")
Back to Journals » International Journal of Nanomedicine » Volume 13
Increased viability of fibroblasts when pretreated with ceria nanoparticles during serum deprivation
Authors Genier FS, Bizanek M, Webster TJ , Roy AK
Received 5 August 2017
Accepted for publication 9 October 2017
Published 9 February 2018 Volume 2018:13 Pages 895—901
DOI https://doi.org/10.2147/IJN.S148390
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Carlos Rinaldi
Francielli S Genier, 1 Maximilian Bizanek, 1 Thomas J Webster, 1,2 Amit K Roy 1,2
1Department of Chemical Engineering, Northeastern University, Boston, MA, USA; 2Wenzhou Institute of Biomaterials and Engineering (WIBE), Wenzhou University, Wenzhou, People’s Republic of China
Abstract: Conditions of cellular stress are often the cause of cell death or dysfunction. Sustained cell stress can lead to several health complications, such as extensive inflammatory responses, tumor growth, and necrosis. To prevent disease and protect human tissue during these conditions and to avoid medication side effects, nanomaterials with unique characteristics have been applied to biological systems. This paper introduces the pretreatment in human dermal fibroblasts with cerium oxide nanoparticles during nutritional stress. For this purpose, human dermal fibroblast cells received cell culture media with concentrations of 250 µg/mL and 500 µg/mL of nano-cerium oxide before being exposed to 24, 48, and 72 hours of serum starvation. Contrast images demonstrated higher cell confluence and cell integrity in cells pretreated with ceria nanoparticles compared to untreated cells. It was confirmed by MTS assay after 72 hours of serum starvation that higher cell viability was achieved with ceria nanoparticles. The results demonstrate the potential of cerium oxide nanoparticles as protective agents during cellular starvation.
Keywords: cerium oxide, nanoparticles, serum starvation, human dermal fibroblasts
Corrigendum for this paper has been published
Introduction
Materials and devices with at least one dimension at the nanoscale (under 1,000 nm) present unique characteristics when compared with their equivalents at the micron scale. The reason is the presence of a high surface area per unit mass of material, which increases material reactivity and electrical characteristics significantly. Hence, nanotechnology, the development and use of nanomaterials, has been a widely studied field that encompasses a multidisciplinary range of applications. Especially in the field of nanomedicine, the engineering of biomaterials has shown considerable progress due to improvements in the synthesis and characterization of nanoparticles for their use in therapeutic purposes, such as magnetic biomarkers, drug delivery systems, and medical device coatings.1
Among several other nanomaterials, cerium composts have been reported as powerful antioxidants with high biological activity and relatively low cell toxicity. Although considered a rare earth element, cerium is one of the most abundant materials of this kind, comparable to copper and cobalt, which enables its application.2 It has demonstrated no toxicity at 25 μg/mL, as shown by Xia et al,3 who also verified that cerium compounds induced considerable increase on bronchial cell resistance during oxidative conditions.3 Often combined with oxygen to create cerium oxide, this element can be used in nanoparticulate forms due to its ability to participate in redox reactions. This is a result of its interchangeable valence state between Ce3+ and Ce4+, which creates oxygen vacancies in the crystalline structure. These spots increase ceria nanoparticle (Ce NP) reactivity, contributing as reactive oxygen species scavengers for a wide range of applications.4
Reactive oxygen species are frequently formed during oxidative stress situations, such as inflammation responses. Hence, it is possible to define oxidative stress as the disproportion between the production of these species, such as hydrogen peroxide and superoxide, and their elimination through antioxidant enzymes that are present on human tissues, such as catalase and superoxide dismutase.5 This imbalance often results in the destruction or dysfunction of cellular structures, causing pathological disorders. The prevention of these pathophysiological conditions is a main goal in the field of nanomedicine. For this reason, Ce NPs have been leading the way with their capacity to imitate superoxide dismutase and catalase activity, thus protecting living cells from oxidative compounds and promoting tissue resistance.6 In addition, Ce NPs have been reported as capable of healing dermal wounds in a mouse model by reducing the oxidative stress in the damaged region, enhancing the regenerative process.7
This paper presents the results of Ce NP pretreatment on human dermal fibroblasts before their exposure to induced nutrition stress, which can naturally occur in pathologic conditions such as infections and neurodegenerative diseases.8 Two concentrations of Ce NP were used: 250 μg/mL and 500 μg/mL. For the starvation stress, three concentrations of fetal bovine serum (FBS) media were tested (10%, 0.2%, and 0.1%) for 24, 48, and 72 hours. Contrast images and MTS assays were used to verify the protective effect of nano-ceria in these stress conditions.
