")
Back to Journals » Infection and Drug Resistance » Volume 15
In vitro Antimicrobial Activity and the Mechanism of Berberine Against Methicillin-Resistant Staphylococcus aureus Isolated from Bloodstream Infection Patients
Authors Xia S, Ma L, Wang G, Yang J, Zhang M, Wang X, Su J, Xie M
Received 23 January 2022
Accepted for publication 8 April 2022
Published 19 April 2022 Volume 2022:15 Pages 1933—1944
DOI https://doi.org/10.2147/IDR.S357077
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Shuai Xia,1,* Liyan Ma,2,* Guoxing Wang,1 Jie Yang,1 Meiying Zhang,3 Xuechen Wang,1 Jianrong Su,2 Miaorong Xie1
1Department of Emergency and Critical Care Center, Beijing Friendship Hospital, Capital Medical University, Beijing, 100050, People’s Republic of China; 2Department of Clinical Laboratory, Beijing Friendship Hospital, Capital Medical University, Beijing, 100050, People’s Republic of China; 3Department of Emergency Center, Peking University First Hospital, Peking University, Beijing, 100050, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jianrong Su, Department of Clinical Laboratory, Beijing Friendship Hospital, Capital Medical University, No. 95, Yong’an Road, Xicheng District, Beijing, 100050, People’s Republic of China, Email [email protected] Miaorong Xie, Department of Emergency and Critical Care Center, Beijing Friendship Hospital, Capital Medical University, No. 95, Yong’an Road, Xicheng District, Beijing, 100050, People’s Republic of China, Email [email protected]
Purpose: To investigate the antimicrobial activity of berberine and the mechanism by which it combats methicillin-resistant Staphylococcus aureus (MRSA) strains isolated from patients with bloodstream infections.
Methods: Fifteen clinical MRSA isolates were collected, and their Multi-locus Sequence Types (MLST) were examined. The minimum inhibitory concentration (MIC) and combined antibacterial activity of berberine alone, and when combined with clindamycin and rifampicin separately, were determined. Additionally, two MRSA strains (ST239 and ST5) were selected to perform the time-killing assay and biofilm formation test. Cell wall alterations and cell membrane integrity were measured by confocal laser scanning microscopy (CLSM) and electron microscopy to assess the influence on cell morphology.
Results: Our data showed berberine was effective against MRSA at MIC values varying from 256 to 64 mg*L− 1 for different MLST types. Berberine alone, and when combined with clindamycin and rifampicin separately, displayed excellent antibacterial activity which reduced the bacterial counts by 2lgCFU*mL within 24h and significantly weakened biofilm formation compared with control strain. Additionally, bacterial cytological profiling indicates that berberine destroyed the structure of the cell walls, membrane integrity and further changed the cell morphology with concentration increased.
Conclusion: In our study, berberine has excellent anti-MRSA activities and has synergistic antibacterial property when combined with clindamycin and rifamycin separately, and the mechanism of activities involves the destruction of cell wall and membrane.
Keywords: methicillin-resistant Staphylococcus aureus, berberine, multi-locus sequence types, synergistic effects, biofilm, mechanism
A Letter to the Editor has been published for this article.
A Response to Letter has been published for this article.
Introduction
Staphylococcus aureus (S. aureus), a major pathogen of healthcare-associated infections, causes many infectious diseases in hospital and community (eg, pneumonia, endocarditis, and bacteremia).1 Due to its well-known exotoxins and ability to form biofilms,2 the S. aureus infections are particularly difficult to treat. Furthermore, S. aureus rapidly develops resistance to the β-lactams antibiotics via the acquisition of the staphylococcal chromosome cassette carrying mecA gene.3–5 The Methicillin-resistance S. aureus (MRSA) has become one of the most lethal pathogen in bacteremia. The case fatality rate of MRSA bacteremia has been reported to be up to 27.3%, with a mortality rate approximately 2–3 times higher than those infected with methicillin-susceptible S. aureus.6 Although vancomycin remains the most effective antibiotic currently, the treatment of MRSA infections is still challenging due to the dramatic increase in MRSA isolation and the limited clinical treatment options.7,8 Thus, there is a strong need to develop novel antibiotics for the treatment of MRSA.
Berberine (BER) is an isoquinoline quaternary alkaloid (5,6-dihydrodibenzo quinolizinium derivative) isolated from many medicinal plants, such as Hydrastis canadensis, Berberis aristata, Coptis chinensis, Coptis rhizome, Coptis japonica, Phellodendron amurense, and Phellodendron chinense schneid.9 BER is used to treat diarrhea and bowel disorders in China, with the potential to be used for the treatment of many diseases.10 Furthermore, BER has received extensive attention for its role as an antimicrobial,11–13 antifungal,14 anti-tumor,15,16 and anti-diabetes drug.17 However, the anti-MRSA mechanism of BER is still unclear.
Therefore, the current study evaluated the effects of BER on MRSA using the minimum inhibitory concentration (MIC) and time-killing test. Moreover, the potential mechanism of BER’s activities on MRSA, including biofilm formation, cell membrane integrity, and cell morphology variation were investigated. We also explored the synergistic effect of BER combined with rifampicin and clindamycin against MRSA in vitro.
Materials and Methods
Bacterial Collection
Fifteen MRSA isolates were collected from blood cultures between March and September 2021 at Beijing Friendship Hospital, affiliated with the Capital Medical University of China. After collection, the bacteria were labeled A–O and stored in skim milk at −80°C for further use. Subsequently, American Type Culture Collection (ATCC) 43300 was selected as the control strain. Bacteria were routinely grown in a Columbia blood plate (Thermo Fisher Scientific, Waltham, MA, USA) at 37°C for 18h unless otherwise specified.
Minimum Inhibitory Concentration Assay
The minimum inhibitory concentration (MIC) of each antibacterial agents is defined as the lowest concentration of the drug that resulted in no visible growth.18 Berberine (BER), clindamycin (CLI), and rifampicin (RIF) were acquired from the National Institutes for Food and Drug Control (Beijing, China). The MICs of BER were determined based on the agar dilution method recommended by the Clinical and Laboratory Standards Institute (CLSI, PA, USA). Mueller-Hinton agar (MHA, OXOID) was mixed with BER in dishes, where the final concentration of BER was 1, 2, 4, 8, 16, 32, 64, 128, 256, 512, 1024, 2048 mg*L.−1 The MICs of antibiotics, including RIF and CLI, against MRSA strains were determined using a standard broth microdilution method recommended by the CLSI.19 Cation adjusted Mueller-Hinton broth (CAMHB, OXOID) was mixed with RIF and CLI separately, where the final concentration of RIF and CLI was twofold series dilution concentration from 0.015625 to 2048 mg*L−1 in triplicate. Then, the tested bacterial were spotted on the medium and incubated at 37°C for 18h. All experiments were conducted at least three times on different days, and the most consistent results were presented.
