")
Back to Journals » Infection and Drug Resistance » Volume 12
In vitro and in vivo activity of ciprofloxacin/fosfomycin combination therapy against ciprofloxacin-resistant Shigella flexneri isolates
Authors Liu Y , Li H , Zhang Y, Ye Y, Gao Y , Li J
Received 8 March 2019
Accepted for publication 3 May 2019
Published 11 June 2019 Volume 2019:12 Pages 1619—1628
DOI https://doi.org/10.2147/IDR.S208071
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Joachim Wink
Yanyan Liu,1,2 Hongru Li,3 Yalong Zhang,1,2 Ying Ye,1,2 Yufeng Gao,1,2 Jiabin Li1,2,4
1Department of Infectious Diseases, The First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China; 2Anhui Center for Surveillance of Bacterial Resistance, Hefei, Anhui, People’s Republic of China; 3Department of Neurology, Xiangya Hospital Central South University, Changsha, People’s Republic of China; 4Department of Infectious Diseases, The Chaohu Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China
Objective: Ciprofloxacin resistance (CIPR) for Shigella isolates is becoming more prevalent. This study systematically investigated the antibacterial activity of ciprofloxacin (CIP)/fosfomycin (FOS) combination in vitro and in vivo against CIPR S. flexneri isolates.
Method: Eighty CIPR S. flexneri isolates were selected for synergy studies by the microtiter plate checkerboard assay. Two S. flexneri isolates (GN120471, CIPRFOSR; GN120454, CIPRFOSS) were used to investigate the efficacy of the CIP/FOS combination by the time-kill methodology. Clinically relevant concentrations (CIP, 0.5, 1, or 2.5 μg/mL; FOS, 30, 150, or 300 μg/mL) were combined, and the colony counts were conducted at 3, 5, 8, and 24 hours. The in vivo activity of the CIP/FOS combination was assessed using a Galleria mellonella larvae model.
Results: In checkerboard assays, 31 strains (38.75%) showed synergy for the CIP/FOS combination. For the isolate GN120471, monotherapy with CIP or FOS at all concentrations produced little or no bacterial killing, while the CIP/FOS combination produced enhanced bacterial killing with FOS concentrations of 150 and 300 μg/mL, especially when combined with CIP at 2.5 μg/mL. For the isolate GN120454, the CIP/FOS combination at all concentrations produced more rapid and extensive killing (up to 5log10 colony forming units (CFU)/mL with many combinations) than with either antibiotic alone. Mortality at 96 hours was around 80% at approximately 104 CFU/larva for GN120471 and GN120454. When CIP at 2.5 μg/mL was combined with FOS at 150 μg/mL for the bactericidal activity in vivo, the survival rates for CIP/FOS combination against GN120471-infected and GN120454-infected larvae were significantly higher than that of CIP (68.75% vs 25%, P=0.013; 81.25% vs 37.5%, P=0.012, respectively).
Conclusion: Against CIPR S. flexneri isolates, the CIP/FOS combination induced synergy, and increased bacterial killing in vitro and in a simple invertebrate model of infection.
Keywords: ciprofloxacin, fosfomycin, antimicrobial synergy, Shigella
Introduction
Shigellosis is an acute invasive enteric infection caused by bacteria belonging to the genus Shigella: S. dysenteriae, S. flexneri, S. boydii, and S. sonnei. Shigella flexneri is the chief cause of endemic shigellosis in developing countries, causing nearly two-thirds of infections.1 While shigellosis is typically self-limiting, treatment with appropriate antimicrobial therapy can shorten the duration of illness and prevent transmission. In the guidelines for the control of shigellosis by the WHO in 2005, ciprofloxacin (CIP) was considered the first-line treatment for shigellosis. However, the widespread use of CIP has led to the development of resistance to CIP and other fluoroquinolones for Shigella isolates, especially in Asia.2–4 Thus, combination therapy has been employed to treat shigellosis caused by CIP-resistant (CIPR) Shigella isolates.
Some studies have demonstrated that fosfomycin (FOS) is a promising agent, especially as part of combination therapy.5–7 FOS was discovered in 1969 and is a phosphonic acid derivative with a molecular mass of 138 Da.8 The mechanism of action is to disrupt the formation of the peptidoglycan precursor uridine diphosphate N-acetylmuramic acid, which is unaffected by other antimicrobials, meaning that there is no cross-resistance with other classes of antimicrobials.9,10 It has a broad spectrum of activity against many bacteria, including multidrug-resistant (MDR) strains, and is generally well tolerated at therapeutic doses.11–13 However, FOS resistance (FOSR) emerges quickly when used as monotherapy.14,15 For this reason, FOS is usually administered in combination with other classes of antimicrobials for the treatment of systemic infections.16 More recently, some studies have demonstrated that the CIP/FOS combination exhibits enhanced in vitro activity against CIPR Pseudomonas aeruginosa strains.17,18 However, no study has examined the in vitro and in vivo activity of the CIP/FOS combination against CIPR Shigella isolates.
