")
Back to Journals » Drug Design, Development and Therapy » Volume 15
Lactobacillus plantarum KSFY06 Prevents Inflammatory Response and Oxidative Stress in Acute Liver Injury Induced by D-Gal/LPS in Mice
Authors Li C, Si J, Tan F, Park KY, Zhao X
Received 10 October 2020
Accepted for publication 24 December 2020
Published 6 January 2021 Volume 2021:15 Pages 37—50
DOI https://doi.org/10.2147/DDDT.S286104
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Chong Li,1– 3,* Jun Si,4,* Fang Tan,5 Kun-Young Park,1– 3 Xin Zhao1– 3
1Chongqing Collaborative Innovation Center for Functional Food, Chongqing University of Education, Chongqing 400067, People’s Republic of China; 2Chongqing Engineering Research Center of Functional Food, Chongqing University of Education, Chongqing 400067, People’s Republic of China; 3Chongqing Engineering Laboratory for Research and Development of Functional Food, Chongqing University of Education, Chongqing 400067, People’s Republic of China; 4Pre-Hospital Emergency Department, Chongqing University Central Hospital, Chongqing Emergency Medical Center, Chongqing 400014, People’s Republic of China; 5Department of Public Health, Our Lady of Fatima University, Valenzuela 838, Philippines
*These authors contributed equally to this work
Correspondence: Xin Zhao; Kun-Young Park Tel +86-23-6265-3650
Email [email protected]; [email protected]
Aim: The purpose of this study is to investigate the preventive effect of Lactobacillus plantarum KSFY06 (LP-KSFY06) on D-galactose/lipopolysaccharide (D-Gal/LPS)-induced acute liver injury (ALI) in mice.
Methods: We evaluated the antioxidant capacity of LP-KSFY06 in vitro, detailed the effects of LP-KSFY06 on the organ index, liver function index, biochemical index, cytokines, and related genes, and noted the accompanying pathological changes.
Results: The results clearly showed that LP-KSFY06 can remove 1,1-diphenyl-2-picrylhydrazyl (DPPH) and 2,2′-azino-bis (3-ethylbenzthiazoline − 6-sulphonic acid) diammonium salt (ABTS) free radicals in vitro. The analysis of the organ index and pathology demonstrated that LP-KSFY06 significantly prevented ALI. Biochemical and molecular biological analysis showed that LP-KSFY06 prevented a decrease in the antioxidant-related levels of superoxide dismutase (SOD), glutathione (GSH), glutathione peroxidase (GSH-Px), catalase (CAT), and total antioxidant capacity (T-AOC), and also prevented an increase in aspartate aminotransaminase (AST), alanine aminotransaminase (ALT), malondialdehyde (MDA), myeloperoxidase (MPO), and nitric oxide (NO) levels. LP-KSFY06 upregulated the anti-inflammatory factor interleukin (IL)-10 and downregulated the pro-inflammatory factors IL-6, IL-1β, tumor necrosis factor-α (TNF-α), and interferon-γ (IFN-γ). These oxidative and inflammatory indicators were consistent with the results of gene detections. Furthermore, we determined that LP-KSFY06 downregulated Keap1, NLRP3, apoptosis-associated speck-like protein containing a CARD (ASC), caspase-1, nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB), IL-18, and mitogen-activated protein kinase 14 (MAPK14 or p38), upregulated Nrf2, heme oxygenase-1 (HO-1), NAD(P)H dehydrogenase [quinone] 1 (NQO1), B-cell inhibitor-α (IκB-α), and thioredoxin (Trx) mRNA expression. These may be related to the regulation of the Kelch-like ECH-associated protein-1 (Keap1)-nuclear factor-erythroid-2-related factor (Nrf2)/antioxidant response element (ARE) and NLRP3/NF-κB pathways.
Conclusion: LP-KSFY06 is an effective multifunctional Lactobacillus with strong anti-oxidant and anti-inflammatory ability that can prevent D-gal/LPS-induced ALI in mice and assist in maintaining health.
Keywords: Lactobacillus plantarum KSFY06, D-galactose, lipopolysaccharide, acute liver injury, mice
Introduction
Acute liver injury (ALI) is a group of clinically common liver disease syndromes resulting from the massive death of hepatocytes. If ALI progresses to liver failure, mortality can then ensue.1 The prognosis for acute liver failure is poor because there is no effective treatment other than liver transplantation. Therefore, it is important to study the mechanisms of ALI and develop targeted interventions to increase patient survival.2,3
Oxidative stress results when the amount of reactive oxygen species (ROS) produced in the body exceeds the amount of ROS eliminated by the antioxidant protection mechanism.4 ROS can destroy unsaturated fatty acids in the phospholipids of biofilms. A series of cell peroxidation reactions result in the formation of oxidation products that further act on nucleic acids and proteins to destroy the normal functions of cells, resulting in varying degrees of oxidative damage.5 Previous research has shown that oxidative stress is involved in all aspects of bodily systems, and diseases such as allergies, asthma, Alzheimer’s, diabetes, hypertension, hyperlipidemia, coronary heart disease, and cancer are closely related to oxidative damage.6–8
In addition, a range of cytokines, such as Nrf2, NF-κB, and p38 kinase, are also activated in the process of antioxidative stress. Among them, Nrf2 is one of the core regulators of the endogenous antioxidant system in cells. The lack or activation of Nrf2 will aggravate the cytotoxicity of oxidative stress, thus inducing liver cell apoptosis and inflammatory response, which results in liver injury. Therefore, the Nrf2 signaling pathway is an important target for the treatment of oxidative stress-induced liver injury as the main pathogenesis.9 The signaling pathway of Keap1-Nrf2/ARE is considered to be a key pathway of antioxidant stress in vivo. Antioxidant enzymes and Phase II detoxification enzymes regulated in this way can remove ROS and other harmful substances, thus providing detoxification and neutralization.10
In animal models, D-galactose (D-Gal) is commonly used to induce liver damage. Ingestion of excessive D-Gal that cannot be metabolized will lead to an imbalance in cell function, changes in oxidase activity, and increased production of peroxides and oxidation products, thus causing damage to the structure and function of biomolecules that can cause systemic inflammatory response syndrome, and subsequently lead to acute liver injury and multi-organ failure.11 Lipopolysaccharide (LPS) influences a variety of biological activities and is released after bacteria grow, multiply, die, break, or lyse. By inducing nonspecific immunity in vivo, it can cause inflammatory cytokines to be synthesized and released, which will damage the host’s main systems and organs.12 Liver injury induced by simultaneous D-Gal and LPS injection has been applied to study the pathogenesis of ALI.13,14 D-Gal can increase the sensitivity and thus the lethality of LPS in mice. Although hepatocytes are primarily targeted and produce inflammatory response cells such as macrophages and neutrophils, the mechanism whereby liver damage is induced by D-Gal/LPS is still not fully understood.15
Antioxidants directly scavenge free radicals, and also increase the activity of antioxidant enzymes, reduce the accumulation of peroxides such as malondialdehyde (MDA) and lipofuscin (LP), and play a protective role.16 Studies on the antioxidant functions of Lactobacillus in vivo and in vitro, for example, Lactobacillus rhamnosus, Lactobacillus fermentans, and Lactobacillus plantarum, indicate that antioxidants are produced during the activities of lactobacillus cells and acellular extracts such as in scavenging free radicals, inhibiting lipid peroxidation, and chelating metal ions.17–19 The regulating effect of Lactobacillus on oxidative damage to the body or cells is reflected by increased expression of antioxidant genes and decreased activity of peroxides in vivo.20
A novel Lactobacillus plantarum KSFY06 (LP-KSFY06) was obtained from traditional fermented yoghurt in Kashgar, Xinjiang, China by traditional culture purification. In the current study, D-GalN/LPS-induced ALI model mice were used to study the effect of in vivo lactobacillus prophylaxis, and the mechanism of action was explained, thus providing useful information for the development of functional products and the promotion of human health.
Materials and Methods
Strain and Reagents
A new Lactobacillus (LAB) strain, Lactobacillus plantarum KSFY06 (LP-KSFY06), was registered and stored (preservation number: 15,659) at the China General Microbiological Culture Collection Center (CGMCC, Beijing, China). Lipopolysaccharides (from Escherichia coli 055:B5) were obtained from Beijing Solarbio Science & Technology Co., Ltd. (Beijing, China). D-Galactose was obtained from Sinopharma Chemical Reagent Co., Ltd. (Beijing, China).
