")
Back to Journals » Cancer Management and Research » Volume 11
lncRNA WT1-AS inhibits the aggressiveness of cervical cancer cell via regulating p53 expression via sponging miR-330-5p
Authors Cui LJ, Nai MM, Zhang K, Li L, Li RM
Received 6 June 2018
Accepted for publication 27 September 2018
Published 11 January 2019 Volume 2019:11 Pages 651—667
DOI https://doi.org/10.2147/CMAR.S176525
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Harikrishna Nakshatri
LiJuan Cui,1,* ManMan Nai,2,* Ke Zhang,1 Lu Li,3 RuiMin Li4
1Department of Gynecology, First People’s Hospital of Jiaozuo City, Jiaozuo, Henan Province, China; 2Department of Gynecology, The Third Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan Province, China; 3Department of Gynecology, Second People’s Hospital of Jiaozuo City, Jiaozuo, Henan Province, China; 4Department of Gynecology, Jiaozuo Maternal and Child Care Service Center, Jiaozuo, Henan Province, China
*These authors contributed equally to this work
Background: Emerging evidences have demonstrated that lncRNAs play vital roles in various pathological processes, including cancer. The lncRNA WT1 antisense RNA (WT1-AS) serves as a tumor suppressor in various cancers. Nevertheless, the expression and precise function of WT1-AS in cervical carcinoma still remain not yet investigated. The objective of our study was to explore the expression of WT1-AS and its biological roles in cervical cancer.
Methods: Differences in the lncRNA expression profiles between cervical cancer and adjacent normal tissues were assessed by lncRNA expression microarray analysis. The expression of p53 in cervical cancer cell was assessed by qRT-PCR and immunofluorescence assay. Loss-of-function studies were used to explore the effect of lncRNA WT1-AS on the growth and metastasis of cervical cancer cell in vitro and in vivo.
Results: Our results demonstrated that WT1-AS was remarkably down-regulated in cervical carcinoma. Functional assays proved that up-regulation of WT1-AS significantly suppressed cervical cancer cell proliferation, migration and invasion. In addition, the luciferase reporter assay identified that miR-330-5p was the target of WT1-AS. Moreover, tumor suppressor p53 was identified as the direct target of miR-330-5p and alternation of miR-330-5p/p53 axis reversed the effects of WT1-AS in cervical cancer cell.
Conclusion: Altogether, our findings suggested that WT1-AS was down-regulated in cervical carcinoma and WT1-AS suppressed cervical carcinoma cell- proliferation, migration and invasion through regulating the miR-330-5p/p53 axis.
Keywords: cervical cancer, WT1-AS, miR-330-5p, p53, metastasis
Introduction
Cervical cancer is one of the most deadly cancers, with its incidence increased drastically worldwide.1 Once cervical cancer cell disseminates to distant and internal organs, cervical cancer will be almost resistant to the current clinical treatments and becomes an incurable disease.2 Hence, it is noteworthy that the occurrence of cancer cell metastasis facilitates cervical cancer to be an aggressive and life-threatening disease, and the identification of metastasis-related biomarkers and therapeutic targets is urgent for the treatment of cervical cancer. Recently, accumulating evidence suggest that lncRNAs act as important modulators in the tumorigenesis and progression of cervical carcinoma and may become innovative treatment targets.3 Currently, evidence demonstrate that lncRNAs have crucial actions in various cellular processes, including cancer cell growth, epithelial–mesenchymal transition (EMT), and metastasis.4 Studies reveal that lncRNAs are commonly dysexpressed in multiple types of cancer. Few studies prove that lncRNAs mediate genes expression at transcriptional level, posttranscriptional level, or epigenetic level.5 In addition, extensive lncRNAs act via ceRNA for miRNA targets, thereby preventing miRNAs from binding to their target genes.6
FENDRR, which locates at chr3q13.31, consists of four exons and has a length of 3,099 nucleotides.7 A previous study identifies that the low level of FENDRR is related to the poor prognosis of patients with gastric carcinoma and overregulation of FENDRR suppresses the migration and invasion capacities of gastric carcinoma cell via downregulating the expression of matrix metalloproteinase 2/9 and fibronectin1.8 In addition, FENDRR suppresses breast cancer cell growth, promotes cell apoptosis, and is closely related to the prognosis of patients with breast carcinoma.9 In prostate cancer, the level of FENDRR is remarkably decreased when compared to normal control, and the downregulation of FENDRR is related to the poor prognosis of patients.10 The expression of lncRNA HOTAIR is elevated in a variety of tumors and are closely associated with poor survival and high recurrence rates of tumor.11–13 Downregulation of HOTAIR suppresses cancer cell proliferation and inhibits cell metastasis. Similarly, more lncRNAs are being identified and many are need to thoroughly functionally investigated in cancers.14 WT1 antisense RNA (WT1-AS) is one of the antisense transcripts of the Wilm’s tumor gene, which encodes a zinc finger transcription domain.15 Recently, WT1-AS has proved to be aberrantly dysexpressed in several tumors, including ovarian clear cell adenocarcinoma (OCCA).16–18 The level of WT1-AS is decreased in gastric cancer tissue compared with the nontumor tissue. The growth, migration, and invasion of gastric cancer cell are inhibited when WT1-AS was upregulated in gastric cancer cell.19 Furthermore, the promoter of WT1-AS has been proved to be markedly methylated in OCCA when compared with the serous adenocarcinoma, which suggest the potential role of WT1-AS in prognosis.16 However, the potential roles of lncRNA WT1-AS in cervical cancer invasion and metastasis have not yet been fully investigated.
In this study, we found that WT1-AS was markedly downregulated in cervical cancer, including clinical tissue and cell line. With overregulation of WT1-AS, cell growth and invasion were significantly inhibited and tumor growth was inhibited. Furthermore, we demonstrated that overregulation of WT1-AS significantly decreased the level of miR-330-5p in cervical carcinoma cell. Bioinformatics analysis and the luciferase reporter assays identified that miR-330-5p bound to WT1-AS in a sequence-specific manner. In mechanism, miR-330-5p targeted directly the mRNA of p53 in cervical cancer cell. The viral oncoprotein E6 is an essential factor for cervical cancer induced by “high-risk” mucosal HPV. Among other oncogenic activities, E6 recruits the ubiquitin ligase E6AP to promote the ubiquitination and subsequent proteasomal degradation of p53.20 As expected, we found that overregulation of miR-330-5p increased the expression of HPV E6 in cervical cancer cell, indicating that miR-330-5p inhibited the expression of p53 and was partly associated with it, regulating the level of HPV E6. Finally, we identified that WT1-AS inhibited cervical cancer cell progression and regulated p53 expression via sponging miR-330-5p. Altogether, we demonstrated that WT1-AS/miR-330-5p/p53 axis regulated the growth, migration, and invasion of cervical cancer cell, which might function as a potential target for cervical cancer treatment.
Materials and methods
Cervical cancer tissues and cell culture
The frozen human cervical cancer tissues and paratumor tissues were obtained from First People’s Hospital of Jiaozuo City. These obtained tissue specimens were utilized to generate tissue section slides and tissue microarrays for research purposes. The cervical carcinoma cells (SiHa, C-4-I, C-33-A, and Ca-Ski) and the normal cervical epithelial cell line, ECT1/E6E7, were obtained from the GuangZhou Jennio Biotech Co., Ltd (GuangZhou, GuangDong, China). Cancer cells were cultured in RPMI-1640 or DMEM containing 10% FBS (Wisent, Quebec, Canada), 100 µg/mL of streptomycin, and 100 µg/mL of penicillin and maintained in an incubator with a humidified atmosphere of 95% air and 5% CO2 at 37°C. This research was approved by the Institutional Research Committee of the First People’s Hospital of Jiaozuo City, and the informed consent was obtained from all patients. The written informed consent for participation in the study was obtained from all patients before participating in this study.
Cells’ transfections
miR-330-5p mimics, miR-330-5p inhibitor, and their controls (miR-NC or miR-NC inhibitor) were purchased from Ambion (GeneCopoeia, Guangzhou, China). Lentiviral vector encoding WT1-AS cDNA (LV-WT1-AS) was constructed by Genepharma Co., Ltd (Shanghai, China). shRNA-WT1-AS was chemically synthesized by Realgene (Nanjing, Jiangsu, China). The shRNA targeting p53 was designed by Genepharma Co., Ltd. The lentiviral vector encoding p53 cDNA was constructed by Genepharma Co., Ltd, and named as LV-p53. The empty vector was control (named as LV-vector). In brief, cells were maintained around 50–60% confluence in 10 cm culture dishes when transfected with plasmids. Transfection of WT1-AS or miRNAs was conducted using the Lipofectamine RNAiMAX transfection reagent (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocol. To establish stable cell lines, media were refreshed on transfected cells 4 days later with media containing puromycin (1.5 µg/mL). Transfected cells were treated with puromycin, refreshed every 3 days, for 2 weeks.
Luciferase reporter assay
HEK-293T cells (1×105) were seeded in a six-well plate at 70%–80% confluence. The luciferase reporter vectors for the wild-type (wt) or mutant (mut) p53 3′-UTR containing miR-330-5p binding site were constructed into pMIR-REPORT luciferase system (Thermo Fisher Scientific). The vectors expressing firefly luciferase reporter fused with wt or mut p53 3′-UTR were co-transfected with miR-330-5p or miR-NC into cell. After 24 hours, cells were harvested, and the two distinct luciferase activities were measured using Dual-luciferase reporter assay according to the manufacturer’s instructions (Promega Corporation, Fitchburg, WI, USA). WT1-AS containing the predicted miR-330-5p-binding site was amplified using PCR and subcloned into a pmirGLO luciferase Target Expression Vector (Promega Corporation) to form the WT1-AS wt (pmirGLO-WT1-AS-wt) vector. The mutated miR-330-5p-binding sequence was constructed that was named as pmirGLO-WT1-AS-mut vector. The HEK-293T cells were co-transfected with PrirGLO, pmirGLO-WT1-AS-wt, prirGLO-WT1-AS-mut, and miR-330-5p mimics or negative control using Lipofectamine 2000, and the relative luciferase activity was measured using the Dual-Luciferase Reporter Assay Kit (Promega Corporation) after 24 hours.
MTT assay
Cells (1×103) were seeded in 96-well culture plates and cultured for 1, 2, 3, or 4 days. A total of 5 mg/mL MTT was added into 96-well culture plates after each day of cell growth. Then, the cells were continually incubated for 4 hours. Then, all cell supernatant was removed, dimethyl sulfoxide (DMSO) (200 µL) was added into plate, and the OD values were detected at 490 nm.
Wound-healing analysis
Cells (1×105) were seeded into a six-well plate and cultured for 24 hours. Then, the cells were treated with 10 µg/mL Mitomycin C (BP25312; Thermo Fisher Scientific) in culture media for 2 hours to eliminate proliferation. The monolayer cells were scratched by using a 100 µL tip, and then, the cells were cultured in serum-free medium. The image of wound closure at 0 and 24 hours was recorded under a microscope. The percentage of wound closure was calculated using ImageJ. The wound healing rate = (0 hour width of scratch –24 hour width of scratch)/0 hour width of scratch ×100%.