Animal studies for this combination are necessary to predict its suitability for clinical use in humans. However, mammalian models of infection are associated with high cost, ethical constraints, and specialized training requirements. Therefore, invertebrate models, such as Galleria mellonella larvae, have been proposed as an alternative to investigate the in vivo activity of antimicrobial agents.19–21 In this study, we demonstrate the in vitro and in vivo efficacy of the CIP/FOS combination against CIPR Shigella isolates in an attempt to gain insights into whether it should be explored further for the treatment of CIPR Shigella infections.
Materials and methods
Bacteria and antimicrobial agents
Eighty CIPR S. flexneri isolates were obtained from the stool samples of patients at 34 hospitals in Anhui, People's Republic of China. Two clinical isolates of S. flexneri (GN120471, CIPRFOSR; GN120454, CIPRFOSS) were employed, which had synergistic effects for the CIP/FOS combination. According to the Clinical and Laboratory Standards Institute (CLSI) guidelines in 2017, susceptibility (S) and resistance (R) were defined as minimal inhibitory concentrations (MICs) of ≤1 and ≥4 μg/mL CIP and ≤64 and ≥256 μg/mL FOS, respectively. In addition, 25 μg/mL glucose-6-phosphate was supplemented when the sensitivity of FOS was detected. All antibiotics were obtained from Sigma-Aldrich China(Shanghai, China).
The study was conducted in accordance with the guidelines of the Declaration of Helsinki, the principles of Good Clinical Practice, and Chinese regulatory requirements, and was approved by the local Ethics Committees of the First Affiliated Hospital of Anhui Medical University (Hefei, People's Republic of China). All patients gave written informed consent.
Synergy testing by the checkerboard assay
MIC values of CIP and FOS were determined with the agar dilution method according to CLSI guidelines. Escherichia coli ATCC 25922 was used as a control. Synergy between CIP and FOS was assessed by the microtiter plate checkerboard assay. In brief, 96-well microtiter plates were set up with increasing concentrations of CIP (1/64MIC-4MIC) in the horizontal wells and FOS (1/32MIC-4MIC) in the vertical wells, and were inoculated with 5×105 CFU/mL of S. flexneri isolates prepared in cation-adjusted Mueller–Hinton broth (CAMHB; with 23.0 μg/mL Ca2+ and 12.2 μg/mL Mg2+; Oxoid [Basingstoke, UK]). Plates were incubated at 37°C overnight and were visually inspected for turbidity to determine growth. Synergy was assessed by calculation of the fractional inhibitory concentration index (FICI). An FICI ≤0.5 was defined as showing a synergistic effect, 0.5< FICI <4.0 as showing no interaction, and FICI ≥4.0 as showing an antagonistic effect.22
Time-kill assays and pharmacodynamic analysis
Bacterial killing was examined using time-kill studies with each antibiotic alone or in combination at an initial inoculum of approximately 106 CFU/mL (GN120471 and GN120454) over 24 hours, according to a previously described methodology.17 The employed concentrations were 0.5, 1, or 2.5 μg/mL CIP and 30, 150, or 300 μg/mL FOS. Nine experiments (three concentrations of CIP × three concentrations of FOS) for combination were performed. Each tube was incubated at 37°C. Serial samples were collected for viable counting at 0, 3, 5, 8, and 24 hours.
Microbiological responses were quantified by the log-change method, which compared the change in log10 CFU/mL from 0 hours (CFU0) to time t (3, 5, 8, or 24 hours; CFUt): log change = log10 (CFUt) – log10 (CFU0). Synergy was defined as a reduction of ≥2 log10 CFU/mL for the combination compared with the most active monotherapy at the specified time, and additivity as a reduction of 1 to <2 log10 CFU/mL for the combination.17
Galleria mellonella treatment assays
The G. mellonella infection model for S. flexneri isolates was performed as previously described.20 Batches of G. mellonella (Kaide Ruixin Co., Tianjin, People's Republic of China) in their final instar stage were stored in the dark at 4°C and were used within 7 days. Larvae masses (250–300 mg) were used to calculate treatment doses.
To establish the lethal inoculum required to cause 80% of the larvae to die over 96 hours, 40 larvae were inoculated with 10 μL of bacterial suspensions (10 larvae for 105, 106, 107, and 108 CFU/mL, respectively) and 10 larvae were inoculated with PBS as controls (Figuire S1). Bacteria were injected into the hemocoels through the last left proleg using a 25 μL Hamilton syringe (Hamilton, Shanghai, People's Republic of China). Larvae were incubated at 37°C and were observed daily for 4 days.
Sixteen larvae were infected with a lethal dose of S. flexneri GN120471 or GN120454 as described above. CIP (the steady-state plasma concentration 2.5 μg/mL) and FOS (the steady-state plasma concentration 150 μg/mL) were administered via 10 μL injections for 4 days into the last right proleg within 2 hours of inoculation of bacterial suspensions. The following doses were used: 100 μg/mL every 12 hours for CIP and 1,000 μg/mL every 6 hours for FOS. Sixteen mock-inoculated (sterile PBS) larvae were used as controls. The larvae were observed for survival every 24 hours for 4 days.