Animals
Forty specific-pathogen-free male Kunming mice, 6–8 weeks old, were purchased from the Experimental Animal Center of Chongqing Medical University (Chongqing, China). The mice were raised in standard cages in the laboratory animal room (barrier environment) with room temperature at 20–25°C and relative humidity at 50%-60%. The lights were turned on every 12 hours for illumination, with food and water offered ad libitum.
Cultivation and Treatment of LP-KSFY06
LP-KSFY06 single colonies were inoculated into Man, Rogosa, and Sharpe (MRS, BD, Franklin Lakes, NJ, USA) media contained in liquid culture tubes, cultured at 37°C for 20 h to the initial stage of stabilization, and transferred to MRS liquid medium for 18 h according to the inoculation amount of 3% by volume. The LP-KSFY06 bacteria were collected by centrifugation (4°C, 4000 rpm, 10 min), washed twice with physiological saline, and resuspended to the adjusted cell concentrations of 2.5×109 CFU/kg·bw and 2.5×1010 CFU/kg·bw.
Antioxidant Experiments in vitro
To determine the antioxidant capability, 0.5 mL of LP-KSFY06 diluent (low concentration: 2.5×109 CFU/mL, high concentration: 2.5×1010 CFU/mL) was mixed with 2,2-diphenyl-1-picrylhydrazyl (DPPH, Sinopharm Chemical Reagent Co., Ltd., Beijing, China) working liquid (0.33 mmol/L, 2 mL). The absorbance (517 nm) was measured after reaction at room temperature in the dark for 30 minutes (BioMate 3S; Thermo Fisher Scientific, Inc., Waltham, MA, USA). The positive control was 0.2 mg/mL vitamin C (VC), and there were three parallels in each group.21
Next, 0.4 mL of LP-KSFY06 diluent (low concentration: 2.5×109 CFU/mL, high concentration: 2.5×1010 CFU/mL) was mixed with 2,2ʹ- azino-bis[3-ethylbenzothiazoline-6-sulphonic acid] (ABTS, Sinopharm Chemical Reagent Co., Ltd., Beijing, China) working liquid (1.0 mL), reacted at room temperature in the dark for 10 min, and then the absorbance (734 nm) was measured. The positive control was 0.2 mg/mL VC, and these experimental procedures were repeated three times. For the ABTS working solution preparation, reagent A consisted of 3.0 mg of ABTS mixed with 0.8 mL of double-distilled water; reagent B consisted of 1.0 mg of potassium persulfate mixed with 1.5 mL of double-distilled water; reagents A and B were mixed using 0.2 mL of each, oxidized in dark for 12 h, and diluted with anhydrous ethanol to the OD value of A734 in the range of 0.7 ± 0.02.22
Induction of ALI in Mice
The experimental mice were divided into four groups (normal, model, low dose, high dose) with ten mice/group to study the effect of LP-KSFY06 on D-Gal/LPS-induced liver injury in mice after adaptation (1 week). All groups were fed a normal diet and water for 4 weeks. The normal group received no additional treatment. The model group received D-Gal (250 mg/kg·bw) and LPS (25 mg/kg·bw) once injected intraperitoneally on the last day of the fourth week. The low- and high-dose groups received daily oral administration of LP-KSFY06 (109 CFU/kg·bw for low-dose group, 1010 CFU/kg·bw for high-dose group) from weeks two through four and one intraperitoneal injection of D-Gal (250 mg/kg·bw) and LPS (25 mg/kg·bw) on the last day of the fourth week. The volume of injection or oral administration was 0.1 mL/10 g·bw. The experimental design is shown in Figure 1.
 |
Figure 1 Experimental schedules of a D-Gal/LPS-induced hepatic injury in Kunming mice. Normal group: no additional treatment; Model group: intraperitoneal injection of D-Gal (250 mg/kg·bw)+ LPS (25 mg/kg·bw); Low-dose group: oral administration of LP-KSFY06 (109 CFU/kg·bw), intraperitoneal injection of D-Gal (250 mg/kg·bw) + LPS (25 mg/kg·bw); High-dose group: oral administration of LP-KSFY06 (1010 CFU/kg·bw), intraperitoneal injection of D-Gal (250 mg/kg·bw) + LPS (25 mg/kg·bw). Injection or oral administration: 0.1 mL/10 g·bw. Normal diet composition (mass/%): wheat flour (25%), oatmeal (25%), corn flour (25%), soybean flour (10%), fish meal (8%), hog bone powder (4%), yeast powder (2%) and refined salt (1%). |
Collection of Samples
Two hours after the last gavage, the model group and LP-KSFY06 group were euthanized by 18-hour fasting after intraperitoneal injection of D-Gal/LPS. After taking blood samples, the organs were separated and weighed. Sections (10% formalin) were made of parts of tissues, and the remaining tissues were preserved at −80°C for later experiments. This study was approved by the Ethics Committee of Chongqing Collaborative Innovation Center for Functional Food (201904045B), Chongqing, China and followed the national standard of the People’s Republic of China (GB/T 35,892–2018) laboratory animal guidelines for ethical review of animal welfare.
Histological Observations
The liver and spleen tissues (0.5 cm2) were fixed in 10% neutral formaldehyde for 48 h and then embedded in paraffin and sectioned (4 μm). Hematoxylin & eosin staining was used to observe the pathological changes under a BX43 light microscope (Olympus, Tokyo, Japan).
Determination of Biochemical Indicators in Serum and Liver
Mouse blood was placed at 4°C for 2 hours, centrifuged at 3000 rpm and 4°C for 15 minutes (BY-80C centrifugal separator, Beijing Baiyang Medical Instrument Co., Ltd., Beijing, China), and then, the upper serum was obtained and frozen at −80°C. After 0.1 g of mouse liver tissue was added to 0.9 mL of physiological saline, the tissue was homogenized (Bioprep-24; Hangzhou Aosheng Instrument Co., Ltd., Hangzhou, China) three times at a rate of 6 m/s for 30 seconds. Kits were used (Nanjing Jiancheng Institute of Bioengineering, Nanjing, Jiangsu, China) to determine the amounts of aspartate aminotransaminase (AST), alanine aminotransaminase (ALT), myeloperoxidase (MPO), MDA, nitric oxide (NO), total antioxidant capacity (T-AOC), catalase (CAT), glutathione (GSH), glutathione peroxidase (GSH-Px), and superoxide dismutase (SOD) in mouse serum and liver.
Determination of Cytokines in the Serum and Liver
The levels of cytokines tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ), and interleukin (IL)-1β, IL-6, and IL-10 in serum and liver were determined by using enzyme-linked immunosorbent assay kits (Abcam, Cambridge, MA, USA).
RT-qPCR Analysis
Liver tissue was homogenized (Bioprep-24, All for Life Science, Co., Ltd., Hangzhou, China), TRIzolTM reagent (Thermo Fisher Scientific) was used to extract the total RNA in the liver, and the concentration was diluted to 1 μg/μL. Total RNA was reverse transcribed into cDNA. Then, 1.0 μL cDNA, 2.0 μL forward and reverse primers (10 μm), and 10.0 μL premix were mixed to react under the following conditions: 3 minutes at 95°C, 30 seconds at 60°C, and 1 minute at 95°C for 40 cycles. β-Actin was used as the internal reference gene, and the data were calculated by the 2−∆∆CT method.23 The primer sequences used are shown in Table 1.
 |
Table 1 Sequences of Primers Used for RT-qPCR Analysis |
Statistical Analysis
All the experimental data were analyzed by SPSS 20.0 software (SPSS Inc, Chicago, IL, USA), combined with one-way ANOVA and Duncan’s multiple range tests; P < 0.05 was considered to be statistically significant. All the experimental data were obtained in triplicate, and the data are expressed as the mean ± standard deviation (SD).
