")
Back to Journals » Cancer Management and Research » Volume 13
lncRNA MCF2L-AS1/miR-105/ IL-1β Axis Regulates Colorectal Cancer Cell Oxaliplatin Resistance
Authors Cai M, Hu W, Huang C, Zhou C, Li J, Chen Y, Yu Y
Received 1 April 2021
Accepted for publication 6 July 2021
Published 19 November 2021 Volume 2021:13 Pages 8685—8694
DOI https://doi.org/10.2147/CMAR.S313905
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Bilikere Dwarakanath
Mao Cai,* Wanle Hu,* Chongjie Huang, Chongjun Zhou, Jiante Li, Yanyu Chen, Yaojun Yu
Department of Anorectal Surgery, The Second Affiliated Hospital of Wenzhou Medical University, Wenzhou, 325027, Zhejiang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yaojun Yu
Department of Anorectal Surgery, The Second Affiliated Hospital of Wenzhou Medical University, 109 West Xueyuan Road, Wenzhou, 325027, Zhejiang, People’s Republic of China
Email [email protected]
Background: Interactions between non-coding RNAs and mRNAs have been shown to play key roles in colorectal cancer (CRC) resistance to chemotherapeutic drugs, but the regulatory network of these ncRNA/mRNA interactions in the context of CRC cell resistance to oxaliplatin has yet to be fully defined.
Methods: MCF2L-AS1, miR-105, and IL-1β expression levels were measured in cells and serum samples via qPCR, while ELISAs were additionally used to quantify IL-1β levels in these samples. Interactions between MCF2L-AS1, miR-105, and IL-1β were detected through pull-down, RNA immunoprecipitation, and luciferase reporter assays. Cellular viability and OXA IC50 values were established through MTT assays, while in vivo OXA resistance was assessed using a tumor xenograft model system.
Results: MCF2L-AS1 levels were significantly elevated in CRC patients that did not respond to chemotherapy and in CRC/OXA cells relative to responders and chemosensitive CRC cells. From a mechanistic perspective, miR-105 was identified as a MCF2L-AS1 target, with this miRNA, in turn, suppressing the expression of IL-1β. Knocking down MCF2L-AS1 or overexpressing miR-105 was sufficient to alleviate CRC/OXA cell chemoresistance, while overexpressing IL-1β reversed this effect.
Conclusion: The MCF2L-AS1/miR-105/IL-1β regulatory axis regulates the resistance of CRC cells to OXA treatment.
Keywords: CRC, oxaliplatin, OXA resistance, MCF2L-AS1, miR-105, IL-1β
Background
Colorectal cancer (CRC) is among the most prevalent and deadly cancers in the world, and affected patients often have poor outcomes.1,2 Oxaliplatin (OXA)-based chemotherapy following tumor resection is the primary approach to treating this cancer type.3 The administration of OXA in combination with 5-fluorouracil (5-FU) and leucovorin (FOLFOX) is associated with >50% response rates in CRC patients, who exhibit a median survival of roughly two years.4 However, many of these patients develop chemoresistance, which represents a major barrier to therapeutic efficacy.5 Clarifying the mechanistic basis for such chemoresistance thus has the potential to highlight new means of reliably treating patients with this deadly cancer type.
Long non-coding RNAs (lncRNAs) have been reported to regulate the progression of diverse forms of cancer, including non-small cell lung cancer (NSCLC), gastric cancer, breast cancer, and CRC.6–9 These lncRNAs can regulate chemoresistance and disease progression in the context of CRC.10–12 For example, the downregulation of LINC00958 can impair the proliferative, migratory, and invasive properties of CRC cells, instead inducing their apoptotic death through a mechanism associated with the ability of this lncRNA to target miR-3619-5p.13 By sequestering miR-145 and thus indirectly controlling ABCC1 expression, the lncRNA CACS15 can facilitate OXA chemoresistance in CRC cells.14 This interaction whereby lncRNAs serve as competing endogenous RNAs (ceRNAs) which sequester miRNAs in a sequence-specific fashion is thought to influence diverse cancer types,15–18 influencing oncogenesis, chemoresistance, tumor cell survival, and progression in the context of CRC. The BLACAT1/miR-519d-3p/CREB1 regulatory axis, for example, can control CRC cell migratory, invasive, proliferative, chemoresistant, and survival properties.19
In one recent report, the lncRNA MCF2L-AS1 was shown to be related to CRC progression,6 yet its influence on the chemoresistance properties of these same tumor cells has yet to be assessed. Herein, we found that CRC patients that were unresponsive to OXA-based chemotherapy exhibited MCF2L-AS1 upregulation, as did OXA-resistant CRC cells, suggesting this lncRNA to be closely related to chemoresistance. To explore the underlying regulatory mechanisms, we conducted bioinformatics analyses when identified miR-105 as a putative MCF2L-AS1 target miRNA which in turn suppressed the expression of IL-1β. Consistent with these regulatory relationships, CRC/OXA tissues and cells exhibited reduced miR-105 expression and increased expression of IL-1β, suggesting that MCF2L-AS1 may indirectly control IL-1β expression by serving as a ceRNA for miR-105, thus regulating CRC cell resistance to OXA.