Invasion assay
In brief, the Boyden chamber inserts were precoated with 80 µL of Matrigel (1:8 dilution) and incubated at 37°C for 30 minutes allowing a gel bed to form. The inserts were then placed onto the wells of a 24-well plate and cells (1×105) were seeded into the upper chamber. A total of 600 µL of medium supplemented with 10% FBS had been added into lower chamber. After 24 hours, the invaded cells were stained with crystal violet. The cells migrated to the bottom of the inserts are counted.20, 21
Colony formation assay
A total of 1×103 cells were mixed with DMEM containing soft agar (Difco, Mumbai, India) to a final concentration of 0.35%. These cells were then plated in a 25 mm3 dish coated with 0.6% soft agar in DMEM. Cells were cultured for 3 weeks. The colonies were stained with crystal violet (0.1%), and the number of cell colonies was counted.22
Apoptosis analysis
A total of 1×106 cells were collected and were re-suspended in 1 mL of binding buffer (BioLegend, San Diego, CA, USA) and incubated with Annexin V (10 µL) and PI solution (5 µL) for total 15 minutes at room temperature. Then, the cells were suspended in 500 µL of binding buffer, and apoptosis was analyzed using the FACScan flow cytometer (BD Biosciences, San Jose, CA, USA).
In situ hybridization (ISH)
The expression of WT1-AS in cervical cancer tissues was determined using the biotin-labeled WT1-AS probe. Paraffinized cancer sections were deparaffinized with xylene and 100% ethanol. Then, the sections were incubated with biotin-labeled probe at 40°C for 18 hours. DAB substrate was selected for the colorimetric measure of WT1-AS. Finally, sections were stained with hematoxylin followed by dehydration in graded alcohols and xylene.
Immunofluorescence
Cells (1×103) on glass coverslips were fixed by precold acetone and then rinsed with PBS. Cells were incubated with 1% BSA/PBS for avoiding nonspecific binding. Then, cells were incubated with anti-p53 antibody (1:500; Epitomics, Burlingame, CA, USA) at 4°C for 24 hours. Next, cells were incubated with fluorescein isothiocyanate (FITC)-conjugated goat antirabbit secondary antibody (Boster Biotechnology, Wuhan, Hubei, China). Nuclei were stained with DAPI (Boster Biotechnology).
Quantitative real-time PCR (qRT-PCR) assay
The total RNAs were extracted using a TRIzol kit (Thermo Fisher Scientific). The level of WT1-AS was detected using the SYBR Premix EX Taq™ II kit (TaKaRa, TakaraBio, Tokyo, Japan) on ABI Prism®7500 (ABI, Foster, CA, USA). miRNA was collected using the PureLink™ miRNA isolation kit (Thermo Fisher Scientific), and its level was determined using the TaqMan microRNA assay kit. The relative level of miRNA and lncRNA was calculated using the 2–ΔΔCT. GAPDH and U6 were the internal controls. The sequences of the primers were as follows: WT1-AS-F: GCCTCTCTGTCCTCTTCTTTGT and WT1-AS-R: GCTGTGAGTCCTGGTGCTTAG; GAPDH-F: GCTCTCTGCTCCTCCTGTTC and GAPDH-R: ACGACCAAATCCGTTGACTC; miR-330-5p-F: TCTCTGGGCCTGTGTCTTAGGC and miR-330-5p-R: GCTATCTCAGGGCTTGTTGCTTCAGTCCTCCTGGG; and U6-F: CTCGCTTCGGCAGCACA and U6-R: AACGCTTCACGAATTTGCGT.
Xenograft model of tumor growth in vivo
All animal experiments were approved by the University Committee on the Use and Care of Animals at First People’s Hospital of Jiaozuo City in accordance with Institutional Guidelines and the Guide for the Care and Use of Laboratory Animals (NIH publication no 85-23, revised 1996). A total of 100 µL of WT1-AS or its positive control (empty vector) cell suspension (3×106) was subcutaneously inoculated into the flank of nude mice. The tumor volume was measured each week using a caliper and calculated as the following formula: length × width2/2.
Experimental metastasis assay
For experimental metastasis analysis, the nude mice were injected at the lateral tail vein with (5×105) SiHa cells carrying shRNA targeting WT1-AS (sh-WT1-AS) or LV-WT1-AS. Mice were sacrificed 6 weeks after inoculation, and all organs were examined for the presence of macroscopic metastases. Lung metastatic nodules were determined under a dissecting microscope.
Statistical analysis
Results are presented as mean ± SD for the three repeated tests, the difference of two samples was compared used one-way ANOVA followed by post hoc Dunnett’s test or two-tailed Student’s t-test. P<0.05 was considered as significant.
Results
WT1-AS is downregulated in cervical carcinoma
In order to explore the expression of WT1-AS in the cervical carcinoma and control normal tissues, the levels of WT1-AS were analyzed in 63 pairs of cervical cancer tissues and matched paratumor tissues. WT1-AS was markedly downregulated in cervical carcinoma tissues than that in the control tissues. Additionally, the relatively lower level of WT1-AS was related to the advanced stages of cervical cancer and lymphatic metastasis. Next, we determined the prognostic significance of WT1-AS in patients with cervical cancer. We divided patients into WT1-AS low and WT1-AS high groups using the median value of 0.37 as a cutoff value. Kaplan–Meier analysis of 63 cases of patients with cervical cancer suggested that patients who had a lower level of WT1-AS exhibited a poor overall survival (Table S1). Then, we performed the ISH assay to further explore the level of WT1-AS in cervical carcinoma tissues. The level of WT1-AS was lower in tumor tissue than in paired peritumor tissue. To investigate the relationship of WT1-AS with the progression of cervical cancer, we analyzed the levels of WT1-AS in early cervical cancer (I) and advanced cervical cancer (II and III) tissues using the ISH assay. We found that advanced stages of cervical cancer (II and III) had relatively low levels of WT1-AS. Finally, WT1-AS levels were also detected in a panel of cervical cancer cell lines by qRT-PCR. Similar results were obtained in four cervical cancer cell lines in which the level of WT1-AS was lower in cervical cancer cell when compared with ECT1/E6E7. To identify the functions of WT1-AS in the growth of cervical cancer, we increased the expression of WT1-AS in cervical cancer cell using LV-WT1-AS. WT1-AS was significantly overregulated in LV-WT1-AS transfected SiHa and Ca-Ski cell. The MTT assay results showed that the overregulated of WT1-AS remarkably suppressed the growth of cervical cancer cell in vitro. Consistently, colony formation growth analysis revealed that upregulation of WT1-AS decreased the colony growth of cervical cancer cell. In order to confirm the role of WT1-AS on the proliferation of cervical cancer cell, SiHa and Ca-Ski cells were transfected with sh-Con or sh-WT1-AS. Then, the proliferation of SiHa and Ca-Ski cells was measured by the MTT assay. As shown in Figure S1, downregulation of WT1-AS remarkably increased the growth of cervical cancer cell in vitro. Altogether, these results indicated that WT1-AS was downexpressed and associated with cervical cancer progression.
Overregulation of WT1-AS inhibits cervical carcinoma cell migration and invasion
In order to explore the impact of WT1-AS on the migration ability of SiHa and Ca-Ski cells, we performed the wound-healing assay, in which we found that the distance between wound edge of LV-WT1-AS transfected SiHa and Ca-Ski cells was markedly longer than those of control cell (Figure 1A). However, we found shorter distance between wound edge of sh-WT1-AS transfected SiHa and Ca-Ski cells compared with control cell (Figure 1C). To investigate the effect of WT1-AS on the invasion of cervical carcinoma cell, we performed a Transwell invasion assay, in which we found that overexpression of WT1-AS remarkably suppressed SiHa and Ca-Ski cell invasion (Figure 1B). As expected, we observed that sh-WT1-AS significantly accelerated the invasion of cervical carcinoma cell (Figure 1D). Altogether, these findings suggested that WT1-AS inhibited cervical carcinoma cell migration and invasion in vitro.