MLST Determination
MLST was determined by the PCR amplification of seven housekeeper genes (aro, arc, glp, gmk, pta, tpi, and yqi), according to the conditions published in https://pubmlst.org/. The PCR products were bidirectionally sequenced, and all sequences in the online database were analyzed to determine the allele number and respective ST types.
Synergistic Effects on Checkerboard Assay
Checkerboard assays were used to determine the interactions between BER and antibiotics on the tested MRSA strains, as recommended by the CLSI guidelines. The concentration gradients of BER and antibiotics were selected based on the MICs. After MRSA cells were cultured in CAMHB medium with continuous shaking at 37°C, the cells were grown to 0.5McF (1.5×108 colony forming unit*mL−1, CFU*mL−1); then, the cultures were diluted 32-fold with sterile broth (approximately 5×106CFU*mL−1). Delivering 10μL aliquots of cultures to 96-well plates containing 100μL of two-fold diluted BER and antibiotics in cation-adjusted Mueller Hinton broth, where the final concentration of bacterial culture was approximately 5×105 CFU*mL−1.20 The plates were incubated for 18 h at 37°C. Finally, each well was examined for signs of MRSA growth. For each interaction, the fractional inhibitory concentration (FIC) was calculated for each agent: synergism, antagonism, and indifference correlated with FIC ≤ 0.5, FIC > 4, and 0.5 < FIC ≤ 4, respectively.21
Time-Killing Curves
Growth kinetics were used to assess the antimicrobial effect of BER against MRSA and sensitization to other antibiotics.22 The tested MRSA strain was selected based on MLST types. The experiments were performed using CAMHB in 10mL with an initial inoculum of 5×105 CFU/mL. The cultures were transferred to tubes with individual compounds and in combination (2 × MIC, 1 × MIC, 1/2 × MIC, 1/4 × MIC of BER, 1/8 × MIC, 1/32 × MIC of RIF, or 1/4 × MIC of CLI) treated and incubated. The concentration of BER and antibiotics were chosen based on the MIC assay and synergistic effects assay. A kinetic growth or inactivation curve was constructed for each treatment using the viable cell counts performed at 0 h, 2 h, 4 h, 6 h, 8 h, 10 h, 12 h, 24h of incubation at 37°C. The cultures containing 1% DMSO and bacterial inoculum served as the bacterial DMSO control (DC) and growth control (GC) separately.
Biofilm Formation
The effect of BER on MRSA biofilm formation was tested. Static biofilm formation assay was performed in 96-well microtiter plates, as previously reported.23 Delivering approximately 5×105 CFU*mL−1 of MRSA cells to 96-well plates containing diluted BER and antibiotics in brain heart infusion broth (BHI, OXIOD) supplemented with 0.5% glucose.21 Wells containing only the BHI were used as the negative control, whereas wells containing bacteria grown in BHI were used as the positive control. The suspension was transferred to 96-well plates with individual compounds and combinations (4 × MIC, 2 × MIC, 1 × MIC, 1/2 × MIC, 1/4 × MIC of BER, 1/16 × MIC, 1/32 × MIC of RIF, or 1/4 × MIC of CLI) treated and incubated for 18 h at 37°C. The concentrations of BER, RIF, and CLI were selected based on the previous MIC assays and synergistic effects assays. The tested MRSA strain was selected based on the MLST types, consistent with kinetics assay. After incubation, each well was washed three times with PBS to remove the planktonic cells, fixed for 15 min with methanol to stabilize biofilm, stained for 5 min with crystal violet (0.5% in methanol), and washed two times again with deionized water. The adhering dye was dissolved in 200μL of 33% acetic acid and the absorbance was measured at OD600 to quantify biofilm formation.
Cell Membrane Integrity
Cell membrane permeability of MRSA cells was assessed by confocal laser scanning microscopy (CLSM) (TCS-SP5, Leica, GER), similar to the previous description.24 To test membrane integrity, fluorescence images were acquired using fluorochrome, which were carboxyfluorescein diacetate (CFDA SE; Sigma-Aldrich, St. Louis, MO, USA) and propidium iodide (PI; Sigma-Aldrich, St. Louis, MO, USA) at final concentrations of 10 mmol*L−1 and 5 mg*L−1, separately. CFDA SE is a nonfluorescent precursor that only passing through intact cell membranes, producing a fluorescent compound. When the cell membrane is damaged, PI can enter the cells and bind to the DNA to form a red fluorescent DNA-complex.25 S. aureus are gram-positive cocci with a cytoderm structure. However, cytomembrane was essential to maintain cell function and prevent cytoplasmic leakage. Fluorescent staining was used to examine cell membrane integrity as previously reported.24 After MRSA cells were grown to an approximately 1.5×108CFU*mL−1 in CAMHB at 37°C, delivering 10 mL of volume cultures to tubes. Then, the cells were treated with BER at 0 × MIC, 1 × MIC, or 2 × MIC for 4 h, centrifuged at 5000×g for 5 min, and resuspended with equivalent volumes of 0.9% NaCl. After incubation with cFDA SE for 15 min and PI for 1 min at room temperature away from light, the suspension was fixed on a glass slide using agar. Finally, the bacteria were examined by confocal laser scanning microscopy (CLSM, OLYMPUS, FV1000, Japan) at the excitation-emission wavelengths of 530nm for PI and 488nm for CFDA SE.
Cell Morphology by Electron Microscopy
Field emission scanning electron microscopy (FESEM, Joel JSM-7900F, Japan) was used to detect morphological changes after BER treatment, according to the published protocols.26 Subsequently, approximately 1.5×108CFU*mL−1 of MRSA cells were incubated in CAMHB of 10mL volume supplemented with BER at 0 × MIC, 1 × MIC, 2 × MIC, and 4 × MIC for 4 h at 37°C. The treated cells were washed with PBS three times and fixed with 2.5% glutaraldehyde for 4 h and 1% osmic acid for 2 h. Then, the cells were washed with PBS again. After dehydrating with different concentrations of ethanol, the cells were fixed on silicon slices, sputter-coated with gold under vacuum, and observed using FESEM.