Materials and Methods
Clinical Samples
In total, 120 samples of tumor tissues were collected from patients with CRC who were undergoing standard OXA-based chemotherapy. All patients provided written informed consent, and the Research Ethics Committee of The Second Affiliated Hospital of Wenzhou Medical University approved this study, which was consistent with the guidelines of the Declaration of Helsinki.
Cell Culture and Transfection
The HT-29 and LOVO CRC cell lines were obtained from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China), and both the parental and OXA-resistant versions of these cells (HT-29/OXA and LOVO/OXA) were cultured in RPMI-1640 (Thermo Fisher Scientific, DE, USA) containing 10% FBS (Sigma-Aldrich, MO, USA) at 37°C in a 5% CO2 incubator.
RIBOBIO (China) was the source of all RNAi vectors (sh-MCF2L-AS1 and miR-105 inhibitor), overexpression vectors (pcDNA3.1-IL-1β and miR-105 mimic), and corresponding controls (sh-NC, NC inhibitor, pcDNA3.1, and NC mimic). Cells were transfected with these constructs by plating them in 6-well plates (4×105/well) for 24 h, after which Lipofectamine 3000 (Invitrogen, USA) was used based on provided directions to transfect cells with 5 µg of an appropriate plasmid.
qPCR
For qPCR analyses, a TaqMan miRNA Reverse Transcription Kit (Applied Biosystems, CA, USA) or Reverse Transcription Reagents (Applied Biosystems) was used to prepare cDNA for miRNAs and for mRNAs/lncRNAs, respectively. All qPCR reactions were then performed with SYBR Green (Takara Bio Inc, Japan), using GAPDH and U6 as respective normalization controls for mRNAs/lncRNAs and miRNAs. Relative target expression was assessed via the 2−ΔΔCt approach. Primers used herein were as follows: MCF2L-AS1, forward, 5′- GATCAACGTTCAATCCACCG-3′ and reverse, 5′-A CGTCAAGATAGCGCAGCTTCC-3′; GAPDH, forward, 5′-TGTTCGTCATGGGTGTGAAC-3′ and reverse, 5′-ATGGCATGGACTGTGGTCAT-3′; IL-1β, forward, 5′-AAATACCTGTGGCCTTGGGC-3′ and reverse, 5′-TTTGGGATCTACACTCTCCAGCT-3′; miR-105, forward, 5′-TCGGCAGGTCAAATGCTCAGACTCC-3′ and reverse, 5′-CTCAACTGGTGTCGTGGA-3′; and U6, forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
MTT Assay
To assess cell viability, cells were added to 96-well plates (2×103/well) following transfection or other treatment. After 48 h, MTT solution was added to each well (20 μL; 5 mg/mL; Sigma-Aldrich). After 4 h, 200 µL of DMSO (Sigma-Aldrich) was added per well, and absorbance at 490 nm was quantified.
Biotin RNA Pulldown Assay
The interactions between MCF2L-AS1 and miR-105 in CRC cells were assessed via biotin RNA pulldown assay. Briefly, biotinylated sense or antisense were combined with lysates of HT-29/OXA and LOVO/OXA cells for 2 h, after which streptavidin-agarose beads (Invitrogen) was used for immunoprecipitation. MCF2L-AS1 and miR-105 expression was then assessed via qPCR.
RNA Immunoprecipitation (RIP)
A Magna RIP Kit (Millipore) was used based on provided directions. Briefly, anti-Ago2 (Cell Signaling Technology) was used to precipitate RNA, with qPCR being used for subsequent analyses and with data being shown as fold enrichment of MCF2L-AS1. Precipitated RNA Ct values were normalized to Ct values for the input RNA fractions.
Luciferase Reporter Assays
WT or mutant MCF2L-AS1 and IL-1β 3ʹ-UTR sequences containing putative miR-105 binding sites were amplified and cloned into the pGL3 vector to yield the WT pGL3-MCF2L-AS1 (MCF2L-AS1-WT) and pGL3-IL-1β (IL-1β-WT) or mutant pGL3-MCF2L-AS (MCF2L-AS-MUT) and pGL3-IL-1β (IL-1β-MUT) vectors. Next, cells were co-transfected with vectors and miR-105 mimics or corresponding controls. At 48 h post-transfection, a dual-luciferase reporter assay system (Promega, Shenzhen, Guangdong, China) was utilized based on provided directions to assess luciferase activity.
ELISAs
Levels of IL-1β were assessed in HT-29/OXA and LOVO/OXA cell supernatants using a commercial ELISA kit (R&D systems).