Methods
Ceria NPs preparation
All chemicals were procured from Sigma (St Louis MO, USA) with no further purification. The Ce NPs used were synthesized by mixing a cerium nitrate hexahydrate solution and a sodium hydroxide solution for an hour and drying it for 24 hours, as described earlier.9 The stock solutions of cerium and sodium hydroxide were prepared separately by dissolving and mixing 21.7 g of cerium nitrate hexahydrate in 100 mL of water and by adding 2 g of sodium hydroxide to 50 mL of distilled water in beaker. In another beaker, 85 mL of ceria nitrate, 5 mL of sodium hydroxide, and 75 mL of distilled water were added. The solution was stirred for 1 hour and heated at 150°C for 24 hours.
Sizing
Samples of cerium oxide nanoparticles were assessed for size by transmission electron microscopy (TEM). In order to prepare the sample for the TEM examination, a suspension was prepared in ethanol. The suspension was then placed on a carbon grid for further analysis. A JEOL JEM-101 TEM was used to characterize the average size, and dynamic light scattering (Zetasizer, NanoZS, Malvern Instruments, Malvern, UK) experiments were performed to analyze the particle size and distribution of Ce NPs.
Cell culture
Neonatal human dermal fibroblasts (HDF) (ATTC, Manassas, VA, USA, Cat # PCS-201-010) were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco, Carlsbad, CA, USA) with high glucose and L-glutamine, supplemented with 10% heat-inactivated FBS (from Gibco), 100 U/mL penicillin G, 100 mg/mL streptomycin (Gibco), and 250 μg/mL amphotericin B and were grown to confluency on 100 mm plates. The cells were maintained at 37°C in a humidified chamber at 5% CO2 and 95% air atmosphere. Fibroblasts were used at passages 6–8 in every experiment, and the culture medium was changed every 2 days. The growth characteristics and light microscopic appearance of the cells were unchanged up to passage 10. When the cultures reached 90% confluence, the cells were then detached from the plates with 500 μL of TripleE (Gibco). Next, the single cell suspension was placed in a 15 mL screw cap tube and centrifuged. The cell pellet was reconstituted in a FBS-supplemented DMEM complete medium. Cell morphology was visualized under a microscope.
Serum starvation
HDF cells were seeded at a uniform concentration of 200,000 cells/well for 48 hours on six-well plates in complete media containing DMEM with high glucose and L-glutamine supplemented with 10% heat-inactivated FBS, 100 U/mL penicillin G, 100 mg/mL streptomycin, 250 μg/mL amphotericin B.
For the pretreatment experiment, the old medium was replaced with the complete medium containing 250 μg/mL and 500 μg/mL of Ce NPs in respective wells. After 24 hours of incubation, the serum starvation process was initiated by adding low serum medium (0.2% and 0.1% FBS in the same DMEM, 0.1% penicillin/streptomycin, and 250 μg/mL amphotericin B) to the respective wells. The plates containing (10%, 0.2%, and 0.1%) media were incubated for 24, 48, and 72 hours at 37°C in a humidified chamber at 5% CO2 and 95% air atmosphere.
Contrast images
Images of six-well plates were obtained through phase contrast microscopy 0, 24, 48, and 72 hours after the stress had been initiated. A Nikon (Eclipse TS100, Nikon Instruments Inc, Melville, NY, USA) microscope was used for acquiring phase contrast images. For each well, four images were acquired in the central region of the sample to avoid edge effects. All images were used to evaluate the cell morphology, although only one for each sample is showed in this report.
Cell viability assay
HDF cells at 5,000 cells/well seeded on 96-well plates went through the same incubation process described in the section “Serum Starvation”. Then, after 72 hours of stress, the MTS Cell Proliferation Assay was applied and the plates were read by a spectrometer to determine the dependence of HDF cell viability on the level of starvation. The viability following various treatments was determined by normalizing the treated to untreated cells. HDF test cells were then pretreated with nanoparticles for 24 hours followed by 72 hours of stress. Hence, the results were collected without any time to recovery, observing mainly the protective effect of Ce NP pretreatment during stressed conditions.