Statistical analysis
Data were analyzed using SPSS, version 16.0 (SPSS, Chicago, IL, USA). Univariate analysis was performed by the chi-squared test or Fisher’s exact test when appropriate. P-values were based on two-tailed test results, and P-values <0.05 were considered statistically significant.
Results
FICI of CIP/FOS combination treatment
The MICs of CIP and/or FOS against 80 CIPR S. flexneri are shown in Table 1. The CIP/FOS combination showed a synergistic effect in 31 (38.75%) strains and no interaction in 49 (61.25%) strains, while no antagonism was observed in any strains.
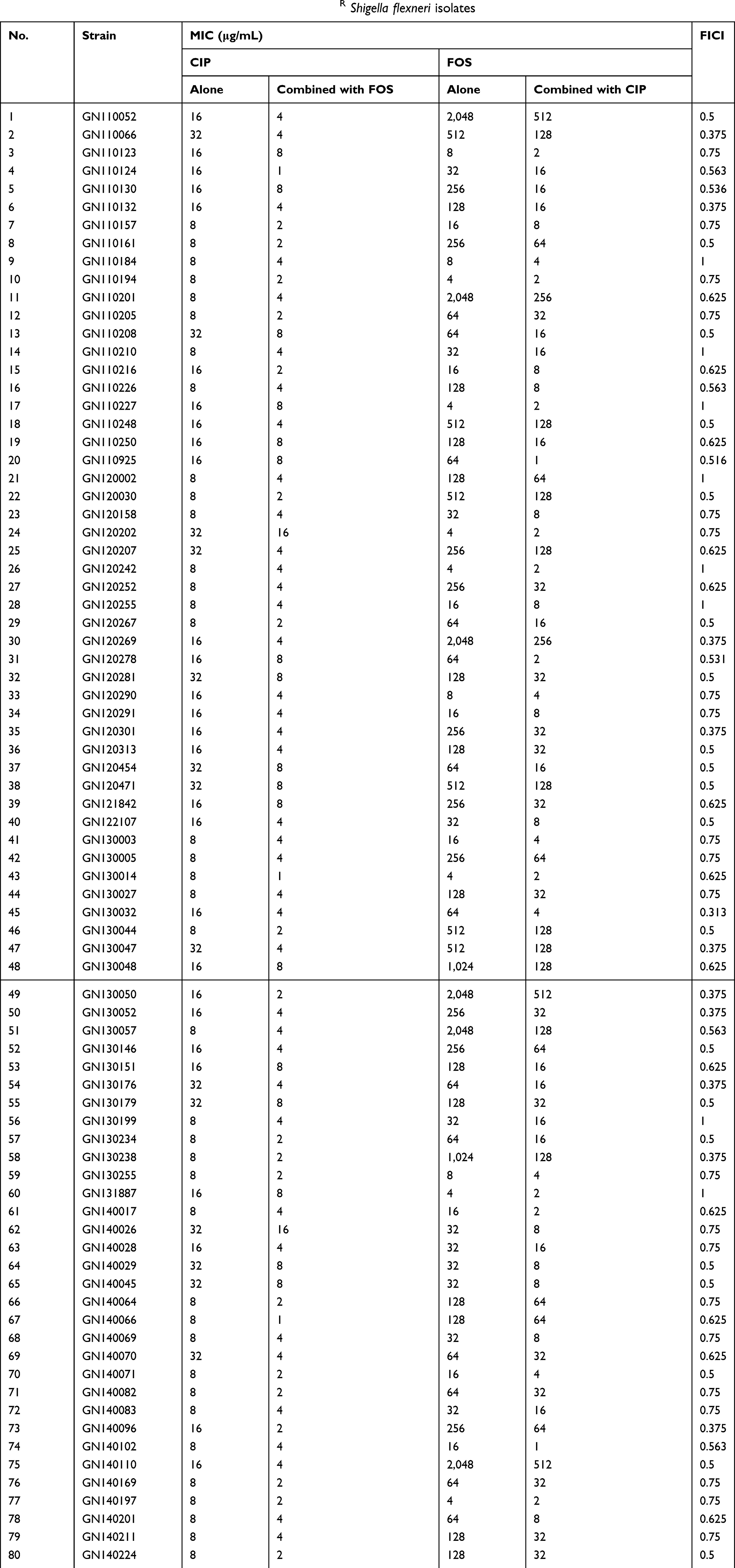 | Table 1 MICs of ciprofloxacin and/or fosfomycin against CIPR Shigella flexneri isolates |
Time-kill assays
The complete time-kill data for GN120471 and GN120454 and concentrations (monotherapy and combination therapy) are presented in Figure 1. Log changes in viable cell counts with monotherapy and combination therapy are shown in Table 2.
