Diagnosis
Due to the lack of stable and reliable biomarkers, many patients are currently diagnosed at an advanced stage. Difficulty in early diagnosis is the main reason for the poor prognosis of lung cancer. In the past, serum proteins such as CEA and CYFRA21-1 have been the main targets of tumor biomarker research, but their diagnostic applications are still limited. With in-depth research on the mechanism and functions of lncRNAs, there is increasing interest in the role of lncRNAs in early diagnosis.
Since circulating lncRNAs are contained within lipid or lipoprotein vesicles (eg, apoptotic cells, exosomes, or microvesicles), they exhibit high stability even under repeated freezing and thawing, incubation at 45 °C for 24 hours, incubation at room temperature for 24 hours, and other harsh conditions.82,83 Current studies have confirmed that multiple lncRNAs are aberrantly expressed in NSCLC. The most fully evaluated candidate blood biomarker for NSCLC diagnosis is MALAT1.84 The expression of MALAT1 is downregulated in blood samples from NSCLC patients compared to those from healthy volunteers, with a sensitivity of 56% and a specificity of 96% as a lung cancer diagnosis marker.85 Moreover, the expression level of MALAT1 is also correlated with the presence of metastasis and the sites of metastatic lesions.85,86 However, the diagnostic value of MALAT1 alone is limited by the lack of significant differences in MALAT1 expression between adenocarcinoma and squamous cell carcinoma and the low diagnostic sensitivity of MALAT1 alone.85,87 Simultaneous detection of multiple lncRNAs can improve the sensitivity and specificity of the diagnosis.88 Similarly, serum GAS5 expression is significantly decreased in NSCLC patients compared with healthy volunteers, and it has a sensitivity of 82.2% and specificity of 72.7% as a marker to distinguish patients with lung cancer. When combined with CEA levels, the sensitivity and specificity are improved.89
In a recent study, 84 dysregulated lncRNAs in lung cancer were identified by comparing RNA sequencing data with TCGA data, among which 10 lncRNAs were meaningfully associated with patient survival. LINC01537 was considered the most important and its downregulation was verified in a validation analysis.90
LncRNAs in Lung Cancer Treatment
The clinical value of lncRNAs is not limited to their biomarker potential; they also have therapeutic aspects. LncRNAs are key regulators of many physiological and pathological processes, and they play a role in the regulation of gene expression and molecular pathways through interactions with proteins, miRNAs and other factors. Thus, lncRNAs may be a potential target for cancer treatment in the future.91 Targeting lncRNA therapy will be highly dependent on the function of lncRNAs, where advances in technologies offer the possibility of selectively targeting the expression of lncRNAs.92,93 For oncogenic lncRNAs, reducing their expression or affecting their functions can restore the sensitivity of cancer cells to antitumor therapy, inhibit the ability of cancer cells to invade and metastasize, or even achieve a cure for tumors. For example, inhibition of MALAT1 and HOTAIR expression in vivo by ASO effectively reduced metastasis and cancer cell migration or invasion in mice.25,94 Small-molecule inhibitors have provided us with another approach to regulate lncRNAs with the advantages of good solubility, bioavailability and metabolic stability compared to ASOs. Small molecule inhibitors target lncRNA-protein interactions by directly interfering with the secondary structure of lncRNAs or by masking the binding sites of their interactors.95–98 However, this approach requires a better understanding of RNA-protein interactions. Another direction is to use RNA interference-mediated gene silencing to selectively silence oncogenic lncRNAs.99,100 For tumor suppressor lncRNAs, there is currently no defined therapeutic targeting method, and in general, increasing their expression or enhancing their effects should be the direction of exploration. Approaches targeting lncRNAs are summarized in Table 2. Tumor progression was conspicuously impeded in triple-negative breast cancer and esophageal squamous cell carcinoma when siRNA nanoparticles were systematically administered in mouse model.101,102 Off-targeting is, however, now a major challenge for siRNAs.