Statistical Analysis
Assays were conducted with at least two biological replications. The Kruskal–Wallis analysis was used to determine the differences between the multiple groups. Data are expressed as mean ± SD, and p-values <0.05 are considered statistically significant. No adjustments were made for the p-values reported in this study.
Results
MLST Determination, MICs, and FIC Index
The MLST determination showed 15 MRSA clinical isolates and distinguished them into five subtypes. The MIC values of BER varying from 256 to 64 mg*L−1 (Table 1); however, among ST5 subtypes, only one strain had a MIC significantly different from the others (ie, >2048 mg*L−1). Excluding this strain, the highest MIC value was 256 mg*L−1 for ATCC 43300, and the lowest MIC value was 64 mg*L−1 for ST239 and ST5. Six clinical isolates were resistant to CLI, and eight strains were susceptible, whereas 11 isolates were resistant to RIF, and three isolates were susceptible. The MRSA control strain was resistant to CLI at MIC of 1024 mg*L−1 and susceptible to RIF at MIC of 0.06 mg*L−1 (Table 2). Using the checkerboard assay, BER in combination with RIF and CLI showed a significant effect against the MRSA strain, leading to a reduction in MIC values. Synergistic activity was observed for the combination of BER with CLI or RIF with FIC index values of 0.19–0.5 (Table 2). However, two isolates showed indifferent effects compared to others when BER was combined with CLI, and one isolate showed indifferent effects when BER was combined with RIF (Table 3).
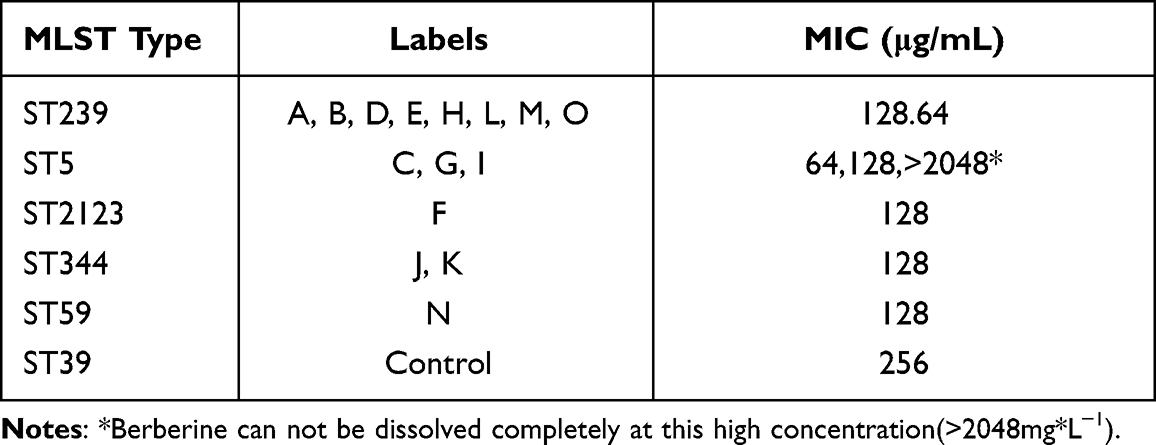 |
Table 1 The MIC Value of Berberine Among Different MLST Types of MRSA |
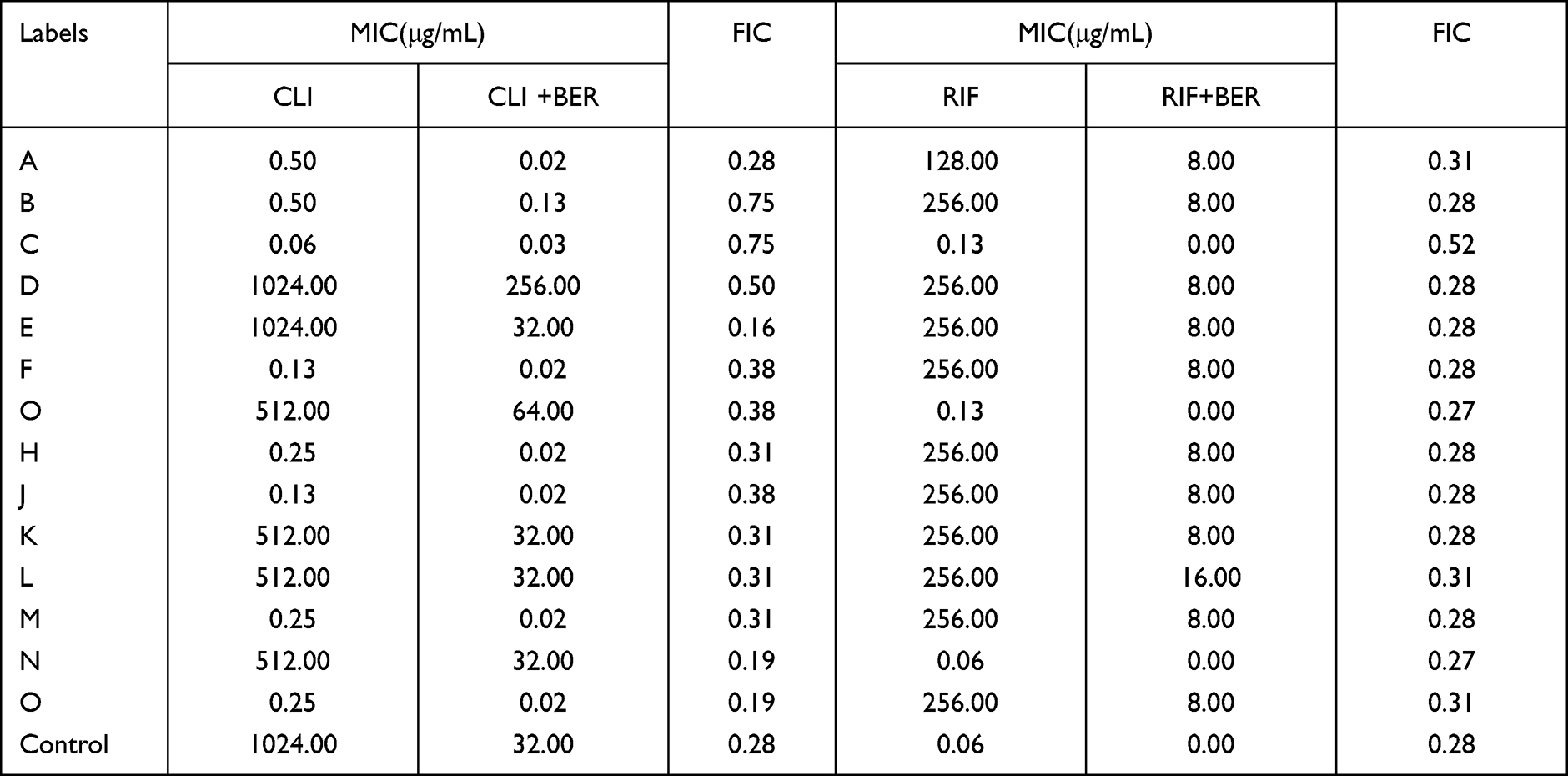 |
Table 2 MIC and FIC Index of CLI and RIF in the Presence of BER Against MRSA Strains |
 |
Table 3 The Antibacterial Activity of BER in Combination with CLI and RIF Against MRSA Strains (%, n/N) |
Time-Killing Curves