Animal Experiments
BALB/c nude mice (4- to 6-week-old) were obtained from the SLAC Laboratory Animal Co., Ltd (Shanghai, China) and housed under specific pathogen-free conditions at 28°C. The Institutional Animal Care and Use Committee of The Second Affiliated Hospital of Wenzhou Medical University approved this study, which was consistent with the NIH guidelines for the care and use of laboratory animals. Briefly, mice were subcutaneously implanted with 5×106 T-29/OXA and LOVO/OXA cells that had been transfected with si-MCF2L-AS1 or si-NC constructs in the right flank (n=5/group). At 6 days post-implantation, mice were intraperitoneally injected with OXA (2 mg/kg) every 7 days five times. Tumor volume was assessed every 7 days for 28 days as follows: volume = length × width2/2.
Statistical Analysis
Data are means ± SD, and were compared between groups using Student’s t-tests and one-way ANOVAs. Associations between MCF2L-AS1 and miR-105 expression were assessed through Pearson’s correlation analyses. Data were prepared and analyzed with GraphPad Prism 7.0 (GraphPad Software Inc, CA, USA), with P < 0.05 as the significance threshold.
Results
CRC/OXA Cells and Serum Samples Exhibit Increased MCF2L-AS1 Expression
Relative to their corresponding parental cell lines, OXA IC50 values in HT-29/OXA and LOVO/OXA cells was significantly increased (Fig. S1) and MCF2L-AS1 expression was significantly increased in HT-29/OXA and LOVO/OXA cells (Figure 1A). We then assessed relationships between the expression of this lncRNA and chemotherapy responses in CRC patients by analyzing tissue samples (Fig. S2) and serum samples (Figure 1B) from individuals being treated with OXA via qPCR after separating patients as per RECIST criteria into chemotherapy responders (n=56) and non-responders (n=64). This analysis revealed MCF2L-AS1 to be upregulated in non-responders relative to responders with CRC. Kaplan-Meier analyses also revealed the high expression of MCF2L-AS1 levels were associated with shorter CRC patient overall survival (Fig. S3) and non-responders were associated with shorter CRC patient overall survival (Figure 1C).
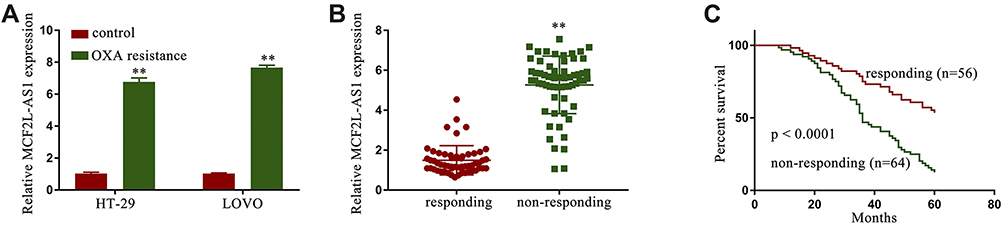 |
Figure 1 The expression of MCF2L-AS1 was enhanced in CRC/OXA cells and samples of serum. MCF2L-AS1 levels were assessed in CRC/OXA cells (A) and serum (B) via qPCR. (C) CRC patient overall survival was assessed via the Kaplan-Meier approach. **p < 0.01. |
MCF2L-AS1 Knockdown Suppresses CRC/OXA Cell Survival and Chemoresistance
To assess the importance of MCF2L-AS1 as a mediator of OXA resistance in CRC, we next transfected sh-MCF2L-AS1 into HT-29/OXA and LOVO/OXA cells to knock down the expression of this lncRNA (Figure 2A). We then treated cells that had been transfected with this shRNA or a corresponding control construct (sh-NC) with a range of OXA concentrations, revealing higher cell viability and OXA IC50 values in the sh-MCF2L-AS1 group as compared to the sh-NC group (Figure 2B and C). These data indicated that knocking down MCF2L-AS1 was sufficient to reduce the survival and chemoresistance of CRC/OXA cells.
