")
Back to Journals » OncoTargets and Therapy » Volume 10
let-7d suppresses proliferation and invasion and promotes apoptosis of meningioma by targeting AEG-1
Received 3 May 2017
Accepted for publication 28 June 2017
Published 6 October 2017 Volume 2017:10 Pages 4895—4904
DOI https://doi.org/10.2147/OTT.S141008
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Yao Dai
Hui Li, Jianmin Zhao
Department of Neurology, Xinxiang Central Hospital, Xinxiang, China
Background: let-7d has been indicated to act as a tumor suppressor in various cancers. However, the function and molecular mechanism of let-7d in meningioma progression have not been elucidated.
Materials and methods: Quantitative real-time polymerase chain reaction (qRT-PCR) was performed to detect the expression levels of let-7d and AEG-1 mRNA in meningioma tissues and cell lines. The protein level of AEG-1 was measured by Western blot analysis. MTT assay, Transwell invasion assay and flow cytometry analysis were carried out to determine the proliferation, invasion and apoptosis of IOMM-Lee and CH-157MN cells, respectively. Target gene of let-7d was verified by luciferase reporter analysis.
Results: let-7d expression was downregulated, and AEG-1 expression was upregulated in meningioma tumor tissues. let-7d overexpression suppressed proliferation and invasion and induced apoptosis in IOMM-Lee and CH-157MN cells. Moreover, AEG-1 was a direct target of let-7d. Restoration of AEG-1 expression reversed let-7d-mediated suppression of the proliferation and invasion and let-7d-induced apoptosis in IOMM-Lee and CH-157MN cells.
Conclusion: let-7d repressed proliferation and invasion and promoted apoptosis of meningioma cells by targeting AEG-1. The present study provided a better understanding of the meningioma pathogenesis and a promising therapeutic target for meningioma patients.
Keywords: let-7d, AEG-1, meningioma cells, therapeutic target
Introduction
With an incidence rate of 4.4/100,000 individuals, meningiomas are the common prevalent neoplasms of central nervous system and occupied ~30% of all intracranial tumors.1,2 According to the World Health Organization (WHO) classification, meningiomas have been divided into three grades, including benign (Grade I), atypical (Grade II) or anaplastic/malignant (Grade III).3 Owing to the invasion and recurrence of malignant meningioma, the estimated overall 10-year survival rate is only ~15%.4 Over the last few years, an increasing attention has been paid to the molecular genetics of meningiomas.5 Specific gene dysfunctions have been implicated in the pathogenesis of these tumors, including the inactivation of tumor suppressor genes or excessive expression of oncogenes.6 However, a thorough understanding of the molecular basis of this tumor remains far from being well established. Therefore, further efforts are urgently required to provide a foundation for potential gene therapies and individualized treatments for meningiomas.
MicroRNAs (miRNAs), a class of naturally occurring and highly conserved small noncoding RNAs with ~22 nucleotides in length, are able to interact with the 3′ untranslated region (UTR) of target mRNAs based on sequence complementarity, leading to mRNA degradation or translation inhibition.7 Increasing evidence has demonstrated that miRNAs play a crucial role in the development and progression of multiple cancers by serving as oncogenes or as tumor suppressors.8 let-7 family members, highly conserved across diverse animal species, have been found to be low expressed in many cancer types.9,10 let-7d, a member of the let-7 family, has been described to be involved in the development and progression of leukemia,11 prostate cancer,12 ovarian cancer13 and head and neck squamous cell carcinoma (HNSCC).14 Moreover, let-7d was reported to be downregulated in meningioma tumor tissues.15 However, the functional action and mechanism of let-7d in meningioma development and progression have not been elucidated.
Astrocyte elevated gene-1 (AEG-1), known as metadherin (MDTH), was initially characterized as a neuropathology-related gene after HIV-1 infection or exposure to HIV envelope glycoprotein.16 Ample evidence revealed that AEG-1 functions as an oncogene by promoting cell proliferation, invasiveness and epithelial–mesenchymal transition (EMT) in many kinds of cancers, such as lung cancer,17 cervical cancer18 and esophageal squamous cell carcinoma.19 Recently, a report showed that AEG-1 contributed to the malignant progression of meningiomas.20 However, how AEG-1 is regulated in meningiomas is still unclear.
In the present study, the function and action mechanism of let-7d in meningioma progression were investigated.
Materials and methods
Tissue specimens
Seventeen meningioma samples (six WHO grade I, five WHO grade II and six WHO grade III) were obtained from resected tumor tissues during surgery, and seven normal arachnoidal samples were acquired from fresh autopsies. All tissue specimens were snap-frozen and maintained at −80°C for subsequent analysis. Ethic review committees of Xinxiang Central Hospital approved the experimental procedures. Informed consent form was signed by each patient.
Cell culture
Human meningioma cell lines IOMM-Lee and CH-157MN were obtained from Shanghai Institute of Cell Research (Shanghai, China). Human benign meningioma cell line BenMen1 was purchased from Chinese Academy of Sciences (Shanghai, China). Human benign meningioma cell line HBL52 was provided by Cell Lines Service and Cellbank (Heidelberg, Germany). All cells were grown in Dulbecco’s Modified Eagle’s Medium (DMEM; Thermo Fisher Scientific, Waltham, MA, USA) with 10% fetal bovine serum (FBS; Thermo Fisher Scientific) and 1% penicillin/streptomycin (Thermo Fisher Scientific) at 37°C in humidified 5% CO2.