Results
Antioxidant Effects of KSFY06 in vitro
As shown in Figure 2, compared to the VC (ascorbic acid) group (28.39% and 32.26%), the scavenging rate of KSFY06 (2.5×109 CFU/mL and 2.5×1010 CFU/mL) was 19.91%-48.49% and 38.46%-59.62% for the ABTS and DPPH free radical, respectively (p<0.05). The free radical scavenging rate of KSFY06 was significantly enhanced with increasing concentration, showing a dose-dependent relationship and satisfactory antioxidant properties.
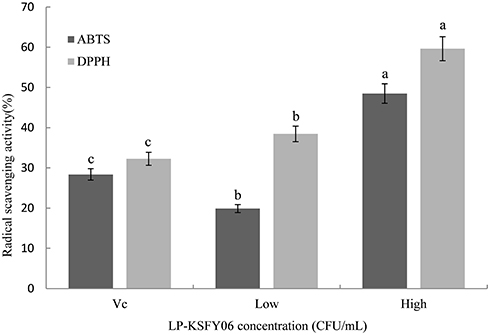 |
Figure 2 Scavenging activity of KSFY06 towards the DPPH and ABTS free radicals. KSFY06: Lactobacillus plantarum KSFY06. Low: mouse treated with 2.5×109 CFU/mL KSFY06; High: mouse treated with 2.5×1010 CFU/mL KSFY06. a-cThere was significant difference in different letters in the same column (P < 0.05), which was determined by Duncan’s multiple range test. |
Body Weight and Organ Index of Mice
The organ index indirectly reflects the change in organ function. The weights of the mice in each group were similar (Figure 3), there was no significant difference (p < 0.05), and none of them died. As shown in Table 2, compared with the normal group, the indexes of the heart, liver, spleen, lung, and kidney in the model groups were significantly increased due to the stimulation of D-Gal/LPS (p < 0.05). Compared with the model group, the organ indexes of the low-dose group and high-dose group decreased after Lactobacillus plantarum KSFY06 treatment, and the organ index of the high-dose group decreased more significantly after treatment (p < 0.05), indicating that the treatment of Lactobacillus plantarum KSFY06 decreased the organ hypertrophy of mice and also reduced the organ damage induced by D-Gal/LPS to a certain extent.24
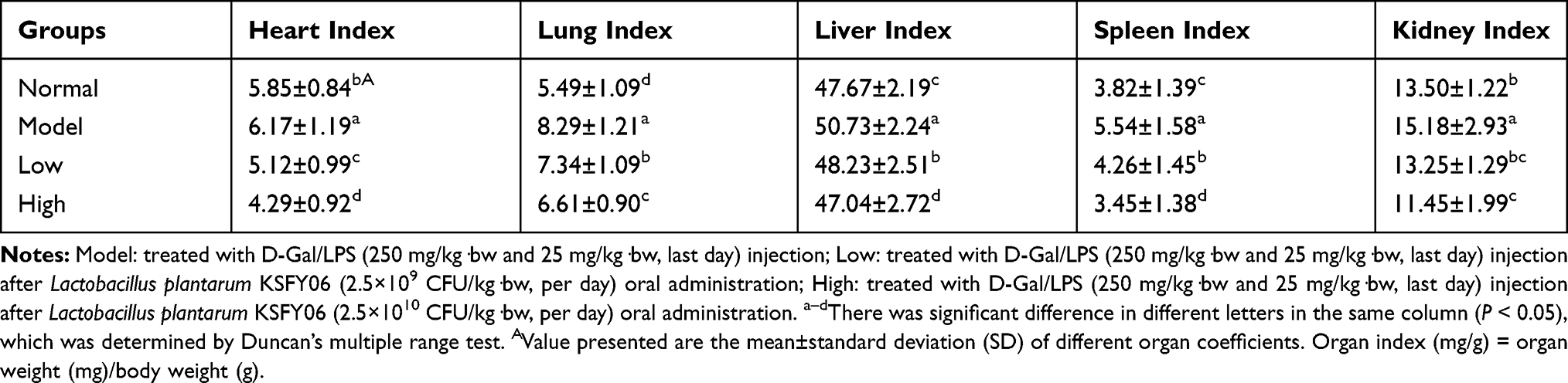 |
Table 2 Effects of Lactobacillus plantarum KSFY06 on Organ Indices in Mice with D-Gal/LPS-Induced Organ Injury (N=10/Group) |
 |
Figure 3 The changes in the weight of mice during the experiment. Low: treated with 2.5×109 CFU/mL KSFY06; High: treated with 2.5×1010 CFU/mL KSFY06. |
Histological Analyses
Harmful chemicals are metabolized in the liver, and the morphology of mouse liver tissue with injury induced by D-Gal/LPS will also change.25 As shown in Figure 4, the liver tissue of the normal group of mice was intact, the cell morphology was normal, the hepatocytes were arranged in order, and the central vein was radial. In contrast, the model group exhibited abnormal liver tissue structure, diffuse infiltration of inflammatory cells, uneven staining, irregular central veins, swollen cells, loose cytoplasm, and some cell necrosis. Compared with the model group, the liver cells of mice in the Lactobacillus plantarum KSFY06 high- and low-dose groups were neatly rearranged, the morphology was slightly altered, and the hepatocellular edema, necrosis, and inflammation were all reduced to varying degrees.
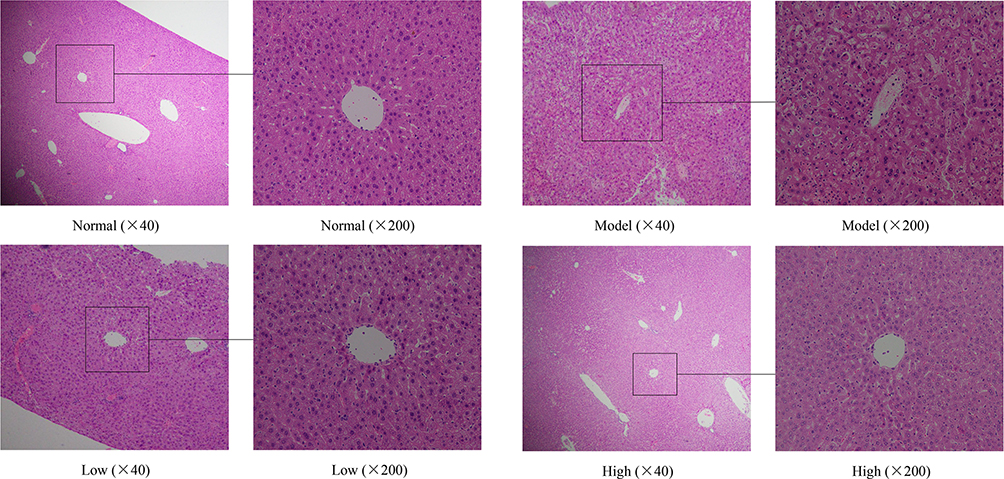 |
Figure 4 Effects of Lactobacillus plantarum KSFY06 on the liver morphology of injured mice. Model: group induced by D-Gal/LPS (250 mg/kg·bw, 25 mg/kg·bw); Low: treated with LP-KSFY06 (2.5×109 CFU/kg·bw); High: treated with LP-KSFY06 (2.5×1010 CFU/kg·bw). |
In the normal control group, the spleen cells were intact, the tissue structure was clear, the medulla cortex connection was clear, the cells were arranged in an orderly fashion, and no degeneration or necrosis was found (Figure 5). In contrast, model mice exhibited obvious spleen injury, abnormal morphology, structural disorder, enlarged splenic cord, enlarged red pulp sinus, congested spleen, increased red blood cells, decreased white pulp lymphocytes, nuclear fragmentation in white pulp, visibly narrowed red cord, and the arrangement was not dense. In the Lactobacillus plantarum KSFY06 groups, the spleen injury was significantly alleviated, the spleen was not congested, there was scattered white pulp in the splenic parenchyma, and the boundary between white pulp and red pulp was clear. Histological observations of the liver and spleen indicated that Lactobacillus plantarum KSFY06 may prevent oxidative aging-related tissue damage.