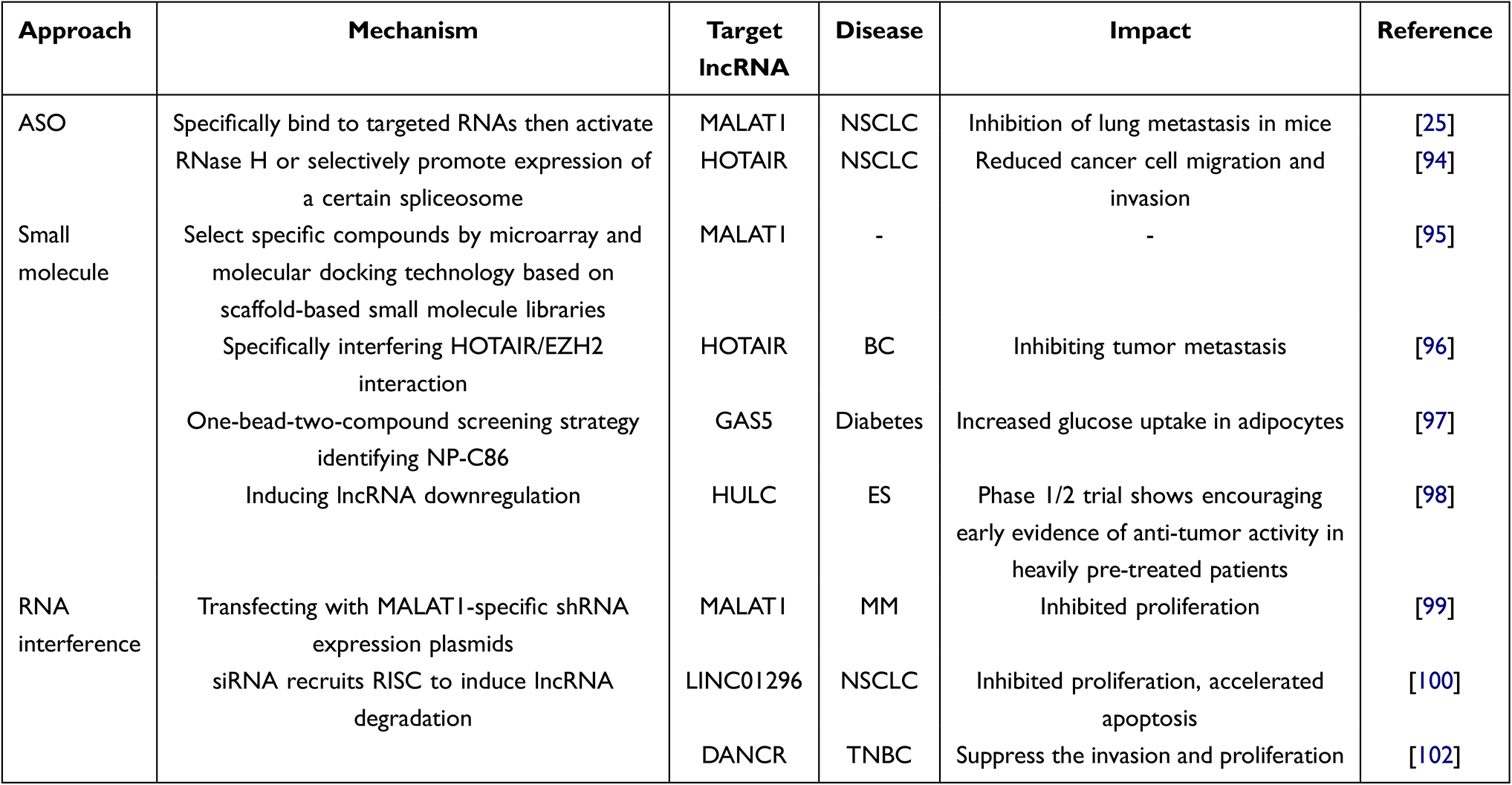 |
Table 2 Examples of Strategies in lncRNA-Based Therapies |
Overall, many common challenges remain in the development of directly targeting lncRNA therapy, including the lack of safe, specific and effective delivery methods, the absence of a defined optimal dosing regimen and the prevention of off-targeting. Although most of the researches on lncRNAs are still in the preclinical stage, researches on mitochondrial long non-coding RNA (mtlncRNA) have progressed rapidly and some are already in clinical trials. For example, a phase Ia trial evaluating Andes-1537, a short single-chain ASO, for the treatment of patients with advanced solid tumors (NCT02508441). The results showed that Andes-1537 was well tolerated, with no dose-limiting toxicities observed except for two injection site reactions in the high-dose group, and potential antitumor effects, with 2 patients (11.8%) had stable disease on scans beyond six months.103 Following the preliminary signals of efficacy observed in phase Ia trial, an open, multicentric, two stage phase Ib trial (NCT03985072) was initiated to investigate the safety and tolerability of the drug in cancer patients from 6 cohorts, associated to six different cancer type.
In addition to directly targeting lncRNA therapy, lncRNAs such as HOTAIR and H19 and their roles in tumor staging can also be used as biomarkers for reference in conventional therapy and patient monitoring. Gong et al evaluated the association between polymorphisms of some lncRNAs, the lung cancer risk and the patient response to treatment modalities in a Chinese population.104 Among them, polymorphisms of H19, MALAT1 and HOTAIR were associated with the patient response to platinum-containing chemotherapy. Subgroup analysis showed that ANRIL polymorphisms were associated with the response to platinum-based chemotherapy in adenocarcinoma patients, while ANRIL and H19 polymorphisms were associated with the treatment response in SCLC patients. In addition, polymorphisms within lncRNAs have been associated with the toxicity of platinum-based chemotherapy. For example, ANRIL polymorphisms are associated with severe overall toxicity and severe gastrointestinal toxicity, and MEG3 polymorphisms are associated with severe gastrointestinal toxicity.105
We have listed several lncRNAs that are the most promising for clinical use in lung cancer and their possible regulatory mechanisms. However, there are far more aberrant lncRNAs in lung cancer. Our previous study showed that LINC00460 is significantly upregulated and acts as an oncogene in NSCLC and is associated with a poor prognosis. Further study revealed that LINC00460 is involved in cell migration and invasion through physical interactions with hnRNP K.106 We also identified and characterized a fusion circRNA (F-circEA) produced from EML4-ALK-positive NSCLC that promotes cell migration and invasion.81 In addition, the currently discovered and validated lncRNAs can be used in combination, such as using lncRNAs+lncRNAs and lncRNAs+proteins, to form detection kits to improve their clinical value.
