")
Back to Journals » Journal of Inflammation Research » Volume 16
Impact of Toll-Like Receptor 2 and 9 Gene Polymorphisms on COVID-19: Susceptibility, Severity, and Thrombosis
Authors Alhabibi AM , Hassan AS , Abd Elbaky NM, Eid HA , Khalifa MAAA, Wahab MA , Althoqapy AA, Abdou AE, Zakaria DM, Nassef EM , Kasim SA , Saleh OI, Elsheikh AA , Lotfy M, Sayed A
Received 31 October 2022
Accepted for publication 26 January 2023
Published 17 February 2023 Volume 2023:16 Pages 665—675
DOI https://doi.org/10.2147/JIR.S394927
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ning Quan
Alshaymaa M Alhabibi,1 Asmaa S Hassan,1 Nashwa Mohamed Abd Elbaky,1 Hoda Asaad Eid,2 Mohie Aldeen Abd Alzaher Khalifa,3 Maisa A Wahab,4 Azza Ali Althoqapy,5 Aml E Abdou,5 Doaa Mohammed Zakaria,6 Eman Mostafa Nassef,6 Sammar Ahmed Kasim,6 Ola I Saleh,7 Asmaa Abdelghany Elsheikh,8 Mahmoud Lotfy,9 Alaa Sayed10
1Departments of Clinical Pathology, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt; 2Chest Disease, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt; 3Department of Chest Disease, General Organization for Teaching Hospitals and institutes, Cairo, Egypt; 4Vascular Surgery, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt; 5Medical Microbiology and Immunology Department, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt; 6Internal Medicine, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt; 7Radio-Diagnosis, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt; 8Community and Occupational Medicine, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt; 9Molecular Biology Department, Genetic Engineering & Biotechnology Research Institute, University of Sadat City, Sadat City, Minufiya, Egypt; 10Hormones Department, Medical Research and Clinical Studies Institute, National Research Centre, Cairo, Egypt
Correspondence: Alshaymaa M Alhabibi, Tel +201002894075, Email [email protected]
Background: Toll-like receptors (TLRs) play an important role in activation of innate and adaptive immune responses.
Aim: We aimed to detect the association between TLR2 rs5743708 G>A and TLR9 rs5743836 C>T variants and COVID-19 disease susceptibility, severity, and thrombosis by using neutrophil extracellular traps (NETs).
Subjects and Methods: We included 100 adult COVID-19 patients as well as 100 age- and gender-matched normal controls. Participants were genotyped for TLR2 rs5743708 and TLR9 rs5743836. Citrullinated Histone (H3) was detected as an indicator of NETs.
Results: The mutant (G/A and C/C) genotypes and (A and C) alleles of TLR2 rs5743708 and TLR9 rs5743836, respectively, have been significantly related to a higher risk of COVID-19 infection, representing a significant risk factor for the severity of COVID-19. There was no significant association between the two variants and citrullinated histone (H3).
Conclusion: TLR2 rs5743708 and TLR9 rs5743836 variants have been significantly related to a higher risk and severity of COVID-19 infection but had no effect on thrombus formation.
Keywords: coronavirus disease 2019, single-nucleotide polymorphism, toll-like receptor, high resolution-CT, susceptibility, severity
Introduction
The WHO Director General proclaimed the COVID-19 (coronavirus disease 2019) pandemic, caused by the new severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), a global health emergency of concern worldwide in January 2020. SARS-CoV-2 is an enveloped, positive-sense single-stranded RNA (ssRNA) virus.1
The immune system protects the host against foreign pathogens. Adaptive immune responses are primed by innate immunological reactions, which are the body’s first line of defence versus invaders. Toll-like receptors (TLRs) as well as retinoic acid-inducible gene-I-like receptors, which recognise pathogen-associated molecular patterns (PAMPs), are various pattern-recognition receptors expressed by innate immune cells.2
TLR2 is a surface receptor that identifies various ligands generated by bacteria, viruses, fungi, and parasites and is implicated in sensing PAMPs via SARS-CoV-2.3 Likewise, TLR9 identifies CpG-rich fragments of DNA produced by viruses, bacteria, and mitochondrial DNA.4
According to Zheng et al,5 TLR2 expression was found to be increased with COVID-19 severity, and El Kebir et al6 suggested number of mechanisms explaining the link between stimulation of TLR2 and the coagulation system in COVID-19. Similarly, TLR9 was proposed as an important factor explaining hyper-inflammation as well as multi-organ and thrombotic complications in COVID-19 patients.7
Infection-related neutrophil extracellular traps (NETs), which form a scaffold and stimulate adhesion of platelets and formation of thrombus, represent a component of the innate immunity response.8 Interestingly, in COVID-19, NETs were suggested to lead to organ damage and death.9 Mitochondrial DNA, which activates polymorphonuclear leukocytes via TLR9, is a strong inducer of NETs. A TLR9 antagonist was found to completely block the formation of mitochondrial DNA-induced NETs.10 Additionally, previous studies have demonstrated that various types of viruses are involved in NETosis through the activation of platelet receptors (C-type lectin), enhancing NETosis, and production of pro-inflammatory cytokines via TLR2.11–13
Even in the absence of apparent hazard variables like elderly age, high BMI, and associated morbidities, disease development may happen quickly in some patients. As a result, it seems likely that additional variables, including the genetic makeup heterogeneity and the host’s various immune responses, may contribute to disease severity.14 Interestingly, recent studies found that there is upregulation of some genes (as ACE2, CD147, FURIN and TMPRSS1) and down regulation of others (as BMAL1) in SARS-CoV-2 infection and periodontitis conditions.15
Individuals exhibit a wide variety of susceptibilities to infections due to genetic changes like single nucleotide polymorphisms (SNPs), which strongly affect innate immune responses to pathogen challenges and illness outcomes. As a result, some people are susceptible to specific diseases, whereas others are protected.16
Thus, it is crucial to pinpoint the genes involved in the incidence and severity of COVID-19 cases to develop preventive measures for individuals who are predisposed to disease and complications.17
We therefore aimed to detect the connection between these variants of TLR2 and TLR9 and COVID-19 disease susceptibility, severity, and thrombosis. To our knowledge, no study has investigated these variants in COVID-19 disease in the population of Egypt.