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNAs were isolated from meningioma tissues and cell lines using TRIzol (Thermo Fisher Scientific). The expression of miRNA was quantified using TaqMan MicroRNA Assays (Thermo Fisher Scientific) and normalized to U6 snRNA expression. Relative levels of mRNA were measured with SYBR green qRT-PCR (Thermo Fisher Scientific) and normalized to β-actin expression. The primers for DNA amplification were as follows: let-7d, 5′ ACACTCCAGCTGGGTGAGGTAGTAGATTGAATA 3′ (forward) and 5′AACTGGTGTCGTGGAG 3′ (reverse); U6, 5′ CTCGCTTCGGCAGCACA 3′ (forward) and 5′ TGGTGTCGTGGAGTCG 3′ (reverse); AEG-1, 5′ TGCCTCCTTCACAGACCAA 3′ (forward) and 5′ TCGGCTGCAGATGAGATAG 3′ (reverse) and β-actin, 5′ TGAGAGGGAAATCGTGCGTGAC 3′ (forward) and 5′ AAGAAGGAAGGCTGGAAAAGAG 3′ (reverse).
The relative level of let-7d and AEG-1 was analyzed using the 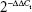 method.21
method.21
Western blot analysis
Western blot analysis was performed as previously described.22 All proteins were resolved on a 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto a nitrocellulose membrane, followed by incubating with primary antibodies anti-AEG-1 (Abcam, Cambridge, MA, USA) and anti-β-actin (Abcam). Protein bands were detected with enhanced chemiluminescence reagent (EMD Millipore, Billerica, MA, USA).
Cell transfection
pcDNA-AEG-1 plasmid was established by inserting the full-length sequence of AEG-1 into pcDNA vector (Thermo Fisher Scientific). To manipulate the expression of AEG-1 and let-7d, IOMM-Lee and CH-157MN cells were transfected with let-7d mimics (GenePharma, Shanghai, China), let-7d inhibitor (GenePharma) or pcDNA-AEG-1 using Lipofectamine 2000 (Thermo Fisher Scientific). Scramble RNA (miR-control) and pcDNA empty vector were used as negative controls.
Cell viability assay
The cell viability of IOMM-Lee and CH-157MN was assessed by MTT assay. Transfected cells were cultured for 48 h in 96-well plates and then incubated with 20 μL MTT (0.5 mg/mL; Sigma-Aldrich Co., St Louis, MO, USA) for 4 h, followed by discarding of the culture supernatant and addition of 150 μL of dimethyl sulfoxide (DMSO; Sigma-Aldrich Co.). The absorbance at 450 nm was determined using a microtiter plate reader (Molecular Devices LLC, Sunnyvale, CA, USA).
Cell invasion assay
The invasion ability of IOMM-Lee and CH-157MN cells was determined by Transwell invasion assay. Transfected cells in DMEM serum-free medium were placed into the upper chamber coated with Matrigel (BD Biosciences, San Jose, CA, USA). DMEM medium with 5% FBS was added to the lower chamber. After 48 h of incubation, cells passing through the lower surface of the membrane were fixed with 4% paraformaldehyde, stained with 0.5% crystal violet and counted using a microscope (Olympus Corporation, Tokyo, Japan). The experiments were independently repeated three times.
Apoptosis assay
Apoptosis was determined using Annexin V/FITC Apoptosis Detection kit (BD Biosciences). Briefly, IOMM-Lee and CH-157MN cells were cultured for 48 h after transfection. Then, the cells were stained by incubating with Annexin V-FITC and propidium iodide (PI) in the dark for 15 min. Finally, the data were acquired by FACSCanto II flow cytometer with BD FACSDiva software V6.1.3 (BD Biosciences).
Luciferase reporter assays
The wild-type 3′ UTR of AEG-1 (AEG-1-wt) was chemically synthesized and cloned into the luciferase reporter vector PGL3 (Promega Corporation, Fitchburg, WI, USA). Mutant AEG-1 3′ UTR (AEG-1-mut) construct was performed using QuickChange II Site-Directed Mutagenesis Kit (Agilent Technologies, Santa Clara, CA, USA) and introduced into pGL3 vector. Subsequently, IOMM-Lee and CH-157MN cells were co-transfected with miR-control or let-7d and wild- or mutant-type reporter plasmids (AEG-1-wt or AEG-1-mut). Luciferase activities were measured using the Dual-Luciferase Reporter Assay System (Promega Corporation) at 48 h post transfection.
Statistical analysis
Data are presented as mean ± SD values for three independent experiments. Data were analyzed using SPSS 19.0 software (SPSS, IBM Corporation, Armonk, NY, USA). Statistical comparisons were assessed using Student’s t-test or one-way ANOVA. Differences were considered to be statistically significant when P<0.05.
Results
let-7d expression is decreased and AEG-1 expression is elevated in meningiomas
To determine whether let-7d and AEG-1 were involved in meningioma carcinogenesis, the expression of let-7d and AEG-1 mRNA was first detected by qRT-PCR analysis in 17 meningioma tissue samples (six WHO grade I, five WHO grade II and six WHO grade III) and seven normal arachnoidal tissue samples. As shown in Figure 1A and B, let-7d expression was significantly downregulated and AEG-1 mRNA level was significantly upregulated in meningioma tissues when compared with that in non-neoplastic arachnoidal tissues. In addition, an inverse correlation between let-7d and AEG-1 expressions was observed in meningioma tissues (r=−0.6993, P=0.0018) (Figure 1C). Moreover, our results displayed that let-7d expression in atypical (WHO grade II) and anaplastic meningiomas (WHO grade III) was significantly lower than that in benign meningiomas (WHO grade I; Figure 1D). On the contrary, AEG-1 level increased with the ascending of pathological tumor grade (Figure 1E). Next, qRT-PCR analysis was performed to measure let-7d expression in four meningioma cell lines (IOMM-Lee, CH-157MN, BenMen1 and HBL52). Malignant cell lines (IOMM-Lee and CH-157MN) exhibited lower let-7d expression than BenMen1 and HBL52 cells derived from benign meningiomas (Figure 1F). Thus, IOMM-Lee and CH-157MN cells were used in the subsequent experiments. All these results suggested that dysregulated expression of let-7d and AEG-1 may be involved in the pathogenesis and progression of meningioma.