This review also has several limitations. Briefly, with more than 3000 recent reports of lncRNAs associated with lung cancer, we have highlighted only a few types that currently appear most likely to translate into clinical applications. Second, no systematic reviews and meta-analyses have been performed to quantify the value of lncRNAs as biomarkers and therapeutic targets, especially for targeted therapies, anti-angiogenic, and immunotherapies.
Conclusion
LncRNAs are biomarkers with great potential. In this review, we summarize the current findings of the most important lncRNAs and their use in the diagnosis and treatment of lung cancer. There is an urgent need for early diagnosis and alternative treatment options to improve patient prognosis. LncRNAs open up a new world. However, most lncRNAs remain elusive in terms of their mechanism of action. More in-depth studies on lncRNAs are urgently needed to pave the way for rational cancer diagnosis and treatment.
Abbreviations
5-AzadC, 5-Aza-2’-deoxycytidine; ASO, antisense oligonucleotides; CNEs, conserved noncoding elements; GAS5, growth arrest-specific transcript 5; H3K, HOTAIR triggered histone 3 lysine; HCC, hepatocellular carcinoma; hnRNP C, heterogeneous nuclear ribonucleoprotein C; HOTAIR, HOX transcript antisense RNA; HOXA5, Homeobox protein A5; ICIs, Immune checkpoint inhibitors; LncRNAs, long noncoding RNAs; LSD1, lysine-specific demethylase 1; MALAT1, metastasis-associated lung adenocarcinoma transcript 1; MEG3, maternally expressed gene 3; miRNA, microRNAs; MMP, matrix metalloproteinase; NEAT2, noncoding nuclear-enriched abundant transcript 2; NSCLC, non-small-cell lung cancer; PRC2, polycomb repressive complex 2; qRT–PCR, quantitative reverse transcription PCR; SCLC, small cell lung cancer; SPRY4-IT1, sprouty 4 intronic transcript 1; SR, serine/arginine.
Data Sharing Statement
Not applicable to this study.
Ethics Approval and Consent to Participate
This report did not meet criteria for IRB approval.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Youth Program of National Natural Science Foundation of China (81902723).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 Countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
2. Brahmer JR, Tykodi SS, Chow LQ, et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N Engl J Med. 2012;366(26):2455–2465. doi:10.1056/NEJMoa1200694
3. Roth A, Diederichs S. Long noncoding RNAs in lung cancer. Curr Top Microbiol Immunol. 2016;394:57–110. doi:10.1007/82_2015_444
4. Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–159. doi:10.1038/nrg2521
5. Zhang K, Shi ZM, Chang YN, Hu ZM, Qi HX, Hong W. The ways of action of long non-coding RNAs in cytoplasm and nucleus. Gene. 2014;547(1):1–9. doi:10.1016/j.gene.2014.06.043
6. Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. doi:10.1016/j.cell.2009.02.006
7. Sati S, Ghosh S, Jain V, Scaria V, Sengupta S. Genome-wide analysis reveals distinct patterns of epigenetic features in long non-coding RNA loci. Nucleic Acids Res. 2012;40(20):10018–10031. doi:10.1093/nar/gks776
8. Khalil AM, Guttman M, Huarte M, et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc Natl Acad Sci U S A. 2009;106(28):11667–11672. doi:10.1073/pnas.0904715106
9. Cech TR, Steitz JA. The noncoding RNA revolution-trashing old rules to forge new ones. Cell. 2014;157(1):77–94. doi:10.1016/j.cell.2014.03.008