Subjects and Methods
Selection of Participants
This case–control research enrolled 200 participants selected from Al-Zahraa University Hospital and categorized them into 2 groups: Group I (n = 100), consisting of individuals diagnosed with COVID-19 infection; and Group II (n = 100), consisting of healthy individuals with similar ages and sexes who served as controls. Group I was subdivided into two groups according to disease severity: group IA (n = 50), diagnosed with moderate to severe COVID-19 infection, and group IB (n = 50), diagnosed with mild COVID-19.
Calculation of Sample Size
The sample size was calculated by G*Power (version 3.1.9.2; Germany). Up to our knowledge, there are no previously published studies about association between TLR2 rs5743708, TLR9 rs5743836 SNP and COVID-19 disease, thus we conducted a pilot study on 20 COVID-19 patients and 20 healthy controls to calculate the sample size. Pilot study data revealed that the probability of exposure to A allele of rs5743708 TLR2 and C allele of rs5743836 TLR9 among controls was 7.5% and 10% respectively, probability of exposure among cases was 25% and 30% respectively, thus the minimum required sample size was 80 patients and 80 controls for TLR2 rs5743708 and 72 patients and 72 controls for TLR9 rs5743836 with a power of 80% and a level of significance of 5%. We add 20 individuals in each group for better statistical analysis.
Inclusion Criteria
This research included adult patients aged 20–60 years, diagnosed using SARS-COV-2 -reverse-transcriptase polymerase chain reaction (RT-PCR). Patients have been classified according to disease severity based on the interim guidance of the WHO18 and the CO-RADS classifications based on CT findings.19
Exclusion Criteria
Patients with known chronic lung disease, chronic liver or kidney disease, malignancies, autoimmune diseases, diabetes, hypertension, hypo/hyperthyroidism, and those receiving any medical treatment were excluded from this research. Children and pregnant females were also excluded from this study.
Ethical Consideration
The research has been carried out in accordance with the World Medical Association’s Helsinki Declaration for Human Subjects Studies. This study has been accepted by Al-Azhar University’s Institutional Review Board (IRB) (study code 1203, 12th. January 2022), and each patient gave their written informed consent.
Assessment and Procedures
All patients underwent a detailed history-assessment, thorough clinical and radiological evaluation, and the following investigations:
- Routine laboratory investigations:
- The automated haematology analyser Sysmex KX 21N (Kobe, Japan) was used to perform a complete blood count (CBC), coagulation profile using Stago STA (Diagnostica Stago, Paris, France), D-dimer using Immulite 1000 (Siemens Healthcare Diagnostics, New York, USA), and biochemical analyses using the Cobas c311 system (Roche Diagnostics, Mannheim, Germany) for kidney and liver function parameters; for ferritin, use the Cobas e411 system; and for C-reactive protein (CRP), use the Cobas Integra 400 plus.
- Human citrullinated histone H3 was measured by ELISA using an ELISA kit supplied by the Bioassay Technology Laboratory (lot No 202203013, Zhejiang, China) in accordance with the manufacturer ‘s directions. The assay had a sensitivity of 1.352 ng/mL with a range of 32–105 ng/mL.
- Molecular analyses of TLR2 rs5743708 and TLR9 rs5743836 variants:
Genomic DNA Extraction
A Thermo Scientific GeneJET Whole Blood Genomic DNA Purification Mini Kit (Lot. No.01176830) was used to extract DNA from blood specimens. DNA samples were tested for quality and concentration using a QIAxpert (QIAGEN, Hilden, Germany) and preserved at −20°C.
Genotyping of TLR2 rs5743708 and TLR9 rs5743836 by TaqMan Genotyping Assay
The PCR process consists of 25–50 cycles with the following steps: initial denaturation at 95°C to separate the nucleic acid double chain, annealing at 58°C for binding of primers, and extension by DNA polymerase at 72°C. The genotyping PCR reaction has been conducted in a 20 μL reaction volume having 10 uL of TaqMan Genotyping Master Mix (Lot. No. 01187540), 0.5 uL of TaqMan Genotyping Assay Mix, and 9.5 uL of diluted DNA template (5.5 μL nuclease free water, Lot No. 01069419, and 4 μL template). After amplification, data was collected and read based on fluorescence signals using a Rotor Gene real-time system (QIAGEN, Hilden, Germany). Every TaqMan® SNP Genotyping Assay contains sequence-specific forward and reverse primers to amplify the polymorphic sequence of interest and two TaqMan® minor groove binder (MGB) probes with non-fluorescent quenchers (NFQ): one VIC™ labelled probe to identify the allele 1 sequence and one FAM™ labelled probe to identify the allele 2 sequence. The TaqMan® SNP Genotyping Assay of TLR2 rs5743708 and TLR9 rs5743836 were obtained from Applied Biosystems by Thermo Scientific, USA (Lot No. P211222-005A10 and P211222-005A11respectively).
The allelic discrimination data were plotted as a comparison of allele 1 (VICTM dye) and allele 2 (FAMTM dye) using real-time PCR instrument software (QIAGEN, Hilden, Germany). Every specimen is represented as a separate point on the allelic discrimination (AD) plot, sometimes referred to as a cluster or scatter plot (Supplementary Data Figures 1–6).
(B) Radiological examination by high resolution – CT (HRCT) scan (Toshiba 160 multidetector CT scan) (Supplementary Data Figure 7).