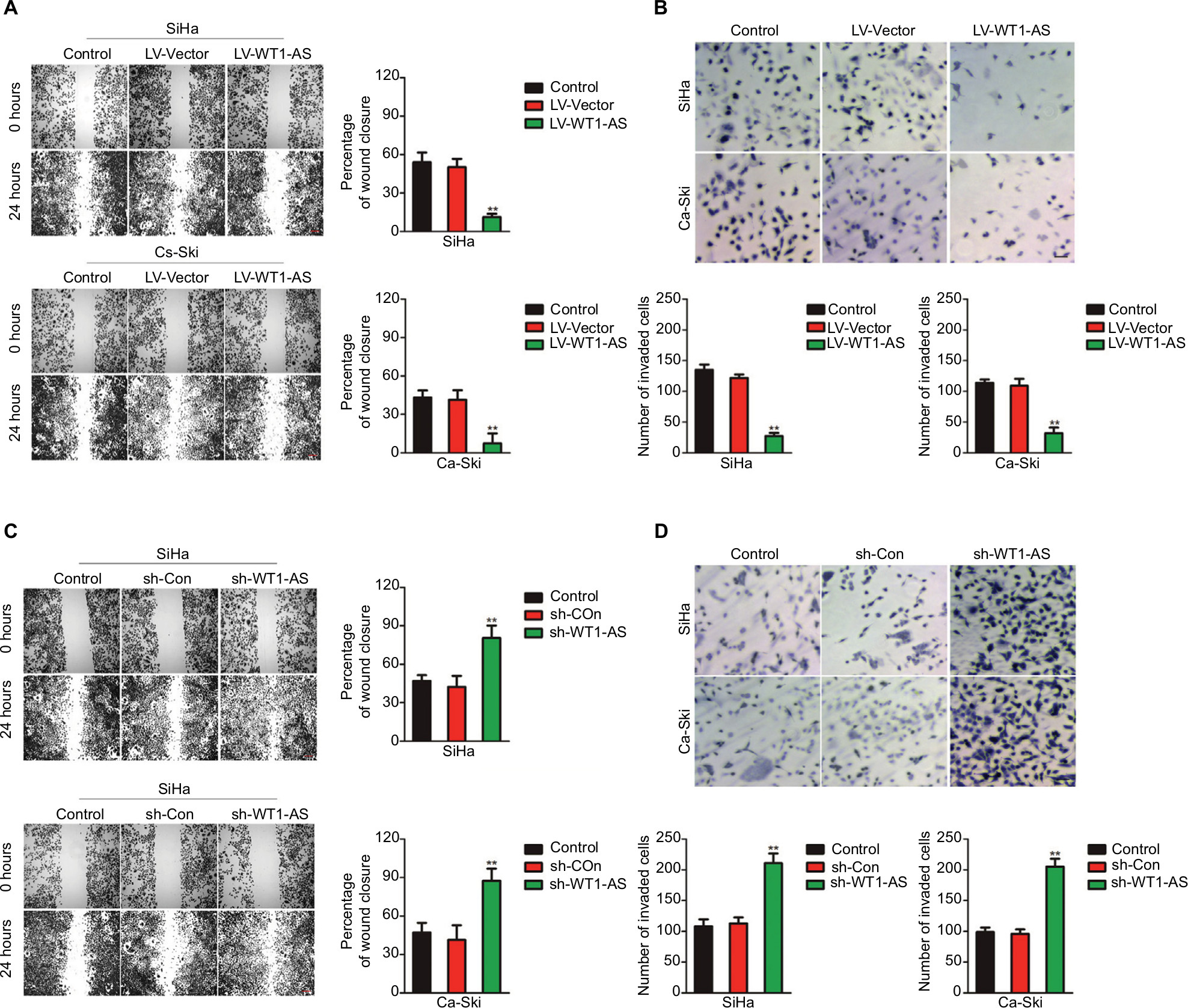 | Figure 1 Effects of WT1-AS on the migration and invasion in cervical cancer cell. Notes: (A) Wound healing assay with SiHa and Ca-Ski cells. Microscopic observations were recorded 0 and 24 hours after scratching the cell surface. A representative image was shown (left panel). Histograms showed the percentage of wound closure (right panel). (B) Transwell assay. Photographs show cervical cancer cell that invaded through the Transwell membrane (upper panel). Histograms showed the numbers of invaded cell (lower panel). (C) Wound healing assay with SiHa and Ca-Ski cells. Microscopic observations were recorded 0 and 24 hours after scratching the cell surface. A representative image was shown (left panel). Histograms showed the percentage of wound closure (right panel). (D) Transwell assay. Photographs show cell that invaded through the membrane (upper panel). Histograms showed the numbers of invaded cell (lower panel). **P<0.01, compared to control. Abbreviations: LV-WT1-AS, lentiviral vector encoding WT1-AS cDNA; sh-WT1-AS, shRNA targeting WT1-AS; WT1-AS, WT1 antisense RNA. |
Overregulation of WT1-AS inhibits the growth of cervical carcinoma cells in vivo
In vitro, WT1-AS inhibited cervical carcinoma cell growth, migration, and invasion. To investigate whether WT1-AS regulated the tumor growth of cervical carcinoma cell in vivo, SiHa cells that were transfected with LV-WT1-AS or LV-Vector were subcutaneously implanted into the nude mice. As shown in Figure 2A–2C, overexpression of WT1-AS remarkably inhibited the tumor growth of SiHa cell in vivo. To further analysis the impacts of WT1-AS on the tumor growth of SiHa cell in vivo, parental or WT1-AS downexpression SiHa cells were implanted into the nude mice to establish xenograft model. Downregulation of WT1-AS markedly accelerated the tumor growth of SiHa cell in vivo (Figure 2D–2F). To further investigate the role of WT1-AS in the metastasis of SiHa cell in vivo, an experimental metastasis assay was performed. SiHa cells that were transfected with sh-WT1-AS or LV-WT1-AS were injected into the lateral tail vein of nude mice. Six weeks postinoculation, animals were sacrificed, and all the major organs were checked for the generation of tumor metastasis. The tumor metastasis was mainly observed in the lungs. We found that sh-WT1-AS resulted in the formation of numerous lung colonies, whereas upregulation of WT1-AS significantly suppressed pulmonary metastasis (Figure 2G). Finally, the Annexin-FITC/PI staining was conducted to determine the effect of WT1-AS on the apoptosis of SiHa cell. As shown in Figure 2H, the apoptosis of cell that was transfected LV-WT1-AS was markedly increased when compared with parental cell. All these results demonstrated that WT1-AS inhibited the growth, migration, and invasion of cervical carcinoma cell in vivo.
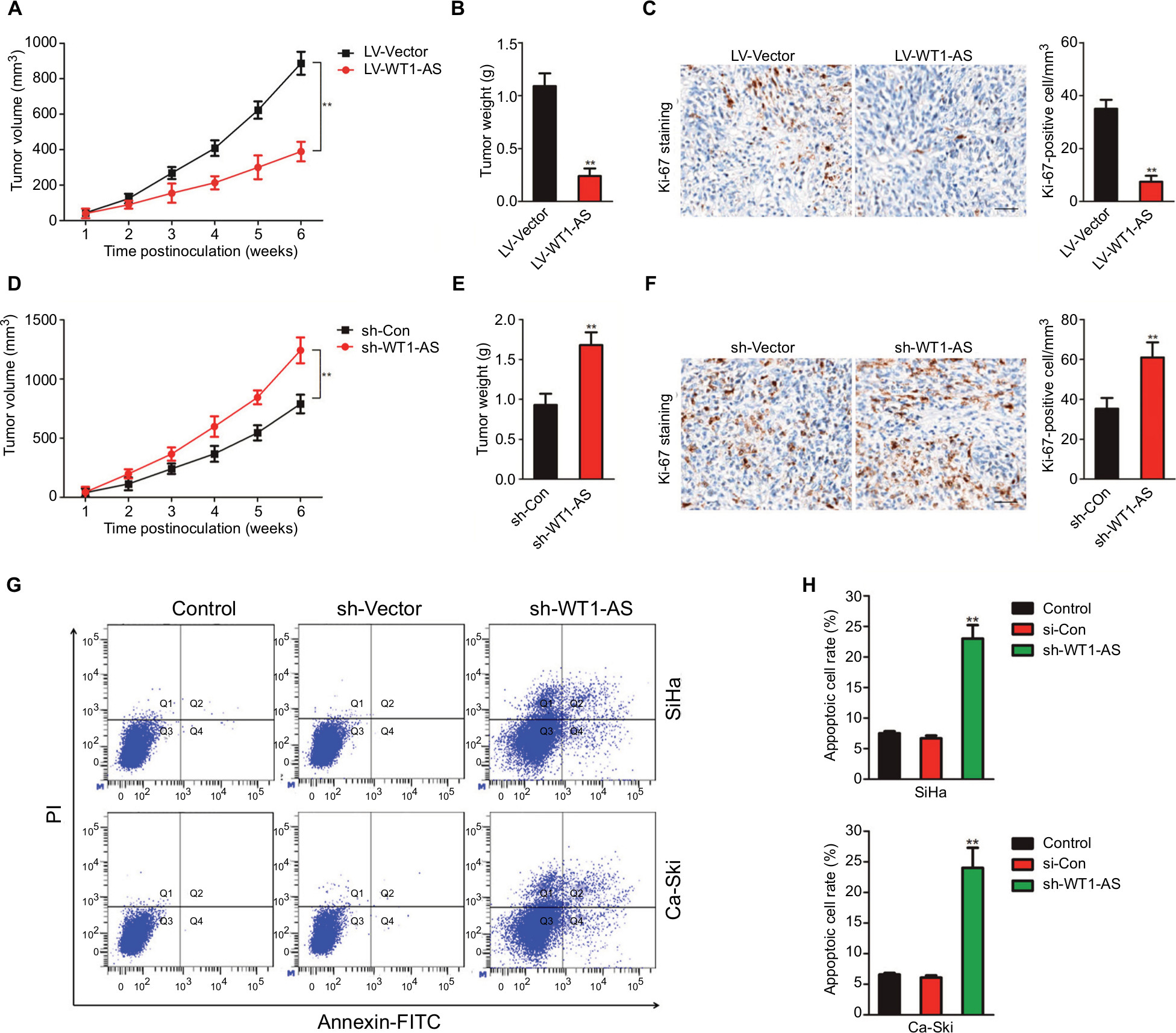 | Figure 2 Effect of WT1-AS on the tumor growth of SiHa cell in vivo. Notes: (A) SiHa cell was transfected with LV-WT1-AS and then was implanted into the nude mice. The tumor volume was measured each week. (B) Tumor weight. (C) Immunohistochemical staining of Ki-67 in tumor tissue. (D) SiHa cell was transfected with sh-Con or sh-WT1-AS and, then, was implanted into the nude mice. The tumor volume was measured each week, and the tumor volume was shown. (E) Tumor weight. (F) Immunohistochemical staining of Ki-67 in tumor tissue from sh-Con and sh-WT1-AS groups. (G) Metastatic lesions in the lungs of the models were shown at 6 weeks after tail vein. (H) Flow cytometry analysis of SiHa and Ca-Ski cells that were transfected with LV-WT1-AS or LV-Vector. **P<0.01, compared to control. Abbreviations: LV-WT1-AS, lentiviral vector encoding WT1-AS cDNA; PI, propidium iodide; sh-WT1-AS, shRNA targeting WT1-AS; WT1-AS, WT1 antisense RNA. |
WT1-AS is a target of miR-330-5p
In order to better uncover the underlying mechanisms by which WT1-AS acts in cervical cancer, we selected the microRNA.org (http://34.236.212.39/microrna/home.do), an online bioinformatics tool, to predict the potential downstream target of WT1-AS.21 As shown in Figure 3A, miR-330-5p was the target of WT1-AS with binding sites. We then explored the level of miR-330-5p in both Ca-Ski and SiHa cells that were transfected with LV-WT1-AS. As shown in Figure 3B, the level of miR-330-5p was inhibited when cell was transfected with LV-WT1-AS. Next, we transfected two cervical cancer cell lines with miR-330-5p mimics and found that miR-330-5p remarkably inhibited the expression of WT1-AS (Figure 3C and D). To future prove that WT1-AS directly binds to miR-330-5p, the luciferase reporter assay was carried out. The wt-WT1-AS or mut-WT1-AS was inserted into pmirGLO luciferase target expression vector. Then, plasmid was transfected into HEK-293T cell combine with miR-330-5p or miR-NC. The luciferase activity of HEK-293T cell that was transfected with wt-WT1-AS was decreased by miR-330-5p. Nevertheless, the inhibition effect of miR-330-5p was abolished in HEK-293T cell that was transfected with mut-WT1-AS (Figure 3E). Finally, the level of miR-330-5p was found inversely associated with the level of WT1-AS in cervical cancer tissue (Figure 3F). Therefore, these results demonstrated that lncRNA WT1-AS directly bound to miR-330-5p. We then examined whether miR-330-5p could affect cervical cancer cell growth and aggressiveness. As shown in Figure 3G–J, upregulation of miR-330-5p strongly promoted, while silencing miR-330-5p decreased the colony formation and invasion of SiHa cell, which indicated that miR-330-5p facilitated cervical carcinoma cell aggressiveness malignant phenotypes in vitro. E6/E7 protein is important for proliferation in cervical cancer cell. We next detected the role of miR-330-5p on the expression of HPV E6 and HPV E7. As shown in Figure S2, the protein expression of HPV E6 and HPV E7 in SiHa cell were slightly increased by miR-330-5p transfection.