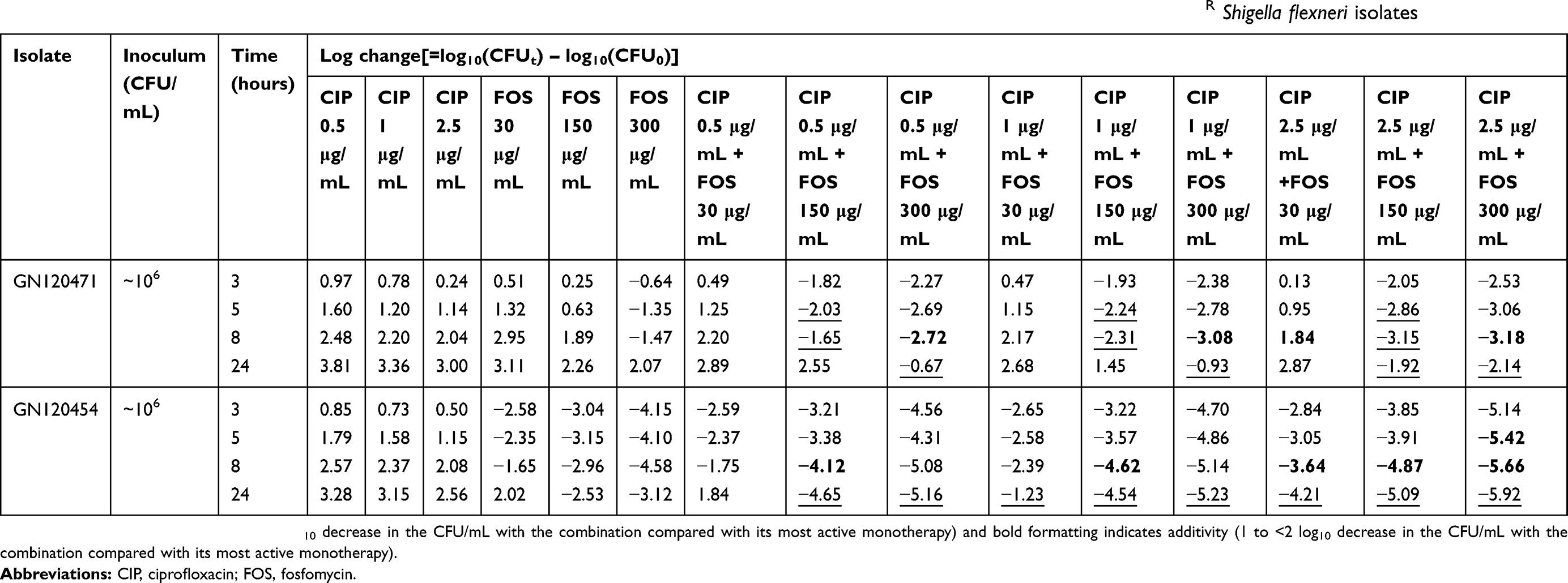 | Table 2 Log changes at 3, 5, 8, and 24 hours with various clinically relevant concentrations of ciprofloxacin and/or fosfomycin against CIPR Shigella flexneri isolates |
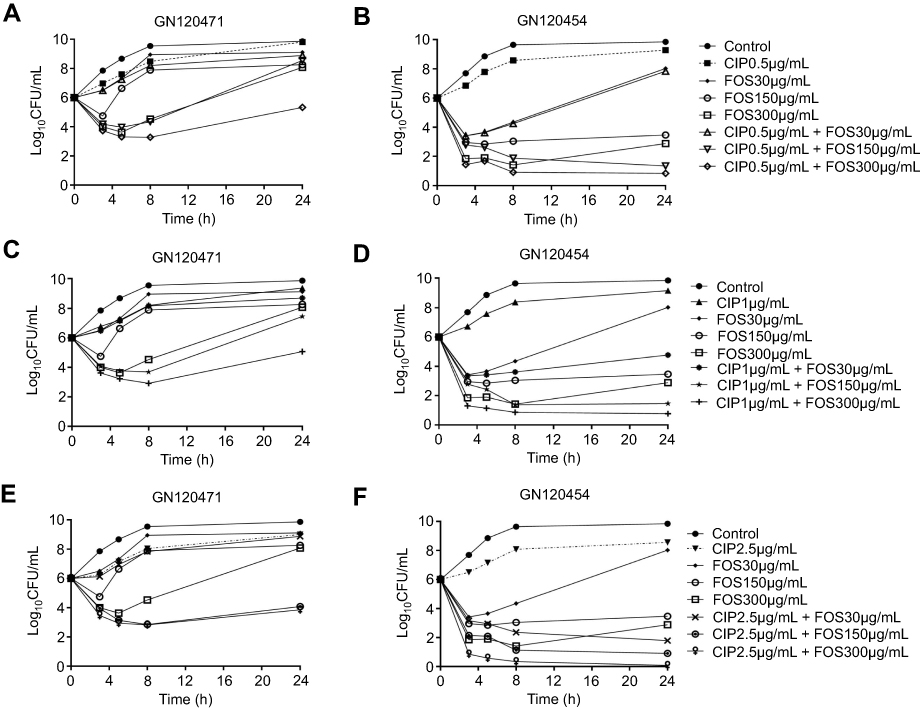 | Figure 1 Representative time-kill curves with low, medium, and high concentrations of ciprofloxacin and fosfomycin alone and in combination against CIPR Shigella flexneri isolates. (A) CIP (0.5 μg/mL) and FOS (30, 150, 300 μg/mL) alone and in combination against GN120471 (CIPRFOSR); (B) CIP (0.5 μg/mL) and FOS (30, 150, 300 μg/mL) alone and in combination against GN120454 (CIPRFOSS); (C) CIP (1 μg/mL) and FOS (30, 150, 300 μg/mL) alone and in combination against GN120471 (CIPRFOSR); (D) CIP (1 μg/mL) and FOS (30, 150, 300 μg/mL) alone and in combination against GN120454 (CIPRFOSS); (E) CIP (2.5 μg/mL) and FOS (30, 150, 300 μg/mL) alone and in combination against GN120471 (CIPRFOSR); (F) CIP (2.5 μg/mL) and FOS (30, 150, 300 μg/mL) alone and in combination against GN120454 (CIPRFOSS). Abbreviations: CIP, ciprofloxacin; FOS, fosfomycin; R, resistance; S, susceptibility. |
For the isolate GN120471 (CIPRFOSR), monotherapy with CIP or FOS at all concentrations produced little or no bacterial killing (Figure 1A, C and E). The CIP/FOS combination produced enhanced bacterial killing with FOS concentrations of 150 and 300 μg/mL, especially when combined with CIP at 2.5 μg/mL (Figure 1E). With these two combinations (CIP at 2.5 μg/mL plus FOS at 150 or 300 μg/mL), >5 log10 CFU/mL additional killing was achieved over equivalent monotherapy at 24 hours (Table 2). For this isolate, the combination was additive or synergistic in four (11.1%) and 10 (27.8%) of 36 cases (nine combinations at four time-points: 3, 5, 8, and 24 hours), respectively. Furthermore, all combinations containing FOS at 150 or 300 μg/mL at some time-points were additive or synergistic, with enhanced killing primarily across the initial 8 hours of therapy (Table 2 and Figure 1A, C and E). At 24 hours, regrowth was observed with all combinations.