Statistics
The Statistical Package for Social Sciences, Windows, version 21.0 (SPSS Inc., Armonk, NY, USA) has been employed to analyse the data. Normally distributed quantitative data has been represented as the mean and standard deviation. The Student’s t-test was used for comparison between two groups, whereas the Mann–Whitney test was for non-parametric data. Qualitative variables were presented as frequencies and percentages, and the Fisher's exact test or the χ2 test was employed to compare qualitative variables. TLR2 and TLR9 genetic variants were assessed for Hardy-Weinberg equilibrium (HWE) with a χ2 test before genotypic and allelic disease correlation analysis. The odds ratio (OR) of genotypes between groups was assessed using logistic regression. The level of statistical significance was fixed at P <0.05.
Results
Genotype and Allelic Frequencies of TLR2 rs5743708and TLR9 rs5743836 Variants in Patients with COVID-19 and the Control Group
In the control group, we found that TLR2 rs5743708 and TLR9 rs5743836 were in Hardy-Weinberg equilibrium (HWE) (p = 0.74 and 0.49, respectively), as indicated by the respective test.
Table 1 illustrates the association between TLR2 rs5743708 and TLR9 rs5743836 genotypes as well as the risk of COVID-19 infection. We observed that wild (G/G and T/T) genotypes have been the most prevalent genotypes in the studied population and have been related to a significantly lower risk of COVID-19 infection. We detected the mutant homozygous (A/A) genotype in 3% of patients with COVID-19, whereas the mutant homozygous (C/C) genotype was detected in 16% of COVID-19 patients and 6% of the control group, representing a significant risk factor for COVID-19 infection (p = 0.02). We found that patients with the (G/A) genotype of TLR2 rs5743708 were 3.7 times more at an increased risk of being infected with COVID-19 than those with wild homozygous genotypes. Moreover, we detected that the (A and C) alleles of TLR2 rs5743708 and TLR9 rs5743836, respectively, have been related to a significantly increased risk of COVID-19 infection.
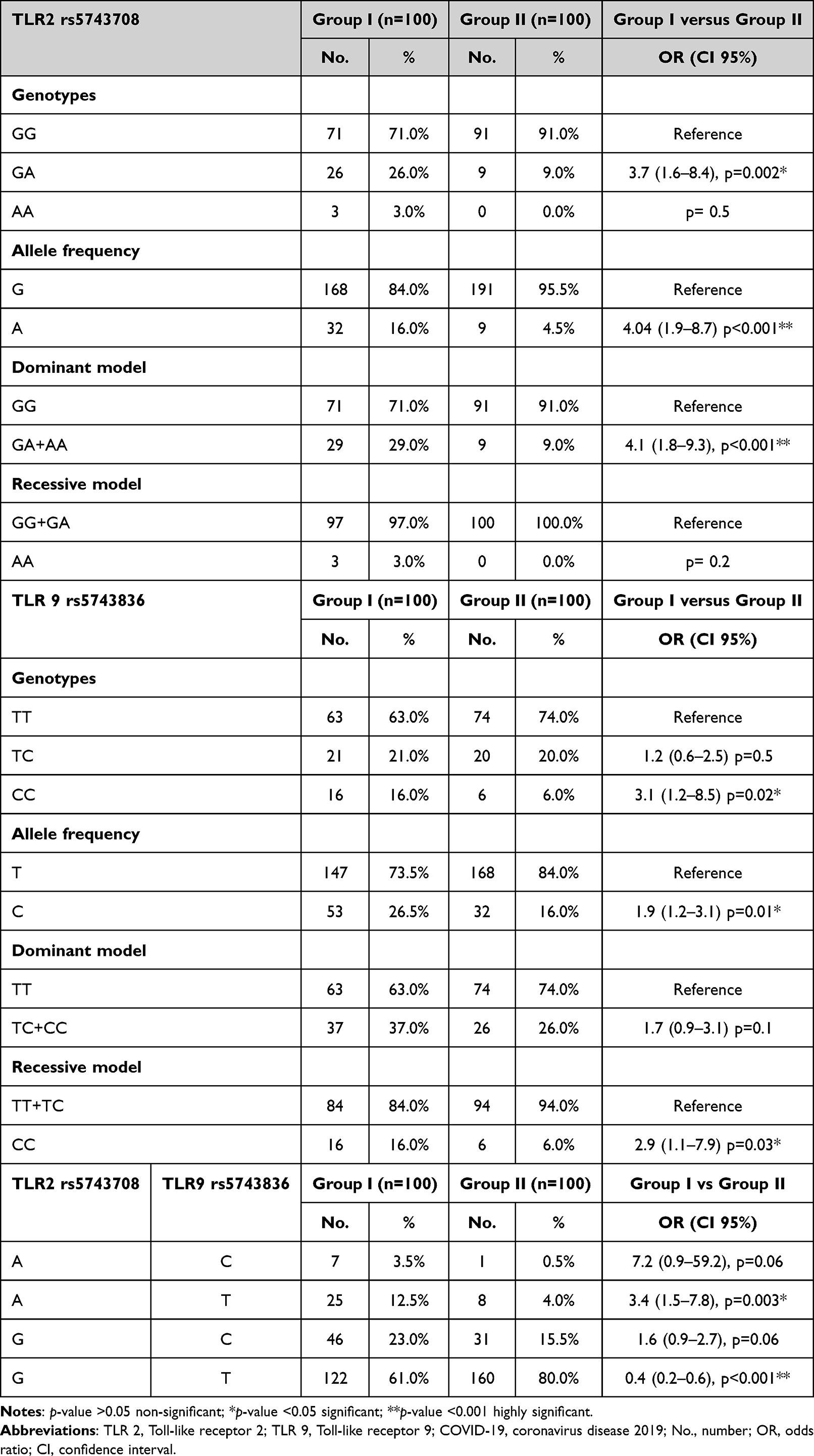 |
Table 1 The Association Between TLR2 rs5743708 and TLR9 rs5743836 Genotypes and the Risk for COVID-19 Infection |
The association of different models of inheritance for TLR9 rs5743836 revealed that the recessive model in which patients with the mutant (C/C) genotype of TLR9 were linked to a significantly increased likelihood of COVID-19 infection was the best inheritance model.