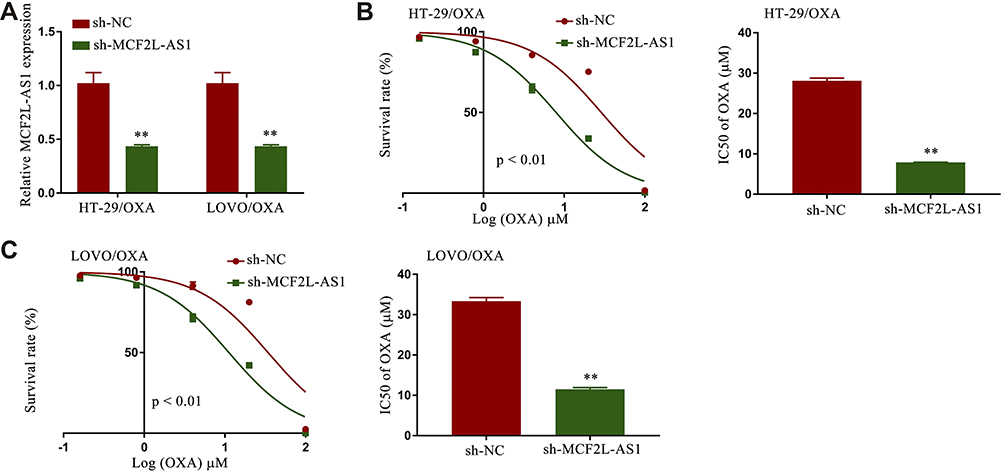 |
Figure 2 Knocking down MCF2L-AS1 suppressed CRC/OXA cell survival and resistance to oxaliplatin. (A) MCF2L-AS1 levels in sh-NC and sh-MCF2L-AS1 CRC/OXA cell groups were assessed via qPCR. OXA IC50 values and cell survival in sh-NC and sh-MCF2L-AS1 groups treated with a range of OXA concentrations were determined via MTT assay for HT-29/OXA (B) and LOVO/OXA (C) cells. **p < 0.01. |
Knocking Down MCF2L-AS1 Suppresses in vivo CRC Tumor OXA Resistance
To more fully understand how MCF2L-AS1 impacts tumor growth, nude mice were subcutaneously implanted with CRC/OXA/sh- MCF2L-AS1 cells and were then treated with OXA. Tumor volumes were significantly lower in mice in the sh-MCF2L-AS1 group that were treated with OXA relative to mice in the sh-NC group that were treated with OXA (Figure 3A and B). Following OXA treatment, overall survival was higher for mice in the sh-MCF2L-AS1 group (Figure 3C and D). Knocking down MCF2L-AS1 thus impaired in vivo OXA resistance.
 |
Figure 3 MCF2L-AS1 knockdown suppresses in vivo OXA resistance. Tumor volume (A and B) and overall survival (C and D) were examined in vivo in OXA-treated sh-NC and sh-MCF2L-AS1 groups. **p < 0.01. |
miR-105 Inhibition Reverses the Effects of MCF2L-AS1 Knockdown on Survival and Chemoresistance in CRC/OXA Cells
To explore whether lncRNA interacted directly with miRNA to regulate the expression of miRNA’s targets in CRC/OXA cells, the position distribution of MCF2L-AS1 in CRC/OXA cells was firstly examined. The results revealed that MCF2L-AS1 was mostly located in cytoplasm of CRC/OXA cells (Fig. S4). Potential interactions between miR-105 and MCF2L-AS1 were predicted using the StartBase 2.0 (http://starbase.sysu.edu.cn/) tool, which identified sequence complementarity between these two ncRNAs (Figure 4A). Additionally, We observed significant reductions in miR-105 expression in serum samples (Fig. S5A) from non-responders and in CRC/OXA cells (Fig. S5B) relative to responders and parental cells, respectively. To validate this putative binding interaction, HEK293T cells were co-transfected with miR-105 mimic or control constructs (NC) and MCF2L-AS1-wt or MCF2L-AS1-mut reporters. This analysis revealed that miR-105 overexpression was sufficient to impair MCF2L-AS1-wt, but not MCF2L-AS1-mut, luciferase activity (Figure 4B). In subsequent pulldown and RIP assays, miR-105 was further confirmed to bind directly to MCF2L-AS1 (Figure 4C–F). These data indicated that MCF2L-AS1 can target miR-105 in CRC. TO better understand the role of this MCF2L-AS1/miR-105 ais in CRC/OXA cells, miR-105 levels were next assessed in cells treated under four different conditions (sh-NC, sh-MCF2L-AS1, sh-MCF2L-AS1 + NC inhibitor, and sh-MCF2L-AS1 + miR-105 inhibitor), revealing a significant increase in miR-105 expression upon r sh-MCF2L-AS1 transfection that could be reversed via miR-105 inhibitor transfection in these CRC/OXA cells (Figure 4G). Survival rates and IC50 values in these groups were then assessed, revealing that the inhibition of miR-105 in both tested cell lines reversed these effects (Figure 4H and I). As such, our findings identified miR-105 as an MCF2L-AS1 target, suggesting that this lncRNA regulates miR-105 to thereby control CRC cell activity.