The time-killing curves for BER alone and in combination with RIF or CLI against ATCC 43300 (control strain) and clinical isolate ST239 are presented in Figure 1. Growth cultures without BER treatment increased to 7lg (CFU*mL−1) for the control strain and 9lg (CFU*mL−1) at ST239 after 24 h of incubation. The DMSO control group showed similar results to the growth cultures, suggesting that DMSO does not affect the growth of cells. Furthermore, BER at its MIC value showed dramatic inhibition of bacterial growth during 24 h of incubation, and showed stronger inhibition at 2 × MIC than at MIC after 12 h of incubation with similar bacterial growth compared to that at MIC after 24 h of incubation (reduction of 2lg (CFU*mL−1) and 1lg (CFU*mL−1) for ATCC 43300 and ST239, separately). BER at sub-MIC had a weak effect on the growth of the MRSA strain over 24 h; however, bacterial growth was positive (Figure 1A and D). Importantly, the combination of BER with CLI showed a synergistic effect against the MRSA control strain and clinical isolate ST239 with ~2lg (CFU*mL−1) killing compared to the single drug applied during 24 h (Figure 1B and E). In addition, the combination of BER with RIF showed more remarkable synergistic effects against MRSA than those of BER and CLI. As such, the combination of BER with RIF resulted in a > 2L g (CFU*mL−1) inhibition compared to the combination of BER with CLI at 24 h (Figure 1C and F).
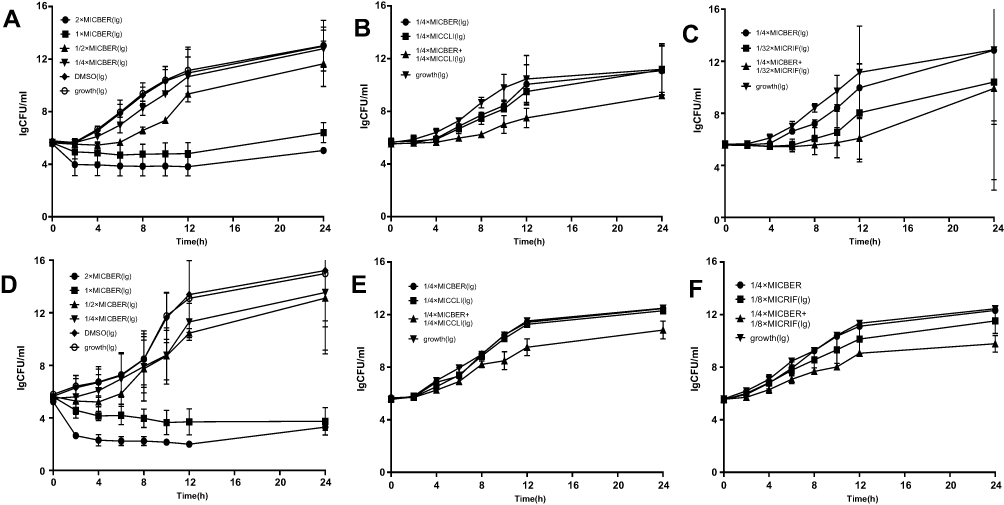 |
Figure 1 The time-killing curves for BER alone and in combination with RIF or CLI against MRSA strains; (A–C): Control (D–F): ST239. The data are the mean ±SD of colony counts at least two biological replicates in each group. |
Effects of BER Alone and in Combination with RIF or CLI on Biofilm Formation
According to the assay of biofilm formation, BER possessed an excellent inhibitory effect on the biofilm in a dose-dependent manner, especially at the MIC or higher concentrations, which was more prominent than that of the sub-MIC and control groups. Interestingly, biofilm formation sharply increased at sub-MIC in the ST 239 strain (Figure 2). In addition, the combination of BER with RIF or CLI at sub-MIC significantly inhibited biofilm formation compared to each drug separately in the control strain and clinical isolate (Figure 3). The percentages of biofilm reduction, which were compared with positive control in each group, are presented at Table 4.