Regarding haplotype analysis, we observed that the frequency of the (A-T) haplotype was linked to an increased risk of COVID-19 infection. In contrast, the frequency of the (G-T) haplotype has been linked to a reduced risk of COVID-19 infection.
Genotype and Allelic Frequencies of TLR2 rs5743708 and TLR9 rs5743836 Variants and Disease Severity
Table 2 shows the association between TLR2 rs5743708 and TLR9 rs5743836 genotypes as well as the likelihood of severe COVID-19 infection. We detected the mutant homozygous (A/A) genotype in 6% of Group IA patients (moderate to severe COVID-19), whereas the mutant homozygous (C/C) genotype was detected in 26% of Group IA patients with COVID-19 and 6% of Group IB patients (mild COVID-19), representing a 7-times elevated risk of severe COVID-19 infection (p < 0.05). We further observed that the presence of (A and C) alleles of TLR2 rs5743708 and TLR9 rs5743836, respectively, were risk factors for severe infection of COVID-19. In particular, we found that the (G/A) genotype of TLR2 was at a 2.6-times elevated risk of moderate to severe infection of COVID-19 than the wild homozygous genotypes.
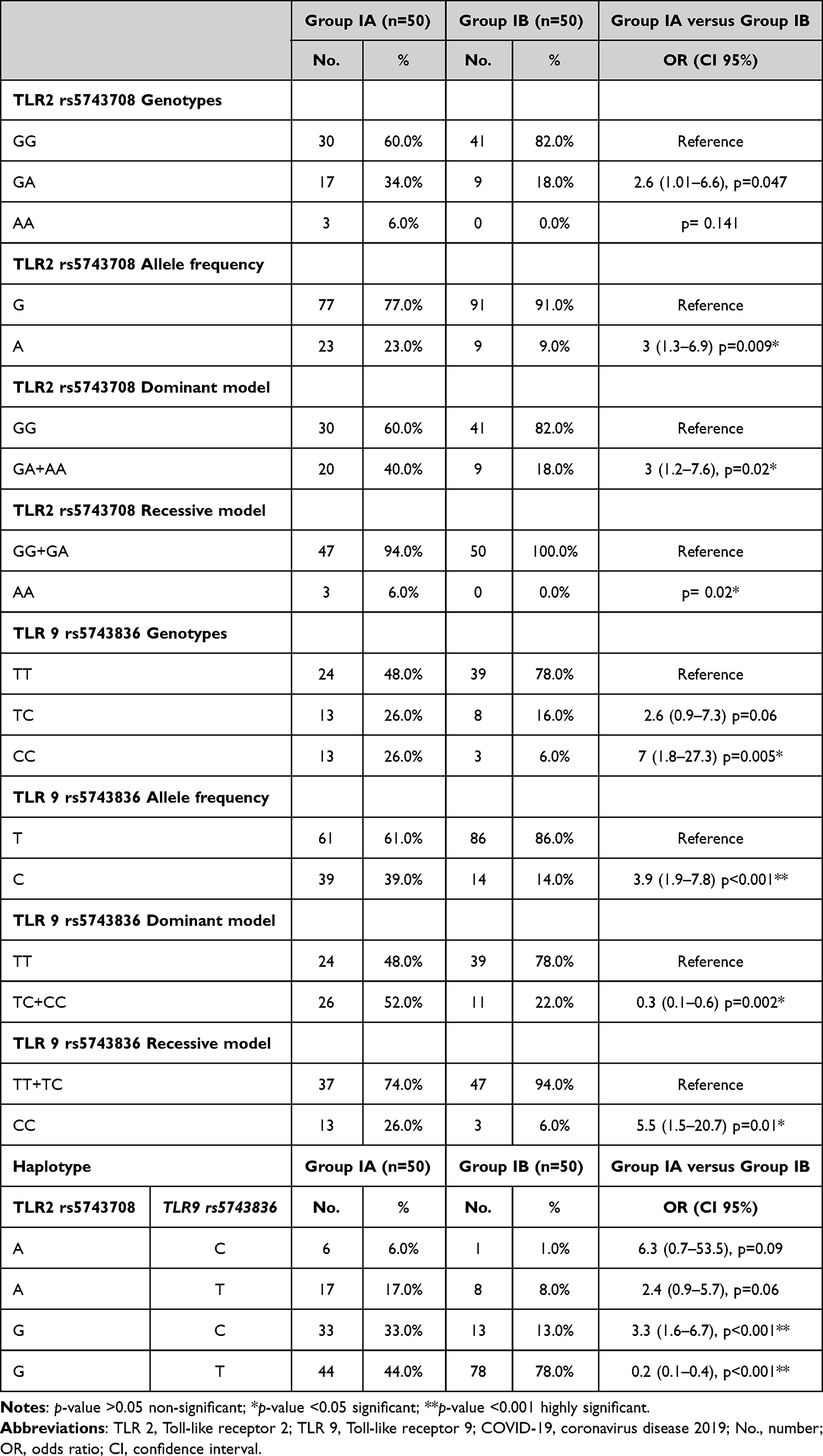 |
Table 2 The Association Between TLR2 rs5743708 and TLR9 rs5743836 Genotypes and the Risk for Severity of COVID-19 Infection |
The association of various inheritance models for TLR2 rs5743708 and TLR9 rs5743836 revealed that the recessive model, in which patients with mutant (A/A) and (C/C) TLR2 and TLR9 genotypes, respectively, were linked to a significantly increased risk of moderate to severe COVID-19 infection, was the best inheritance model.
Regarding haplotype analysis, we noticed that the frequency of the (G-C) haplotype was strongly linked to an increased chance of developing a severe COVID-19 infection. In contrast, the frequency of the (G-T) haplotype was linked to a lower chance of severe COVID-19 infection.