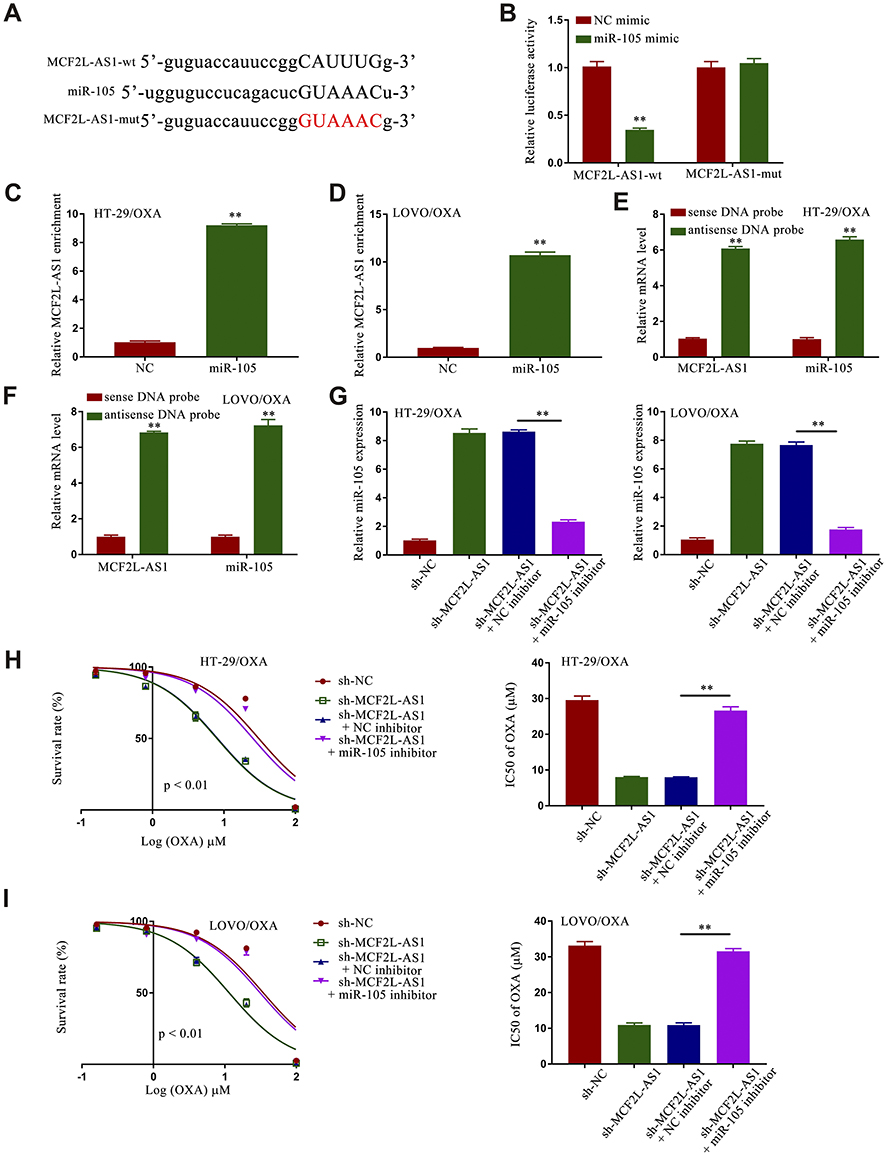 |
Figure 4 Sh-MCF2L-AS1 suppresses CRC/OXA cell survival and oxaliplatin resistance, while miR-105 inhibition reverses this effect. (A) Interactions between miR-105 and MCF2L-AS1 were predicted with StartBase 2.0. (B) Luciferase reporter assays were conducted as a means of measuring luciferase activity. (C and D). MCF2L-AS1 enrichment within CRC/OXA cells was assessed via a RIP assay approach. (E and F). Biotin-coupled sense or antisense DNA probes specific for MCF2L-AS1 were combined with lysates prepared from CRC/OXA cells, after which RNA was pulled down and qPCR was conducted. (G) miR-105 levels in CRC/OXA cells were assessed via qPCR. (H and I). Cell survival and OXA IC50 values were measured via MTT assay for CRC/OXA cells. **p < 0.01. |
miR-105 Overexpression Suppresses IL-1β and Thereby Impairs the Survival and Oxaliplatin Resistance of CRC/OXA Cells
Given that miRNAs can regulate the expression of specific genes, we used the TargetScan application to identify IL-1β as a putative miR-105 target (Figure 5A). Additionally, IL-1β expression was significantly increased in HT-29/OXA and LOVO/OXA cells compared with HT-29 and LOVO cells (Figure S6). In a luciferase reporter gene assay, we confirmed that miR-105 mimic transfection was sufficient to suppress the activity of the WT but not the mutant IL-1β reporter construct (Figure 5B). Overexpressing miR-105 also suppressed IL-1β expression, whereas knocking down this miRNA resulted in the opposite phenotype (Figure 5C and D). The effect of miR-105 mimic transfection on the expression of IL-1β was reversed upon IL-1β transfection (Figure 5E and F). In subsequent experiments, we determined that miR-105 overexpression was sufficient to suppress HT-29/OXA and LOVO/OXA cell survival and to decrease OXA IC50 values in these cells, while IL-1β overexpression partially reversed these changes (Figure 5G and H). Together, these results suggested that IL-1β is a miR-105 target gene, and indicated that miR-105 overexpression can suppress the growth and chemoresistance of CRC/OXA cells growth and drug resistance, while IL-1β upregulation can reverse this phenotype.