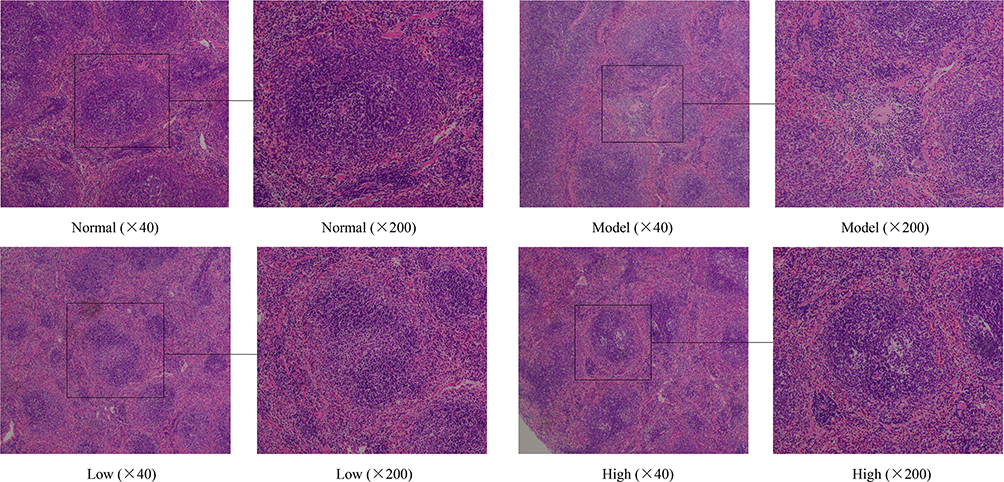 |
Figure 5 Effects of Lactobacillus plantarum KSFY06 on spleen morphology of injured mice. Model: group induced by D-Gal/LPS (250 mg/kg·bw, 25 mg/kg·bw); Low: treated with LP-KSFY06 (2.5×109 CFU/kg·bw); High: treated with LP-KSFY06 (2.5×1010 CFU/kg·bw). |
Liver Function-Related Indexes in Serum and Liver Tissue of Mice
ALT and AST are very sensitive to liver injury.26 Through detection, we found that the AST and ALT levels in serum and liver tissue of the model group significantly increased (P < 0.05), with normalization after LP-KSFY06 intervention (Table 3).
 |
Table 3 Levels of AST and ALT in Serum and Liver Tissue of Mice (N=10/Group) |
SOD, CAT, GSH-Px, GSH, T-AOC, NO, MDA, and MPO Levels in Serum and Liver of Mice
As shown in Figures 6 and 7, the SOD, CAT, GSH-Px, and T-AOC enzymatic activity and GSH content in the model group were lower than those in the normal group, while MPO activity and MDA and NO content were higher than those in the normal group (p < 0.05). Lactobacillus plantarum KSFY06 treatment effectively inhibited the decrease in serum and liver oxidation-related enzymes and complexes (SOD, CAT, GSH-Px, GSH, and T-AOC) caused by liver injury, and decreased the content of MDA, NO, and MPO, with a stronger effect being observed for the high concentration of Lactobacillus plantarum KSFY06. By fully supplementing with exogenous antioxidants (Lactobacillus plantarum KSFY06), we can increase the amount and activity of antioxidants in vivo, thus effectively inhibiting free radical oxidation damage, and subsequently preventing various aging diseases caused by oxidative damage.27
 |
Figure 6 Effects of Lactobacillus plantarum KSFY06 on SOD, CAT, GSH-Px, GSH, T-AOC, MPO, MDA, and NO in serum of mice injured by D-Gal/LPS. a–dThere was significant difference in different letters in the same column (P < 0.05), which was determined by Duncan’s multiple range test. Model: group induced by D-Gal/LPS (250 mg/kg·bw and 25 mg/kg·bw); Low: treated with LP-KSFY06 (2.5×109 CFU/kg·bw); High: treated with LP-KSFY06 (2.5×1010 CFU/kg·bw). |
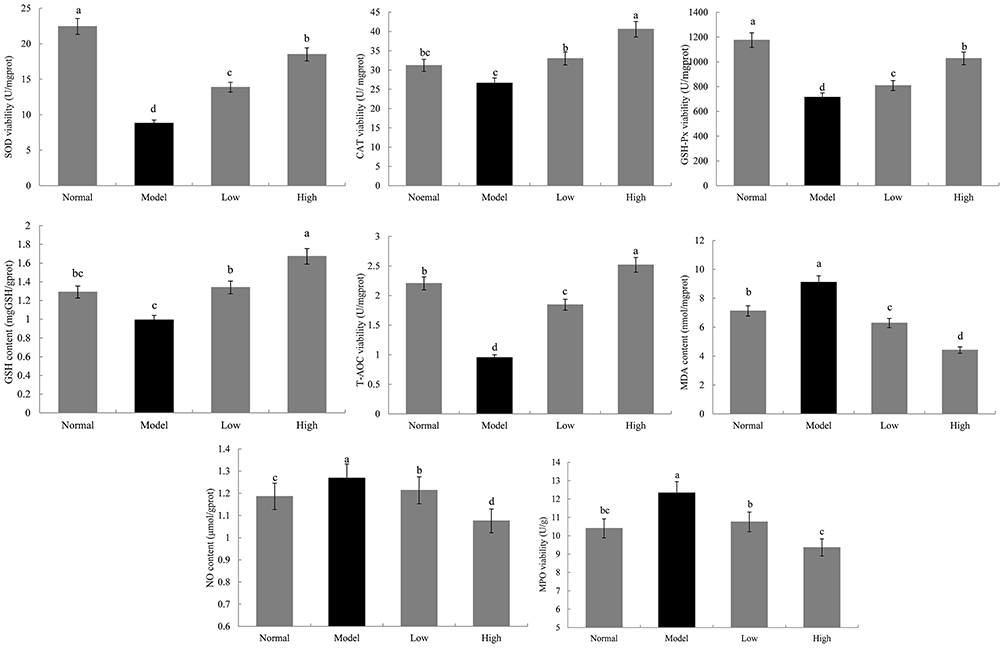 |
Figure 7 Effects of Lactobacillus plantarum KSFY06 on SOD, CAT, GSH-Px, GSH, T-AOC, MPO, MDA, and NO in liver of mice injured by D-Gal/LPS. a–dThere was significant difference in different letters in the same column (P < 0.05), which was determined by Duncan’s multiple range test. Model: group induced by D-Gal/LPS (250 mg/kg·bw and 25 mg/kg·bw); Low: treated with LP-KSFY06 (2.5×109 CFU/kg·bw); High: treated with LP-KSFY06 (2.5×1010 CFU/kg·bw). |
Cytokine Levels in Serum and Liver of Mice
As shown in Table 4, the expression of TNF-α, IL-6, IL-1β, and IFN-γ in serum and liver of the model group was higher than that of the model group, while the expression of IL-10 was lower than that of the model group (P < 0.05). In contrast, the Lactobacillus plantarum KSFY06 group ameliorated this adverse condition, and the expression of an inflammatory response in mice was decreased.