Association Between TLR2 rs5743708 and TLR9 rs5743836 Genotypes and Laboratory Characteristics
We discovered a statistically significant relation among different TLR2 genotypes and erythrocyte sedimentation rate (ESR) in Group IA (p = 0.026), and between different TLR2 genotypes and O2 saturation and serum alanine transaminase (ALT) in Group IB (p = 0.03 and p = 0.048, respectively).
With regard to TLR9, we detected a statistically significant relation among various TLR9 genotypes, haemoglobin and total protein (p = 0.024 and p = 0.029, respectively) in Group IA, and between different genotypes of TLR9 and serum aspartate transaminase (AST) in Group IB (p = 0.032). However, we detected no link between various TLR2 genotypes and any of the coagulation indicators. Likewise, there has been no significant association among various TLR9 genotypes and any of the coagulation indicators, except for a highly significant association with D-dimer (p = 0.006).
Association Between TLR2 rs5743708 and TLR9 rs5743836 Genotypes and Citrullinated Histone (H3)
Table 3 illustrates the association between different TLR2 rs5743708 and TLR9 rs5743836 genotypes and citrullinated histone (H3). We discovered no link between various TLR2 and TLR9 genotypes and citrullinated histones.
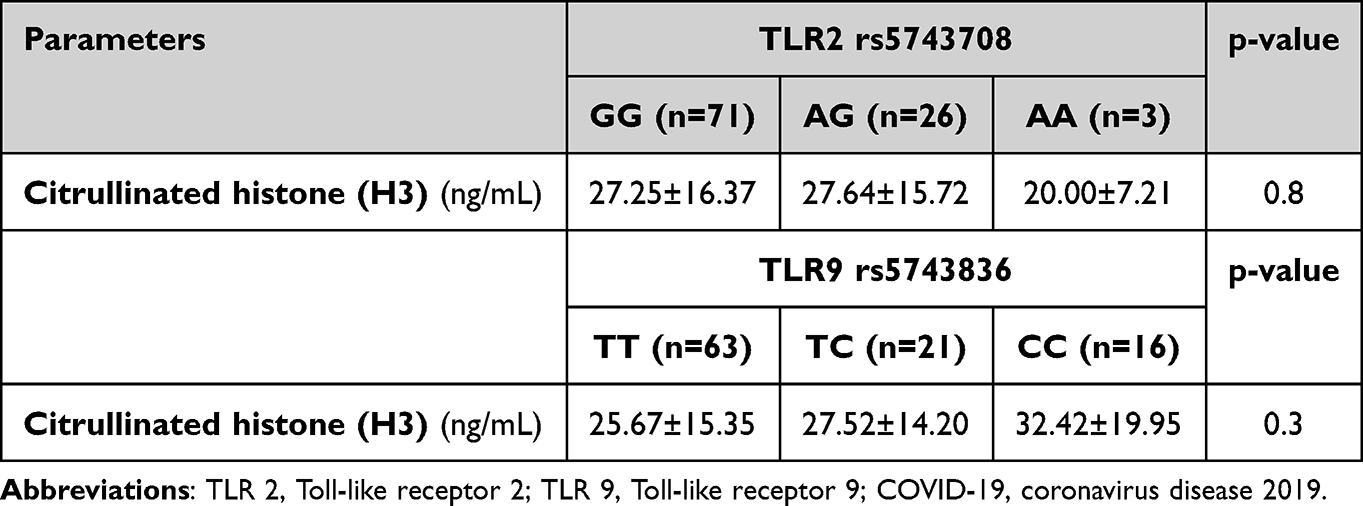 |
Table 3 The Association of Different TLR2 rs5743708 and TLR9 rs5743836 Genotypes and Citrullinated Histone |
Analysis of Linkage Disequilibrium
The analysis of allelic variants (rs5743708 and rs5743836) showed no evidence of linkage disequilibrium (X²=0.062 and p > 0.05).
Discussion
Toll-like receptors (TLRs) are a 13-transmembrane receptor family, which is at the forefront of guiding adaptive and innate immune responses to invaders such as viruses, fungi, bacteria, as well as parasites.20–22
When β-coronavirus infection occurs, TLR2 is necessary for the release of inflammatory cytokines because it can detect the SARS-CoV-2 envelope protein. Hence, previous study identified TLR2 as a possible target in treatment intervention strategies versus this lethal pandemic and has been shown to play an essential role in the development of COVID-19.5
At 4q32, there are two exons of the TLR2 gene. At 2258 bp in the TLR2 rs5743708 variant, a substitution of guanine for an adenine occurred, resulting in the substitution of an arginine-to-glutamine at residue 753.23
Our research found that the TLR2 rs5743708 mutant (G/A) genotype and the A allele were significantly related to an elevated risk of COVID-19 infection and were represented as a major risk factor for infection severity.
Ogus et al24 demonstrated that this 753Q allele was linked to an increased risk of developing TB, also having an impact on the severity of the disease because it resulted in a reduced responsiveness of macrophages to bacterial peptides, leading to a weakened immunological response.
According to Schroder et al,25 heterozygosity for the Arg753Gln variant exhibited a decreased induction of tumour necrosis factor-alpha as well as interferon-gamma when stimulated with Borrelia lysate in comparison to those not showing this variant. This seemed to protect against the onset of late-stage Lyme illness instead of being a risk factor for the illness.
Kijpittayarit et al26 showed that this variant has been linked to an elevated susceptibility to cytomegalovirus infection after liver transplantation. In addition, Tabel et al27 found that this variant was a risk factor for gram-positive pathogen-caused children’s urinary tract infections.
In addition, some studies reported that this variant made children more likely to develop rheumatic fever and recurring febrile infections,28,29 while it also increased the risk of developing familial Mediterranean fever, potentially affecting disease severity and the development of complications in adults.30
Similarly, El-Nabi et al31 reported that the Arg753Gln variant in the TLR2 gene was linked to CMV infection in Egyptian bone marrow recipients. They showed that 85% of the patients screened were G/G homozygous, whereas 15% were G/A heterozygous; there were no patients that were homozygous for (A/A).