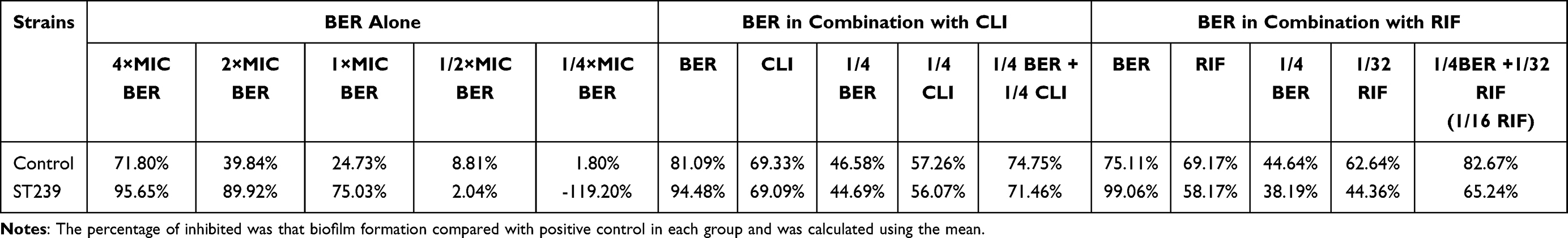 |
Table 4 The Percentage of Biofilm Formation Inhibited |
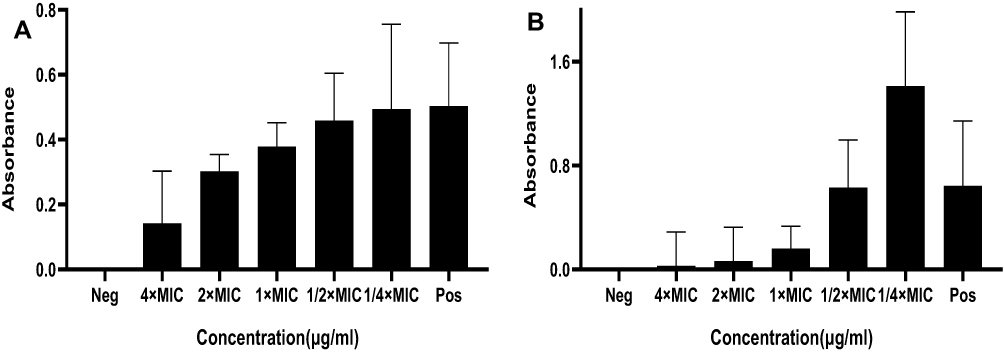 |
Figure 2 The effects of BER alone on biofilm formation of MRSA strains. (A) Control (B) ST 239; relative biofilm formation levels were represented as mean±SD of at least two biological replicates; significant difference was determined at P < 0.0001 with comparison between the groups. |
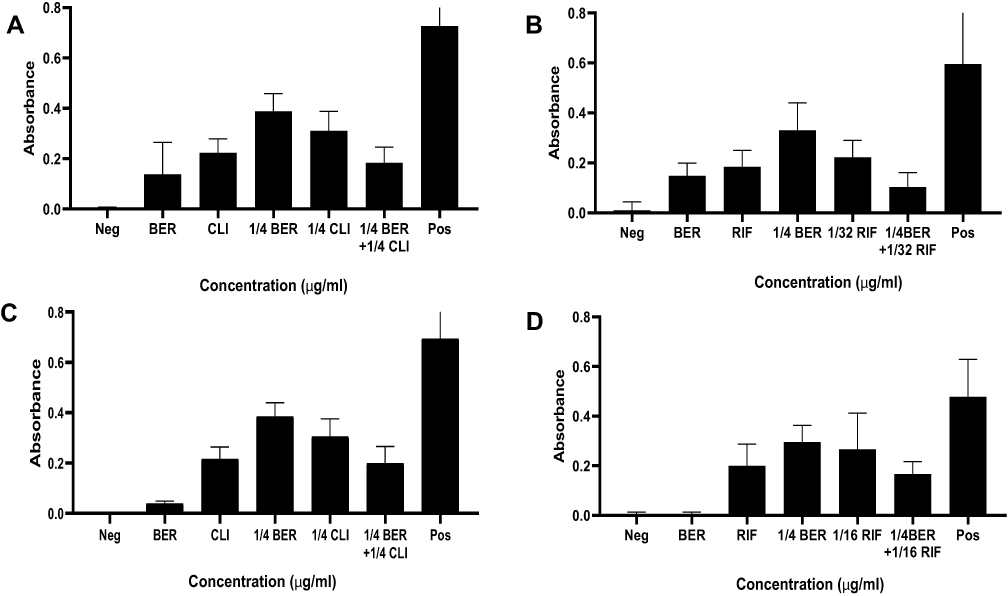 |
Figure 3 The effects of BER in combination with RIF or CLI on biofilm formation of MRSA strains. (A and B): Control (C and D): ST 239; relative biofilm formation levels were represented as mean±SD of at least two biological replicates; significant difference was determined at P < 0.0001 with comparison between the groups. |
Cell Membrane Integrity
According to the images, untreated MRSA cells emitted green fluorescence, showing that the cell membrane was intact. Conversely, BER-treated MRSA appeared red and the proportion of red fluorescence increased with increasing BER concentrations, suggesting a damaged cell membrane (Figure 4). Meanwhile, the total number of cells declined after BER treatment, and the reduction was proportional to the BER concentration.
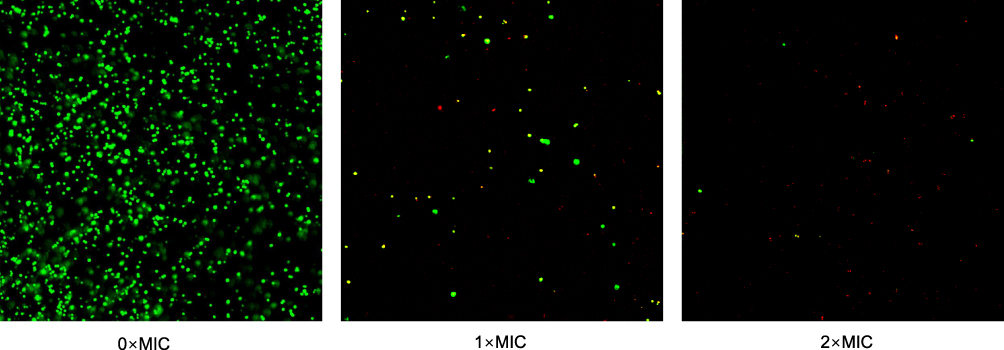 |
Figure 4 The effect of BER on cell membrane integrity of MRSA strain by confocal laser scanning microscopy. |
Alternation in Morphology
The FESEM images illustrated significant changes in the morphology of MRSA cells with BER (Figure 5) at low magnification (10,000–15,000×) and high magnification (45,000–50,000×). Untreated control S. aureus exhibited arrangement in clusters and smooth surface and spherical contour at 10,000× and 50,000×, separately (Figure 5A and E). However, the number of MRSA cells in the field of vision decreased and the morphology of the clusters was changed with increasing BER concentrations (Figure 5B–D). Remarkably, numerous flocculent substances were adhering to the cells that were not significantly cleaned by PBS. These flocculent substances increased with the increase in BER concentration compared to the untreated control. Furthermore, the cells were less uniform in size and had a rough surface (50,000×) compared to the untreated cells. When treated with MIC or higher concentrations of BER, the cell morphology exhibited shrinkage and deformation (Figure 5F); some cells even had a defective and punched surface, suggesting that the content was released (Figure 5G and H).