For the isolate GN120454 (CIPRFOSS), FOS at the different concentrations produced varying degrees of initial killing, and CIP at all concentrations produced little or no bacterial killing (Figure 1B, D and F). The CIP/FOS combination at all concentrations produced more rapid and extensive killing (up to 5 log10 CFU/mL with many combinations) than with either antibiotic alone, especially when combined with FOS at 150 or 300 μg/mL (Table 2 and Figure 1B, D and F). For this isolate, the combination was additive or synergistic in six (16.7%) and eight (22.2%) of 36 cases, respectively (Table 2). Furthermore, all combinations at 24 hours were synergistic, except for CIP at 0.5 μg/mL plus FOS at 30 μg/mL.
Activities of CIP and FOS in the G. mellonella infection model
The concentration of bacteria was approximately 104 CFU in the body that could cause approximately 80% of the larvae to die at 96 hours for GN120471 (Figure 2A) and GN120454 (Figure 2B) (bacterial suspension concentration for ~106 CFU/mL). Based on these data, 1×104 CFU/larva was selected as the inoculum for the subsequent treatment experiments. According to the above research results, the following antimicrobial concentrations were used, simulating human doses: CIP (the steady-state plasma concentration 2.5 μg/mL) and FOS (the steady-state plasma concentration 150 μg/mL). The survival rate for the CIP/FOS combination against GN120471-infected larvae was 68.75% compared with 25% for CIP (χ2=6.149, P=0.013) and 43.75% for FOS (χ2=2.032, P=0.154) (Figure 3A). The survival rate for the CIP/FOS combination against GN120454-infected larvae was 81.25% compared with 37.5% for CIP (χ2= 6.348, P=0.012) and 62.5% for FOS (P>0.05) (Figure 3B).