Previous studies demonstrated that this variant was also linked to non-infectious diseases, like type 1 diabetes, as this variant might drive the innate immune response to prime and promote an aggressive adaptive immune response;32 and coronary restenosis, as this variant leads to a loss-of-function mutation that shifts the balance toward a TH1 response, promoting an inflammatory process that results in restenosis.33 Moreover, this variant was also shown to confer an increased risk of allograft failure and death as it results in poor intracellular signalling and reduced cytokine release, increasing predisposition to microbial sepsis.34
TLR9 is mostly found in intracellular compartments and serves as a crucial gatekeeper in the detection and treatment of viral infection.35 Thus, it was explained that the intensification of hyper-inflammation and thrombotic problems brought on by SARS-CoV-2 may be caused by TLR9 activation, which is a quiet but powerful driving force.7
TLR9 gene is encoded via 2 exons, the second of which represents the main coding area and maps to 3p21.3. There are numerous cases of genetic variants that result in TLR9 gain-of-function. For instance, the variant of the C allele of rs5743836 (T-1237C) that causes the activation of pro-inflammatory chemokines and cytokines as well as adaptive immunological responses, has been linked to a higher incidence of ICU-acquired infection and immune-mediated disease.36,37 The amplification of TLR9/IL-6 signalling by T-1237C was particularly found to result in a deregulation of B-cell stimulation and proliferation in response to CpG stimulation.38
The mutant (C/C) genotype and TLR9 rs5743836 C allele were found to be significantly linked with an elevated risk and severity of infection with COVID-19 in our study.
This result was in concordance with the study by Elsherif et al,39 which found that the TLR9-1237T/C variant was a risk factor for the infection-to-severe-sepsis development in paediatric patients.
This variant was also linked to non-infectious disorders, including a higher risk of asthma in a European-American cohort, a significant increase in the prevalence of Crohn’s disease among patients,40,41 and a higher risk of systemic lupus erythematosus among Asian communities.42
Regarding polymorphisms that were found to be associated with COVID-19 disease susceptibility and severity, Delanghe et al, showed that polymorphisms in angiotensin-converting enzyme-1 (ACE-1), complement C3, human homeostatic iron regulator protein (HFE) and cystic fibrosis transmembrane conductance regulator (CFTR) genes were associated with prevalence and severity.43
Alseoudy et al showed that TLR3 rs3775290 and TLR7 rs179008 polymorphisms were associated with increased susceptibility to COVID-19 disease.17
Recent research illustrated that some genetic mutations increase the severity of COVID-19 disease as D614G which increases the viral load as it enhances the interaction of spike and ACE2. In addition, other mutations increase spike-ACE2 affinity as L452R, S477G and S477N.44–48
Our study found that there has been no relation between TLR2 rs5743708 and TLR9 rs5743836 variants and citrullinated histones (H3), which are strong indicators of NETs and thrombosis. Therefore, these variants do not seem to have an impact on the formation and effects of NETs and, hence, thrombus formation. Furthermore, there has been no significant correlation between the two variants and other pro-inflammatory and coagulation indicators.
Thalin et al showed that quantification of citrullinated histone (H3) can be used as a specific, precise, and reliable indicator of NETs.49
Among the limitations of this study is that it does not include different age groups as infants, children, and elderly population. More studies involving pregnant females and patients with associated comorbidities are also required.
Conclusion
We concluded that TLR2 rs5743708 and TLR9 rs5743836 variants might be independent risk factors influencing COVID-19 disease susceptibility and severity, but they do not seem to be associated with markers of thrombosis.
This finding might be useful for the early detection of the disease’s course and progression as well as for prioritising patients who might profit from TLR2 and TLR9 antagonists. In addition, individuals could be categorized according to their genetic testing of TLR2 and TLR9 SNPs to specify who are in great need for the vaccine.
Abbreviations
TLRs, toll-like receptors; COVID-19, coronavirus disease 2019; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2; ssRNA, single-stranded RNA; PAMPs, pathogen-associated molecular pattern; SNPs; single nucleotide polymorphisms; NETs, neutrophil extracellular traps; RT-PCR, reverse-transcriptase polymerase chain reaction; WHO, World Health Organization; IRB, institutional review board; CBC, complete blood count; CRP, C-reactive protein; MGB, minor groove binder; NFQ, non-fluorescent quenchers; AD, allele discrimination; HRCT, high resolution-CT; SPSS, statistical package for social sciences; HWE, Hardy-Weinberg equilibrium; OR, odds ratio; ESR, erythrocyte sedimentation rate; ALT, alanine transaminase; AST, aspartate transaminase.