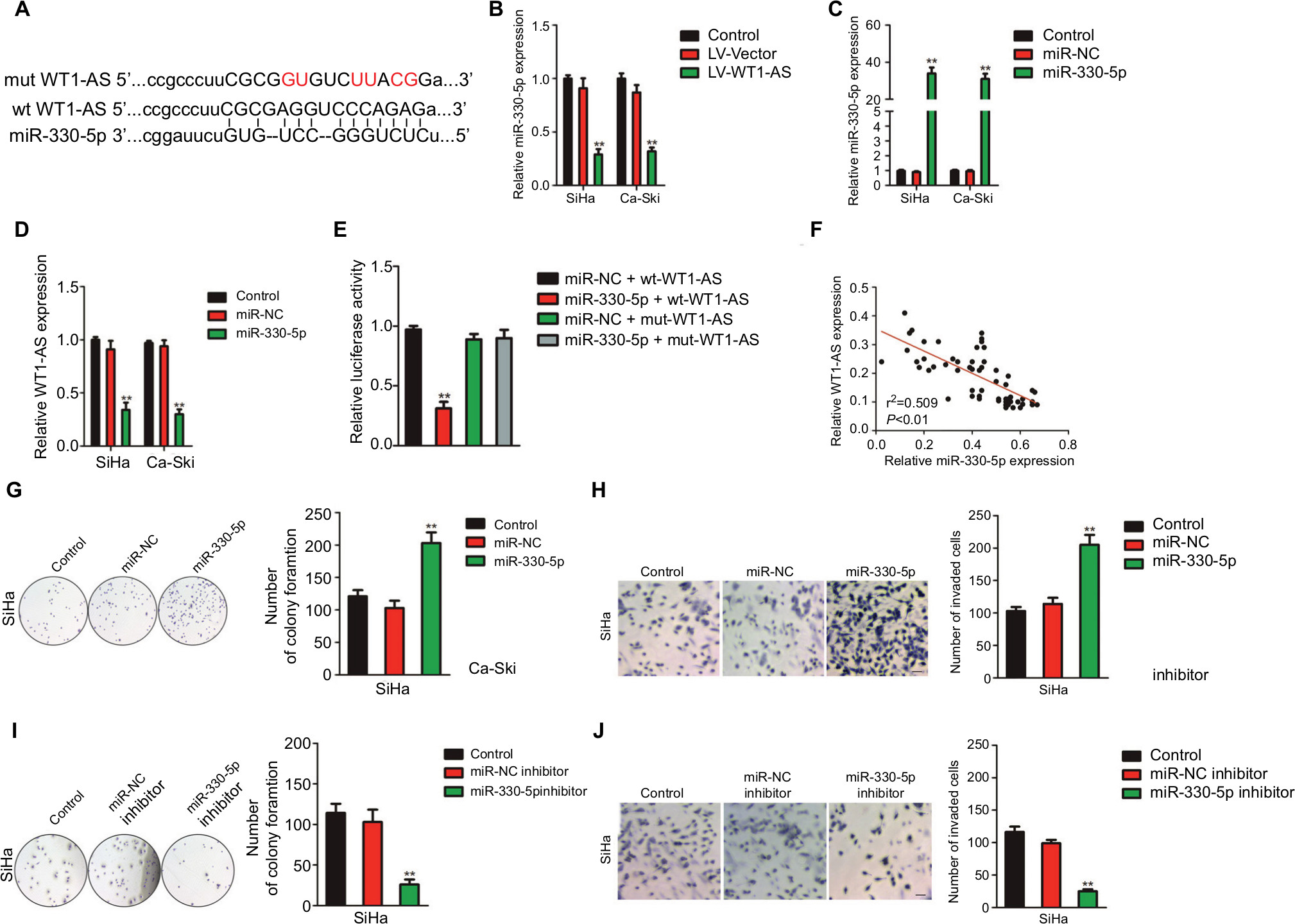 | Figure 3 WT1-AS is the downstream target of miR-330-5p. Notes: (A) The potential binding site was identified between WT1-AS and miR-330-5p. (B) Overexpression of WT1-AS inhibited the level of miR-330-5p in SiHa and Ca-Ski cells. (C) Cell was transfected with miR-NC or miR-330-5p, and the level of miR-330-5p was detected by qRT-PCR. (D) miR-330-5p transfection remarkably suppressed the level of WT1-AS in SiHa and Ca-Ski cells. **P<0.01, compared to control. (E) Luciferase reporter analysis suggested that WT1-AS directly targeted miR-330-5p. **P<0.01, compared to miR-MC + wt-WT1-AS. (F) Association between WT1-AS and miR-330-5p in cervical cancer tissue was evaluated. (G) miR-330-5p overexpressing promoted SiHa cell growth ability. Cell growth was measured by colony formation assay. (H) miR-330-5p overexpressing promoted cell invasion ability. Cell invasion was measured by Transwell invasion assay. (I) Downregulation of miR-330-5p inhibited SiHa cell colony formation. (J) Downexpression of miR-330-5p suppressed SiHa cell invasion in vitro. **P<0.01, compared to control. Abbreviations: LV-WT1-AS, lentiviral vector encoding WT1-AS cDNA; qRT-PCR, quantitative real-time PCR; wt, wild-type; WT1-AS, WT1 antisense RNA. |
Upregulation of miR-330-5p neutralizes the inhibitory effect of WT1-AS on cervical cancer cell
In order to analysis the levels of miR-330-5p in cervical carcinoma and control normal tissues, the levels of miR-330-5p were detected in 63 pairs of cervical cancer tissues and matched paratumor tissues. As shown in Figure 4A, the levels of miR-330-5p in cervical carcinoma tissues were much higher than that in the control. Additionally, the relatively higher level of miR-330-5p was related to the advanced stages of cervical cancer and lymphatic metastasis (Figure 4B and C). Kaplan–Meier analysis of 63 cases of patients with cervical cancer suggested that higher level of miR-330-5p was associated with the poor overall survival (Figure 4D and Table S2). Similar results were obtained in four cervical cancer cell lines in which the level of miR-330-5p was increased in cervical cancer cell when compared with ECT1/E6E7 cell (Figure 4E). Although we have identified that WT1-AS was the target of miR-330-5p, the precise roles of miR-330-5p in WT1-AS-mediated cervical cancer cell growth and aggressiveness were unknown. To solve this problem, SiHa cell was cotransfected with miR-330-5p along with WT1-AS (data not shown) and, then, colony formation and invasion assays were conducted. As shown in Figure 4F and G, overregulation of miR-330-5p in SiHa cell, which stably pretransfected with LV-WT1-AS, remarkably counteracted the suppressive effect of WT1-AS on SiHa cell colony formation and invasion. Moreover, SiHa cell was co-transfected with miR-330-5p inhibitor along with sh-WT1-AS and both colony formation and Transwell assays were conducted. As expected, miR-330-5p inhibitor significantly decreased the growth and invasion of SiHa cell that was increased by sh-WT1-AS (Figure 4H and I). Altogether, our study demonstrated that WT1-AS regulated cervical carcinoma cell growth, migration, and invasion through miR-330-5p.
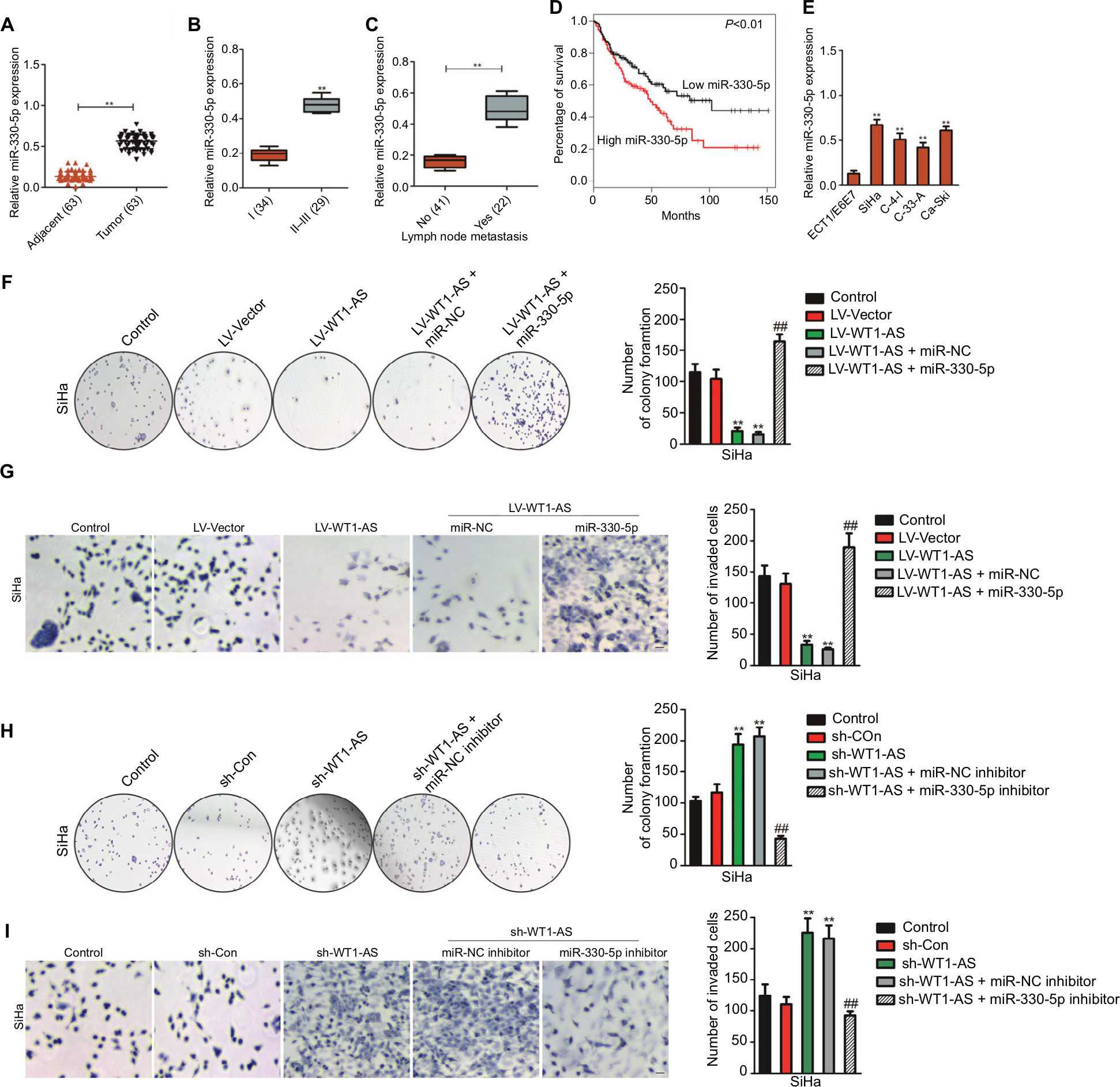 | Figure 4 Regulation of cervical cancer cell colony formation and invasion by WT1-AS requires miR-330-5p. Notes: (A) Total miRNAs was extracted from the paratumor and tumor tissues from patients with cervical cancer, and the level of miR-330-5p was determined by qRT-PCR assay. (B) The association between miR-330-5p expression and I or II–III tumor stage was shown. **P<0.01 compared to I. (C) Relative levels of miR-330-5p in metastatic and nonmetastatic cervical cancer tissues. **P<0.01 compared to nonmetastasis. (D) Overall survival analysis of cervical cancer patients with high or low level of miR-330-5p. (E) The levels of miR-330-5p in four cervical cancer cell lines and ECT1/E6E7 cell were detected by qRT-PCR analysis. **P<0.01 compared to ECT1/E6E7 cell. (F) Colony formation assay was performed in SiHa cell transfected with LV-WT1-AS, miR-330-5p, or LV-WT1-AS combination with miR-330-5p. (G) Transwell invasion assay was performed in SiHa cell transfected with LV-WT1-AS, miR-330-5p, or LV-WT1-AS combination with miR-330-5p. (H) Colony formation assay was conducted in SiHa cell transfected with sh-WT1-AS, miR-330-5p inhibitor, or sh-WT1-AS combination with miR-330-5p inhibitor. (I) Transwell invasion assay was conducted in SiHa cell transfected with sh-WT1-AS, miR-330-5p inhibitor, or sh-WT1-AS combination with miR-330-5p inhibitor. **P<0.01, compared to control. ##P<0.01, compared to LV-WT1-AS + miR-330-5p or sh-WT1-AS + miR-330-5p inhibitor. Abbreviations: LV-WT1-AS, lentiviral vector encoding WT1-AS cDNA; qRT-PCR, quantitative real-time PCR; sh-WT1-AS, shRNA targeting WT1-AS; WT1-AS, WT1 antisense RNA. |
WT1-AS serves as a ceRNA of p53 through binding to miR-330-5p