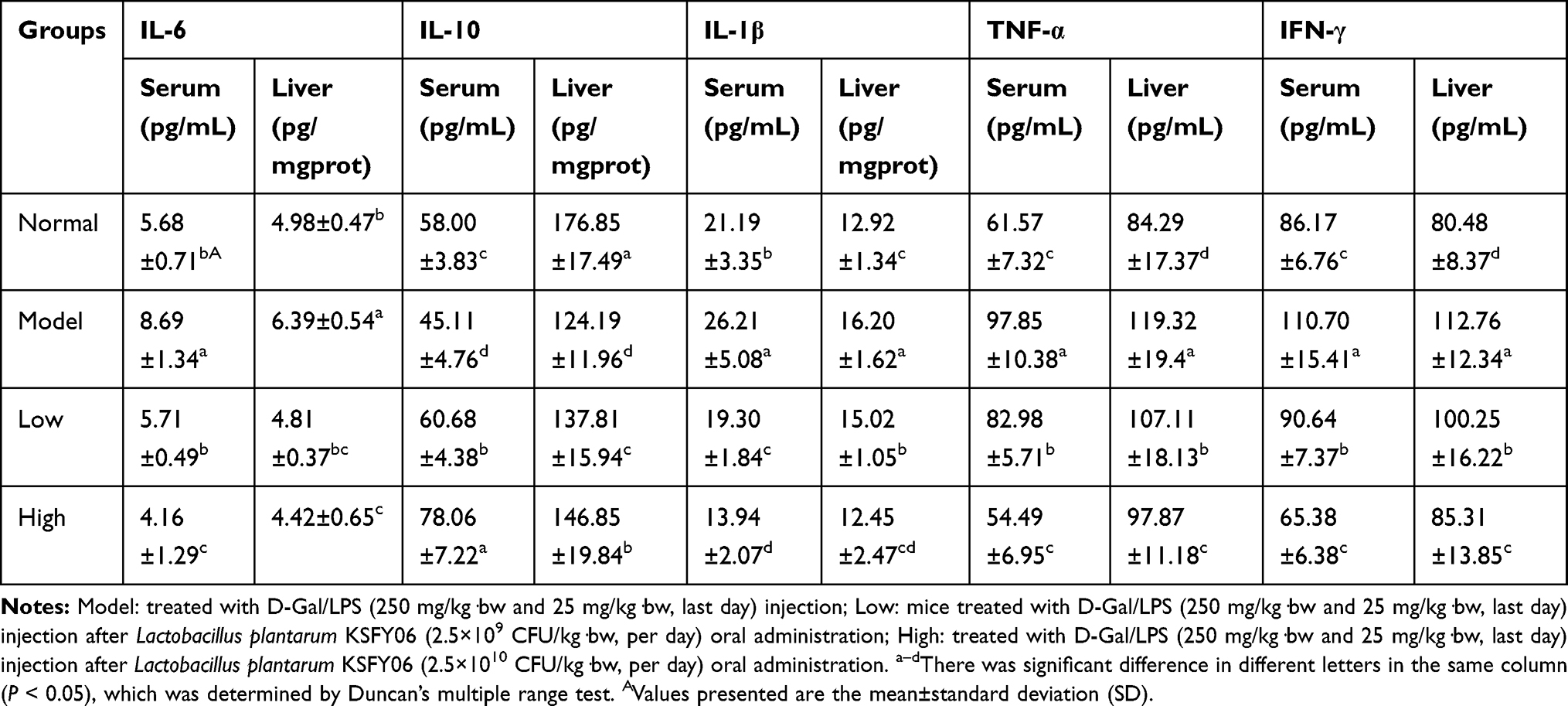 |
Table 4 Levels of IL-6, IL-10, IL-1β, TNF-α and IFN-γ in Serum and Liver Tissue of Mice (N=10/Group) |
Effects of Lactobacillus plantarum KSFY06 on the Gene Expression of Oxidation, Inflammation, and Apoptosis-Related Markers in Mouse Livers (RT-qPCR Assay)
Compared with the normal group (Figure 8), the expression levels of cuprozinc-superoxide dismutase (Cu/Zn-SOD or SOD1), manganese-superoxide dismutase (Mn-SOD or SOD2), GSH, GSH-Px, CAT, Trx, and IL-10 mRNA in the acute liver injury group (model group) decreased, while IFN-γ, IL-6, IL-18 IL-1β, TNF-α, and Mapek14 mRNA levels increased, and the expression levels of mRNA after LP-KSFY06 treatment were significantly decreased, indicating that LP-KSFY06 exerted a significant protective effect on acute liver injury caused by D-Gal/LPS (P<0.05). In addition, the expression of NLRP3, ASC, caspase-1, and NF-κB mRNA in the model group was higher than that in the normal group, while the expression of IκB-α mRNA was lower than that in the normal group. The expression of related genes (NLRP3, ASC, caspase-1, and NF-κB) in the high- and low-dose LP-KSFY06 group was significantly decreased except that the IκB-α gene was increased (P<0.05), which indicates that the intake of LP-KSFY06 may function through the NLRP3/NF-κB pathway. Furthermore, the expression of Keap1 increased, while the expression of Nrf2, HO-1, and NQO1 decreased in the model group (compared to the normal group). After treatment with LP-KSFY06, the expression of Keap1 decreased, resulting in the release and increased expression of nuclear transcription factor Nrf2, activation of downstream antioxidants, and increased expression of HO-1 and NQO1 (P<0.05), which indicates that the intake of LP-KSFY06 through the Keap1-Nrf2/ARE pathway has the effect of inhibiting the oxidative stress injury in mice.
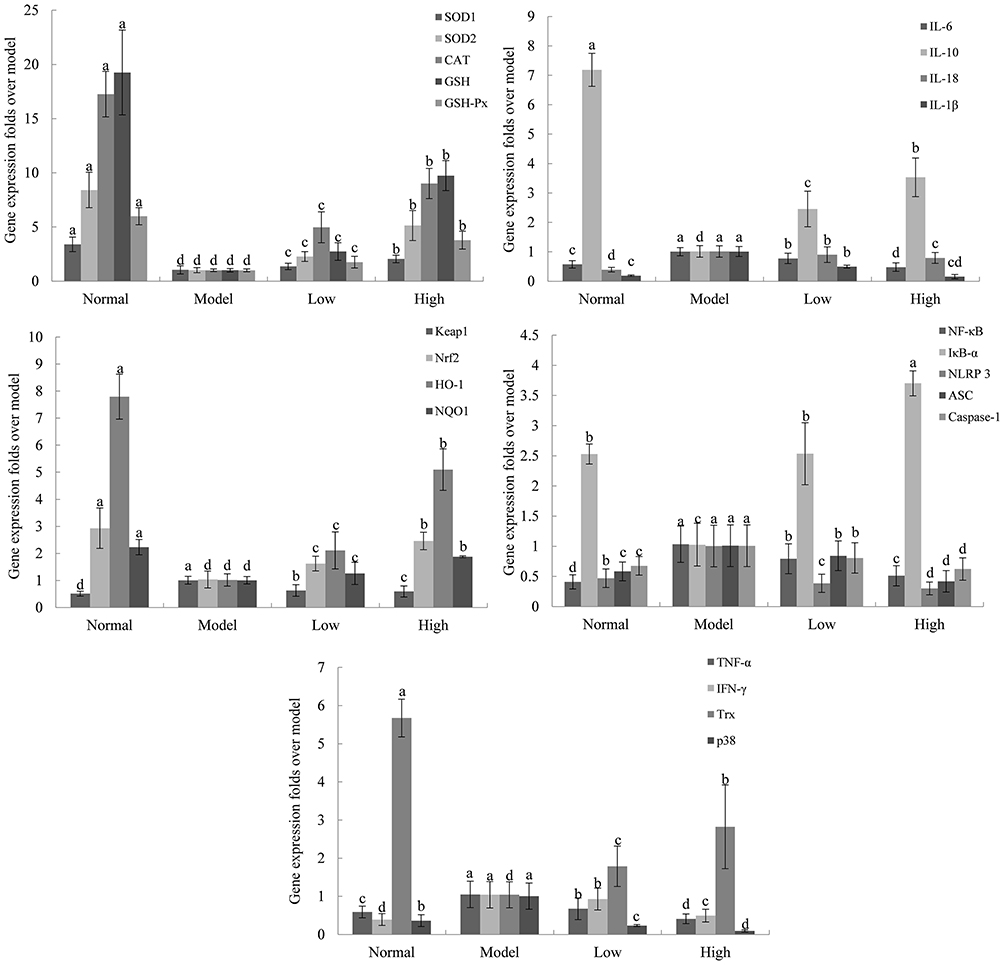 |
Figure 8 Effects of Lactobacillus plantarum KSFY06 on the mRNA expression in liver tissue of mice. a–dThere was significant difference in different letters in the same column (P < 0.05), which was determined by Duncan’s multiple range test. Model: group induced by D-Gal/LPS (250 mg/kg·bw, 25 mg/kg·bw); Low: low-dose group, 2.5×109 CFU/kg·bw; High: high-dose group, 2.5×1010 CFU/kg·bw. |
Discussion
Lactobacillus is a probiotic that is widely distributed in nature with species diversity. Some Lactobacillus strains have important biological functions such as anti-oxidation, endotoxin control, anti-aging, anti-cancer, and immunity enhancement.28–32 The Lactobacillus plantarum KSFY06 strain employed in the current study was isolated and identified from naturally fermented milk of cattle produced by the Kashi Herdsman Family in Xinjiang. This high-quality yoghurt was produced in a unique ecological environment with large temperature difference and strong ultraviolet radiation in the Kashi pastoral area, and the separated microorganism also has specific properties that are different from the general microorganism of the same type.33 Our previous experiments showed that LP-KSFY06 remained active in simulated gastric juice and a bile salt environment.