Acknowledgments
We would like to express our thanks to all voluntary participants in this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Al-Tawfiq JA, Petersen E, Memish ZA, Perlman S, Zumla A. Middle East respiratory syndrome coronavirus—The need for global proactive surveillance, sequencing, and modeling. Travel Med Infect Dis. 2021;43:102118. doi:10.1016/j.tmaid.2021.102118
2. Ghosh S, Dellibovi-Ragheb TA, Kerviel A, et al. ß-coronaviruses use lysosomes for egress instead of the biosynthetic secretory pathway. Cell. 2020;183(6):1520–1535.e14. doi:10.1016/j.cell.2020.10.039
3. Oliveira-Nascimento L, Massari P, Wetzler LM. The role of TLR2 in infection and immunity. Front Immunol. 2012;18(3):79. doi:10.3389/fimmu.2012.00079
4. Sariol A, Perlman S. SARS-COV-2 takes its Toll. Nat Immunol. 2021;22:801–802. doi:10.1038/s41590-021-00962-w
5. Zheng M, Karki R, Williams EP, et al. TLR2 senses the SARS-CoV-2 envelope protein to produce inflammatory cytokines. Nat Immunol. 2021;22:829–838. doi:10.1038/s41590-021-00937-x
6. El Kebir D, Damlaj A, Makhezer N, Filep JG. Toll-like receptor-9 signaling regulates tissue factor and tissue factor pathway inhibitor expression in human endothelial cells and coagulation in mice. Crit Care Med. 2015;43(6):e179–e189. doi:10.1097/CCM.0000000000001005
7. Bezemer GPG, Garssen J. TLR9 and COVID-19: a multidisciplinary theory of a multifaceted therapeutic target. Front Pharmacol. 2021;11:601685. doi:10.3389/fphar.2020.601685
8. Fuchs TA, Brill A, Duerschmied D, et al. Extracellular DNA traps promote thrombosis. Proc Natl Acad Sci U S A. 2010;107(36):15880–15885. doi:10.1073/pnas.1005743107
9. Barnes BJ, Adrover JM, Baxter-Stoltzfus A, et al. Targeting potential drivers of COVID-19: neutrophil extracellular traps. J Exp Med. 2020;217(6):e20200652. doi:10.1084/jem.20200652
10. Itagaki K, Kaczmarek E, Lee YT, et al. Mitochondrial DNA released by trauma Induces neutrophil extracellular traps. PLoS One. 2015;10(3):e0120549. doi:10.1371/journal.pone.0120549
11. Schellekens GA, Visser H, de Jong BA, et al. The diagnostic properties of rheumatoid arthritis antibodies recognizing a cyclic citrullinated peptide. Arthritis Rheum. 2000;43(1):155–163. doi:10.1002/1529-0131(200001)43:1<155::AID-ANR20>3.0.CO;2-3
12. Sung PS, Hsieh SL. CLEC2 and CLEC5A. Pathogenic host factors in acute viral infections. Front Immunol. 2019;10:2867. doi:10.3389/fimmu.2019.02867
13. Sung P, Yang S, Peng Y, Sun C, Tao M, Hsieh S. CLEC5A and TLR2 a 1 re critical in SARS-CoV-2-induced NET formation and lung inflammation. J Biomed Sci. 2022;29:52. doi:10.1186/s12929-022-00832-z
14. Taha SI, Shata AK, Baioumy SA, et al. Toll-like receptor 4 polymorphisms (896A/G and 1196C/T) as an indicator of COVID-19 severity in a convenience sample of Egyptian patients. J Inflamm Res. 2021;14:6293–6303. doi:10.2147/JIR.S343246
15. Khandia R, Pandey MK, Khan AA, Rzhepakovsky IV, Gurjar P, Karobari MI. Codon usage and context analysis of genes modulated during SARS-CoV-2. Infection and dental inflammation. Vaccines. 2022;10(11):1874. doi:10.3390/vaccines10111874
16. Hill AV. The genomics and genetics of human infectious disease susceptibility. Annu Rev Genomics Hum Genet. 2001;2:373–400. doi:10.1146/annurev.genom.2.1.373
17. Alseoudy MM, Elgamal M, Abdelghany DA, et al. Prognostic impact of toll-like receptors gene polymorphism on outcome of COVID-19 pneumonia: a case-control study. Clin Immunol. 2022;235:108929. doi:10.1016/j.clim.2022.108929
18. World Health Organization. Coronavirus disease (COVID-19) weekly epidemiological update and weekly operational update; 2019. Available from: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports.
19. Bai HX, Hsieh B, Xiong Z, et al. Performance of radiologists in differentiating COVID 19 from viral pneumonia on chest CT. Radiology. 2020;296(2):E46–E54. doi:10.1148/radiol.2020200823
20. Akira S. Toll-like receptor signaling. J Biol Chem. 2003;278(40):38105–38108. doi:10.1074/jbc.r300028200
21. Takeda K, Akira S. TLR signaling pathways. Semin Immunol. 2004;16(1):3–9. doi:10.1016/j.smim.2003.10.003
22. Pasare C, Medzhitov R. Toll-like receptors: linking innate and adaptive immunity. Adv Exp Med Biol. 2005;560:11–18. doi:10.1007/0-387-24180-9_2
23. Lorenz E, Mira JP, Cornish KL, Arbour NC, Schwartz DA, Tuomanen EI. A novel polymorphism in the toll-like receptor 2 gene and its potential association with staphylococcal infection. Infect Immunol. 2000;68:6398–6401. doi:10.1128/IAI.68.11.6398-6401.2000
24. Ogus AC, Yoldas B, Ozdemir T, et al. The Arg753GLn polymorphism of the human toll-like receptor 2 gene in tuberculosis disease. Eur Respir J. 2004;23(2):219–223. doi:10.1183/09031936.03.00061703
25. Schroder NW, Diterich I, Zinke A, et al. Heterozygous Arg753Gln polymorphism of human TLR-2 impairs immune activation by Borrelia burgdorferi and protects from late-stage Lyme disease. J Immunol. 2005;175:2534–2540. doi:10.4049/jimmunol.175.4.2534
26. Kijpittayarit S, Eid AJ, Brown RA, Paya CV, Razonable RR. Relationship between Toll-like receptor 2 polymorphism and cytomegalovirus disease after liver transplantation. Clin Infect Dis. 2007;44:1315–1320. doi:10.1086/514339
27. Tabel Y, Berdeli A, Mir S. Association of TLR2 gene Arg753Gln polymorphism with urinary tract infection in children. Int J Immunogenet. 2007;34:399–405. doi:10.1111/j.1744-313X.2007.00709.x
28. Berdeli A, Celin HA, Ozyurek R, Dogrusoz B, Aydin HH. TLR-2 gene Arg753Gln polymorphism is strongly associated with acute rheumatic fever in children. J Mol Med. 2005;83:535–541. doi:10.1007/s00109-005-0677-x
29. Kutukculer N, Yeniay BS, Aksu G, Berdeli A. Arg753Gln polymorphism of the human toll-like receptor-2 gene in children with recurrent febrile infections. Biochem Genet. 2007;45:507–514. doi:10.1007/s10528-007-9091-0
30. Ozen S, Berdeli A, Turel B, et al. Arg753Gln TLR-2 polymorphism in familial Mediterranean fever: linking the environment to the phenotype in a monogenic inflammatory disease. J Rheumatol. 2006;33:2498–2500. PMID: 17013994.
