")
Back to Journals » OncoTargets and Therapy » Volume 7
IL-17A (-197G/A) and IL-17F (7488T/C) gene polymorphisms and cancer risk in Asian population: a meta-analysis
Authors Dai W, Zhou Q, Tan X, Sun C
Received 20 February 2014
Accepted for publication 17 March 2014
Published 13 May 2014 Volume 2014:7 Pages 703—711
DOI https://doi.org/10.2147/OTT.S62781
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Wei Dai, Qing Zhou, Xuexin Tan, Changfu Sun
Department of Oromaxillofacial, Head and Neck Surgery, Department of Oral and Maxillofacial Surgery, School of Stomatology, China Medical University, Shenyang, People's Republic of China
Abstract: Interleukin (IL)-17 has been shown to play an important role in the pathogenesis of inflammation and cancer. The IL-17A (-197G/A) and IL-17F (7488T/C) polymorphisms have been extensively investigated with cancer risk, but individually published results have been inconclusive. The aim of this study was to clarify the effects of the IL-17A (-197G/A) and IL-17F (7488T/C) polymorphisms on cancer risk in Asian populations. Relevant studies were identified by searching databases extensively. The association between the IL-17A (-197G/A) and IL-17F (7488T/C) polymorphisms and cancer risk was assessed by odds ratios (ORs) together with their 95% confidence intervals (CIs). A total of 12 articles with adequate information satisfied our inclusion criteria; these included 12 studies, with 4,540 cases and 5,875 controls, of IL-17A (-197G/A) polymorphism and seven studies, with 1,960 cases and 3,226 controls, of IL-17F (7488T/C) polymorphism. In the overall analysis, the IL-17A (-197G/A) polymorphism was significantly associated with increased cancer risk (P<0.05), for all genetic models. However, there was no statistically significant association between IL-17F (7488T/C) and cancer risk (P>0.05), for any genetic models. Furthermore, stratification by cancer type revealed a significant correlation between the IL-17A (-197G/A) polymorphism and cancer risk for all cancer types. When stratified by source of controls, a significant correlation was observed between the IL-17A (-197G/A) polymorphism and cancer risk in the population-based control subgroup but not in hospital-based control subgroup. In conclusion, our meta-analysis provides evidence that the IL-17A (-197G/A) polymorphism might be associated with cancer risk, while no evidence suggested a significant association between IL-17F (7488T/C) polymorphism and cancer risk.
Keywords: interleukin-17, meta-analysis, cancer, polymprphism
Introduction
Interleukin (IL)-17A, initially termed as cytotoxic T lymphocyte-associated antigen (CTLA)-8, is the founding member of IL-17 cytokine family, consisting of six homologous proteins (from IL-17A to IL-17F).1 IL-17 is a pleiotropic cytokine that acts on multiple cell types to enhance the production of proinflammatory molecules, including the induction of IL-6, IL-8, and prostaglandin E2, as well as enhancing granulocyte colony-stimulating factor and CXC chemokine production.2,3 In addition, IL-17 induces secretion of tumor necrosis factor (TNF)-α, IL-1β, and stromelysin, by macrophages,4,5 and activates the nuclear factor κB and activator protein 1 transcription factors, which may explain its proinflammatory properties.6,7
Recently, several studies have shown that IL-17 can have either a protumor or antitumor role in different cancer models.8 Accumulating evidence indicates that IL-17 expression is aberrant in several human tumors, such as ovarian cancer,9 cervical cancer,10 breast cancer,11 hepatocellular carcinoma,12 esophageal cancer,13 gastric cancer,14 and colorectal cancer.15 But the underlying mechanism of IL-17 in tumor initiation and progression is not yet completely clear. Of note, Kryczek et al suggested that endogenous IL-17 or/and T helper (Th)17 cells may play a protective role in tumor immunity.16 Martin-Orozco et al reported that IL-17A-deficient mice were more susceptible to developing lung melanoma.17 These studies showed an opposite role of IL-17; specifically, they suggest an antitumor effect, by promoting cytotoxic T lymphocyte generation.
A large number of previous studies have suggested the possible correlation between genetic polymorphisms of cancer susceptibility genes and higher risk of human malignant tumors. There are several single nucleotide polymorphisms (SNPs) that are reported for the IL-17 gene, and the IL-17A gene polymorphism IL-17A (-197G/A, rs2275913) and IL-17F gene polymorphism IL-17F (7488T/C, rs763780) are reported to be associated in gastric carcinogenesis.18 IL-17A (-197G/A), located at a position -197 from the starting codon of the IL-17A gene, may regulate the expression of messenger ribonucleic acid (mRNA).19 Recently, Kawaguchi et al reported that IL-17F (7488T/C) causes a histidine (His)-to-arginine (Arg) substitution at amino acid 161 (H161R) variant.20 To date, many studies have been published that assessed the association between the IL-17A (-197G/A) and IL-17F (7488T/C) polymorphisms and cancer susceptibility, but they have reported controversial results. A single case-control study may fail to completely demonstrate the complicated genetic relationship because of the small sample size.21 In order to provide strong evidence of the effects of the IL-17A (-197G/A) and IL-17F (7488T/C) polymorphisms on cancer susceptibility, we performed a meta-analysis, by combining data from numerous published studies.