10. Pang KC, Frith MC, Mattick JS. Rapid evolution of noncoding RNAs: lack of conservation does not mean lack of function. Trends Genet. 2006;22(1):1–5. doi:10.1016/j.tig.2005.10.003
11. Kim TK, Hemberg M, Gray JM, et al. Widespread transcription at neuronal activity-regulated enhancers. Nature. 2010;465(7295):182–187. doi:10.1038/nature09033
12. Huang T, Wang G, Yang L, et al. Transcription factor YY1 modulates lung cancer progression by activating lncRNA-PVT1. DNA Cell Biol. 2017;36(11):947–958. doi:10.1089/dna.2017.3857
13. Yao RW, Wang Y, Chen LL. Cellular functions of long noncoding RNAs. Nat Cell Biol. 2019;21(5):542–551. doi:10.1038/s41556-019-0311-8
14. Hutchinson JN, Ensminger AW, Clemson CM, Lynch CR, Lawrence JB, Chess A. A screen for nuclear transcripts identifies two linked noncoding RNAs associated with SC35 splicing domains. BMC Genomics. 2007;8(1):39. doi:10.1186/1471-2164-8-39
15. Wu Y, Huang C, Meng X, Li J. Long noncoding RNA MALAT1: insights into its biogenesis and implications in human disease. Curr Pharm Des. 2015;21(34):5017–5028. doi:10.2174/1381612821666150724115625
16. Yoshimoto R, Mayeda A, Yoshida M, Nakagawa S. MALAT1 long non-coding RNA in cancer. Biochim Biophys Acta. 2016;1859(1):192–199. doi:10.1016/j.bbagrm.2015.09.012
17. Gutschner T, Hammerle M, Diederichs S. MALAT1 – a paradigm for long noncoding RNA function in cancer. J Mol Med. 2013;91(7):791–801. doi:10.1007/s00109-013-1028-y
18. Ji P, Diederichs S, Wang W, et al. MALAT-1, a novel noncoding RNA, and thymosin beta 4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene. 2003;22(39):8031–8041. doi:10.1038/sj.onc.1206928
19. Zhang H-M, Yang F-Q, Chen S-J, Che J, Zheng J-H. Upregulation of long non-coding RNA MALAT1 correlates with tumor progression and poor prognosis in clear cell renal cell carcinoma. Tumor Biol. 2015;36(4):2947–2955. doi:10.1007/s13277-014-2925-6
20. Pang E-J, Yang R, X-b F, Y-f L. Overexpression of long non-coding RNA MALAT1 is correlated with clinical progression and unfavorable prognosis in pancreatic cancer. Tumor Biol. 2015;36(4):2403–2407. doi:10.1007/s13277-014-2850-8
21. Lai MC, Yang Z, Zhou L, et al. Long non-coding RNA MALAT-1 overexpression predicts tumor recurrence of hepatocellular carcinoma after liver transplantation. Med Oncol. 2012;29(3):1810–1816. doi:10.1007/s12032-011-0004-z
22. Karimpour M, Ravanbakhsh R, Maydanchi M, Rajabi A, Azizi F, Saber A. Cancer driver gene and non-coding RNA alterations as biomarkers of brain metastasis in lung cancer: a review of the literature. Biomed Pharmacother. 2021;143:112190. doi:10.1016/j.biopha.2021.112190
23. Gutschner T, Diederichs S. The hallmarks of cancer: a long non-coding RNA point of view. RNA Biol. 2012;9(6):703–719. doi:10.4161/rna.20481
24. Tano K, Mizuno R, Okada T, et al. MALAT-1 enhances cell motility of lung adenocarcinoma cells by influencing the expression of motility-related genes. FEBS Lett. 2010;584(22):4575–4580. doi:10.1016/j.febslet.2010.10.008
25. Gutschner T, Hammerle M, Eissmann M, et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 2013;73(3):1180–1189. doi:10.1158/0008-5472.CAN-12-2850
26. Shen L, Chen L, Wang Y, Jiang X, Xia H, Zhuang Z. Long noncoding RNA MALAT1 promotes brain metastasis by inducing epithelial-mesenchymal transition in lung cancer. J Neurooncol. 2015;121(1):101–108. doi:10.1007/s11060-014-1613-0
27. Li J, Wang J, Chen Y, et al. LncRNA MALAT1 exerts oncogenic functions in lung adenocarcinoma by targeting miR-204. Am J Cancer Res. 2016;6(5):1099–1107.