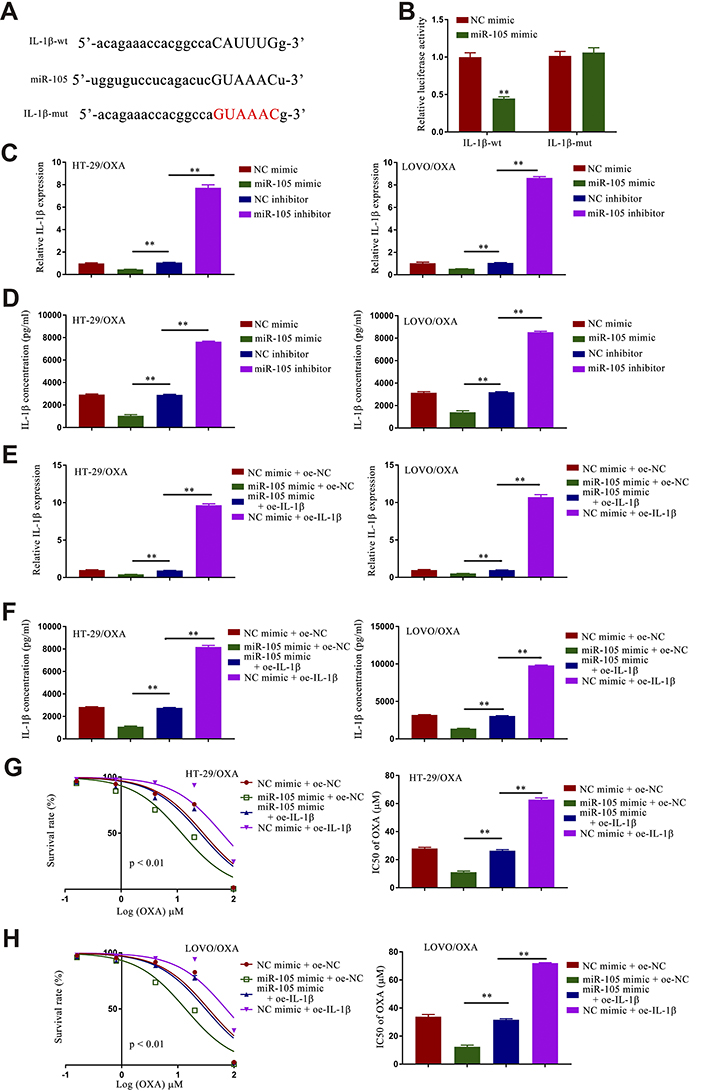 |
Figure 5 miR-105 overexpression results in the inhibition of IL-1β and the suppression of CRC/OXA cell survival and oxaliplatin resistance. (A) Binding interactions between miR-105 and IL-1β were predicted with TargetScanHuman 2.0. (B) Luciferase assays were used to quantify luciferase activity. Levels of IL-1β expression in CRC/OXA cells were quantified via qPCR (C and E) and ELISA (D and F). (G and H). Cell survival and OXA IC50 values were measured via MTT assay for CRC/OXA cells. **p < 0.01. |
MCF2L-AS1 Targets miR-105 to Regulate the Expression of IL-1β in CRC/OXA Cells
To better understand how MCF2L-AS1, miR-105, and IL-1β are associated with one another, we next measured IL-1β expression in the sh-NC + NC inhibitor, sh-MCF2L-AS1 + NC inhibitor, sh-NC + miR-105 inhibitor, and sh-MCF2L-AS1 + miR-105 inhibitor groups via qPCR and ELISA. This revealed that IL-1β was suppressed upon MCF2L-AS1 downregulation in these OXA cells, while miR-105 inhibitor transfection was sufficient to reverse this phenotype (Figure 6A and B), indicating that MCF2L-AS1 can indirectly control the expression of this inflammatory cytokine by binding miR-105.
 |
Figure 6 MCF2L-AS1 targets miR-105 and thereby regulates IL-1β in CRC/OXA cells. The expression of IL-1β in these cells was assessed via qPCR (A) and ELISA (B). **p < 0.01. |
Discussion
Regulatory interactions between different non-coding RNA species and target mRNAs have been identified as key regulators of chemoresistance in a range of oncogenic contexts. In CRC, for example, 5FU resistance is at least partially driven by the lncRNA HOTAIR, which can suppress the expression of miR-218 and thereby promote NF-κB/TS signaling.20 The BLACAT1/miR-519d-3p/CREB1 signaling axis has similarly been shown to control the proliferative, migratory, invasive, survival, and chemoresistance activities of CRC cells,19 while knockdown of the lncRNA SLC25A25-AS1 can enhance CRC cell proliferation, chemoresistance, and EMT progression.21 MCF2L-AS1 has increasingly been identified as an important lncRNA associated with cancer-related glycolysis, proliferation, and invasive activity.6 Herein, we determined that therapeutic non-responder CRC patients expressed substantially higher MCF2L-AS1 levels as compared to those that did respond to chemotherapy. Functional analyses revealed that knocking down MCF2L-AS1 was sufficient to compromise the chemoresistance of CRC/OXA cells in vitro and in vivo, indicating that a loss of this lncRNA can disrupt the mechanisms governing OXA resistance in this oncogenic context.