ALI is a common group of liver syndromes. Viruses, alcohol, drugs, and other factors can cause liver cell swelling, inflammation, and necrosis.34 D-Gal/LPS-induced liver injury models have been widely used in the screening of antioxidants, anti-aging drugs, plant extracts, and probiotics. The mechanism of ALI induced by this model is mainly manifested in oxidative stress and inflammatory response.35–38 Long-term injection of galactose in mice resulted in incomplete metabolic functions and accumulation in vivo, with changes in cell osmotic pressure, cell swelling, metabolic disorder, and ultimately, oxidative stress response to liver injury.39 LPS is an endotoxin that is mainly eliminated by Kupffer cells, which are activated in endotoxemia and produce ROS such as hydrogen peroxide to cause oxidative stress, and also secrete inflammatory factors such as IL and TNF-α, leading to cell necrosis.40 In our study, we found that D-Gal/LPS induced severe hepatocyte necrosis, inflammatory cell production (lymphocytes and macrophages), and disorder of liver cells around the central vein. However, after LP-KSFY06 treatment, the pathological degree of liver injury was significantly decreased, which had a positive preventive effect on liver injury.
Organ weight and organ index reflect the damage status of the body to a certain extent. LP-KSFY06 can prevent the increase in the organ index induced by D-Gal/LPS.41 ALT is the most sensitive in the early stage of ALI, and it is the main index used to diagnose the damage to the hepatocyte parenchyma, whereas AST is the main index that reflects the degree of liver injury.42 The presence of transaminase in serum and liver tissue indicated that LP-KSFY06 inhibited ALT and AST from increasing. SOD catalyzes the superoxide anion free radical dismutation to produce oxygen and hydrogen peroxide through the alternative electron gain and loss of metal ions Mn+1 (oxidation state) and Mn (reduction state), which plays a role in the oxidation balance of the body.43 CAT catalyzes the decomposition of H2O2 into H2O and O2, protects cells from H2O2, and interacts with SOD.44 GSH is a major nonenzymatic scavenger that regulates redox homeostasis in cells.45 GSH-Px can catalyze GSH to produce GSSG, and they work together to inhibit the systemic edema caused by oxidative stress, thus delaying the oxidative aging of the body.46 In our study, antioxidant enzymes (SOD, CAT, and GSH-Px) and nonenzymatic antioxidants (GSH) interacted with LP-KSFY06 to act as antioxidants.
Free radicals act on lipids to produce peroxidation reactions, and the end product of oxidation is MDA, which will cause cross-linking polymerization of proteins, nucleic acids, and other macromolecules, and has cytotoxicity, thus affecting the biochemical reactions of normal tissues.47 NO is fat soluble, rapidly diffuses through biofilm, and plays an important role in the regulation of heart and cerebral vessels.48 MPO is derived from polymorphonuclear neutrophils (PMNs), monocytes, and macrophages, which are stored in azurophilic granules. When leukocytes are activated and degranulated, MPO is released, and this occurs in response to immune defense and tissue damage in many diseases such as leukemia, vasculitis, and Alzheimer’s disease.49 When external stimulation causes oxidative stress damage (D-Gal/LPS), the metabolism of the body is disordered, and the initial balance state cannot be maintained. The lipid peroxidation products (MDA, NO, MPO) were significantly increased, and LP-KSFY06 acted as an antioxidant by reducing the content and activity of these products.
With the aggravation of liver injury, peroxides increase, and simultaneously, oxidative stress injury leads to low immune function and secretion of inflammatory cytokines. Oxidative stress injury can also act on hepatic stellate cell (HSC) and Kupffer cells, forming positive feedback regulation, leading to the increase and aggregation of cytokines, and thus promoting the development of liver injury.50 IL-1β, IL-6, IL-10, TNF-α, and IFN-γ in serum are the most important immunoregulatory cytokines in the body. The function of the immune system is related to Th1 or Th2 cells, the secretion of IFN-γ is related to Th1 cells, and the secretion of IL-6 and IL-10 is related to Th2 cells.51 TNF-α and IL-6 can act on the receptors located on the surface of hepatocytes, and they are clearly toxic to the liver and cause necrosis of a large number of hepatocytes. TNF-α is the earliest and most important mediator in the inflammation process, and IL-6 promotes the accumulation of acute proteins and T cells at the inflammation site.52,53 IL-1β is mainly expressed by innate immune cells in inflammatory injury, and its precursor intracellular processing depends on caspase-1.54 INF-γ inhibits Th2 development, and IL-10 inhibits Th1 reaction, and while these two normally regulate each other, this homeostasis is destroyed in the process of inflammation.55 In the current study, LP-KSFY06 downregulated TNF-α, IFN-γ, IL-6, and IL-1β, and upregulated IL-10, thereby reducing the expression of stress factors in injured mice.
The Keap1-Nrf2/ARE pathway is considered to be the most important endogenous antioxidant signaling pathway. Under normal physiological conditions, Nrf2 mainly binds to its inhibitor Keapl, exists in the cytoplasm in its inactive state, and is rapidly degraded under the action of the ubiquitin proteasome pathway to maintain the low transcriptional activity of Nrf2 under physiological conditions. D-Gal/LPS enhanced the expression of negative regulator Keap1 and inhibited the expression of Nrf2, HO-1, and NQO1. LP-KSFY06 ingestion causes Nrf2/ARE signal activation, lower relative expression of Keap1, release of nuclear transcription factor Nrf2, enhanced binding of Nrf2 and ARE, and further activation of downstream antioxidant enzymes and type II detoxification enzyme genes, and the body participates by regulating antioxidant activity. Nrf2 is an important transcription factor that regulates the oxidative stress response of cells, which can reduce the cell damage caused by ROS and electrophilics, maintain cellular stability, and maintain the dynamic balance of redox by inducing and regulating the expression of a series of antioxidant proteins. HO-1, as a stress protein, plays a role in hemoglobin metabolism, inflammation, and the antioxidant process, and also has a protective effect on the cardiovascular and nervous systems.56–58
NF-κB belongs to the Rel protein family, and the heterodimer composed of p65/P50 is the main biologically active form, which combines with IκB-α to form a trimer (inactive form). D-Gal/LPS can activate a series of protein kinases, such as activation of the IκB kinase complex, which leads to IκB-α ubiquitination degradation and the production of NLRP3 inflammatory bodies, and then mediates an inflammatory response. NLRP3 inflammatory corpuscles can lead to the self-shearing of the inflammatory aspartate-specific cysteine proteolytic enzyme, which initiates the host’s inflammatory response by activating proinflammatory factors such as IL-1β.59–61 Previous studies have documented that the NF-κB/NLRP3 pathway plays an inflammatory role in the process of ALI.62 We found that LP-KSFY-6 treatment enhanced the expression of IκB-α, which inhibited the expression of NF-κB, NLRP3, ASC, and caspase-1 in the NF-κB/NLRP3 pathway.
In addition, we demonstrated the anti-oxidant and anti-inflammatory effects of LP-KSFY06 in molecular biology, histology and at the gene level. We have not examined these effects at the protein level, but we plan to do so in future experiments.
Conclusion
LP-KSFY06 ameliorated the ALI induced by D-Gal/LPS, increased the activity of antioxidant enzymes in the liver and serum of injured mice, reduced the amount of peroxides, and increased the level of immunoregulatory factors. Its mechanism is related to increasing the body’s antioxidant capacity and enhancing the body’s immunoregulatory function. LP-KSFY06 activated the Nrf2/ARE signaling pathway, and inhibition of the NF-κB/NLRP3 signaling pathway plays a role.
Acknowledgments
This research was funded by Chongqing University Innovation Research Group Project (CXQTP20033), the Science and Technology Project of Chongqing Education Commission (KJQN202001604) and the Research Project of Chongqing University of Education (KY201920C), China.
Disclosure
The authors report no conflicts of interest for this work.