28. Tripathi V, Ellis JD, Shen Z, et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol Cell. 2010;39(6):925–938. doi:10.1016/j.molcel.2010.08.011
29. Tripathi V, Shen Z, Chakraborty A, et al. Long noncoding RNA MALAT1 controls cell cycle progression by regulating the expression of oncogenic transcription factor B-MYB. PLoS Genet. 2013;9(3):e1003368. doi:10.1371/journal.pgen.1003368
30. Yang L, Lin C, Liu W, et al. ncRNA- and Pc2 methylation-dependent gene relocation between nuclear structures mediates gene activation programs. Cell. 2011;147(4):773–788. doi:10.1016/j.cell.2011.08.054
31. He S, Liu S, Zhu H. The sequence, structure and evolutionary features of HOTAIR in mammals. BMC Evol Biol. 2011;11(1):102. doi:10.1186/1471-2148-11-102
32. Nepal C, Taranta A, Hadzhiev Y, et al. Ancestrally duplicated conserved noncoding element suggests dual regulatory roles of HOTAIR in cis and trans. iScience. 2020;23(4):101008. doi:10.1016/j.isci.2020.101008
33. Rinn JL, Kertesz M, Wang JK, et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129(7):1311–1323. doi:10.1016/j.cell.2007.05.022
34. Tsai MC, Manor O, Wan Y, et al. Long noncoding RNA as modular scaffold of histone modification complexes. Science. 2010;329(5992):689–693. doi:10.1126/science.1192002
35. Gupta RA, Shah N, Wang KC, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464(7291):1071–1076. doi:10.1038/nature08975
36. Liu XH, Sun M, Nie FQ, et al. Lnc RNA HOTAIR functions as a competing endogenous RNA to regulate HER2 expression by sponging miR-331-3p in gastric cancer. Mol Cancer. 2014;13(1):92. doi:10.1186/1476-4598-13-92
37. Ishibashi M, Kogo R, Shibata K, et al. Clinical significance of the expression of long non-coding RNA HOTAIR in primary hepatocellular carcinoma. Oncol Rep. 2013;29(3):946–950. doi:10.3892/or.2012.2219
38. Kogo R, Shimamura T, Mimori K, et al. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res. 2011;71(20):6320–6326. doi:10.1158/0008-5472.CAN-11-1021
39. Nakagawa T, Endo H, Yokoyama M, et al. Large noncoding RNA HOTAIR enhances aggressive biological behavior and is associated with short disease-free survival in human non-small cell lung cancer. Biochem Biophys Res Commun. 2013;436(2):319–324. doi:10.1016/j.bbrc.2013.05.101
40. Liu MY, Li XQ, Gao TH, et al. Elevated HOTAIR expression associated with cisplatin resistance in non-small cell lung cancer patients. J Thorac Dis. 2016;8(11):3314–3322. doi:10.21037/jtd.2016.11.75
41. Liu Z, Sun M, Lu K, et al. The long noncoding RNA HOTAIR contributes to cisplatin resistance of human lung adenocarcinoma cells via downregualtion of p21(WAF1/CIP1) expression. PLoS One. 2013;8(10):e77293. doi:10.1371/journal.pone.0077293
42. Yang Y, Jiang C, Yang Y, et al. Silencing of LncRNA-HOTAIR decreases drug resistance of non-small cell lung cancer cells by inactivating autophagy via suppressing the phosphorylation of ULK1. Biochem Biophys Res Commun. 2018;497(4):1003–1010. doi:10.1016/j.bbrc.2018.02.141
43. Fang S, Gao H, Tong Y, et al. Long noncoding RNA-HOTAIR affects chemoresistance by regulating HOXA1 methylation in small cell lung cancer cells. Lab Invest. 2016;96(1):60–68. doi:10.1038/labinvest.2015.123
44. Leibovitch MP, Nguyen VC, Gross MS, Solhonne B, Leibovitch SA, Bernheim A. The human ASM (adult skeletal muscle) gene: expression and chromosomal assignment to 11p15. Biochem Biophys Res Commun. 1991;180(3):1241–1250. doi:10.1016/S0006-291X(05)81329-4
45. Liu F, Killian JK, Yang M, et al. Epigenomic alterations and gene expression profiles in respiratory epithelia exposed to cigarette smoke condensate. Oncogene. 2010;29(25):3650–3664. doi:10.1038/onc.2010.129
46. Zhang Y, Lu Y, Yuan BZ, et al. The Human mineral dust-induced gene, mdig, is a cell growth regulating gene associated with lung cancer. Oncogene. 2005;24(31):4873–4882. doi:10.1038/sj.onc.1208668
47. Ghafouri-Fard S, Esmaeili M, Taheri M. H19 lncRNA: roles in tumorigenesis. Biomed Pharmacother. 2020;123:109774. doi:10.1016/j.biopha.2019.109774
48. Qian B, Wang DM, Gu XS, et al. LncRNA H19 serves as a ceRNA and participates in non-small cell lung cancer development by regulating microRNA-107. Eur Rev Med Pharmacol Sci. 2018;22(18):5946–5953. doi:10.26355/eurrev_201809_15925
49. Huang T, Wen Y, Peng B, Ding G, Yang L, Wang Z. Upregulated lncRNA H19 promotes non-small cell lung cancer cell proliferation through miR-138/PDK1 axis. Int J Clin Exp Pathol. 2017;10(8):9012–9020.
50. Zhao Y, Feng C, Li Y, Ma Y, Cai R. LncRNA H19 promotes lung cancer proliferation and metastasis by inhibiting miR-200a function. Mol Cell Biochem. 2019;460(1–2):1–8. doi:10.1007/s11010-019-03564-1
51. Liu L, Liu L, Lu S. lncRNA H19 promotes viability and epithelial-mesenchymal transition of lung adenocarcinoma cells by targeting miR-29b-3p and modifying STAT3. Int J Oncol. 2019;54(3):929–941. doi:10.3892/ijo.2019.4695
52. Matouk IJ, DeGroot N, Mezan S, et al. The H19 non-coding RNA is essential for human tumor growth. PLoS One. 2007;2(9):e845.