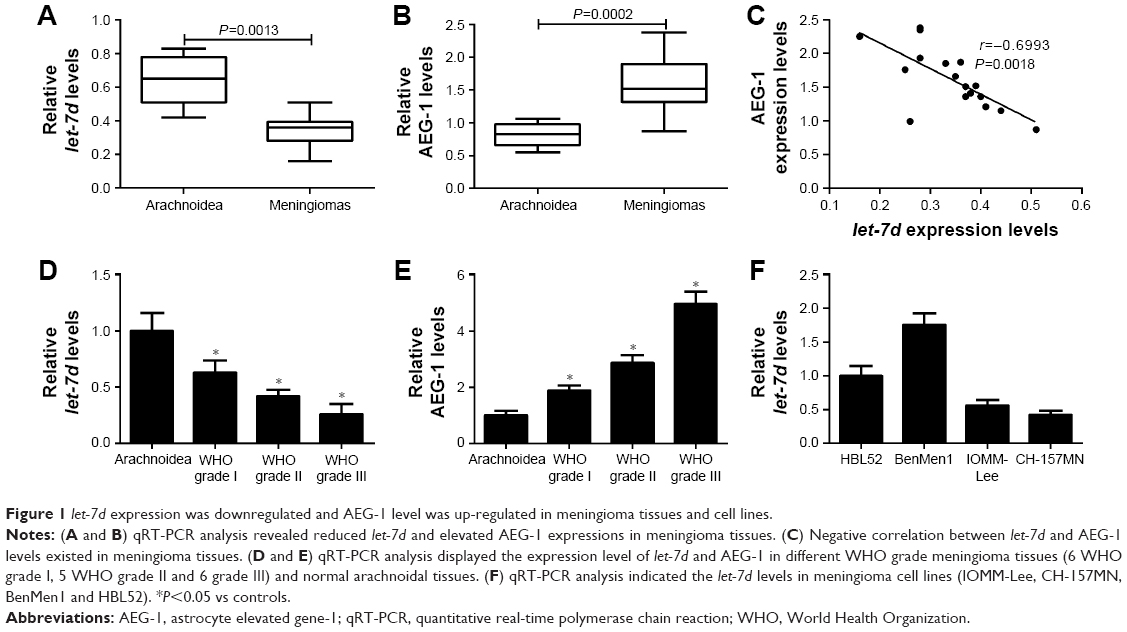 | Figure 1 let-7d expression was downregulated and AEG-1 level was up-regulated in meningioma tissues and cell lines. |
let-7d overexpression inhibits cell viability and invasion and induces apoptosis in meningioma cells
To investigate the functional role of let-7d in meningioma cells, gain-of-function experiments were performed in IOMM-Lee and CH-157MN cells by transfecting with miR-control or let-7d mimics. First, the transfection efficiency was validated with the fact that let-7d expression was significantly upregulated in IOMM-Lee and CH-157MN cells transfected with let-7d mimics (Figure 2A). Then, the effect of let-7d overexpression on the cell viability, invasive ability and apoptotic activity was determined by MTT, Transwell invasion assays and flow cytometry analysis, respectively. The results of MTT assay indicated that let-7d-overexpressing IOMM-Lee and CH-157MN cells displayed a substantially lower cell viability compared with that in the control groups (Figure 2B). Transwell invasion assay revealed that let-7d overexpression led to a significant reduction in the invasion ability of IOMM-Lee and CH-157MN cells when compared with the miR-control-transfected cells (Figure 2C). Furthermore, the apoptotic rate of IOMM-Lee and CH-157MN cells was remarkedly elevated by let-7d mimics transfection (Figure 2D). Taken together, these results demonstrated that let-7d suppressed the development of meningioma.