The binding site between miR-330-5p in p53 was predicted using the bioinformatics analysis (Figure 5A). The eleven common potential target genes that were obtained from three bioinformatics analysis tools (TargetScan, miRTarBase, and miRDB) are summarized in Figure S3A and B. miR-330-5p was found to bind p53 with a minimum free energy of –32.3 kcal/mol (Figure S3B). In order to identify the direct gene of miR-330-5p, the mRNA levels of these genes in miR-330-5p- or miR-NC-transfected SiHa cell were detected using the qRT-PCR assay. As shown in Figure S3C, the mRNA level of p53 was significantly inhibited by miR-330-5p in SiHa cell. Then, transfection of miR-330-5p into HEK-293T cell significantly inhibited the luciferase activity of p53-wt, whereas miR-330-5p transfection did not suppress the luciferase activity of p53 when the binding sites were mutated (Figure 5B). To confirm that p53 was the potential target of miR-330p, the expression of p53 in SiHa and Ca-Ski cells that were transfected with miR-NC or miR-330-5p was measured by qRT-PCR and immunofluorescence. As shown in Figures 5C and D, miR-330-5p transfection markedly decreased the level of p53 in cervical cancer cell. In addition, the expression of p53 was analyzed in 63 pairs of cervical cancer tissues and matched paratumor tissues. As shown in Figure 5E, the levels of p53 in cervical carcinoma tissues were much lower than those in the control tissues. Additionally, the relatively lower level of p53 was related to advanced stages of cervical cancer and lymphatic metastasis (Figure 5F and G). Kaplan–Meier analysis of 63 cases of patients with cervical cancer suggested that patients had a lower level of p53 exhibited the poor overall survival (Figure 5H and Table S3). To define the expression of p53 in cervical carcinoma, we checked the expression of p53 in peritumor and cervical carcinoma tissues by immunohistochemical staining with p53 antibody. We found that p53 exhibited lower level in cervical cancer tissue (Figure 5I). Importantly, the levels of miR-330-5p were found inversely associated with the expression of p53 in cervical carcinoma tissues (Figure 5J). Finally, similar results were obtained in four cervical cancer cell lines in which p53 was downexpressed in cervical cancer cell compared to ECT1/E6E7 cell (Figure 5K).
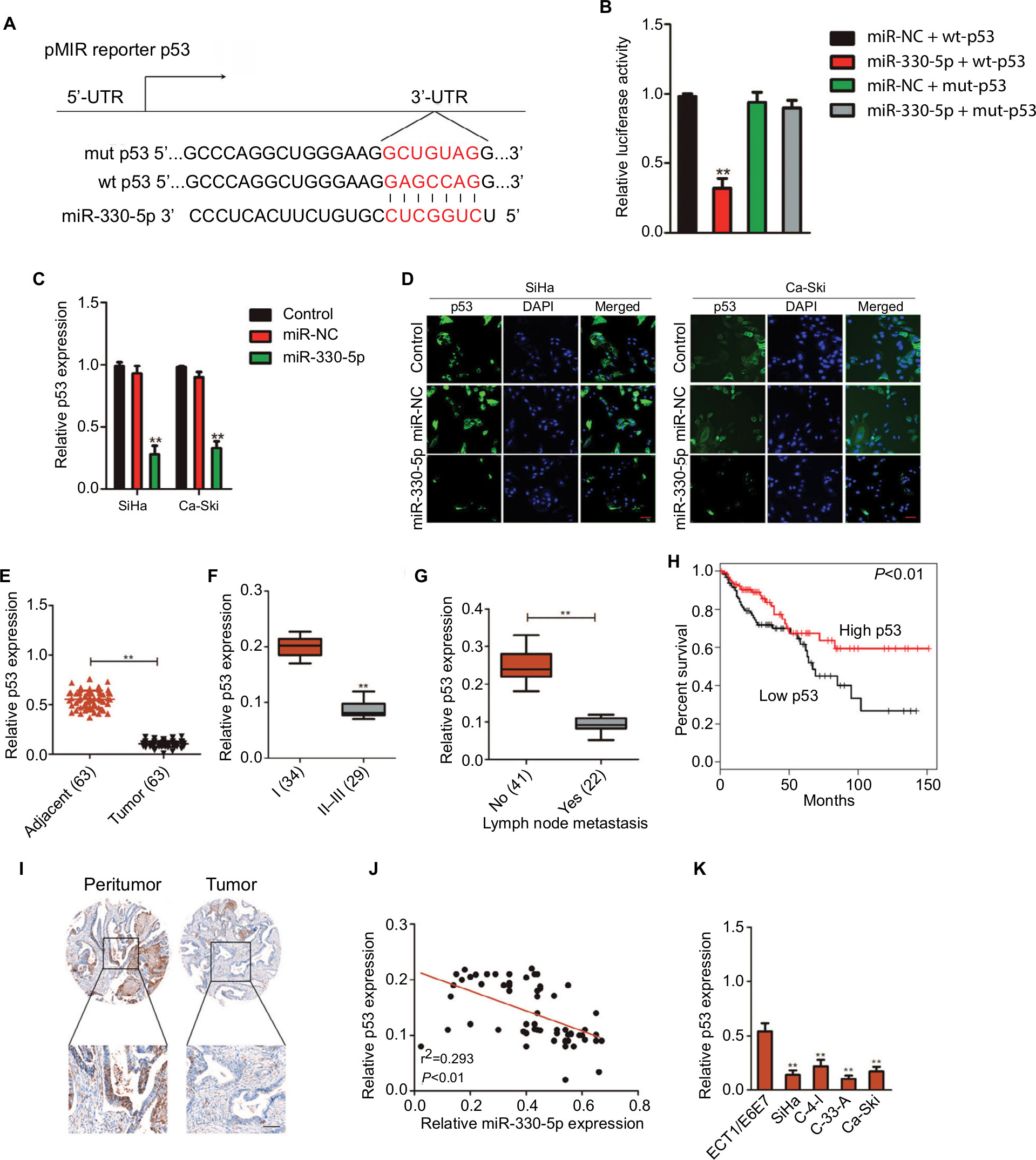 | Figure 5 p53 is the directly target of miR-330-5p and associated with the progression of cervical cancer. Notes: (A) Bioinformatics analysis revealed the putative binding sequences of miR-330-5p on p53 and the mutant sequences. (B) Luciferase activity was determined in HEK-293T cell co-transfected with miR-330-5p mimic and pMIR luciferase reporters containing p53-wt or p53-mut sequences. (C) miR-330-5p reduced the mRNA expression of p53 in SiHa and Ca-Ski cell. (D) miR-330-5p inhibited the expression of p53 in SiHa and Ca-Ski cell as shown in immunofluorescence analysis. (E) Total RNA was extracted from the paratumor and tumor tissues from patients with cervical cancer, and the mRNA level of p53 was determined by qRT-PCR assay. (F) Representative association between p53 expression and I or II–III tumor stage was shown. **P<0.01 compared to I. (G) Relative levels of p53 in metastatic and nonmetastatic cervical cancer tissues. **P<0.01 compared to nonmetastasis. (H) Overall survival analysis of cervical cancer patients with high or low level of p53. (I) The expression of p53 in peritumor and cervical cancer tissue was analyzed by immunohistochemical assay. (J) Association between p53 and miR-330-5p in cervical cancer tissue was evaluated. (K) The level of p53 in four cervical cancer cell lines and ECT1/E6E7 cell was detected by qRT-PCR analysis. **P<0.01 compared to ECT1/E6E7 cell. Abbreviation: qRT-PCR, quantitative real-time PCR. |
To further confirm whether WT1-AS regulated p53 expression by sponging miR-330-5p, SiHa, or Ca-Ski cell was transfected with LV-WT1-AS. We found that WT1-AS overexpressing increased the level of p53, whereas upregulation of miR-330-5p in the meantime reversed this trend (Figure 6A). In order to prove that WT1-AS-mediated regulation on the expression of p53 is through miR-330-5p, SiHa or Ca-Ski cell was transfected with sh-WT1-AS alone or cotransfected with sh-WT1-AS and miR-330-5p inhibitor. Then, the level of p53 was assayed using the qRT-PCR assay. As shown in Figure S4, sh-WT1-AS decreased the level of p53, whereas downregulation of miR-330-5p rescued the inhibitory effect of WT1-AS on the level of p53. Previous assays demonstrated that miR-330-5p attenuated the suppressive impacts of LV-WT1-AS on cervical cancer cell (Figure 4F–I). Then, we analyzed whether the impacts of WT1-AS on cervical cancer cell were regulated by miR-330-5p/p53. Two cervical cancer cell lines were cotransfected with sh-WT1-AS and LV-Vector or LV-p53 (Figure 6B). We revealed that the aggressiveness abilities of SiHa cell that were increased by the downregulation of WT1-AS were inhibited by the overexpression of p53. Upregulation of p53 markedly decreased the migration and invasion abilities of cervical cancer cell (Figure 6C–E). In general, these results indicated that WT1-AS served as a ceRNA to regulate the expression of p53 via sponging miR-330-5p (Figure 6F).