Statistical analysis
All experiments were conducted three times, and differences between means were determined using analysis of variance followed by post hoc Tukey’s test. Figure 1 summarizes the experimental paradigm.
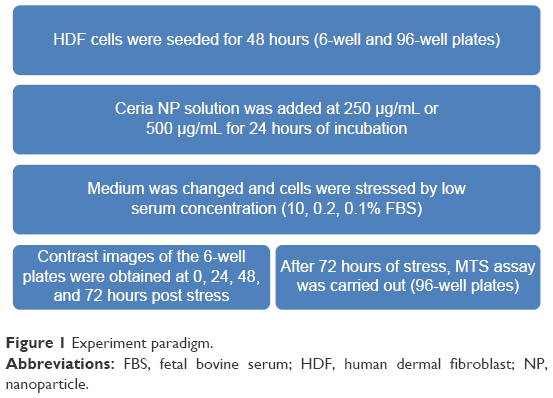 | Figure 1 Experiment paradigm. |
Results
Samples of Ce NP were characterized through TEM. Figure 2 shows a TEM image in which it is possible to observe the diameter of four nanoparticles, which were 8.34 nm, 9.44 nm, 9.74 nm, and 10.2 nm.
 | Figure 2 TEM image of CeriaNPs. |
Contrast images of HDF were obtained after 0, 24, 48, and 72 hours after the serum starvation was initiated. At 0 hour of stress, cells were of similar confluence for all treatments. After 24 hours, the wells with different serum concentrations (10%, 0.2%, and 0.1%) showed significant differences in cell population. Figure 3 presents images of the control (10% FBS), including wells with no Ce NPs pretreatment, as well as the ones pretreated with concentrations of 250 μg/mL and 500 μg/mL.
 | Figure 3 HDF cells exposed to ceria NP (250 and 500 μg/mL for 24, 48, and 72 hours). |
Phase contrast images of the serum-starved cells at 0.2% FBS were also obtained. Figure 4 shows these images such that each column represents a different concentration of Ce NP in the pretreatment (no ceria NP, 250 μg/mL, and 500 μg/mL), and each row represents a different period of stress (24, 48, and 72 hours).
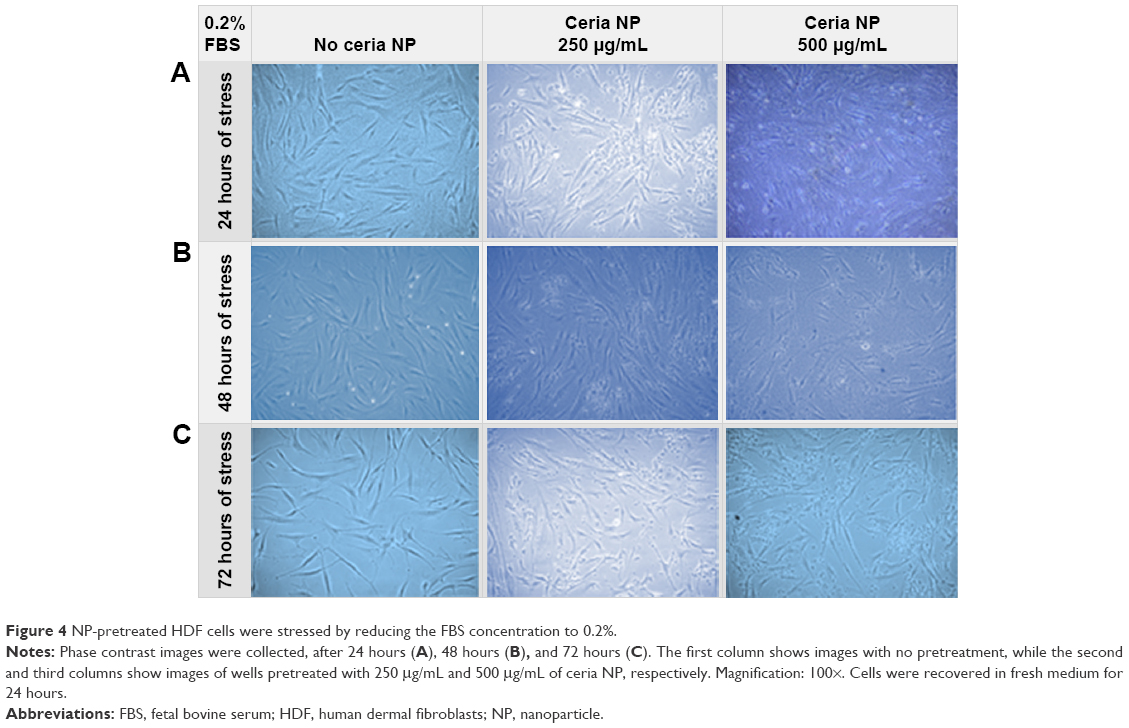 | Figure 4 NP-pretreated HDF cells were stressed by reducing the FBS concentration to 0.2%. |