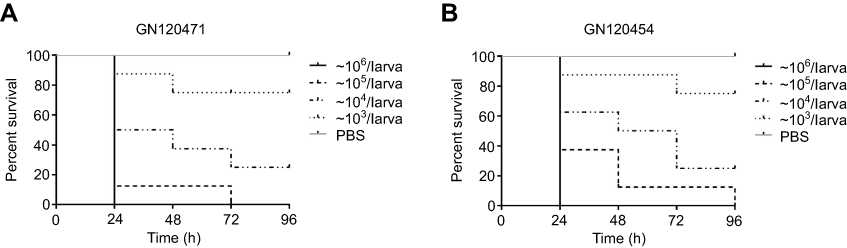 | Figure 2 Survival curves of various inoculum doses of Shigella flexneri GN120471 (A) or GN120454 (B) in Galleria mellonella larvae during incubation at 37°C for 96 hours. Curves were plotted using 10 larvae for every experiment. Larvae were inoculated with 10 μL of bacterial suspensions. Mortality at 24 hours was 100% at 106 CFU/larva (bacterial suspension concentration for ~108 CFU/mL). |
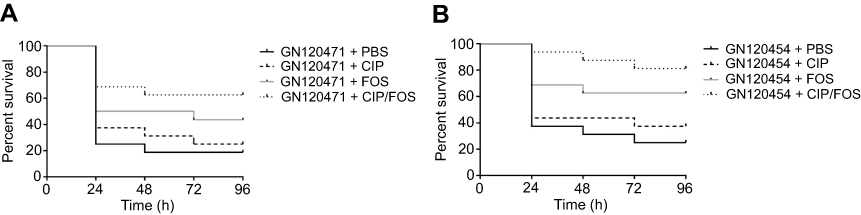 | Figure 3 Survival curves for Galleria mellonella larvae by CIP (steady-state plasma concentration 2.5 μg/mL) and FOS (steady-state plasma concentration 150 μg/mL) alone and in combination against Shigella flexneriisolates GN120471 (A) or GN120454 (B). Curves were plotted using 16 larvae for every experiment during incubation at 37°C for 96 hours. Bacterial suspension concentration for 1×106 CFU/mL (1×104 CFU/larva) was selected as the inoculum for the treatment experiments. Univariate analysis was performed by the chi-squared test or Fisher’s exact test when appropriate. The survival rate for CIP/FOS combination against GN120471-infected larvae was 68.75% compared with 25% for CIP (χ2=6.149, P=0.013) and 43.75% for FOS (χ2=2.032, P=0.154). The survival rate for CIP/FOS combination against GN120454-infected larvae was 81.25% compared with 37.5% for CIP (χ2=6.348, P=0.012) and 62.5% for FOS (P>0.05). Abbreviations: CIP, ciprofloxacin; FOS, fosfomycin. |
Discussion
With the increased resistance to CIP, unorthodox combination therapies are increasingly being considered against CIPR Shigella infections. FOS, an old antibiotic, is increasingly used for treatment of infections due to MDR organisms.23,24 Some studies have shown that the CIP/FOS combination had potent synergy effect in vitro against CIPR P. aeruginosa.17,18 Although this combination appears to be a promising treatment option based on in vitro data, further preclinical work is clearly needed before it can be considered for clinical use. Therefore, we systematically investigated the effectiveness of the CIP/FOS combination in vitro and in vivo against CIPR S. flexneri isolates.
To the best of our knowledge, this study is the first to confirm that the CIP/FOS combination enhanced in vitro and in vivo activity against CIPR S. flexneri isolates. This may be due to enhancement of FOS cell-wall permeability, which could enhance the activity of CIP. In checkerboard assays, our results provided evidence that CIP possessed a considerable synergy with FOS against CIPR S. flexneri isolates. In time-kill assays, the concentrations of CIP and FOS represented low, medium, and high average steady-state plasma concentrations typically achieved in patients receiving standard doses.25,26 With the majority of combinations, the addition of FOS to CIP generally resulted in substantial improvements in bacterial killing (up to 5 log10 CFU/mL) over 24 hours for CIPR S. flexneri isolates, especially when CIP at 2.5 μg/mL was combined with FOS at 150 or 300 μg/mL. Moreover, all combinations of CIP/FOS at some time-points were synergistic in vitro for CIPR S. flexneri isolates, with the exception of combinations containing CIP at all concentrations plus FOS at 30 μg/mL for CIPR S. flexneri isolate. This indicated that substantial improvements in bacterial killing were possible with the higher concentrations of the CIP/FOS combination against CIPR S. flexneri isolates.
Larvae of G. mellonella have been used as an alternative to study a number of important human pathogens.27–30 Barnoy et al31 initially developed a G. mellonella model to investigate the pathogenicity of Shigella infection and to show that S. flexneri reside within a vacuole of the insect hemocytes that ultrastructurally resemble vacuoles, with mouse and human macrophage cell lines. We applied this model to study the in vivo activities of the CIP/FOS combination against CIPR S. flexneri isolates. In this study, mortality at 96 hours was around 80% at approximately 104 CFU/larva for GN120471 and GN120454 (bacterial suspension concentration for ~106 CFU/mL). The CIP/FOS combination (CIP 2.5 μg/mL and FOS 150 μg/mL) was significantly more effective than CIP monotherapy (P<0.05) in protecting larvae from lethal infections with CIPR S. flexneri isolates, which was consistent with the in vitro results of the CIP/FOS combination against CIPR S. flexneri isolates. An interesting finding was that the CIP/FOS combination in the larvae model showed no significant difference compared to FOS monotherapy (P>0.05), despite its performance in vitro being better than any drug individually. It was speculated that this phenomenon might result from immunomodulatory activities of FOS in G. mellonella larvae because FOS exerted immunomodulatory effects by altering lymphocyte, monocyte, and neutrophil function.32 It is possible that certain immunological effects augment the antibacterial activity of FOS, leading to better efficacy than observed in vitro. Furthermore, although preliminary evidence of in vivo efficacy has been obtained through the use of invertebrate infection models, additional studies using mammalian infection models are required.