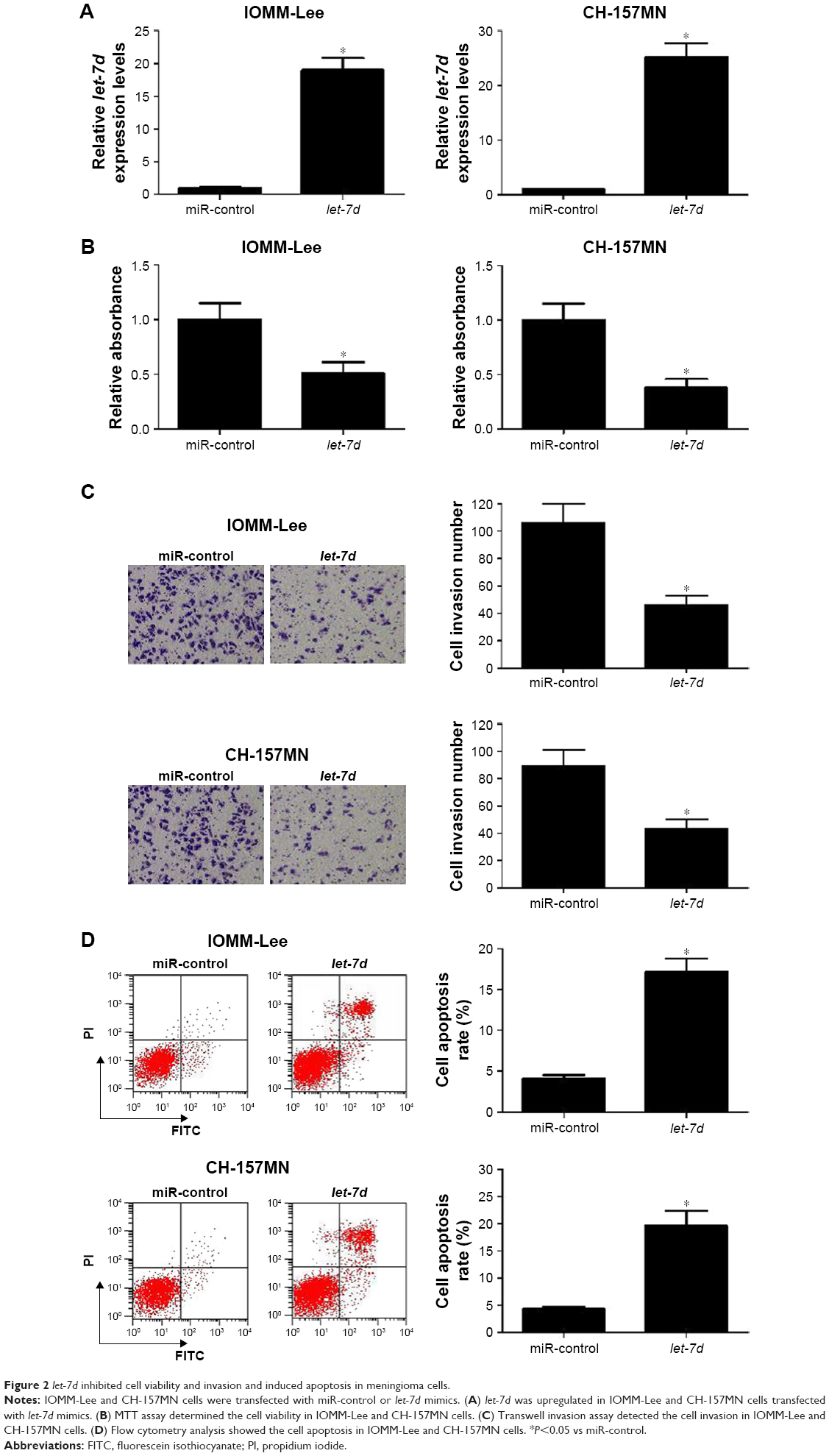 | Figure 2 let-7d inhibited cell viability and invasion and induced apoptosis in meningioma cells. |
let-7d targets AEG-1
Collecting evidence suggested that miRNAs play functional actions by suppressing their special target genes. Therefore, target prediction of let-7d was conducted by using online TargetScan software (Whitehead Institute for Biomedical Research, Cambridge, MA, USA). As shown in Figure 3A, AEG-1 3′ UTR contains a binding site of let-7d. To further verify that AEG-1 was a target gene of let-7d, dual luciferase reporter assay was performed. let-7d mimics remarkedly reduced the luciferase activity of AEG-1-wt reporter in IOMM-Lee and CH-157MN cells; however, no significant change was observed in the luciferase activity of AEG-1-mut reporter between let-7d and miR-control groups (Figure 3B). To further confirm the actual effect of let-7d on AEG-1 expression, qRT-PCR and Western blot analyses were performed to detect the mRNA and protein level of AEG-1 in let-7d mimics- or inhibitor-transfected cells. The results indicated that ectopic expression of let-7d suppressed AEG-1 expression at miRNA and protein levels, while let-7d inhibition improved the AEG-1 level in IOMM-Lee and CH-157MN cells (Figure 3C and D). Collectively, these data implied that let-7d directly inhibited AEG-1 expression.