Similarly, Figure 5 presents contrast images of cells under serum starvation at the level of 0.1% FBS. Each column represents a different concentration of Ce NP (no ceria NP, 250 μg/mL, and 500 μg/mL) during pretreatment, and each row represents a different period of stress (24, 48, and 72 hours).
 | Figure 5 NP-pretreated HDF cells were stressed by reducing the FBS concentration to 0.1%. |
An MTS assay was carried out for HDF cells under 72 hours of serum starvation in order to evaluate quantitatively the effect of pretreatment with Ce NP at concentrations of 0 μg/mL, 250 μg/mL, and 500 μg/mL. Figure 6 presents the results in terms of cell viability for the control (cells in an environment with 10% FBS) and for serum concentrations of 0.2% and 0.1%.
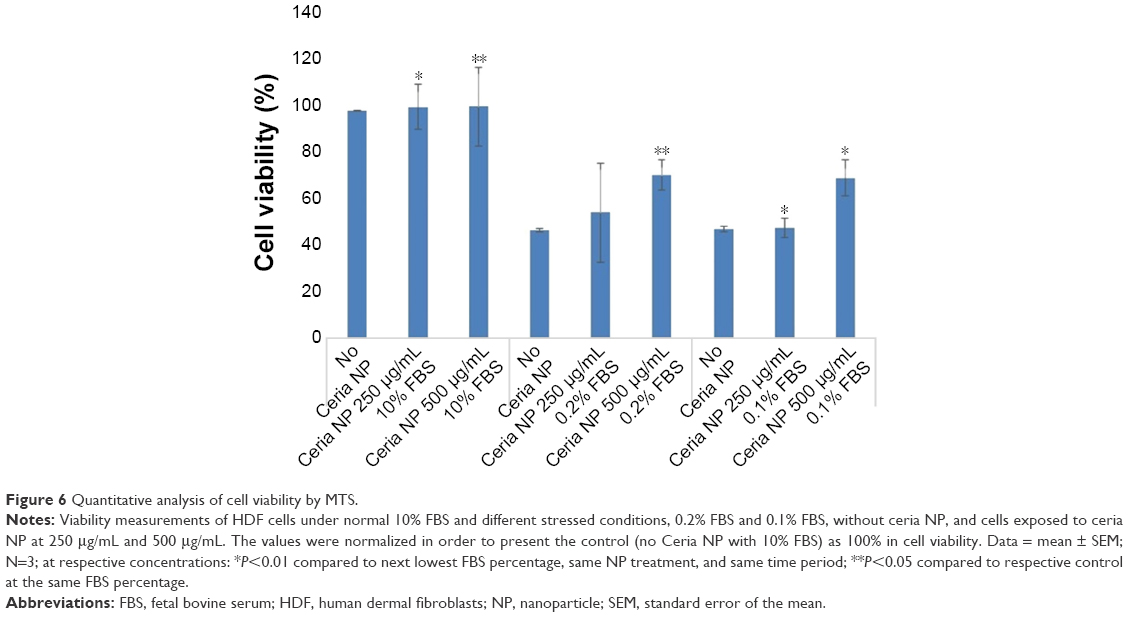 | Figure 6 Quantitative analysis of cell viability by MTS. |
After 72 hours of serum starvation stress at a concentration of 0.2% FBS, MTS assays were carried out in order to evaluate the effect of pretreatment with Ce NPs at 250 μg/mL and 500 μg/mL, as well as non-pretreated cells. Results for all three concentrations of Ce NPs used (0 μg/mL, 250 μg/mL, and 500 μg/mL) during pretreatment are presented. As the serum concentration reduced to 0.1%, there was no significant change in cell number from no ceria to 250 μg/mL ceria, which could mean that under this condition, the concentration is too low to induce any effect. However, there was a 50% increase in cell population at 500 μg/mL.