Conclusion
Our in vitro and in vivo results provide clues to understanding the synergistic effect of CIP combined with FOS and to developing more effective therapy against CIPR S. flexneri isolates. One possible mechanism for the enhanced bacterial killing is through the increased uptake of one antibiotic by the other. Further studies are required to clarify the underlying mechanism of their synergistic effect.
Acknowledgment
This work was supported by grant 81673242 from the National Natural Science Foundation of China.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Livio S, Strockbine NA, Panchalingam S, et al. Shigella isolates from the global enteric multicenter study inform vaccine development. Clin Infect Dis. 2014;59(7):933–941. doi:10.1093/cid/ciu468
2. Zhang W, Luo Y, Li J, et al. Wide dissemination of multidrug-resistant Shigella isolates in China. J Antimicrob Chemother. 2011;66(11):2527–2535. doi:10.1093/jac/dkr341
3. Yang H, Chen G, Zhu Y, et al. Surveillance of antimicrobial susceptibility patterns among Shigella species isolated in China during the 7-year period of 2005–2011. Ann Lab Med. 2013;33(2):111–115. doi:10.3343/alm.2013.33.2.111
4. Troeger C, Forouzanfar M, Rao PC, et al.Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: a systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect Dis. 2017;17(9):909–948. doi:10.1016/S1473-3099(17)30276-1
5. Falagas ME, Maraki S, Karageorgopoulos DE, Kastoris AC, Mavromanolakis E, Samonis G. Antimicrobial susceptibility of multidrug-resistant (MDR) and extensively drug-resistant (XDR) Enterobacteriaceae isolates to fosfomycin. Int J Antimicrob Agents. 2010;35(3):240–243. doi:10.1016/j.ijantimicag.2009.10.019
6. Falagas ME, Kastoris AC, Kapaskelis AM, Karageorgopoulos DE. Fosfomycin for the treatment of multidrug-resistant, including extended-spectrum beta-lactamase producing, Enterobacteriaceae infections: a systematic review. Lancet Infect Dis. 2010;10(1):43–50. doi:10.1016/S1473-3099(09)70325-1
7. Samonis G, Maraki S, Karageorgopoulos DE, Vouloumanou EK, Falagas ME. Synergy of fosfomycin with carbapenems, colistin, netilmicin, and tigecycline against multidrug-resistant Klebsiella pneumoniae, Escherichia coli, and Pseudomonas aeruginosa clinical isolates. Eur J Clin Microbiol Infect Dis. 2012;31(5):695–701. doi:10.1007/s10096-011-1360-5
8. Hendlin D, Stapley EO, Jackson M, et al. Phosphonomycin, a new antibiotic produced by strains of streptomyces. Science. 1969;166(3901):122–123.
9. Kahan FM, Kahan JS, Cassidy PJ, Kropp H. The mechanism of action of fosfomycin (phosphonomycin). Ann N Y Acad Sci. 1974;235:364–386.