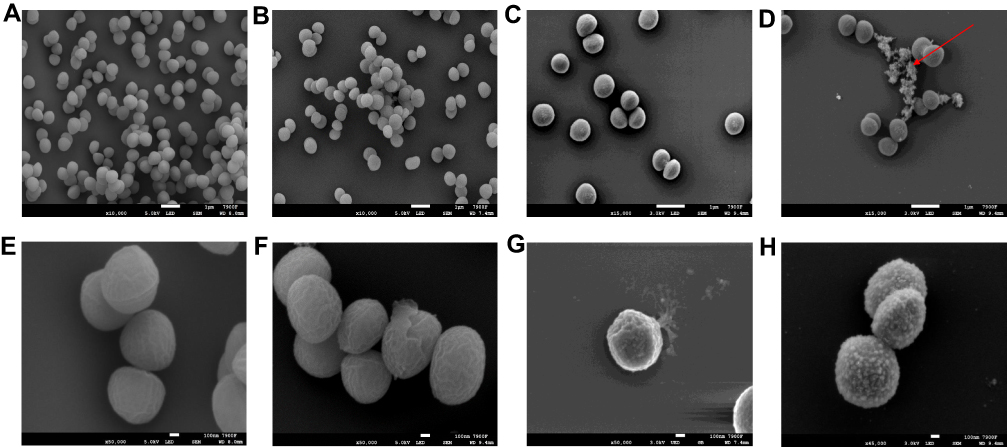 |
Figure 5 The changes in the morphology of MRSA cells with BER by field emission scanning electron microscope. (A and E): 0×MIC; (B and F): 1×MIC; (C and G): 2×MIC; (D and H): 4×MIC. Shown by the arrow is flocculent substances. |
Discussion
The dramatic increase and global spread of MRSA strains pose a serious threat to human health since it was first identified in 1960.27 Vancomycin and daptomycin are recommended as the first-line antibiotics by the 2011 Infectious Diseases Society of America guidelines for MRSA.28 However, each antimicrobial agent has limitations29 related to its effectiveness against bacteria30 or its increasing resistance.31,32 What is more, the emergence of vancomycin-resistant S. aureus leaves physicians with limited therapeutic options. Therefore, there is an urgent need in the development of new therapeutics to aid in the treatment of infectious diseases caused by MRSA. BER, an alkaloid extracted from a traditional Chinese herb, has emerged as a promising antibacterial agent.
In the current study, we observed that BER had anti-MRSA activities at MIC of 64–256 mg*L−1, which related to the subtype of MRSA strains. As previously reported, subtype strains within the same bacterial species may have different clinical uses.2 In this study, 15 strains were divided into five subtypes. We found that the MIC value of BER differed among various MLST types, and many strains showed diverse BER MIC values even in the same subtype. The MIC showed three values for ST5 and two values for ST239, unlike what has been reported in previous studies.2 In this study, a subtype was matched with a MIC value. However, the MIC value of ST239 and ST39 in our study which was 64 or 128 mg*L−1 and 256mg*L−1, was different with this study (64 mg*L−1 and 512 mg*L−1).2 Therefore, we recommend that the types of MLST should be expanded to verify the results.
In the current investigation, a synergistic effect of BER with CLI and RIF was observed against MRSA. As such, BER can markedly reduce the MIC value of antibacterial agents against MRSA. A synergistic effect of BER has also been reported in combination with oxacillin,33 azithromycin,34 and levofloxacin,34 whereas an additive effect has been reported for BER in combination with ampicillin33,34 and cefazolin.34 This study is the first one to report the synergistic effects of BER in combination with CLI and RIF.
The time-killing assays showed similar results. The time-killing test of the control strains, the strain ST239 for BER alone, and the combination of BER with CLI or RIF separately showed significant growth inhibition. The inhibition of growth by BER alone strengthened with the increasing concentration. Notably, the test strain ST239 and the control strain showed attenuation of inhibition with the increase in curve after 12 h, suggesting that BER alone may not kill all the cells, but it maintains the bacteria at low levels when the BER gradually expands. When this occurs, the remaining cells are reduced, which is similar to the results of a previous study on the antimicrobial activity of a Chinese herb.35 Therefore, there is a significant effect of BER concentration on its antimicrobial activity, suggesting that BER must maintain a persistently high blood concentration. In addition, the combined treatment resulted in obvious reduction in the cell quantity within 8–24 h of incubation in the control strain and 10–24 h of incubation in ST239, compared to CLI or RIF separately. These results suggest that BER has excellent anti-MRSA activity, which is enhanced when combined with other antibiotics, such as CLI and RIF.33,34,36–39
Biofilm formation plays an essential role in the persistence of pathogens.34,40 Biofilms harbor bacteria that are irreversibly attached to cell surfaces and prevent antibiotic exposure.41 In light of this, BER showed excellent inhibition of biofilm formation with increasing concentrations, similar to previous studies.2,42 However, biofilm formation increased at sub-MIC in the ST 239 strain in our study, similar to other studies.2 The concentration of BER cannot increase slowly when treating MRSA infections, because low concentration of BER may promote biofilm formation. Importantly, BER, in combination with CLI or RIF separately, also inhibited biofilm formation better than either drug alone. These data are consistent with previous studies that suggest the existence of synergistic effects of BER with those of CLI and RIF against MRSA. Therefore, antibacterial activity and anti-biofilm formation character of BER can be used as the basis of clinical practice and the synergistic effect of BER with CLI and RIF can be used as the alternative treatment when the bacterial infection was difficult to treat. However, the specific mechanism underlying the effects of BER in combination with CLI or RIF separately remains unclear and should be explored in future studies.
Morphological analysis was used to explore the mechanism of the BER effect against MRSA. In the current study, we observed cell wall alteration and its punched appearance due to BER exposure at high magnification. Flocculent substances were observed to adhere to the cells. Zhang et al found that BER could damage the cell surface of MRSA.43 Therefore, it is reasonable to speculate that the substances may include the contents of punched bacterial cells due to BER treatment. This observation is consistent with the above findings, illustrating that BER can destroy MRSA cell walls in a concentration-dependent manner.
The cytoplasmic membrane is an important dynamic structure that allows cells to interact with other cells and environmental molecules.44 While the cytoplasmic membrane integrity of MRSA has not been studied previously, the function of the gram-positive bacterial membrane is essential for subsisting membrane proteins and transportation.45 As assessed by CLSM, our results showed that the integrity of the MRSA cytoplasmic membrane was compromised at increased BER concentrations. This finding showed that BER possessed anti-MRSA activities by damaging the cell wall and cytoplasmic membrane to perturb cell functions and release intracellular contents. Although BER could exert an influence on the cell wall and cytoplasmic membrane, the mechanism here is unclear and needs to be investigated further. Meanwhile, combination antibacterial assays would verify the hypothesis that BER could damage the cytoderm and cytoplasmic membrane to kill MRSA. Because CLI and RIF play antibacterial roles inside the cells,46,47 they may easily enter the cells in the presence of BER.