In several prior reports, miR-105 was shown to be associated with diverse oncogenic processes in the prostate, gastric, non-small-cell lung, and hepatocellular carcinoma (HCC) cancer types.22–25 In prostate cancer, for example, miR-105 can suppress the expression of CDK6 and thereby inhibit tumor growth,22 while in gastric cancer it can similarly suppress SOX9 expression to impair the EMT and metastatic progression.23 In HCC, miR-105 can also impair PI3K/AKT activity and thereby disrupt tumor cell proliferation.24 The importance of miR-105 as a regulator of CRC cell chemoresistance, however, has not been assessed previously. Herein, we detected significant miR-105 downregulation in non-responder CRC patients and CRC/OXA cells compared with responders and chemosensitive cells. Through luciferase reporter, RIP, and pull-down assays, we further confirmed the ability of MCF2L-AS1 to target miR-105 such that knocking down this miRNA in CRC/OXA cells was sufficient to reverse the suppressive effects of sh-MCF2L-AS1 on OXA resistance in these cells, confirming that MCF2L-AS1 controls the chemoresistance of CRC/OXA cells by regulating miR-105.
Many studies have highlighted a role for IL-1β as a regulator of chemoresistance. In breast cancer patients, for example, inflammatory IL-1β responses are associated with tumor metastasis and colonization.26 Herein, we found that miR-105 was able to target and suppress IL-1β expression, with IL-1β overexpression being sufficient to overcome the inhibitory effects of miR-105 overexpression in CRC/OXA cells. MCF2L-AS1 was also shown to regulate the expression of IL-1β by binding to miR-105 within these same cells.
Conclusion
In summary, the knockdown of MCF2L-AS1 was sufficient to overcome the chemoresistance of CRC/OXA cells owing to consequent miR-105 upregulation and IL-1β downregulation, indicating that this MCF2L-AS1/miR-105/IL-1β regulatory axis is an important pathway controlling the therapeutic resistance of CRC/OXA cells.
Acknowledgements
This study was supported by the Wenzhou Basic Science and Technology Project (Y20210925).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Han D, Wang M, Ma N, Xu Y, Jiang Y, Gao X. Long noncoding RNAs: novel players in colorectal cancer. Cancer Lett. 2015;361(1):13–21. doi:10.1016/j.canlet.2015.03.002
2. Li PL, Zhang X, Wang LL, et al. MicroRNA-218 is a prognostic indicator in colorectal cancer and enhances 5-fluorouracil-induced apoptosis by targeting BIRC5. Carcinogenesis. 2015;36(12):1484–1493.
3. Yang AD, Fan F, Camp ER, et al. Chronic oxaliplatin resistance induces epithelial-to-mesenchymal transition in colorectal cancer cell lines. Clin Cancer Res. 2006;12(14 Pt 1):4147–4153. doi:10.1158/1078-0432.CCR-06-0038
4. Alberts SR, Horvath WL, Sternfeld WC, et al. Oxaliplatin, fluorouracil, and leucovorin for patients with unresectable liver-only metastases from colorectal cancer: a North Central Cancer Treatment Group Phase II study. J Clin Oncol. 2005;23(36):9243–9249. doi:10.1200/JCO.2005.07.740
5. Goldberg RM, Sargent DJ, Morton RF, et al. A randomized controlled trial of fluorouracil plus leucovorin, irinotecan, and oxaliplatin combinations in patients with previously untreated metastatic colorectal cancer. J Clin Oncol. 2004;22(1):23–30. doi:10.1200/JCO.2004.09.046
6. Zhang Z, Yang W, Li N, et al. LncRNA MCF2L-AS1 aggravates proliferation, invasion and glycolysis of colorectal cancer cells via the crosstalk with miR-874-3p/FOXM1 signaling axis. Carcinogenesis. 2021;42(2):263–271. doi:10.1093/carcin/bgaa093
7. Wang Z, Xu R. lncRNA PART1 promotes breast cancer cell progression by directly targeting miR-4516. Cancer Manag Res. 2020;12:7753–7760. doi:10.2147/CMAR.S249296
8. Li H, Yao G, Zhai J, Hu D, Fan Y. LncRNA FTX promotes proliferation and invasion of gastric cancer via miR-144/ZFX axis. Onco Targets Ther. 2019;12:11701–11713. doi:10.2147/OTT.S220998
9. Wu C, Yang J, Li R, Lin X, Wu J, Wu J. LncRNA WT1-AS/miR-494-3p regulates cell proliferation, apoptosis, migration and invasion via PTEN/PI3K/AKT signaling pathway in non-small cell lung cancer. Onco Targets Ther. 2021;14:891–904. doi:10.2147/OTT.S278233
10. Zhang W, Yuan W, Song J, Wang S, Gu X. LncRna CPS1-IT1 suppresses cell proliferation, invasion and metastasis in colorectal cancer. Cell Physiol Biochem. 2017;44(2):567–580. doi:10.1159/000485091
11. Qi FF, Yang Y, Zhang H, Chen H. Long non-coding RNAs: key regulators in oxaliplatin resistance of colorectal cancer. Biomed Pharmacother. 2020;128:110329. doi:10.1016/j.biopha.2020.110329
12. Ni Y, Li C, Bo C, et al. LncRNA EGOT regulates the proliferation and apoptosis of colorectal cancer by miR-33b-5p/CROT axis. Biosci Rep. 2020. doi:10.1042/BSR20193893
13. Sun Y, Liu Y, Cai Y, et al. Downregulation of LINC00958 inhibits proliferation, invasion and migration, and promotes apoptosis of colorectal cancer cells by targeting miR36195p. Oncol Rep. 2020;44(4):1574–1582.