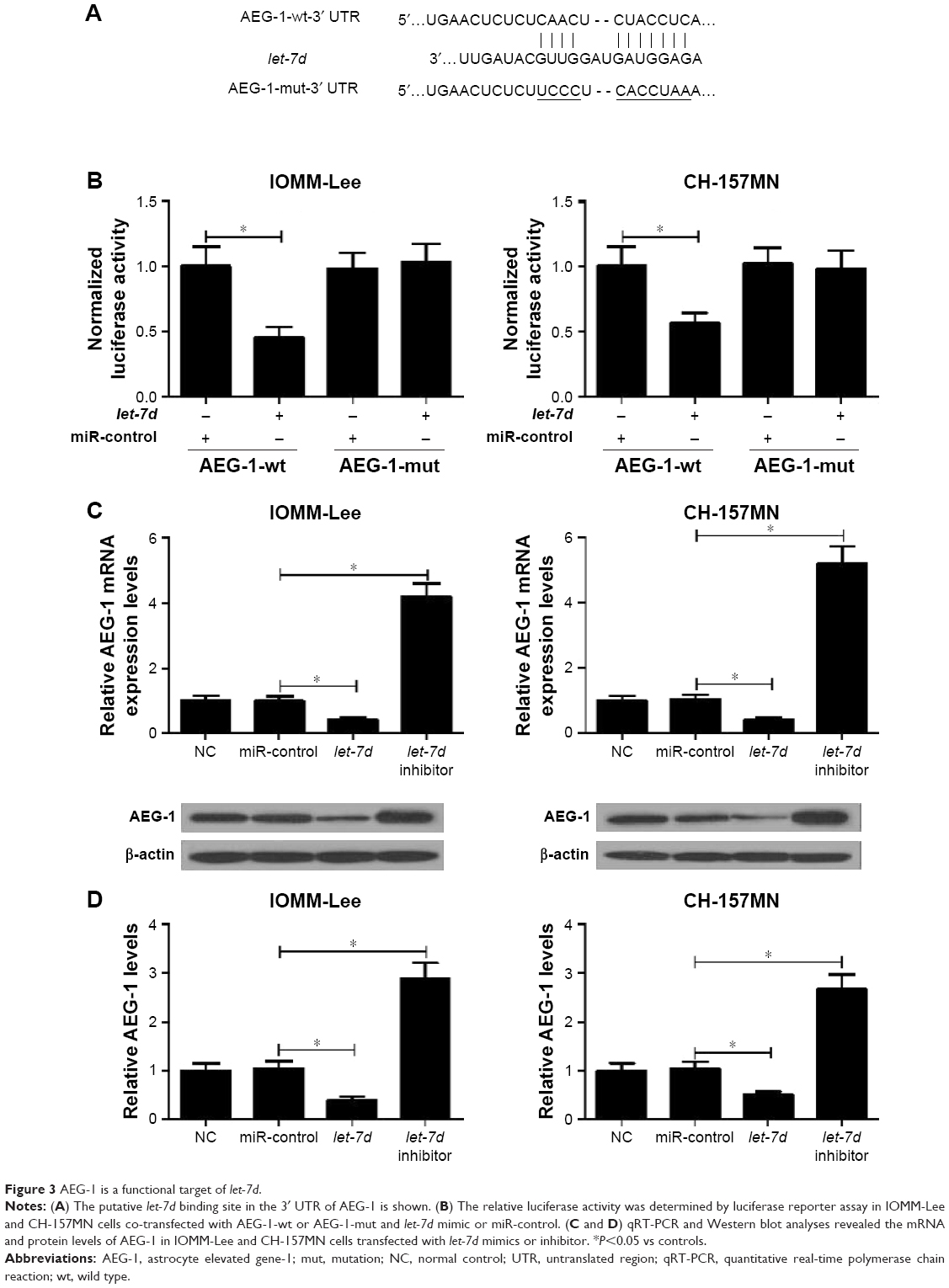 | Figure 3 AEG-1 is a functional target of let-7d. |
Upregulation of AEG-1 reverses the effects of let-7d overexpression on proliferation, invasion and apoptosis of meningioma cells
To further investigate whether let-7d exerted its antitumor effect in meningioma progression by targeting AEG-1, IOMM-Lee and CH-157MN cells were transfected with let-7d or co-transfected with let-7d and pcDNA-AEG-1. MTT assay revealed that the inhibitory effect of let-7d on the cell viability of IOMM-Lee and CH-157MN cells was eliminated after pcDNA-AEG-1 transfection (Figure 4A). Transwell invasion assay showed that exogenetic expression of AEG-1 expression abolished let-7d-mediated suppression on IOMM-Lee and CH-157MN cell invasion (Figure 4B). Furthermore, overexpression of AEG-1 abrogated let-7d-induced apoptosis of IOMM-Lee and CH-157MN cells (Figure 4C). Taken together, the data confirmed that let-7d suppressed the tumorigenesis of meningioma by targeting AEG-1.
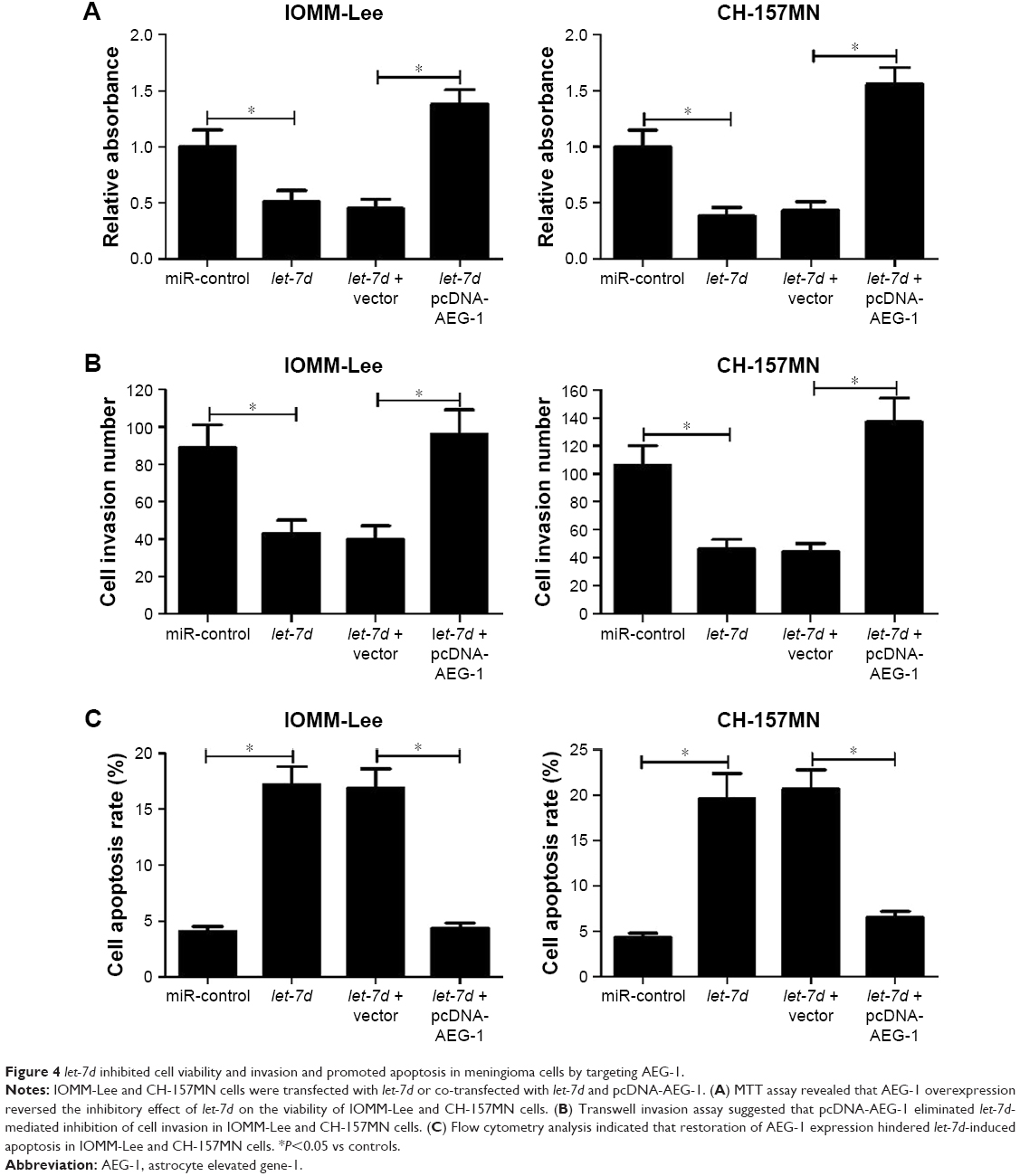 | Figure 4 let-7d inhibited cell viability and invasion and promoted apoptosis in meningioma cells by targeting AEG-1. |
To further confirm the functional role of AEG-1 in meningioma, IOMM-Lee cells were transfected with pcDNA-AEG-1 or pcDNA-control. First, the result of Western blot analysis indicated that AEG-1 was successfully overexpressed in pcDNA-AEG-1 transfecting IOMM-Lee cells (Figure 5A). Then, the cell viability, invasion and apoptosis were determined in IOMM-Lee cells. As expected, AEG-1 overexpression increased the viability and invasion of IOMM-Lee cells (Figure 5B and C). However, AEG-1 overexpression did not lead to significant decrease in IOMM-Lee cell apoptosis (Figure 5D), which may due to that the apoptotic rate in IOMM-Lee cells had reached the lowermost.
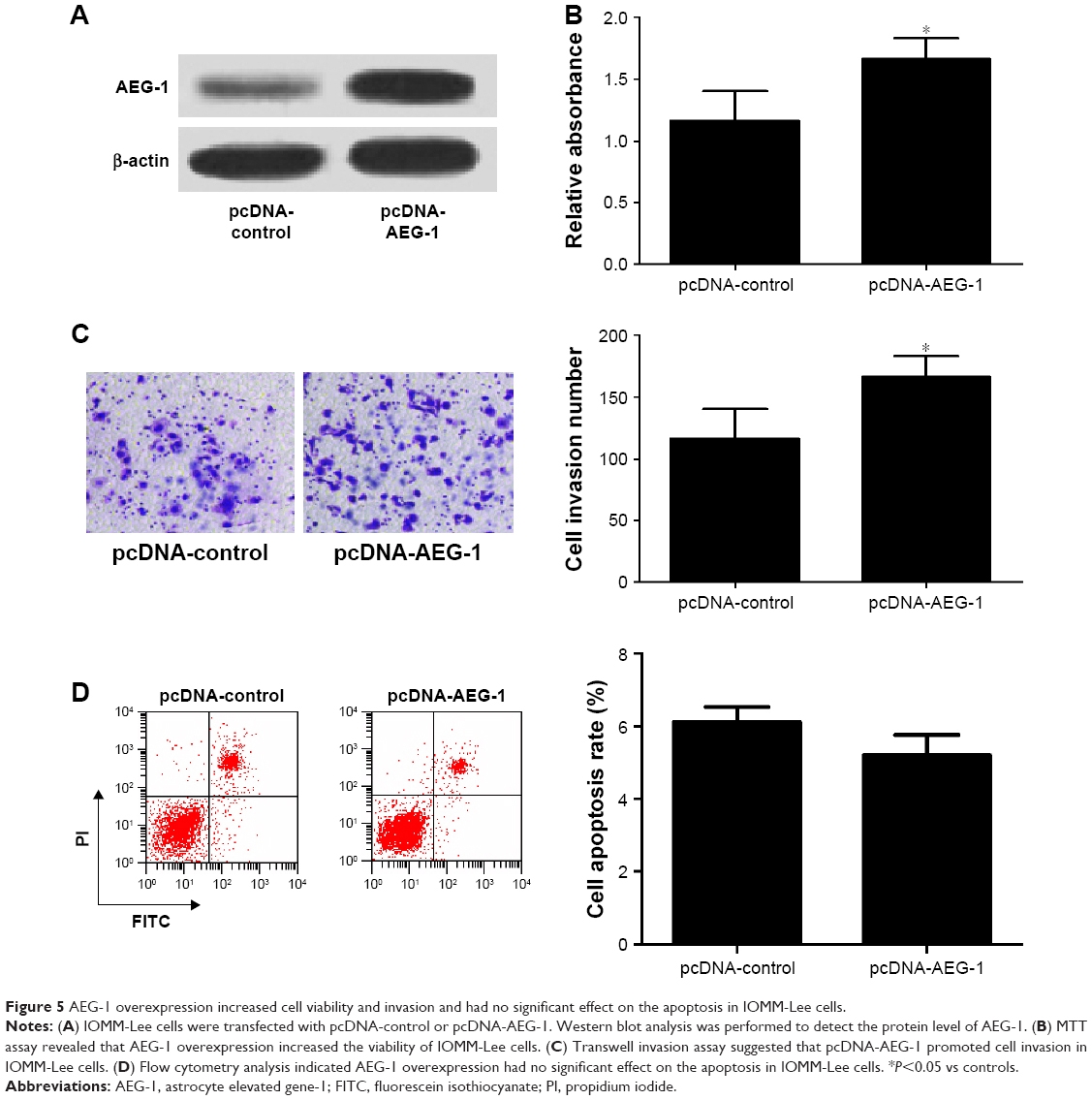 | Figure 5 AEG-1 overexpression increased cell viability and invasion and had no significant effect on the apoptosis in IOMM-Lee cells. |