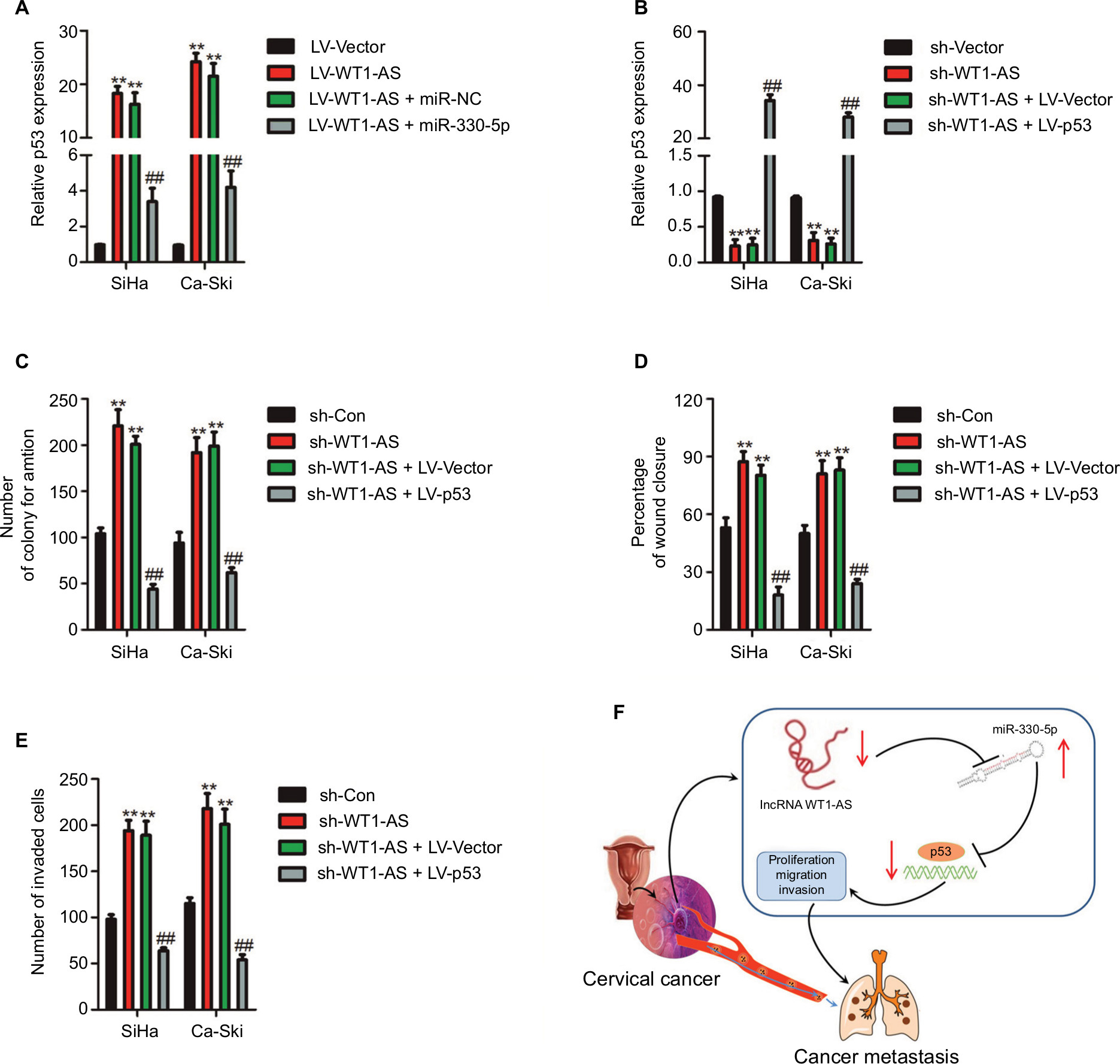 | Figure 6 WT1-AS inhibits cervical cancer cell colony formation, migration, and invasion via regulation of miR-330-5p/p53 axis. Notes: (A) The level of p53 was assayed in cell co-transfected with miR-330-5p alone or LV-WT1-AS combination with miR-330-5p. (B) Cell was co-transfected with sh-WT1-AS alone or sh-WT1-AS combination with LV-p53, and the level of p53 was determined by qRT-PCR analysis. (C) Cell was co-transfected with sh-WT1-AS alone or sh-WT1-AS combination with LV-p53, and colony formation assay was conducted. (D) Cell was co-transfected with sh-WT1-AS alone or sh-WT1-AS combination with LV-p53, and wound-healing assay was performed. (E) The invasion of transfected cell was determined by Transwell assay. (F) Schematic illustration of the positive feedback loop. WT1-AS upregulated p53 by binding to and suppressing miR-330-5p. **P<0.01 compared to sh-Con, ##P<0.01 compared to sh-WT1-AS + LV-Vector. Abbreviations: LV-WT1-AS, lentiviral vector encoding WT1-AS cDNA; qRT-PCR, quantitative real-time PCR; sh-WT1-AS, shRNA targeting WT1-AS; WT1-AS, WT1 antisense RNA. |
Discussion
Cervical cancer is one of the most aggressive tumors, and the occurrence of metastasis led to a sharp decline in survival rate of patients. Thus, identification of metastatic-related biomarkers and therapeutic targets will help improve the prognosis of patients with cervical cancer.23 Recently, lncRNAs have been proved to be involved in the invasion and metastasis of cancer and have been shown to be potentially noninvasive biomarkers in various cancers.14 Here, we reported that WT1-AS was a novel metastasis-associated lncRNA that played a significant role in the regulation of cervical carcinoma growth and metastasis. We found that WT1-AS was significantly downregulated in cervical carcinoma tissue and cervical cancer cell. In addition, overexpression of WT1-AS inhibited the aggressiveness and migration of cervical cancer cell in vitro. Consistently, overexpression of WT1-AS suppressed the growth of cervical cancer cell in vivo. In conclusion, our results suggested that metastatic-associated WT1-AS was not only a potential biomarker but also a valuable therapeutic target for cervical cancer.
Recently, substantive gene expression investigations of cancer have identify that a great quantity of lncRNAs are dysregulated in diverse cancer types.24 For example, lncRNA H19 inhibits cells’ growth, mobility, and invasion through downregulation of IRS-1 in thyroid cancer cell.25 lncRNA ATB is upregulated in osteosarcoma and positively associated with Enneking stage, metastasis, and recurrence.25 Meanwhile, overexpression of lncRNA ATB promotes osteosarcoma cell growth and aggressiveness by inhibiting miR-200s.26 In addition, lncRNA GPX4 participates into metastasis and regulates EMT process in colorectal carcinoma through competing for miR-200b-3p to future modulate the expression of ZEB1.27 Previous studies have convinced that the level of FENDRR is lower in breast cancer samples than that in the matched noncancerous specimens, and downregulation of FENDRR is correlated with the poor overall survival rate of patients with breast cancer.9 In vitro, upregulation of FENDRR inhibits breast carcinoma cell growth and increases apoptosis, while FENDRR knock-down increases breast carcinoma cell growth and restrains apoptosis.9 Additionally, WT1-AS is remarkably downexpressed in gastric carcinoma, and the expression of WT1-AS is associated with the tumor size and the clinicopathological stage of gastric carcinoma.19 Similarly, in this work, WT1-AS was downexpressed in cervical carcinoma tissue and cell when compared with the corresponding normal specimen and normal epithelial cell line.
To explore the precise roles of WT1-AS in cervical cancer cell, Ca-Ski and SiHa cells were selected and transfected with sh-WT1-AS and LV-WT1-AS, respectively. Overregulated WT1-AS inhibited the growth of cervical cancer cell as demonstrated in MTT and colony formation analysis, whereas downexpression of WT1-AS facilitated cell growth and colony formation. After demonstrating that WT1-AS inhibited the growth of cervical cancer cell in vitro, we next investigated the role of WT1-AS on cervical cancer cell in vivo. As expected, overregulation of WT1-AS remarkably decrease cervical cancer cell growth in vivo. Cancer cell metastasis is the major cause of death in patients with cancer. The initial steps of metastatic cascade include local invasion and migration, EMT, and intravasation.28 lncRNAs are emerging as key regulators governing biological processes of metastasis, including migration and invasion. In this work, wound healing and Transwell experiments were executed to verify whether WT1-AS regulated the migration and invasion ability of cervical cancer cell. All these findings suggested that both migration and invasion abilities of cervical carcinoma cell were inhibited by overregulation of WT1-AS.
Emerging evidence prove that lncRNAs are the endogenous miRNA sponges and competitively bind to miRNAs to future mediate genes’ expression.5 In order to explore whether WT1-AS acts as a miRNA sponge, the bioinformatics method was utilized and the result suggested that there were binding sites between miR-330-5p and WT1-AS and the following investigations confirmed that miR-330-5p was negatively regulated by WT1-AS. The luciferase activity assays verified that the luciferase activity was remarkably reduced in miR-330-5p and lncRNA WT1-AS wt cotransfection cell. Subsequently, miR-330-5p was demonstrated to reverse the roles of WT1-AS on the growth and malignant phenotypes of cervical cancer cell. miR-330-5p inhibitor transfected significantly decreased the growth and aggressiveness of cervical carcinoma cell that were increased by sh-WT1-AS, whereas overexpression of miR-330-5p reversed the inhibitory impacts of WT1-AS on the colony formation and invasion of cervical carcinoma cell.
One function of lncRNAs is to act as a ceRNA to sponge miRNAs. It is well known that miRNAs regulate the expression of target gene by binding to the 3′-UTR of gene.29 In our study, bioinformatics analysis was selected to predict the target gene and p53 was selected as the potential target gene. The luciferase reporter analysis, which is the direct method for target validation, also confirmed that p53 was the direct target of miR-330-5p. In cervical carcinoma, two viral genes, HPV E6 and HPV E7, are expressed and act cooperatively to promote tumorogenesis.30 The HPV E6 oncoprotein interacts with several cellular proteins, thereby activating a number of oncogenic pathways that lead to the blockage of apoptosis, alterations of the transcription machinery, interference with cell–cell interactions, and cell immortalization.31 One of the most investigated oncogenic activities of “high-risk” mucosal HPV E6 proteins is the ability to inactivate the tumor suppressor p53 by targeting it to degradation.32 We also explored the impact of miR-330-5p on the expression of HPV E6 and E7 and found that the expression of E6 and E7 were slightly increased by miR-330-5p in cervical cancer cell, suggesting that the suppressive role of miR-330-5p on the expression of p53 was associated with its regulation of HPV E6/E7. More importantly, the rescue experiments proved that miR-330-5p/p53 mediated the impacts of WT1-AS on cervical cancer cell, which revealed an lncRNA WT1-AS/miR-330-5p/p53 axis. In conclusion, WT1-AS inhibited cell growth and aggressiveness in cervical cancer cell, miR-330-5p was a target of WT1-AS, and alternation in miR-330-5p reversed the functions of WT1-AS on the progression of cervical cancer cell. Our study first revealed the possible relationship between WT1-AS and miR-330-5p in cervical cancer.
Disclosure
The authors report no conflicts of interest in this work.