14. Gao R, Fang C, Xu J, Tan H, Li P, Ma L. LncRNA CACS15 contributes to oxaliplatin resistance in colorectal cancer by positively regulating ABCC1 through sponging miR-145. Arch Biochem Biophys. 2019;663:183–191. doi:10.1016/j.abb.2019.01.005
15. Karreth FA, Pandolfi PP. ceRNA cross-talk in cancer: when ce-bling rivalries go awry. Cancer Discov. 2013;3(10):1113–1121. doi:10.1158/2159-8290.CD-13-0202
16. Jiang Z, Li L, Hou Z, et al. LncRNA HAND2-AS1 inhibits 5-fluorouracil resistance by modulating miR-20a/PDCD4 axis in colorectal cancer. Cell Signal. 2020;66:109483. doi:10.1016/j.cellsig.2019.109483
17. Liu A, Liu L, Lu H. LncRNA XIST facilitates proliferation and epithelial-mesenchymal transition of colorectal cancer cells through targeting miR-486-5p and promoting neuropilin-2. J Cell Physiol. 2019;234(8):13747–13761. doi:10.1002/jcp.28054
18. Hui Y, Yang Y, Li D, et al. LncRNA FEZF1-AS1 modulates cancer stem cell properties of human gastric cancer through miR-363-3p/HMGA2. Cell Transplant. 2020;29:963689720925059. doi:10.1177/0963689720925059
19. Chen R, Zhou S, Chen J, Lin S, Ye F, Jiang P. LncRNA BLACAT1/miR-519d-3p/CREB1 axis mediates proliferation, apoptosis, migration, invasion, and drug-resistance in colorectal cancer progression. Cancer Manag Res. 2020;12:13137–13148. doi:10.2147/CMAR.S274447
20. Li P, Zhang X, Wang L, et al. lncRNA HOTAIR contributes to 5FU resistance through suppressing miR-218 and activating NF-kappaB/TS signaling in colorectal cancer. Mol Ther Nucleic Acids. 2020;20:879–880. doi:10.1016/j.omtn.2020.05.016
21. Li Y, Huang S, Li Y, et al. Decreased expression of LncRNA SLC25A25-AS1 promotes proliferation, chemoresistance, and EMT in colorectal cancer cells. Tumour Biol. 2016;37(10):14205–14215. doi:10.1007/s13277-016-5254-0
22. Honeywell DR, Cabrita MA, Zhao H, Dimitroulakos J, Addison CL. miR-105 inhibits prostate tumour growth by suppressing CDK6 levels. PLoS One. 2013;8(8):e70515. doi:10.1371/journal.pone.0070515
23. Shang JC, Yu GZ, Ji ZW, Wang XQ, Xia L. MiR-105 inhibits gastric cancer cells metastasis, epithelial-mesenchymal transition by targeting SOX9. Eur Rev Med Pharmacol Sci. 2019;23(14):6160–6169.
24. Shen G, Rong X, Zhao J, et al. MicroRNA-105 suppresses cell proliferation and inhibits PI3K/AKT signaling in human hepatocellular carcinoma. Carcinogenesis. 2014;35(12):2748–2755. doi:10.1093/carcin/bgu208
25. Lu G, Fu D, Jia C, et al. Reduced miR-105-1 levels are associated with poor survival of patients with non-small cell lung cancer. Oncol Lett. 2017;14(6):7842–7848.
26. Castano Z, San Juan BP, Spiegel A, et al. IL-1beta inflammatory response driven by primary breast cancer prevents metastasis-initiating cell colonization. Nat Cell Biol. 2018;20(9):1084–1097. doi:10.1038/s41556-018-0173-5
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.