References
1. Hernaez R, Sola E, Moreau R, Gines P. Acute-on-chronic liver failure: an update. Gut. 2017;66(3):541–553. doi:10.1136/gutjnl-2016-312670
2. Tian Z, Chen Y, Yao N, Hu C, Wu Y, Guo D. Role of mitophagy regulation by ros in hepatic stellate cells during acute liver failure. Am J Physiol Gastrointest Liver Physiol. 2018;315(3):374–384. doi:10.1152/ajpgi.00032.2018
3. Dechene A, Sowa JP, Gieseler RK, et al. Acute liver failure is associated with elevated liver stiffness and hepatic stellate cell activation. Hepatology. 2010;52(3):1008–1016. doi:10.1002/hep.23754
4. Liguori I, Russo G, Curcio F, et al. Oxidative stress, aging, and diseases. Clin Interv Aging. 2018;13:757–772. doi:10.2147/CIA.S158513
5. Mattout A, Dechat T, Adam SA, Goldman RD, Gruenbaum Y. Nuclear lamins, diseases and aging. Curr Opin Cell Biol. 2006;18(3):335–341. doi:10.1016/j.ceb.2006.03.007
6. Crunkhorn S. Neurodegenerative disorders: restoring the balance. Nat Rev Drug Discov. 2011;10(8):576. doi:10.1038/nrd3521
7. Sas K, Robotka H, Toldi J, Vecsei L. Mitochondria, metabolic disturbances, oxidative stress and the kynurenine system, with focus on neurodegenerative disorders. J Neurol Sci. 2007;257(1–2):221–239. doi:10.1016/j.jns.2007.01.033
8. Kurien BT, Hensley K, Bachmann M, Scofield RH. Oxidatively modified autoantigens in autoimmune diseases. Free Radic Biol Med. 2006;41(4):549–556. doi:10.1016/j.freeradbiomed.2006.05.020
9. Okada K, Warabi E, Sugimoto H, et al. Nrf2 inhibits hepatic iron accumulation and counteracts oxidative stress-induced liver injury in nutritional steatohepatitis. J Gastroenterol. 2012;47(8):924–935. doi:10.1007/s00535-012-0552-9
10. Suzuki M, Otsuki A, Keleku-Lukwete N, Yamamoto M. Overview of redox regulation by Keap1–Nrf2 system in toxicology and cancer. Current Opinion Toxicol. 2016;1:29–36. doi:10.1016/j.cotox.2016.10.001
11. Dong F, Wang S, Wang Y, et al. Quercetin ameliorates learning and memory via the Nrf2-ARE signaling pathway in D-galactose-induced neurotoxicity in mice. Biochem Biophys Res Commun. 2017;491(3):636–641. doi:10.1016/j.bbrc.2017.07.151
12. Liang N, Sang Y, Liu W, Yu W, Wang X. Anti-inflammatory effects of gingerol on lipopolysaccharide-stimulated RAW 264.7 cells by inhibiting NF-kappaB signaling pathway. Inflammation. 2018;41(3):835–845. doi:10.1007/s10753-018-0737-3
13. Wang CC, Cheng PY, Peng YJ, Wu ES, Wei HP, Yen MH. Naltrexone protects against lipopolysaccharide/D-galactosamine-induced hepatitis in mice. J Pharmacol Sci. 2008;108(3):239–247. doi:10.1254/jphs.08096FP
14. Choi JW, Kim IH, Kim YM, Lee MK, Nam TJ. Pyropia yezoensis glycoprotein regulates antioxidant status and prevents hepatotoxicity in a rat model of D-galactosamine/lipopolysaccharide- induced acute liver failure. Mol Med Rep. 2016;13(4):3110–3114. doi:10.3892/mmr.2016.4932
15. Shakil A, Kramer D, Mazariegos G, Fung J, Rakela J. Acute liver failure: clinical features, outcome analysis, and applicability of prognostic criteria. Liver Transplantation. 2000;6(2):163–169. doi:10.1016/S1527-6465(00)80005-2
16. Comert ED, Gokmen V. Evolution of food antioxidants as a core topic of food science for a century. Food Res Int. 2018;105:76–93. doi:10.1016/j.foodres.2017.10.056
17. Ge Q, Ge P, Jiang D, et al. A novel and simple cell-based electrochemical biosensor for evaluating the antioxidant capacity of Lactobacillus plantarum strains isolated from Chinese dry-cured ham. Biosens Bioelectron. 2018;99:555–563. doi:10.1016/j.bios.2017.08.037
18. Marazza JA, Nazareno MA, de Giori GS, Garro MS. Enhancement of the antioxidant capacity of soymilk by fermentation with Lactobacillus rhamnosus. J Funct Foods. 2012;4(3):594–601. doi:10.1016/j.jff.2012.03.005
19. Persichetti E, De Michele A, Codini M, Traina G. Antioxidative capacity of Lactobacillus fermentum LF31 evaluated in vitro by oxygen radical absorbance capacity assay. Nutrition. 2014;30(7–8):936–938. doi:10.1016/j.nut.2013.12.009
20. Fardet A, Rock E. In vitro and in vivo antioxidant potential of milks, yoghurts, fermented milks and cheeses: a narrative review of evidence. Nutr Res Rev. 2017;31(1):52–70. doi:10.1017/S0954422417000191
21. Zhao X, Wang Q, Li G, Chen F, Qian Y, Wang R. In vitro antioxidant, anti-mutagenic, anti-cancer and anti-angiogenic effects of Chinese Bowl tea. J Funct Foods. 2014;7:590–598. doi:10.1016/j.jff.2013.12.026
22. Zhang H, Yang Y-F, Zhou Z-Q. Phenolic and flavonoid contents of mandarin (Citrus reticulata Blanco) fruit tissues and their antioxidant capacity as evaluated by DPPH and ABTS methods. J Integrative Agriculture. 2018;17(1):256–263. doi:10.1016/S2095-3119(17)61664-2
23. Suo H, Sun P, Wang C, Peng D, Zhao X. Apoptotic effects of insect tea in HepG2 human hepatoma cells. CyTA - J Food. 2015;14(2):169–175. doi:10.1080/19476337.2015.1076521
24. Khan SS, Singer BD, Vaughan DE. Molecular and physiological manifestations and measurement of aging in humans. Aging Cell. 2017;16(4):624–633. doi:10.1111/acel.12601
25. Li W, Shi J, Papa F, Maggi F, Chen X. Isofuranodiene, the main volatile constituent of wild celery (Smyrnium olusatrum L.), protects d-galactosamin/lipopolysacchride-induced liver injury in rats. Nat Prod Res. 2016;30(10):1162–1165. doi:10.1080/14786419.2015.1041139
26. Shidfar F, Bahrololumi SS, Doaei S, Mohammadzadeh A, Gholamalizadeh M, Mohammadimanesh A. The effects of extra virgin olive oil on alanine aminotransferase, aspartate aminotransferase, and ultrasonographic indices of hepatic steatosis in nonalcoholic fatty liver disease patients undergoing low calorie diet. Can J Gastroenterol Hepatol. 2018;2018:1–7. doi:10.1155/2018/1053710
27. Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002;7(9):405–410. doi:10.1016/S1360-1385(02)02312-9
28. Chiva M, Soriano G, Rochat I, Rochat F, Teresa Llovet MB. Effect of lactobacillus johnsonii la1 and antioxidants on intestinal flora and bacterial translocation in rats with experimental cirrhosis. J Hepatol. 2002;37(4):456–462. doi:10.1016/S0168-8278(02)00142-3
29. Korhonen R, Kosonen O, Korpela R. Moilanen. The expression of COX2 protein induced by Lactobacillus rhamnosus GG, endotoxin and lipoteichoic acid in T84 epithelial cells. Lett Appl Microbiol. 2004;39(1):19–24. doi:10.1111/j.1472-765X.2004.01531.x
30. Lin X, Xia Y, Wang G, et al. Lactobacillus plantarum AR501 alleviates the oxidative stress of D-Galactose-induced aging mice liver by upregulation of Nrf2-mediated antioxidant enzyme expression. J Food Sci. 2018;83(7):1990–1998. doi:10.1111/1750-3841.14200
31. Otani TT, Briley MR, Geran RI. Use of a lactobacillus casei assay as a prescreen for potential anticancer agents: an update study. J Pharm Sci. 2010;73(2):264–265. doi:10.1002/jps.2600730232