Conclusions
The current study demonstrated that BER has remarkable antimicrobial effects against MRSA with favorable MIC. It can also be used as an adjuvant to RIF or CLI to treat MRSA infections. BER alone and in combination with RIF or CLI reduces biofilm formation. In addition, BER significantly damages the cell wall and membrane of MRSA to kill the bacteria. This study provides novel mechanistic insight into supporting the development of BER as an antibacterial drug.
Data Sharing Statement
The datasets generated for this study are available from the corresponding author Miaorong Xie.
Ethics Statement
This study was approved by the Ethics Committee of Beijing Friendship hospital, Capital Medical University (20210208). Adult patients wrote the informed consent and a parent or legal guidance of patients under 18 years of age provided informed consent prior to the experiment described below. This study was conducted in accordance with the Declaration of Helsinki.
Acknowledgments
We thank Wei-dong Qian from Shaanxi University of Science and Technology providing the experimental consults.
Funding
This work was supported by the National Key New Drug Creation and Manufacturing Program, Ministry of Science and Technology (YFC1702605).
Disclosure
The authors declare no conflicts of interest for this study.
References
1. Bayer MG, Heinrichs JH, Cheung AL. The molecular architecture of the sar locus inStaphylococcus aureus. J Bacteriol. 1996;178(15):4563–4570.
2. Tan J, Wang J, Yang C, et al. Antimicrobial characteristics of Berberine against prosthetic joint infection-related Staphylococcus aureus of different multi-locus sequence types. BMC Complement Altern Med. 2019;19(1):218. doi:10.1186/s12906-019-2558-9
3. Katatama Y. A new class of genetic element, staphylococcus cassette chromosome mec, encodes methicillin resistance in Staphylococcus aureus. Antimicrob Agents Chemother. 2000;44:1549–1555.
4. Pinho MG, Filipe SR, de Lencastre H, Tomasz A. Complementation of the essential peptidoglycan transpeptidase function of penicillin-binding protein 2 (PBP2) by the drug resistance protein PBP2A inStaphylococcus aureus. J Bacteriol. 2001;183(22):6525–6531.
5. Harkins CP, Pichon B, Doumith M, et al. Methicillin-resistant Staphylococcus aureus emerged long before the introduction of methicillin into clinical practice. Genome Biol. 2017;18(1). doi:10.1186/s13059-017-1252-9
6. Paulsen J, Mehl A, Askim A, Solligard E, Asvold BO, Damas JK. Epidemiology and outcome of Staphylococcus aureus bloodstream infection and sepsis in a Norwegian county 1996–2011: an observational study. BMC Infect Dis. 2015;15:116. doi:10.1186/s12879-015-0849-4
7. Wang L, Yang R, Yuan B, Liu Y, Liu C. The antiviral and antimicrobial activities of licorice, a widely-used Chinese herb. Acta Pharm Sin B. 2015;5(4):310–315. doi:10.1016/j.apsb.2015.05.005
8. Boucher H, Miller LG, Razonable RR. Serious infections caused by methicillin-resistant Staphylococcus aureus. Clin Infect Dis. 2010;51(Suppl 2):S183–S197. doi:10.1086/653519
9. Imanshahidi M, Hosseinzadeh H. Pharmacological and therapeutic effects of Berberis vulgaris and its active constituent, berberine. Phytother Res. 2008;22(8):999–1012. doi:10.1002/ptr.2399
10. Tillhon M, Guaman Ortiz LM, Lombardi P, Scovassi AI. Berberine: new perspectives for old remedies. Biochem Pharmacol. 2012;84(10):1260–1267. doi:10.1016/j.bcp.2012.07.018
11. Slobodnikova L, Kost’alova D, Labudova D, Kotulova D, Kettmann V. Antimicrobial activity of Mahonia aquifolium crude extract and its major isolated alkaloids. Phytother Res. 2004;18(8):674–676. doi:10.1002/ptr.1517
12. Yan D, Jin C, Xiao XH, Dong XP. Antimicrobial properties of berberines alkaloids in Coptis chinensis Franch by microcalorimetry. J Biochem Biophys Methods. 2008;70(6):845–849. doi:10.1016/j.jbbm.2007.07.009
13. Roser E, Grundemann C, Engels I, Huber R. Antibacterial in vitro effects of preparations from anthroposophical medicine. BMC Complement Altern Med. 2016;16(1):372. doi:10.1186/s12906-016-1350-3
14. Xiao CW, Ji QA, Wei Q, Liu Y, Bao GL. Antifungal activity of berberine hydrochloride and palmatine hydrochloride against Microsporum canis -induced dermatitis in rabbits and underlying mechanism. BMC Complement Altern Med. 2015;15:177. doi:10.1186/s12906-015-0680-x
15. Gu S, Song X, Xie R, et al. Berberine inhibits cancer cells growth by suppressing fatty acid synthesis and biogenesis of extracellular vesicles. Life Sci. 2020;257:118122. doi:10.1016/j.lfs.2020.118122
16. Shen ZQ, Wang J, Tan WF, Huang TM. Berberine inhibits colorectal tumor growth by suppressing SHH secretion. Acta Pharmacol Sin. 2020;42:1190–1194. doi:10.1038/s41401-020-00514-2
17. Zhang Y, Gu Y, Ren H, et al. Gut microbiome-related effects of berberine and probiotics on type 2 diabetes (the PREMOTE study). Nat Commun. 2020;11(1):5015. doi:10.1038/s41467-020-18414-8
18. Wiegand I, Hilpert K, Hancock REW. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc. 2008;3(2):163–175. doi:10.1038/nprot.2007.521
19. Isenberg HD. Minimum bactericidal concentration testing. In: Clinical Microbiology Procedures Handbook. Wiley Online Library; 2016:
20. Isenberg HD. Synergism testing: broth microdilution checkerboard and broth macrodilution methods. In: Clinical Microbiology Procedures Handbook. Wiley Online Library; 2016:
21. Wang G, Li L, Wang X, et al. Hypericin enhances beta-lactam antibiotics activity by inhibiting sarA expression in methicillin-resistant Staphylococcus aureus Acta Pharm Sin B. 2019;9(6):1174–1182. doi:10.1016/j.apsb.2019.05.002
22. Isenberg HD. Time-kill. In: Clinical Microbiology Procedures Handbook. Wiley Online Library; 2016:
23. Xu Y, Shi C, Wu Q, et al. Antimicrobial activity of punicalagin against Staphylococcus aureus and its effect on biofilm formation. Foodborne Pathog Dis. 2017;14(5):282–287. doi:10.1089/fpd.2016.2226
24. Yanfei L, Xiaobin F, Caiping G, et al. Flow cytometry for viability analysis of three bacteria species with fluorescein diacetate vs propidium iodide staining. Acad J Guangzhou Med Univ. 2015;43(02):29–31.