53. Matouk IJ, Mezan S, Mizrahi A, et al. The oncofetal H19 RNA connection: hypoxia, p53 and cancer. Biochimica et Biophysica Acta Mol Cell Res. 2010;1803(4):443–451. doi:10.1016/j.bbamcr.2010.01.010
54. Zhang E, Li W, Yin D, et al. c-Myc-regulated long non-coding RNA H19 indicates a poor prognosis and affects cell proliferation in non-small-cell lung cancer. Tumour Biol. 2016;37(3):4007–4015. doi:10.1007/s13277-015-4185-5
55. Zhang X, Zhou Y, Mehta KR, et al. A pituitary-derived MEG3 isoform functions as a growth suppressor in tumor cells. J Clin Endocrinol Metab. 2003;88(11):5119–5126. doi:10.1210/jc.2003-030222
56. Zhou Y, Zhong Y, Wang Y, et al. Activation of p53 by MEG3 non-coding RNA. J Biol Chem. 2007;282(34):24731–24742. doi:10.1074/jbc.M702029200
57. Benetatos L, Vartholomatos G, Hatzimichael E. MEG3 imprinted gene contribution in tumorigenesis. Int J Cancer. 2011;129(4):773–779. doi:10.1002/ijc.26052
58. Zhao Y, Zhu Z, Shi S, Wang J, Li N. Long non-coding RNA MEG3 regulates migration and invasion of lung cancer stem cells via miR-650/SLC34A2 axis. Biomed Pharmacother. 2019;120:109457. doi:10.1016/j.biopha.2019.109457
59. Liu J, Wan L, Lu K, et al. The long noncoding RNA MEG3 contributes to cisplatin resistance of human lung adenocarcinoma. PLoS One. 2015;10(5):e0114586. doi:10.1371/journal.pone.0114586
60. Xia H, Qu XL, Liu LY, Qian DH, Jing HY. LncRNA MEG3 promotes the sensitivity of vincristine by inhibiting autophagy in lung cancer chemotherapy. Eur Rev Med Pharmacol Sci. 2018;22(4):1020–1027. doi:10.26355/eurrev_201802_14384
61. Gong X, Huang MY. Tumor-suppressive function of lncRNA-MEG3 in glioma cells by regulating miR-6088/SMARCB1 axis. Biomed Res Int. 2020;2020:4309161. doi:10.1155/2020/4309161
62. Zhao J, Dahle D, Zhou Y, Zhang X, Klibanski A. Hypermethylation of the promoter region is associated with the loss of MEG3 gene expression in human pituitary tumors. J Clin Endocrinol Metab. 2005;90(4):2179–2186. doi:10.1210/jc.2004-1848
63. Zhou Y, Zhang X, Klibanski A. MEG3 noncoding RNA: a tumor suppressor. J Mol Endocrinol. 2012;48(3):R45–53. doi:10.1530/JME-12-0008
64. Li J, Bian EB, He XJ, et al. Epigenetic repression of long non-coding RNA MEG3 mediated by DNMT1 represses the p53 pathway in gliomas. Int J Oncol. 2016;48(2):723–733. doi:10.3892/ijo.2015.3285
65. Zhou C, Huang C, Wang J, et al. LncRNA MEG3 downregulation mediated by DNMT3b contributes to nickel malignant transformation of human bronchial epithelial cells via modulating PHLPP1 transcription and HIF-1alpha translation. Oncogene. 2017;36(27):3878–3889. doi:10.1038/onc.2017.14
66. Schneider C, King RM, Philipson L. Genes specifically expressed at growth arrest of mammalian cells. Cell. 1988;54(6):787–793. doi:10.1016/S0092-8674(88)91065-3
67. Coccia EM, Cicala C, Charlesworth A, et al. Regulation and expression of a growth arrest-specific gene (gas5) during growth, differentiation, and development. Mol Cell Biol. 1992;12(8):3514–3521. doi:10.1128/mcb.12.8.3514-3521.1992
68. Smith CM, Steitz JA. Classification of gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member of the 5’-terminal oligopyrimidine gene family reveals common features of snoRNA host genes. Mol Cell Biol. 1998;18(12):6897–6909. doi:10.1128/MCB.18.12.6897
69. Mourtada-Maarabouni M, Pickard MR, Hedge VL, Farzaneh F, Williams GT. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer. Oncogene. 2009;28(2):195–208. doi:10.1038/onc.2008.373