Discussion
Recently, increasing evidence suggested that dysregulation of miRNAs is involved in the oncogenesis of various cancers, including meningioma. For example, Werner et al23 reported that miR-34a-3p was significantly downregulated in WHO grade II and III meningiomas, and miR-34a-3p overexpression suppressed proliferation and induced apoptosis of meningioma cells in vitro by directly targeting SMAD4, FRAT1 and BCL2. Overexpressed miR-200a inhibited meningioma cell migration and tumor growth by directly targeting non-muscle heavy chain IIb (NMHCIIb), playing key roles in cell division and cell migration.24 Another study revealed that high-expressed miR-21 expression and low-expressed miR-107 were closely associated with histopathological grade of meningiomas.25 miRNA-145 was found to be downregulated in meningioma, and elevated miR-145 expression in IOMM-Lee meningioma cells led to reduced proliferation, induced apoptosis and impaired migratory and invasive potential.15 Moreover, this study also revealed a significant downregulation of let-7d expression in meningioma tumor tissues.15 However, the function and mechanism of let-7d in meningioma development and progression are still unclear. Therefore, the present study mainly investigated the role of dysregulated let-7d in meningioma.
Accumulating documents pointed that let-7d was lowly expressed in several cancers and functions as a tumor suppressor.10 Low expression of let-7d was found in HNSCC patients and was predictive of poor survival.14 let-7d inhibited renal cell carcinoma growth, metastasis and tumor macrophage infiltration at least partially by downregulating COL3A1 and CCL7.26 let-7d was also implied to suppress EMT expression and chemoresistant ability in oral cancer.27 However, the role of let-7d in meningioma is still not illuminated. In the present study, we found that let-7d was lowly expressed