Discussion
Human cells are often exposed to environmental stress capable of stimulating mutations and cell death. For this reason, cell defense mechanisms can be activated to preserve cellular integrity.10 These mechanisms frequently include the expression of specific genes responsible for cell adaptive response, such as the hypoxia-inducible factors, which are activated as a transcriptional response to low concentrations of oxygen.11 The sources of stress can vary: temperature, nutrient deficiency, oxidative stress, ionizing radiation, osmotic shock, and so on. In vitro, artificial stress can be used to test materials and protocols that could stimulate or suppress (in cancer cells, for instance) survival responses.12 In this study, nutritional starvation was used in order to test CeO2 nanoparticles’ protective effects. Concentrations below 10% in FBS were used, precisely 0.2% and 0.1%. As demonstrated, extremely low concentrations of serum can compromise cell proliferation and multipotentiality due to nutrient deprivation.13
During this study, there were no significant differences between pretreated cells and non-pretreated cells in the control wells, as shown on Figure 3. Up until 72 hours of incubation, the level of confluence remained very similar, as determined by MTS assays. This demonstrates that exposure to nano-ceria at concentrations of 250 μg/mL and 500 μg/mL does not result in any substantial negative effect on cell growth.
In the serum starvation experiments, the cells that received pretreatment with Ce NPs at concentrations of 250 μg/mL and 500 μg/mL presented more confluence when compared to the non-pretreated cells, as demonstrated on Figures 4 and 5. This behavior was observed in both starving concentrations of FBS (0.2% in FBS and 0.1% in FBS). The untreated cells also showed an elongated phenotype after 72 hours of stress in an effort to increase their superficial area to absorb the scarce amount of serum in the media. This configuration was less observed on cells submitted to pretreatment. The MTS results (Figure 6) indicated that nano-ceria behaved as a protective material in stress conditions. At a concentration of 0.2% in FBS, pretreatment of 250 μg/mL Ce NPs resulted in 16.3% more cell viability, while pretreatment at 500 μg/mL resulted in 51.1% more. However, the statistical analysis indicated no significant difference between the control and the pretreated cells at 250 μg/mL Ce NPs. At concentrations of 0.1% in FBS, pretreated cells with nano-ceria at 250 μg/mL presented cell viability 2.9% higher than the untreated cells, and 47.4% higher on cells pretreated with Ce NPs at 500 μg/mL. The low cell viability between pretreated cells with Ce NP at 250 μg/mL and untreated cells may be an indication that this concentration of nano-ceria is not as effective as 500 μg/mL.
As demonstrated by Xu et al,14 in case of acute starvation conditions, autophagy can be activated to offer energy and nutrients as an adaptive response. This process is regulated mostly by Bcl-2 family proteins, and its inhibition is responsible for reduced cell viability.14 Hence, the protective effect of nano-CeO2 can be a reflection of its stimulation of Bcl-2 expression, which would explain higher cell viability on pretreated HDF cells compared to non-pretreated ones. Another cause could be an increased expression of p8 protein, highly activated during stress conditions such as starvation.15 In this case, the nanoparticles could be acting on the Nupri1 gene, which encodes p8 protein in order to prevent cell death.16 Therefore, the presented results demonstrate that Ce NP pretreatment prevented cell death and increased cell viability upon starvation stress. This indicates that nano-CeO2 might stimulate cell survivor mechanisms, such as higher expression of p8 and Bcl-2 proteins. Further molecular studies, such as RNA analysis, must be conducted in order to confirm this hypothesis. Since the applications of CeO2 nanoparticles could lead to their distribution in the body through the bloodstream, cytotoxicity assays using other mammalian tissues would be useful to broadly evaluate the effects of nano-ceria in the human body. Moreover, a larger difference between the serum concentrations could be helpful to determine the lowest nutritional level in which HDF cells could survive under Ce NPs protection.