70. Kino T, Hurt DE, Ichijo T, Nader N, Chrousos GP. Noncoding RNA gas5 is a growth arrest- and starvation-associated repressor of the glucocorticoid receptor. Sci Signal. 2010;3(107):ra8. doi:10.1126/scisignal.2000568
71. Qiao HP, Gao WS, Huo JX, Yang ZS. Long non-coding RNA GAS5 functions as a tumor suppressor in renal cell carcinoma. Asian Pac J Cancer Prev. 2013;14(2):1077–1082. doi:10.7314/APJCP.2013.14.2.1077
72. Shi X, Sun M, Liu H, et al. A critical role for the long non-coding RNA GAS5 in proliferation and apoptosis in non-small-cell lung cancer. Mol Carcinog. 2015;54(Suppl 1):E1–E12. doi:10.1002/mc.22120
73. Leeksma OC, Van Achterberg TA, Tsumura Y, et al. Human sprouty 4, a new ras antagonist on 5q31, interacts with the dual specificity kinase TESK1. Eur J Biochem. 2002;269(10):2546–2556. doi:10.1046/j.1432-1033.2002.02921.x
74. Tennis MA, Van Scoyk MM, Freeman SV, Vandervest KM, Nemenoff RA, Winn RA. Sprouty-4 inhibits transformed cell growth, migration and invasion, and epithelial-mesenchymal transition, and is regulated by Wnt7A through PPARgamma in non-small cell lung cancer. Mol Cancer Res. 2010;8(6):833–843. doi:10.1158/1541-7786.MCR-09-0400
75. Sun M, Liu XH, Lu KH, et al. EZH2-mediated epigenetic suppression of long noncoding RNA SPRY4-IT1 promote s NSCLC cell proliferation and metastasis by affecting the epithelial–mesenchymal transition. Cell Death Dis. 2014;5(6):e1298. doi:10.1038/cddis.2014.256
76. Wen X, Han XR, Wang YJ, et al. Effects of long noncoding RNA SPRY4-IT1-mediated EZH2 on the invasion and migration of lung adenocarcinoma. J Cell Biochem. 2018;119(2):1827–1840. doi:10.1002/jcb.26344
77. Khaitan D, Dinger ME, Mazar J, et al. The melanoma-upregulated long noncoding RNA SPRY4-IT1 modulates apoptosis and invasion. Cancer Res. 2011;71(11):3852–3862. doi:10.1158/0008-5472.CAN-10-4460
78. Li H, Liu C, Lu Z, et al. Upregulation of the long non-coding RNA SPRY4-IT1 indicates a poor prognosis and promotes tumorigenesis in ovarian cancer. Biomed Pharmacother. 2017;88:529–534.
79. Liu D, Li Y, Luo G, et al. LncRNA SPRY4-IT1 sponges miR-101-3p to promote proliferation and metastasis of bladder cancer cells through up-regulating EZH2. Cancer Lett. 2017;388:281–291. doi:10.1016/j.canlet.2016.12.005
80. Tan W, Song ZZ, Xu Q, et al. Up-regulated expression of SPRY4-IT1 predicts poor prognosis in colorectal cancer. Med Sci Monit. 2017;23:309–314. doi:10.12659/MSM.898369
81. Tan S, Gou Q, Pu W, et al. Circular RNA F-circEA produced from EML4-ALK fusion gene as a novel liquid biopsy biomarker for non-small cell lung cancer. Cell Res. 2018;28(6):693–695. doi:10.1038/s41422-018-0033-7
82. Arita T, Ichikawa D, Konishi H, et al. Circulating long non-coding RNAs in plasma of patients with gastric cancer. Anticancer Res. 2013;33(8):3185–3193. doi:10.3892/br.2015.422
83. Hewson C, Morris KV. Form and function of exosome-associated long non-coding RNAs in cancer. Curr Top Microbiol Immunol. 2016;394:41–56. doi:10.1007/82_2015_486
84. Pan J, Bian Y, Cao Z, et al. Long noncoding RNA MALAT1 as a candidate serological biomarker for the diagnosis of non-small cell lung cancer: a meta-analysis. Thorac Cancer. 2020;11(2):329–335. doi:10.1111/1759-7714.13265
85. Guo F, Yu F, Wang J, et al. Expression of MALAT1 in the peripheral whole blood of patients with lung cancer. Biomed Rep. 2015;3(3):309–312.