32. Gill HS, Rutherfurd KJ, Prasad J, Gopal PK. Enhancement of natural and acquired immunity by Lactobacillus rhamnosus (HN001), Lactobacillus acidophilus (HN017) and Bifidobacterium lactis (HN019). British J Nutrition. 2000;83:167–176. doi:10.1017/S0007114500000210
33. Wang Y, Zhou J, Xia X, Zhao Y, Shao W. Probiotic potential of Lactobacillus paracasei FM-LP-4 isolated from Xinjiang camel milk yoghurt. Int Dairy J. 2016;62:28–34. doi:10.1016/j.idairyj.2016.07.001
34. Khan R, Koppe S. Modern management of acute liver failure. Gastroenterol Clin North Am. 2018;47(2):313–326. doi:10.1016/j.gtc.2018.01.005
35. Li F, Miao L, Sun H, Zhang Y, Bao X, Zhang D. Establishment of a new acute-on-chronic liver failure model. Acta Pharm Sin B. 2017;7(3):326–333. doi:10.1016/j.apsb.2016.09.003
36. Liu H, Zhang W, Dong S, et al. Protective effects of sea buckthorn polysaccharide extracts against LPS/d-GalN-induced acute liver failure in mice via suppressing TLR4-NF-kappaB signaling. J Ethnopharmacol. 2015;176:69–78. doi:10.1016/j.jep.2015.10.029
37. Wang SF, Fang HY, Qu HB. Optimization of micellar electrokinetic capillary chromatography method using central composite design for the analysis of components in Yangwei granule. J Zhejiang Univ Sci B. 2011;12(3):193–200. doi:10.1631/jzus.B1000135
38. Truusalu K, Kullisaar T, Hutt P, et al. Immunological, antioxidative, and morphological response in combined treatment of ofloxacin and Lactobacillus fermentum ME-3 probiotic in Salmonella Typhimurium murine model. APMIS. 2010;118(11):864–872. doi:10.1111/j.1600-0463.2010.02672.x
39. Zhang ZF, Fan SH, Zheng YL, et al. Purple sweet potato color attenuates oxidative stress and inflammatory response induced by D-galactose in mouse liver. Food Chem Toxicol. 2009;47(2):496–501. doi:10.1016/j.fct.2008.12.005
40. Ohshima T, Mukai R, Nakahara N, et al. HTLV-1 basic leucine-zipper factor, HBZ, interacts with MafB and suppresses transcription through a Maf recognition element. J Cell Biochem. 2010;111(1):187–194. doi:10.1002/jcb.22687
41. Tang T, He B. Treatment of D-galactose induced mouse aging with Lycium barbarum polysaccharides and its mechanism study. Afr J Tradit Complement Altern Med. 2013;10(4):12–17. doi:10.4314/ajtcam.v10i4.3
42. Josekutty J, Iqbal J, Iwawaki T, Kohno K, Hussain MM. Microsomal triglyceride transfer protein inhibition induces endoplasmic reticulum stress and increases gene transcription via Ire1alpha/cJun to enhance plasma ALT/AST. J Biol Chem. 2013;288(20):14372–14383. doi:10.1074/jbc.M113.459602
43. McCord JM, Edeas MA. SOD, oxidative stress and human pathologies: a brief history and a future vision. Biomed Pharmacother. 2005;59(4):139–142. doi:10.1016/j.biopha.2005.03.005
44. Sepasi Tehrani H, Moosavi-Movahedi AA. Catalase and its mysteries. Prog Biophys Mol Biol. 2018;140:5–12. doi:10.1016/j.pbiomolbio.2018.03.001
45. Wilke BC, Vidailhet M, Favierb A, et al. Selenium, glutathione peroxidase (GSH-Px) and lipid peroxidation products before and after selenium supplementation. Clinica Chimica Acta. 1992;207:137–142. doi:10.1016/0009-8981(92)90157-L
46. Li JE, Nie SP, Xie MY, Huang DF, Wang YT, Li C. Chemical composition and antioxidant activities in immunosuppressed mice of polysaccharides isolated from Mosla chinensis Maxim cv. jiangxiangru. Int Immunopharmacol. 2013;17(2):267–274. doi:10.1016/j.intimp.2013.05.033
47. Draper HH, Hadley M. Malondialdehyde Determination as Index of Lipid Peroxidation. Methods Enzymol. 1990;186:421–431. doi:10.1016/0076-6879(90)86135-I
48. Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev. 2007;87(1):315–424. doi:10.1152/physrev.00029.2006
49. Kizaki M, Miller CW, Selsted ME, Koeffler HP. Myeloperoxidase (MPO) gene mutation in hereditary MP0 deficiency. Blood. 1994;83(7):1935–1940. doi:10.1007/BF01834369
50. Xu C, Bailly-Maitre B, Reed JC. Endoplasmic reticulum stress: cell life and death decisions. J Clin Invest. 2005;115(10):2656–2664. doi:10.1172/JCI26373
51. Liu Y, Jiao F, Qiu Y, et al. The effect of Gd@C82(OH)22 nanoparticles on the release of Th1/Th2 cytokines and induction of TNF-alpha mediated cellular immunity. Biomaterials. 2009;30(23–24):3934–3945. doi:10.1172/JCI26373
52. Chen J, Liu GZ, Sun Q, et al. Protective effects of ginsenoside Rg3 on TNF-alpha-induced human nucleus pulposus cells through inhibiting NF-kappaB signaling pathway. Life Sci. 2019;216:1–9. doi:10.1016/j.lfs.2018.11.022
53. Taher MY, Davies DM, Maher J. The role of the interleukin (IL)-6/IL-6 receptor axis in cancer. Biochem Soc Trans. 2018;46(6):1449–1462. doi:10.1042/BST20180136
54. Swaroop S, Mahadevan A, Shankar SK, Adlakha YK, Basu A. Correction to: HSP60 critically regulates endogenous IL-1beta production in activated microglia by stimulating NLRP3 inflammasome pathway. J Neuroinflammation. 2018;15(1):317. doi:10.1186/s12974-018-1355-6
55. Brodacki B, Staszewski J, Toczylowska B, et al. Serum interleukin (IL-2, IL-10, IL-6, IL-4), TNFalpha, and INFgamma concentrations are elevated in patients with atypical and idiopathic parkinsonism. Neurosci Lett. 2008;441(2):158–162. doi:10.1016/j.neulet.2008.06.040
56. De Freitas Silva M, Pruccoli L, Morroni F, et al. The Keap1/Nrf2-ARE pathway as a pharmacological target for chalcones. Molecules. 2018;23(7):1803–1825. doi:10.3390/molecules23071803
57. Bellezza I, Giambanco I, Minelli A. Donato R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim Biophys Acta Mol Cell Res. 2018;1865(5):721–733. doi:10.1016/j.bbamcr.2018.02.010
58. Zhang J, Fu B, Zhang X, et al. Bicyclol upregulates transcription factor Nrf2, HO-1 expression and protects rat brains against focal ischemia. Brain Res Bull. 2014;100:38–43. doi:10.1016/j.brainresbull.2013.11.001
59. Liu RT, Wang A, To E, et al. Vinpocetine inhibits amyloid-beta induced activation of NF-kappaB, NLRP3 inflammasome and cytokine production in retinal pigment epithelial cells. Exp Eye Res. 2014;127:49–58. doi:10.1016/j.exer.2014.07.003
60. Sun Y, Zhao Y, Yao J, et al. Wogonoside protects against dextran sulfate sodium-induced experimental colitis in mice by inhibiting NF-kappaB and NLRP3 inflammasome activation. Biochem Pharmacol. 2015;94(2):142–154. doi:10.1016/j.bcp.2015.02.002
61. Long L, Pang XX, Lei F, et al. SLC52A3 expression is activated by NF-kappaB p65/Rel-B and serves as a prognostic biomarker in esophageal cancer. Cell Mol Life Sci. 2018;75(14):2643–2661. doi:10.1007/s00018-018-2757-4
62. Jia KK, Ding H, Yu HW, et al. Huanglian-Wendan decoction inhibits NF-kappaB/NLRP3 inflammasome activation in liver and brain of rats exposed to chronic unpredictable mild stress. Mediators Inflamm. 2018;2018:1–15. doi:10.1155/2018/3093516
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.