References
Wu Y, Wang A, Zhu B, et al. KIF18B promotes tumor progression through activating the Wnt/β-catenin pathway in cervical cancer. Onco Targets Ther. 2018;11:1707–1720. | ||
Guo H, Yang S, Li S, Yan M, Li L, Zhang H. lncRNA SNHG20 promotes cell proliferation and invasion via miR-140-5p-ADAM10 axis in cervical cancer. Biomed Pharmacother. 2018;102:749–757. | ||
Shi C, Yang Y, Yu J, Meng F, Zhang T, Gao Y. The long noncoding RNA LINC00473, a target of microRNA 34a, promotes tumorigenesis by inhibiting ILF2 degradation in cervical cancer. Am J Cancer Res. 2017;7(11):2157–2168. | ||
Huang G, He X, Wei XL. lncRNA NEAT1 promotes cell proliferation and invasion by regulating miR-365/RGS20 in oral squamous cell carcinoma. Oncol Rep. 2018;39(4):1948–1956. | ||
Liang L, Xu J, Wang M, et al. lncRNA HCP5 promotes follicular thyroid carcinoma progression via miRNAs sponge. Cell Death Dis. 2018;9(3):372. | ||
Liao Z, Zhao J, Yang Y. Downregulation of lncRNA H19 inhibits the migration and invasion of melanoma cells by inactivating the NF-κB and PI3K/Akt signaling pathways. Mol Med Rep. 2018;17(5):7313–7318. | ||
Yang L, Wu D, Chen J, et al. A functional CNVR_3425.1 damping lincRNA FENDRR increases lifetime risk of lung cancer and COPD in Chinese. Carcinogenesis. 2018;39(3):347–359. | ||
Xu TP, Huang MD, Xia R, et al. Decreased expression of the long non-coding RNA FENDRR is associated with poor prognosis in gastric cancer and FENDRR regulates gastric cancer cell metastasis by affecting fibronectin1 expression. J Hematol Oncol. 2014;7:63. | ||
Li Y, Zhang W, Liu P, et al. Long non-coding RNA FENDRR inhibits cell proliferation and is associated with good prognosis in breast cancer. Onco Targets Ther. 2018;11:1403–1412. | ||
Zhang G, Han G, Zhang X, et al. Long non-coding RNA FENDRR reduces prostate cancer malignancy by competitively binding miR-18a-5p with RUNX1. Biomarkers. 2018;23(5):435–445. | ||
Oliveira AI, Xavier-Magalhães A, Moreira-Barbosa C, et al. Influence of HOTAIR rs920778 and rs12826786 genetic variants on prostate cancer risk and progression-free survival. Biomark Med. 2018;12(3):257–264. | ||
Zhou Y, Wang C, Liu X, Wu C, Yin H. Long non-coding RNA HOTAIR enhances radioresistance in MDA-MB231 breast cancer cells. Oncol Lett. 2017;13(3):1143–1148. | ||
Fagoonee S, Durazzo M. HOTAIR and gastric cancer: a lesson from two meta-analyses. Panminerva Med. 2017;59(3):201–202. | ||
Hong Q, Li O, Zheng W, et al. lncRNA HOTAIR regulates HIF-1α/AXL signaling through inhibition of miR-217 in renal cell carcinoma. Cell Death Dis. 2017;8(5):e2772. | ||
Lv L, Chen G, Zhou J, Li J, Gong J. WT1-AS promotes cell apoptosis in hepatocellular carcinoma through down-regulating of WT1. J Exp Clin Cancer Res. 2015;34:119. | ||
Kaneuchi M, Sasaki M, Tanaka Y, et al. WT1 and WT1-AS genes are inactivated by promoter methylation in ovarian clear cell adenocarcinoma. Cancer. 2005;104(9):1924–1930. | ||
Hancock AL, Brown KW, Moorwood K, et al. A CTCF-binding silencer regulates the imprinted genes AWT1 and WT1-AS and exhibits sequential epigenetic defects during Wilms’ tumourigenesis. Hum Mol Genet. 2007;16(3):343–354. | ||
Busch M, Schwindt H, Brandt A, et al. Classification of a frameshift/extended and a stop mutation in WT1 as gain-of-function mutations that activate cell cycle genes and promote Wilms tumour cell proliferation. Hum Mol Genet. 2014;23(15):3958–3974. | ||
du T, Zhang B, Zhang S, et al. Decreased expression of long non-coding RNA WT1-AS promotes cell proliferation and invasion in gastric cancer. Biochim Biophys Acta. 2016;1862(1):12–19. | ||
Zanier K, Ould M’Hamed Ould Sidi A, Boulade-Ladame C, et al. Solution structure analysis of the HPV16 E6 oncoprotein reveals a self-association mechanism required for E6-mediated degradation of p53. Structure. 2012;20(4):604–617. | ||
Zeng HF, Yan S, Wu SF. MicroRNA-153-3p suppress cell proliferation and invasion by targeting SNAI1 in melanoma. Biochem Biophys Res Commun. 2017;487(1):140–145. | ||