in meningioma tissues and cells, and restoration of let-7d expression suppressed cell viability and invasion and induced apoptosis in meningioma cells. Our study first demonstrated that let-7d functioned as a tumor suppressor in meningioma progression, which may provide a novel therapeutic target for patients with meningioma.
Emerging evidence suggests the involvement of AEG-1 in many cancers. For example, AEG-1 knockdown led to the apoptosis induction in prostate cancer through activating forkhead box 3a (FOXO3a).28 Elevated AEG-1 expression was linked to progression of cervical intraepithelial neoplasia and represented a poor prognosis in cervical cancer.29 In human retinoblastoma cells, downregulation of AEG-1 displayed a tumor suppressive effect by inhibiting growth and inducing apoptosis via MAPK pathways.30 In the present study, to investigate the functional role of AEG-1 in the progression of meningioma, AEG-1 was knocked down in IOMM-Lee and CH-157MN cells. AEG-1 downregulation resulted in the inhibition of cell viability and invasive ability and the enhancement of apoptosis. A recent study revealed that knockdown of AEG-1 suppressed cell growth and induced apoptosis and blocked invasion in ovarian cancer cells,31 which was consistent with our findings. Moreover, AEG-1 was verified to be a target of let-7d. Restoration of AEG-1 expression counteracted the inhibitory effect of let-7d on the cell viability and invasion and let-7d-induced apoptosis in meningioma cells.
Conclusion
In summary, our study demonstrates that let-7d overexpression suppresses proliferation and invasion and induces apoptosis in meningioma cells. More importantly, the inhibitory effect of let-7d on meningioma progression is mediated by its functional target AEG-1. Altogether, our study suggests that let-7d may provide a novel therapy for meningioma patients in the future.
Disclosure
The authors report no conflicts of interest in this work.