86. Zhang R, Xia Y, Wang Z, et al. Serum long non coding RNA MALAT-1 protected by exosomes is up-regulated and promotes cell proliferation and migration in non-small cell lung cancer. Biochem Biophys Res Commun. 2017;490(2):406–414. doi:10.1016/j.bbrc.2017.06.055
87. Weber DG, Johnen G, Casjens S, et al. Evaluation of long noncoding RNA MALAT1 as a candidate blood-based biomarker for the diagnosis of non-small cell lung cancer. BMC Res Notes. 2013;6(1):518. doi:10.1186/1756-0500-6-518
88. Yuan S, Xiang Y, Guo X, et al. Circulating long noncoding RNAs act as diagnostic biomarkers in non-small cell lung cancer. Front Oncol. 2020;10:537120. doi:10.3389/fonc.2020.537120
89. Liang W, Lv T, Shi X, et al. Circulating long noncoding RNA GAS5 is a novel biomarker for the diagnosis of nonsmall cell lung cancer. Medicine. 2016;95(37):e4608. doi:10.1097/MD.0000000000004608
90. Gong W, Yang L, Wang Y, et al. Analysis of survival-related lncRNA landscape identifies A role for LINC01537 in energy metabolism and lung cancer progression. Int J Mol Sci. 2019;20(15):3713. doi:10.3390/ijms20153713
91. Zhou M, Zhong L, Xu W, et al. Discovery of potential prognostic long non-coding RNA biomarkers for predicting the risk of tumor recurrence of breast cancer patients. Sci Rep. 2016;6(1):31038. doi:10.1038/srep31038
92. Shan G. RNA interference as a gene knockdown technique. Int J Biochem Cell Biol. 2010;42(8):1243–1251. doi:10.1016/j.biocel.2009.04.023
93. Kole R, Krainer AR, Altman S. RNA therapeutics: beyond RNA interference and antisense oligonucleotides. Nat Rev Drug Discov. 2012;11(2):125–140. doi:10.1038/nrd3625
94. Liang H, Peng J. LncRNA HOTAIR promotes proliferation, invasion and migration in NSCLC cells via the CCL22 signaling pathway. PLoS One. 2022;17(2):e0263997. doi:10.1371/journal.pone.0263997
95. Donlic A, Zafferani M, Padroni G, Puri M, Hargrove AE. Regulation of MALAT1 triple helix stability and in vitro degradation by diphenylfurans. Nucleic Acids Res. 2020;48(14):7653–7664. doi:10.1093/nar/gkaa585
96. Ren Y, Wang YF, Zhang J, et al. Targeted design and identification of AC1NOD4Q to block activity of HOTAIR by abrogating the scaffold interaction with EZH2. Clin Epigenetics. 2019;11(1):29. doi:10.1186/s13148-019-0624-2
97. Shi Y, Parag S, Patel R, et al. Stabilization of lncRNA GAS5 by a small molecule and its implications in diabetic adipocytes. Cell Chem Biol. 2019;26(3):319–330 e316. doi:10.1016/j.chembiol.2018.11.012
98. Ludwig JA, Federman NC, Anderson P, et al. TK216 for relapsed/refractory Ewing sarcoma: interim Phase 1/2 results. J Clin Oncol. 2021;39(15_suppl):11500. doi:10.1200/JCO.2021.39.15_suppl.11500
99. Liu H, Wang H, Wu B, et al. Down-regulation of long non-coding RNA MALAT1 by RNA interference inhibits proliferation and induces apoptosis in multiple myeloma. Clin Exp Pharmacol Physiol. 2017;44(10):1032–1041. doi:10.1111/1440-1681.12804
100. Xu L, Wei B, Hui H, et al. Positive feedback loop of lncRNA LINC01296/miR-598/Twist1 promotes non-small cell lung cancer tumorigenesis. J Cell Physiol. 2019;234(4):4563–4571. doi:10.1002/jcp.27235
101. Liang Y, Chen X, Wu Y, et al. LncRNA CASC9 promotes esophageal squamous cell carcinoma metastasis through upregulating LAMC2 expression by interacting with the CREB-binding protein. Cell Death Differ. 2018;25(11):1980–1995. doi:10.1038/s41418-018-0084-9
102. Vaidya AM, Sun Z, Ayat N, et al. Systemic delivery of tumor-targeting siRNA nanoparticles against an oncogenic LncRNA facilitates effective triple-negative breast cancer therapy. Bioconjug Chem. 2019;30(3):907–919. doi:10.1021/acs.bioconjchem.9b00028
103. Dhawan MS, Aggarwal RR, Boyd E, Comerford K, Munster PN. Phase 1 study of Andes-1537: a novel antisense oligonucleotide against non-coding mitochondrial DNA in advanced solid tumors. J Clin Oncol. 2018;36(15_suppl):2557. doi:10.1200/JCO.2018.36.15_suppl.2557
104. Gong WJ, Yin JY, Li XP, et al. Association of well-characterized lung cancer lncRNA polymorphisms with lung cancer susceptibility and platinum-based chemotherapy response. Tumour Biol. 2016;37(6):8349–8358. doi:10.1007/s13277-015-4497-5
105. Gong WJ, Peng JB, Yin JY, et al. Association between well-characterized lung cancer lncRNA polymorphisms and platinum-based chemotherapy toxicity in Chinese patients with lung cancer. Acta Pharmacol Sin. 2017;38(4):581–590. doi:10.1038/aps.2016.164
106. Li K, Sun D, Gou Q, et al. Long non-coding RNA linc00460 promotes epithelial-mesenchymal transition and cell migration in lung cancer cells. Cancer Lett. 2018;420:80–90. doi:10.1016/j.canlet.2018.01.060
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.