Zhou J, Xu D, Xie H, et al. miR-33a functions as a tumor suppressor in melanoma by targeting HIF-1α. Cancer Biol Ther. 2015;16(6):846–855. | ||
Luo C, Fan W, Jiang Y, Zhou S, Cheng W. Glucose-related protein 78 expression and its effects on cisplatin-resistance in cervical cancer. Med Sci Monit. 2018;24:2197–2209. | ||
Zhou R, Chen KK, Zhang J, et al. The decade of exosomal long RNA species: an emerging cancer antagonist. Mol Cancer. 2018;17(1):75. | ||
Wang P, Liu G, Xu W, Liu H, Bu Q, Sun D. Long noncoding RNA H19 inhibits cell viability, migration, and invasion via downregulation of IRS-1 in thyroid cancer cells. Technol Cancer Res Treat. 2017;16(6):1102–1112. | ||
Han F, Wang C, Wang Y, Zhang L. Long noncoding RNA ATB promotes osteosarcoma cell proliferation, migration and invasion by suppressing miR-200s. Am J Cancer Res. 2017;7(4):770–783. | ||
Chen DL, Chen LZ, Lu YX, et al. Long noncoding RNA XIST expedites metastasis and modulates epithelial-mesenchymal transition in colorectal cancer. Cell Death Dis. 2017;8(8):e3011. | ||
Shroff GS, Viswanathan C, Carter BW, Benveniste MF, Truong MT, Sabloff BS. Staging lung cancer: metastasis. Radiol Clin North Am. 2018;56(3):411–418. | ||
Xue P, Zheng M, Diao Z, et al. miR-155* mediates suppressive effect of PTEN 3’-untranslated region on AP-1/NF-κB pathway in HTR-8/SVneo cells. Placenta. 2013;34(8):650–656. | ||
Martinez-Zapien D, Ruiz FX, Poirson J, et al. Structure of the E6/E6AP/p53 complex required for HPV-mediated degradation of p53. Nature. 2016;529(7587):541–545. | ||
Stutz C, Reinz E, Honegger A, et al. Intracellular analysis of the interaction between the human papillomavirus type 16 E6 oncoprotein and inhibitory peptides. PLoS One. 2015;10(7):e0132339. | ||
Ansari T, Brimer N, vande Pol SB. Peptide interactions stabilize and restructure human papillomavirus type 16 E6 to interact with p53. J Virol. 2012;86(20):11386–11391. |
Supplementary materials
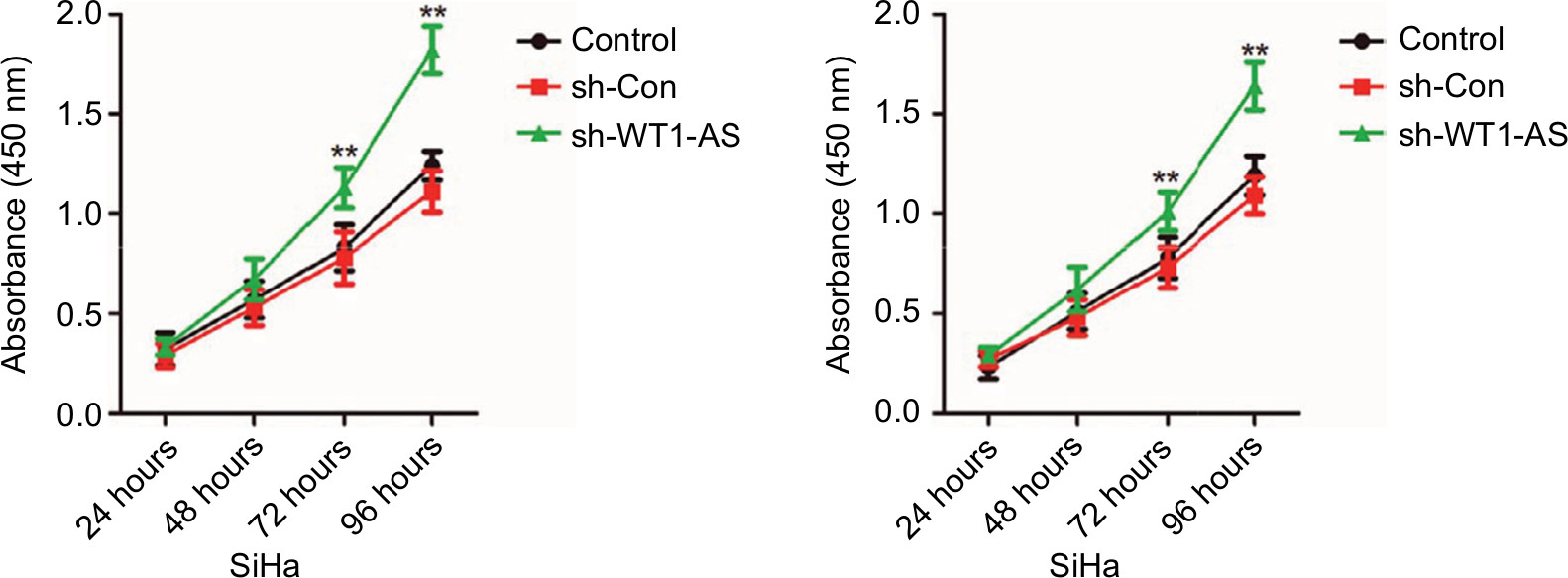 | Figure S1 Cell was transfected with sh-Con or sh-WT1-AS and was plated into 96-well plates and cultured for 1, 2, 3, or 4 days. Notes: MTT assay was performed, and the OD value was measured at 490 nm. **P<0.01 compared to control. |
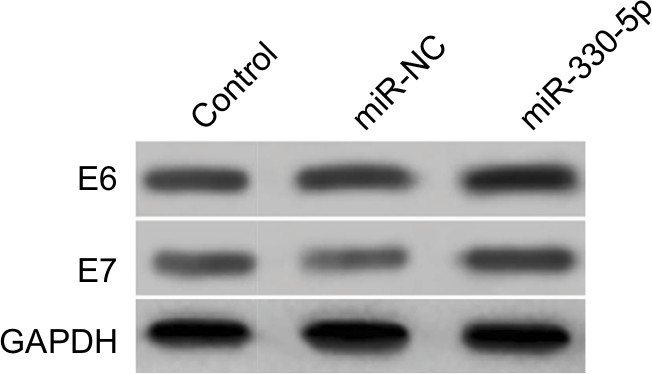 | Figure S2 SiHa cell was transfected with miR-NC or miR-330-5p, and the expression of HPV E6 and HPV E7 were detected using Western blotting assay. |
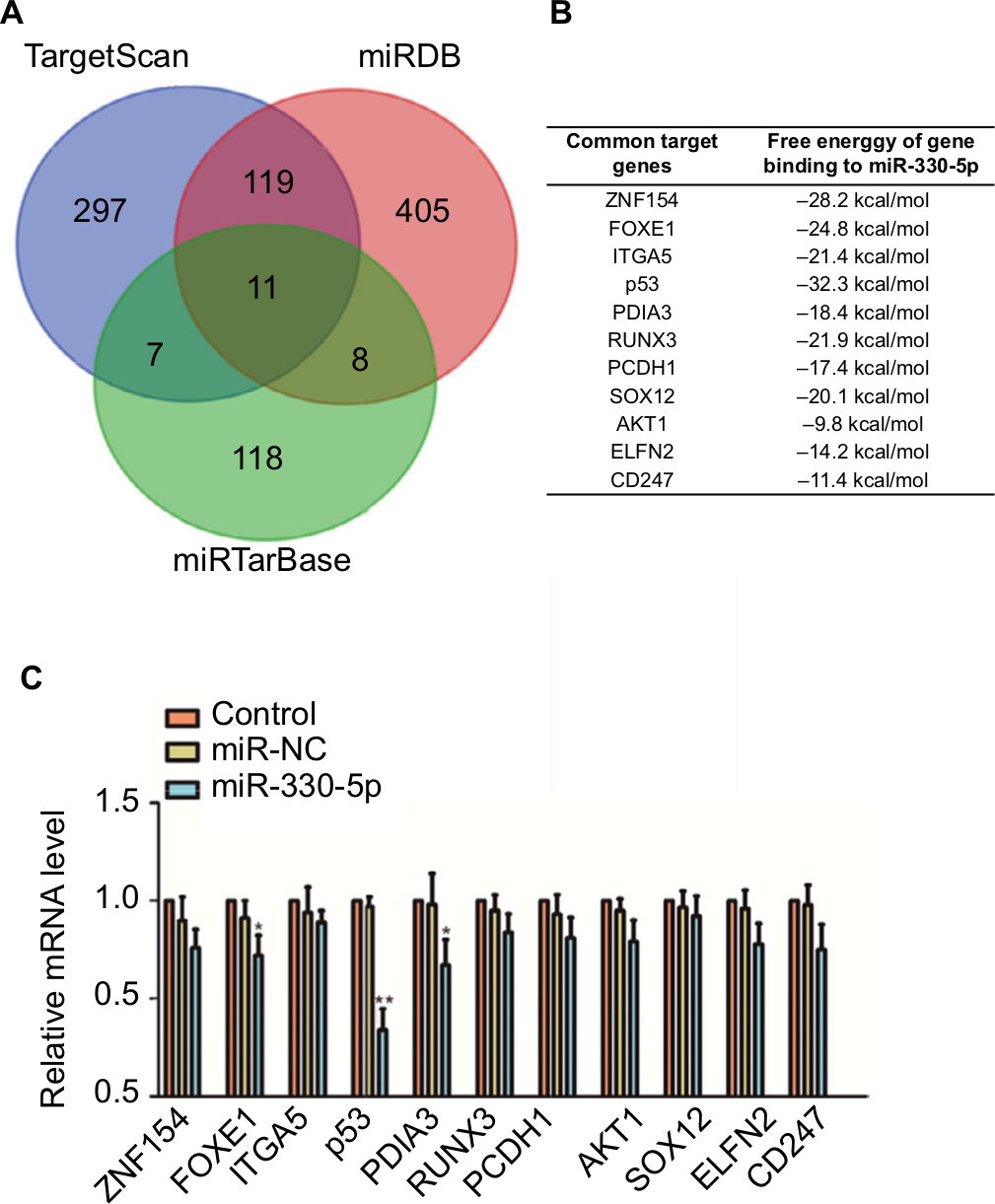 | Figure S3 The mRNA level of p53 is significantly inhibited in cell that is transfeced with miR-330-5p. Notes: (A and B) Venn graph represented the number of candidate common target genes determined by three bioinformatics analysis. (C) SiHa cell was transfected with miR-NC or miR-330-5p, and the levels of common target genes were detected using qRT-PCR assay. *P<0.05, **P<0.01 compared to control. Abbreviation: qRT-PCR, quantitative real-time PCR. |
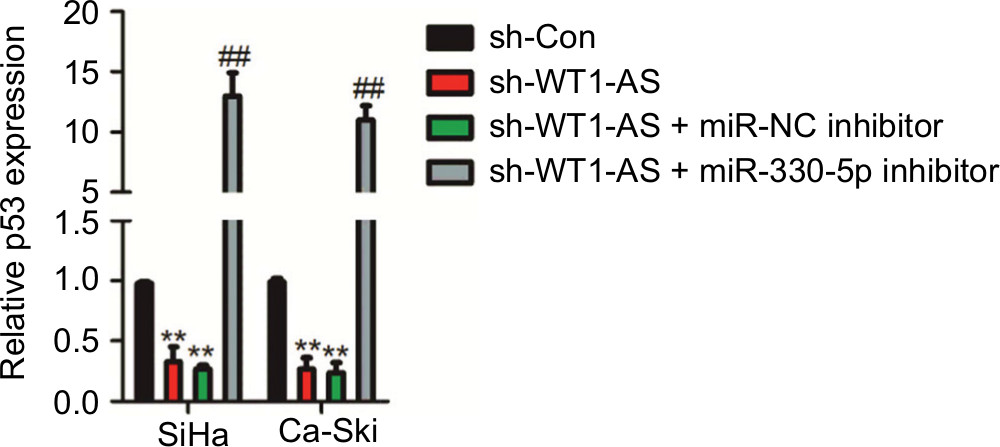 | Figure S4 SiHa or Ca-Ski cell was transfected with sh-WT1-AS alone or cotransfected with sh-WT1-AS and miR-330-5p inhibitor. Notes: The level of p53 was assayed using the qRT-PCR assay. **P<0.01 compared to sh-Con. ##P<0.01 compared to sh-WT1-AS + miR-NC inhibitor. Abbreviation: qRT-PCR, quantitative real-time PCR. |
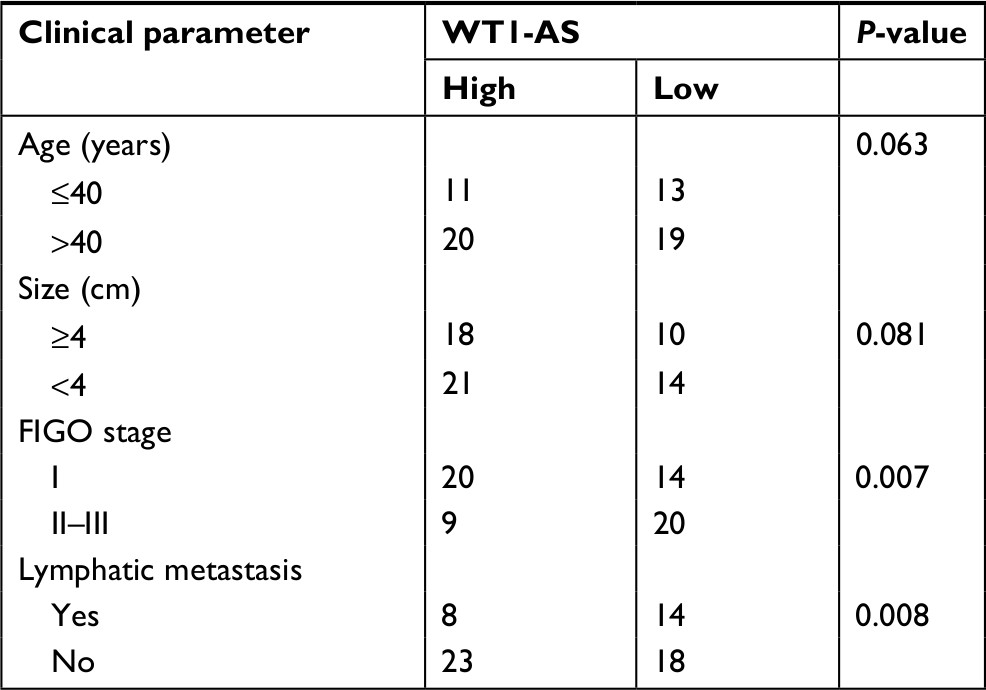 | Table S1 The association between WT1-AS expression and clinicopathological factors in cervical cancer patients Abbreviation: FIGO, International Federation of Gynecology and Obstetrics. |
 | Table S2 The association between miR-330-5p expression and clinicopathological factors in cervical cancer patients Abbreviation: FIGO, International Federation of Gynecology and Obstetrics. |
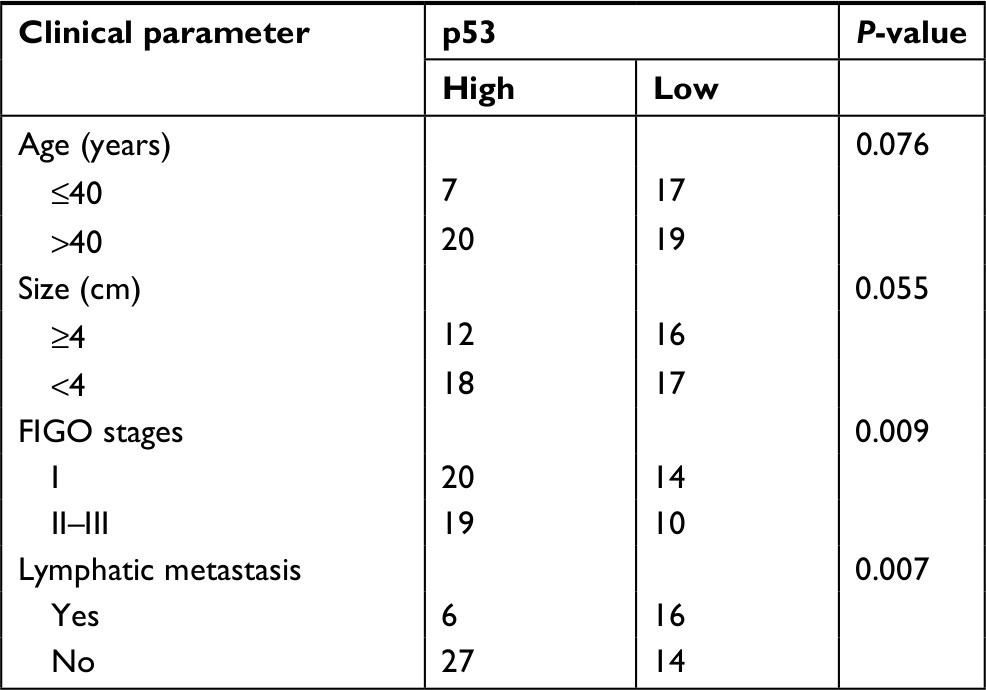 | Table S3 The association between p53 expression and clinicopathological factors in cervical cancer patients Abbreviation: FIGO, International Federation of Gynecology and Obstetrics. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.