10. Reeves DS. Fosfomycin trometamol. J Antimicrob Chemother. 1994;34(6):853–858.
11. Popovic M, Steinort D, Pillai S, Joukhadar C. Fosfomycin: an old, new friend? Eur J Clin Microbiol Infect Dis. 2010;29(2):127–142. doi:10.1007/s10096-009-0833-2
12. Falagas ME, Kanellopoulou MD, Karageorgopoulos DE, et al. Antimicrobial susceptibility of multidrug-resistant Gram negative bacteria to fosfomycin. Eur J Clin Microbiol Infect Dis. 2008;27(6):439–443. doi:10.1007/s10096-007-0456-4
13. Florent A, Chichmanian RM, Cua E, Pulcini C. Adverse events associated with intravenous fosfomycin. Int J Antimicrob Agents. 2011;37(1):82–83. doi:10.1016/j.ijantimicag.2010.09.002
14. Karageorgopoulos DE, Wang R, Yu XH, Falagas ME. Fosfomycin: evaluation of the published evidence on the emergence of antimicrobial resistance in Gram-negative pathogens. J Antimicrob Chemother. 2012;67(2):255–268. doi:10.1093/jac/dkr466
15. Walsh CC, McIntosh MP, Peleg AY, Kirkpatrick CM, Bergen PJ. In vitro pharmacodynamics of fosfomycin against clinical isolates of Pseudomonas aeruginosa. J Antimicrob Chemother. 2015;70(11):3042–3050. doi:10.1093/jac/dkv221
16. Pogue JM, Marchaim D, Kaye D, Kaye KS. Revisiting “older” antimicrobials in the era of multidrug resistance. Pharmacotherapy. 2011;31(9):912–921. doi:10.1592/phco.31.9.912
17. Walsh CC, Landersdorfer CB, McIntosh MP, et al. Clinically relevant concentrations of fosfomycin combined with polymyxin B, tobramycin or ciprofloxacin enhance bacterial killing of Pseudomonas aeruginosa, but do not suppress the emergence of fosfomycin resistance. J Antimicrob Chemother. 2016;71(8):2218–2229. doi:10.1093/jac/dkw115
18. Yamada S, Hyo Y, Ohmori S, Ohuchi M. Role of ciprofloxacin in its synergistic effect with fosfomycin on drug-resistant strains of Pseudomonas aeruginosa. Chemotherapy. 2007;53(3):202–209. doi:10.1159/000100811
19. Hornsey M, Phee L, Longshaw C, Wareham DW. In vivo efficacy of telavancin/colistin combination therapy in a Galleria mellonella model of Acinetobacter baumannii infection. Int J Antimicrob Agents. 2013;41(3):285–287. doi:10.1016/j.ijantimicag.2012.11.013
20. Yang H, Chen G, Hu L, et al. In vivo activity of daptomycin/colistin combination therapy in a Galleria mellonella model of Acinetobacter baumannii infection. Int J Antimicrob Agents. 2015;45(2):188–191. doi:10.1016/j.ijantimicag.2014.10.012
21. Yang H, Chen G, Hu L, et al. Enhanced efficacy of imipenem-colistin combination therapy against multiple-drug-resistant Enterobacter cloacae: in vitro activity and a Galleria mellonella model. J Microbiol Immunol Infect. 2018;51(1):70–75. doi:10.1016/j.jmii.2016.01.003
22. Odds FC. Synergy, antagonism, and what the chequerboard puts between them. J Antimicrob Chemother. 2003;52(1):1. doi:10.1093/jac/dkg486
23. Theuretzbacher U, Van Bambeke F, Canton R, et al. Reviving old antibiotics. J Antimicrob Chemother. 2015;70(8):2177–2181. doi:10.1093/jac/dkv157
24. Pontikis K, Karaiskos I, Bastani S, et al. Outcomes of critically ill intensive care unit patients treated with fosfomycin for infections due to pandrug-resistant and extensively drug-resistant carbapenemase-producing Gram-negative bacteria. Int J Antimicrob Agents. 2014;43(1):52–59. doi:10.1016/j.ijantimicag.2013.09.010
25. Traunmuller F, Popovic M, Konz KH, Vavken P, Leithner A, Joukhadar C. A reappraisal of current dosing strategies for intravenous fosfomycin in children and neonates. Clin Pharmacokinet. 2011;50(8):493–503. doi:10.2165/11592670-000000000-00000
26. Gotfried MH, Danziger LH, Rodvold KA. Steady-state plasma and intrapulmonary concentrations of levofloxacin and ciprofloxacin in healthy adult subjects. Chest. 2001;119(4):1114–1122.
27. Peleg AY, Jara S, Monga D, Eliopoulos GM, Moellering RJ, Mylonakis E. Galleria mellonella as a model system to study Acinetobacter baumannii pathogenesis and therapeutics. Antimicrob Agents Chemother. 2009;53(6):2605–2609. doi:10.1128/AAC.01533-08
28. Wei WJ, Yang HF, Ye Y, Li JB. Galleria mellonella as a model system to assess the efficacy of antimicrobial agents against Klebsiella pneumoniae infection. J Chemother. 2017;29(4):252–256. doi:10.1080/1120009X.2016.1156892
29. Jonsson R, Struve C, Jenssen H, Krogfelt KA. The wax moth Galleria mellonella as a novel model system to study Enteroaggregative Escherichia coli pathogenesis. Virulence. 2017;8(8):1894–1899. doi:10.1080/21505594.2016.1256537
30. Thomas RJ, Hamblin KA, Armstrong SJ, et al. Galleria mellonella as a model system to test the pharmacokinetics and efficacy of antibiotics against Burkholderia pseudomallei. Int J Antimicrob Agents. 2013;41(4):330–336. doi:10.1016/j.ijantimicag.2012.12.009
31. Barnoy S, Gancz H, Zhu Y, Honnold CL, Zurawski DV, Venkatesan MM. The Galleria mellonella larvae as an in vivo model for evaluation of Shigella virulence. Gut Microbes. 2017;8(4):335–350. doi:10.1080/19490976.2017.1293225
32. Falagas ME, Vouloumanou EK, Samonis G, Vardakas KZ. Fosfomycin. Clin Microbiol Rev. 2016;29(2):321–347. doi:10.1128/CMR.00068-15
Supplementary material
 | Figure S1 Larvae infected with Shigella flexneri (103, 105, 106, and 107 CFU/larva) at 96 hours. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.