31. El-Nabi SH, Sayed S, Abd-Elhafez MA, Elfiky M, Abdel Moneim AE, El-Garawani I. Arg753Gln polymorphisms in the TLR2 gene are associated with cytomegalovirus infection in Egyptian bone marrow recipients. Endocr Metab Immune Disord Drug Targets. 2020;20(4):619–624. doi:10.2174/1871530319666191018124710
32. Park Y, Park S, Yoo E, Kim D, Shin H. Association of the polymorphism for Toll-like receptor 2 with type 1 diabetes susceptibility. Ann N Y Acad Sci. 2004;1037:170–174. doi:10.1196/annals.1337.028
33. Hamann L, Gomma A, Schroder NW, et al. A frequent toll-like receptor (TLR)-2 polymorphism is a risk factor for coronary restenosis. J Mol Med. 2005;83:478–485. doi:10.1007/s00109-005-0643-7
34. Eid AJ, Brown RA, Paya CV, Razonable RR. Association between toll-like receptor polymorphisms and the outcome of liver transplantation for chronic hepatitis C virus. Transplantation. 2007;84:511–516. doi:10.1097/01.tp.0000276960.35313.bf
35. Akira S, Hemmi H. Recognition of pathogen-associated molecular patterns by TLR family. Immunol Lett. 2003;85(2):85–95. doi:10.1016/s0165-2478(02)00228-6
36. Ng MTH, Van’t Hof R, Crockett JC, et al. Increase in NF-κB binding affinity of the variant C allele of the toll-like receptor 9 −1237T/C polymorphism Is associated with Helicobacter pylori induced gastric disease. Infect Immun. 2010;78(3):1345–1352. doi:10.1128/iai.01226-09
37. Chatzi M, Papanikolaou J, Makris D, et al. Toll-like receptor 2, 4 and 9 polymorphisms and their association with ICU-acquired infections in Central Greece. J Crit Care. 2018;47:1–8. doi:10.1016/j.jcrc.05.012
38. Carvalho A, Osorio NS, Saraiva M, et al. The C allele of rs5743836 polymorphism in the human TLR9 promoter links IL-6 and TLR9 up-regulation and confers Increased B-cell proliferation. PLoS One. 2011;6(11):e28256. doi:10.1371/journal.pone.0028256
39. Elsherif RH, Algebaly HAF, Ismail DK, et al. Toll-like receptors 2 and 9 gene polymorphisms in severe sepsis and septic shock: a single center study in the pediatric intensive care unit. Int J Clin Exp Med. 2019;12(4):4381–4386.
40. Lazarus R, Klimecki WT, Raby BA, et al. Single-nucleotide polymorphisms in the Toll-like receptor 9 gene (TLR9): frequencies, pairwise linkage disequilibrium, and haplotypes in three U.S. ethnic groups and exploratory case–control disease association studies. Genomics. 2003;81:85–91. doi:10.1016/s0888-7543(02)00022-8
41. Torok HP, Glas J, Tonenchi L, Bruennler G, Folwaczny M, Folwaczny C. Crohn’s disease is associated with a toll-like receptor-9 polymorphism. Gastroenterology. 2004;127:365–366. doi:10.1053/j.gastro.2004.05.051
42. Ng MTW, Lau CS, Chan TM, Wong WH, Lau YL. Polymorphisms of the toll-like receptor 9 (TLR9) gene with systemic lupus erythematosus in Chinese. Rheumatology. 2005;44:1456–1457. doi:10.1093/rheumatology/kei120
43. Delanghe JR, De Buyzere ML, Speeckaert MM. Genetic polymorphisms in the host and COVID-19 infection. Adv Exp Med Biol. 2021;1318:109–118. doi:10.1007/978-3-030-63761-3_7
44. Korber B, Fischer WM, Gnanakaran S, et al. Tracking changes in SARS-CoV-2 spike: evidence that D614G increases infectivity of the COVID-19 virus. Cell. 2020;182(4):812–827.e19. doi:10.1016/j.cell.2020.06.043
45. Kumar BK, Rohit A, Prithvisagar KS, Rai P, Karunasagar I, Karunasagar I. Deletion in the C-terminal region of the envelope glycoprotein in some of the Indian SARS-CoV-2 genome. Virus Res. 2021;291:198222. doi:10.1016/j.virusres.2020.198222
46. Shen L, Bard JD, Triche TJ, Judkins AR, Biegel JA, Gai X. Emerging variants of concern in SARS-CoV-2 membrane protein: a highly conserved target with potential pathological and therapeutic implications. Emerg Microbes Infect. 2021;10(1):885–893. doi:10.1080/22221751.2021.1922097
47. Singh A, Steinkellner G, K¨ochl K, Gruber K, Gruber CC. Serine 477 plays a crucial role in the interaction of the SARS-CoV-2 spike protein with the human receptor ACE2. Sci Rep. 2021;11(1):1–11. doi:10.1038/s41598-021-83761-5
48. Ortuso F, Mercatelli D, Guzzi PH, Giorgi FM. Structural genetics of circulating variants affecting the SARS-CoV-2 spike/human ACE2 complex. J Biomol Struct Dyn. 2021;1–11. doi:10.1080/07391102.2021.1886175
49. Thålin C, Daleskog M, Göransson SP, et al. Validation of an enzyme-linked immunosorbent assay for the quantification of citrullinated histone H3 as a marker for neutrophil extracellular traps in human plasma. Immunol Res. 2017;65(3):706–712. doi:10.1007/s12026-017-8905-3
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.