25. Qian W, Wang W, Zhang J, et al. Antimicrobial and antibiofilm activities of ursolic acid against carbapenem-resistant Klebsiella pneumoniae. J Antibiot. 2020;73(6):382–391. doi:10.1038/s41429-020-0285-6
26. Bozzala JJ. Electron Microscopy Principles and Techniques for Biologists. Jones and Bartlett Publishers; 1999.
27. Kaddurah-Daouk R, Kristal BS, Weinshilboum RM. Metabolomics: a global biochemical approach to drug response and disease. Annu Rev Pharmacol Toxicol. 2008;48:653–683. doi:10.1146/annurev.pharmtox.48.113006.094715
28. Liu C, Bayer A, Cosgrove SE, et al. Clinical practice guidelines by the infectious diseases society of America for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children: executive summary. Clin Infect Dis. 2011;52(3):285–292. doi:10.1093/cid/cir034
29. Hassoun A, Linden PK, Friedman B. Incidence, prevalence, and management of MRSA bacteremia across patient populations-a review of recent developments in MRSA management and treatment. Crit Care. 2017;21(1):211. doi:10.1186/s13054-017-1801-3
30. Rodvold KA, McConeghy KW. Methicillin-resistant Staphylococcus aureus therapy: past, present, and future. Clin Infect Dis. 2014;58(Suppl 1):S20–S27. doi:10.1093/cid/cit614
31. McGuinness WA, Malachowa N, DeLeo FR. Vancomycin resistance in Staphylococcus aureus. Yale J Biol Med. 2017;90(2):269–281.
32. Gomez Casanova N, Siller Ruiz M, Munoz Bellido JL. Mechanisms of resistance to daptomycin in Staphylococcus aureus. Rev Esp Quimioter. 2017;30(6):391–396.
33. Yu HH, Kim KJ, Cha JD, et al. Antimicrobial activity of berberine alone and in combination with ampicillin or oxacillin against methicillin-resistant Staphylococcus aureus. J Med Food. 2005;8(4):454–461. doi:10.1089/jmf.2005.8.454
34. Zuo GY, Li Y, Han J, Wang GC, Zhang YL, Bian ZQ. Antibacterial and synergy of berberines with antibacterial agents against clinical multi-drug resistant isolates of methicillin-resistant Staphylococcus aureus (MRSA). Molecules. 2012;17(9):10322–10330. doi:10.3390/molecules170910322
35. Liu QQ, Han J, Zuo GY, Wang GC, Tang HS. Potentiation activity of multiple antibacterial agents by Salvianolate from the Chinese medicine Danshen against methicillin-resistant Staphylococcus aureus (MRSA). J Pharmacol Sci. 2016;131(1):13–17. doi:10.1016/j.jphs.2015.10.009
36. Wojtyczka RD, Dziedzic A, Kepa M, et al. Berberine enhances the antibacterial activity of selected antibiotics against coagulase-negative Staphylococcus strains in vitro. Molecules. 2014;19(5):6583–6596. doi:10.3390/molecules19056583
37. Aksoy CS, Avci FG, Ugurel OM, Atas B, Sayar NA, Sariyar Akbulut B. Potentiating the activity of berberine for Staphylococcus aureus in a combinatorial treatment with thymol. Microb Pathog. 2020;149:104542. doi:10.1016/j.micpath.2020.104542
38. Gao S, Zhang S, Zhang S. Enhanced in vitro antimicrobial activity of amphotericin B with berberine against dual-species biofilms of Candida albicans and Staphylococcus aureus. J Appl Microbiol. 2021;130(4):1154–1172. doi:10.1111/jam.14872
39. Liang RM, Yong XL, Duan YQ, et al. Potent in vitro synergism of fusidic acid (FA) and berberine chloride (BBR) against clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA). World J Microbiol Biotechnol. 2014;30(11):2861–2869. doi:10.1007/s11274-014-1712-2
40. Rabin N, Zheng Y, Opoku-Temeng C, Du Y, Bonsu E, Sintim HO. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med Chem. 2015;7(4):493–512. doi:10.4155/fmc.15.6
41. Jamal M, Ahmad W, Andleeb S, et al. Bacterial biofilm and associated infections. J Chin Med Assoc. 2018;81(1):7–11. doi:10.1016/j.jcma.2017.07.012
42. Chu M, Zhang MB, Liu YC, et al. Role of berberine in the treatment of methicillin-resistant Staphylococcus aureus infections. Sci Rep. 2016;6:24748. doi:10.1038/srep24748
43. Zhang X, Sun X, Wu J, et al. Berberine damages the cell surface of methicillin-resistant Staphylococcus aureus. Front Microbiol. 2020;11:621. doi:10.3389/fmicb.2020.00621
44. Zhang R, Qin X, Kong F, Chen P, Pan G. Improving cellular uptake of therapeutic entities through interaction with components of cell membrane. Drug Deliv. 2019;26(1):328–342. doi:10.1080/10717544.2019.1582730
45. Wang D, Yu L, Xiang H, et al. Global transcriptional profiles of Staphylococcus aureus treated with berberine chloride. FEMS Microbiol Lett. 2008;279(2):217–225. doi:10.1111/j.1574-6968.2007.01031.x
46. Spizek J, Rezanka T. Lincosamides: chemical structure, biosynthesis, mechanism of action, resistance, and applications. Biochem Pharmacol. 2017;133:20–28. doi:10.1016/j.bcp.2016.12.001
47. Rothstein DM. Alone and in combination. In: Cold Spring Harb Perspect Med. Vol. 6. Rifamycins; 2016:7.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.