References
Porter KR, Mccarthy BJ, Freels S, Kim Y, Davis FG. Prevalence estimates for primary brain tumors in the United States by age, gender, behavior, and histology. Neuro Oncol. 2010;12(6):520–527. | ||
Ostrom QT, Gittleman H, Farah P, et al. CBTRUS statistical report: primary brain and central nervous system tumors diagnosed in the United States in 2006–2010. Neuro Oncol. 2013;15(Sup 2):ii1–ii56. | ||
Louis DN, Ohgaki H, Wiestler OD, et al. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007;114(2):97–109. | ||
He S, Pham MH, Pease M, et al. A review of epigenetic and gene expression alterations associated with intracranial meningiomas. Neurosurg Focus. 2013;35(6):E5. | ||
Vranic A, Peyre M, Kalamarides M. New insights into meningioma: from genetics to trials. Curr Opin Oncol. 2012;24(6):660–665. | ||
Ozer O, Sahin FI, Aydemir F, Ozen O, Yilmaz Z, Altinörs N. Her-2/neu gene amplification in paraffin-embedded tissue sections of meningioma patients. Turk Neurosurg. 2009;19(2):135–138. | ||
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. | ||
Hayes J, Peruzzi PP, Lawler S. MicroRNAs in cancer: biomarkers, functions and therapy. Trends Mol Med. 2014;20(8):460–469. | ||
Schultz J, Lorenz P, Gross G, Ibrahim S, Kunz M. MicroRNA let-7b targets important cell cycle molecules in malignant melanoma cells and interferes with anchorage-independent growth. Cell Res. 2008;18(5):549–557. | ||
Boyerinas B, Park S-M, Hau A, Murmann AE, Peter ME. The role of let-7 in cell differentiation and cancer. Endocr Relat Cancer. 2010;17(1):F19–F36. | ||
Garzon R, Pichiorri F, Palumbo T, et al. MicroRNA gene expression during retinoic acid-induced differentiation of human acute promyelocytic leukemia. Oncogene. 2007;26(28):4148–4157. | ||
Ramberg H, Alshbib A, Berge V, Svindland A, Taskén KA. Regulation of PBX3 expression by androgen and let-7d in prostate cancer. Mol Cancer. 2011;10(1):50. | ||
Dahiya N, Sherman-Baust CA, Wang TL, et al. MicroRNA expression and identification of putative miRNA targets in ovarian cancer. PLoS One. 2008;3(6):e2436. | ||
Childs G, Fazzari M, Kung G, et al. Low-level expression of microRNAs let-7d and miR-205 are prognostic markers of head and neck squamous cell carcinoma. Am J Pathol. 2009;174(3):736–745. | ||
Kliese N, Gobrecht P, Pachow D, et al. miRNA-145 is downregulated in atypical and anaplastic meningiomas and negatively regulates motility and proliferation of meningioma cells. Oncogene. 2013;32(39):4712–4720. | ||
Su ZZ, Kang DC, Chen Y, et al. Identification and cloning of human astrocyte genes displaying elevated expression after infection with HIV-1 or exposure to HIV-1 envelope glycoprotein by rapid subtraction hybridization, RaSH. Oncogene. 2002;21(22):3592–3602. | ||
He W, He S, Wang Z, et al. Astrocyte elevated gene-1 (AEG-1) induces epithelial-mesenchymal transition in lung cancer through activating Wnt/β-catenin signaling. BMC Cancer. 2015;15(1):1–13. | ||
Liu X, Wang D, Liu H, et al. Knockdown of astrocyte elevated gene-1 (AEG-1) in cervical cancer cells decreases their invasiveness, epithelial to mesenchymal transition, and chemoresistance. Cell Cycle. 2014;13(11):1702–1707. | ||
Yu C, Chen K, Zheng H, et al. Overexpression of astrocyte elevated gene-1 (AEG-1) is associated with esophageal squamous cell carcinoma (ESCC) progression and pathogenesis. Carcinogenesis. 2009;30(5):894–901. | ||
Park KJ, Yu MO, Song NH, et al. Expression of astrocyte elevated gene-1 (AEG-1) in human meningiomas and its roles in cell proliferation and survival. J Neurooncol. 2015;121(1):31–39. | ||
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25(4):402–408. | ||
Shanmugam MK, Manu KA, Ong TH, et al. Inhibition of CXCR4/CXCL12 signaling axis by ursolic acid leads to suppression of metastasis in transgenic adenocarcinoma of mouse prostate model. Int J Cancer. 2011;129(7):1552–1563. | ||
Werner TV, Hart M, Nickels R, et al. MiR-34a-3p alters proliferation and apoptosis of meningioma cells in vitro and is directly targeting SMAD4, FRAT1 and BCL2. Aging. 2017;9(3):932–954. | ||
Senol O, Schaaij-Visser T, Erkan E, et al. miR-200a-mediated suppression of non-muscle heavy chain IIb inhibits meningioma cell migration and tumor growth in vivo. Oncogene. 2015;34(14):1790–1798. | ||
Katar S, Baran O, Evran S, et al. Expression of miRNA-21, miRNA-107, miRNA-137 and miRNA-29b in meningioma. Clin Neurol Neurosurg. 2017;156:66–70. | ||
Su B, Zhao W, Shi B, et al. Let-7d suppresses growth, metastasis, and tumor macrophage infiltration in renal cell carcinoma by targeting COL3A1 and CCL7. Mol Cancer. 2014;13(1):206. | ||
Chang CJ, Hsu CC, Chang CH, et al. Let-7d functions as novel regulator of epithelial-mesenchymal transition and chemoresistant property in oral cancer. Oncol Rep. 2011;26(4):1003–1010. | ||
Kikuno N, Shiina H, Urakami S, et al. Knockdown of astrocyte-elevated gene-1 inhibits prostate cancer progression through upregulation of FOXO3a activity. Oncogene. 2007;26(55):7647–7655. | ||
Huang K, Li LA, Meng Y, You Y, Fu X, Song L. High expression of astrocyte elevated gene-1 (AEG-1) is associated with progression of cervical intraepithelial neoplasia and unfavorable prognosis in cervical cancer. World J Surg Oncol. 2013;11(1):297. | ||
Ying C, Li B, Xu X, et al. Lentivirus-mediated knockdown of astrocyte elevated gene-1 inhibits growth and induces apoptosis through MAPK pathways in human retinoblastoma cells. PLoS One. 2016;11(2):e0148763. | ||
Wang J, Chen X, Tong M. Knockdown of astrocyte elevated gene-1 inhibited cell growth and induced apoptosis and suppressed invasion in ovarian cancer cells. Gene. 2017;616:8–15. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.