Conclusion
In summary, we have shown for the first time that Ce NPs protect HDF cells during serum starvation. Specifically, a 24-hour pretreatment with Ce NP at concentrations of 250 and 500 μg/mL resulted in increased cell density under stressful conditions. Cell viability experiments with MTS assays indicate that pretreatment with nano-ceria increased the cell population by 51.1% after starvation. This indicates that nano-ceria somehow abrogates the cellular stress condition by interfering with reactive oxygen species generation. A lot still must be done in order to delineate the molecular mechanism to fully understand the role of nano-ceria in attenuating the cellular death under stressful conditions. Hence, the present study demonstrates the potential of Ce NP to prevent cell death in stress conditions.
Acknowledgment
We would like to thank Northeastern University for funding this research.
Disclosure
The authors report no conflicts of interest in this work.
References
Shcherbakov AB, Zholobak NM, Baranchikov AE, Ryabova AV, Ivanov VK. Cerium fluoride nanoparticles protect cells against oxidative stress. Mat Sci Eng C. 2015;50:151–159. | ||
Castano CE, O’Keefe MJ, Fahrenholtz WG. Cerium-based oxide coatings. Curr Opin Solid State Mater Sci. 2015;19(2):69–76. | ||
Xia T, Kovochich M, Liong M, et al. Comparison of the mechanism of toxicity of zinc oxide and cerium oxide nanoparticles based on dissolution and oxidative stress properties. ACS Nano. 2008;2(12):2121–2134. | ||
Montfort C, Alili L, Teuber-Hanselmann S, Brenneisen P. Redox-active cerium oxide nanoparticles protect human dermal fibroblasts from PQ-induced damage. Redox Biol. 2015;4:1–5. | ||
Weaver JD, Stabler CL. Antioxidant cerium oxide nanoparticle hydrogels for cellular encapsulation. Acta Biomater. 2015;16:136–144. | ||
Pirmohamed T, Dowding J, Singh S, et al. Nanoceria exhibit redox state-dependent catalase mimetic activity. Chem Commun. 2010;46(16):2736–2738. | ||
Chigurupati S, Mughal MR, Okun E, et al. Effects of cerium oxide nanoparticles on the growth of keratinocytes, fibroblasts and vascular endothelial cells in cutaneous wound healing. Biomaterials. 2013;34(9):2194–2201. | ||
Levine B, Kroemer G. Autophagy in the Pathogenesis of Disease. Cell. 2008;132(1):27–42. | ||
Webster T, Roy A, Yuan B. Cytoprotective effects of cerium and selenium nanoparticles on heat-shocked human dermal fibroblasts: an in vitro evaluation. Int J Nanomedicine. 2016;11:1427. | ||
Leong MLL, Maiyar AC, Kim B, O’keeffe BA, Firestone GL. Expression of the serum- and glucocorticoid-inducible protein kinase, Sgk, is a cell survival response to multiple types of environmental stress stimuli in mammary epithelial cells. J Biol Chem. 2002;278(8):5871–5882. | ||
Majmundar AJ, Waihay JW, Simon MC. Hypoxia-inducible factors and the response to hypoxic stress. Mol Cell. 2010;40(2):294–309. | ||
Spriggs KA, Bushell M, Willis AE. Translational regulation of gene expression during conditions of cell stress. Mol Cell. 2010;40(2):228–237. | ||
Vunjak-Novakovic G, Freshney RI. Culture of Cells for Tissue Engineering. 1st ed. Hoboken: Wiley-Liss; 2006. | ||
Xu H, Wu D, Gu J, et al. The pro-survival role of autophagy depends on Bcl-2 under nutrition stress conditions. PLoS One. 2013;8(5):e63232. | ||
Goruppi S, Iovanna JL. Stress-inducible protein P8 is involved in several physiological and pathological processes. J Biol Chem. 2009;285(3):1577–1581. | ||
Cano CE, Iovanna JL. Stress proteins and pancreatic cancer metastasis. Sci World J. 2010;10:1958–1966. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.