Materials and methods
Search strategy
A comprehensive literature search of PubMed, Embase, Web of Science, Science Direct, SpringerLink, and the Chinese National Knowledge Infrastructure databases (up to November 2013) was conducted to identify case-control studies that investigated the association between IL-17 polymorphisms and cancer risk. The search strategies were based on combinations of the following keywords: “cancer or carcinoma or neoplasm or tumor” AND “IL-17 or IL-17A or IL-17F or CTLA” AND “polymorphism or variant or mutation or genotype”. There was no restriction on time period, sample size, population, or language, in order to minimize potential publication bias.
Selection criteria
Studies were included in the meta-analysis if they satisfied the following inclusion criteria: 1) case-control studies focused on association between the IL-17A (-197G/A) or IL-17F (7844T/C) polymorphism and cancer risk; 2) study design that was either retrospective or a nested case-control design; 3) any diagnoses of patients with cancer had to be confirmed by pathological examinations; and 4) sufficient published data to estimate the odds ratio (OR) and corresponding 95% confidence interval (CI). The exclusion criteria of the meta-analysis were: 1) case-control studies not focusing on the correlation between the IL-17A (-197G/A) or IL-17F (7844T/C) polymorphism and cancer risk; 2) studies with duplicate data; 3) studies with incomplete data; and 4) meta-analyses, letters, reviews, and editorial articles. When an individual author published several articles obtained from the same patient population, only the newest or most complete article was included in the analysis.
Data extraction
Two authors independently extracted data from the included studies (Dai and Zhou). For each eligible study, the following information was extracted: first author’s name, year of publication, country, ethnicity, cancer type, numbers of cases and controls, source of controls (hospital-based controls or population-based controls), genotyping method, and the Hardy–Weinberg equilibrium frequency. In cases of discrepancy, a consensus was reached by consulting a third author.
Sensitivity analysis
Sensitivity analysis was employed by sequentially excluding a single study each time, in an attempt to identify the potential influence of the individual data set to the pooled ORs.22
Statistical analysis
Data management and processing were performed using Review Manager (RevMan) 5.0 software (The Nordic Cochrane Centre, Copenhagen) and Stata 11.0 (Stata Corp, College Station, Texas, USA). The strength of the association between the IL-17 gene polymorphisms and cancer risk was measured by crude ORs and 95% CIs. The significance of the combined ORs was determined by a Z test, and two-sided P-value <0.05 was considered significant. The pooled ORs were calculated for the allele model (mutation [M] allele versus [vs] wild [W] allele), dominant model (WM + MM vs WW), recessive model (MM vs WM + WW), homozygote comparison (MM vs WW), and heterozygote comparison (WM vs WW), respectively. Stratified analyses were conducted with respect to cancer type and source of controls. Statistical heterogeneity among the studies was evaluated using the chi square-based Q-test, and heterogeneity was considered significant when P<0.1 for Q statistic. Heterogeneity was quantified by I2 statistics examining the percentage of heterogeneity (0%–25% = no heterogeneity; 25%–50% = moderate heterogeneity; 50%–75% = large heterogeneity; and 75%–100% = extreme heterogeneity).23 The random effects model was used when heterogeneity was significant (I2>50% or P≤0.10); otherwise, the fixed effects model was used. Publication bias was examined with both Begg’s funnel plot24 and the Egger’s regression method,25 and P<0.05 was considered representative of statistically significant publication bias.
Results
Characteristics of eligible studies
A total of 136 relevant studies were reviewed based on our selection strategy. The stepwise selection process is shown in Figure 1. After sequential selection, a total of 12 articles with adequate information satisfied our inclusion criteria, among which were 12 studies, with 4,540 cases and 5,875 controls, of the IL-17A (-197G/A) polymorphism18,26–36 and seven studies, with 1,960 cases and 3,226 controls, of the IL-17F (7488T/C) polymorphism18,29,31–34,36 All studies were case-control designs, with subjects of Asian ancestry. Out of the 12 applicable studies, eight were studies of gastric cancer, one was cervical cancer, one was breast cancer, one was ovarian cancer, and one was bladder cancer. Population-based controls were used in seven studies, and hospital-based controls were used in five studies. The detailed characteristics of the eligible studies are listed in Table 1. The genotype distribution of IL-17 polymorphisms in the controls was in compliance with Hardy–Weinberg equilibrium, except for two studies of IL-17A (-197G/A) polymorphism18,34 and three studies of IL-17F (7488T/C) polymorphism.30,32,33
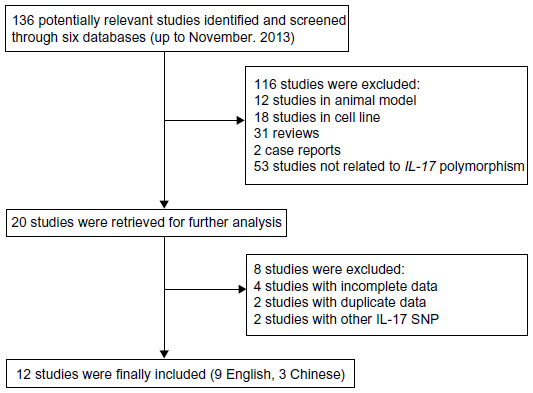 | Figure 1 Flow chart of the literature search and study selection. |
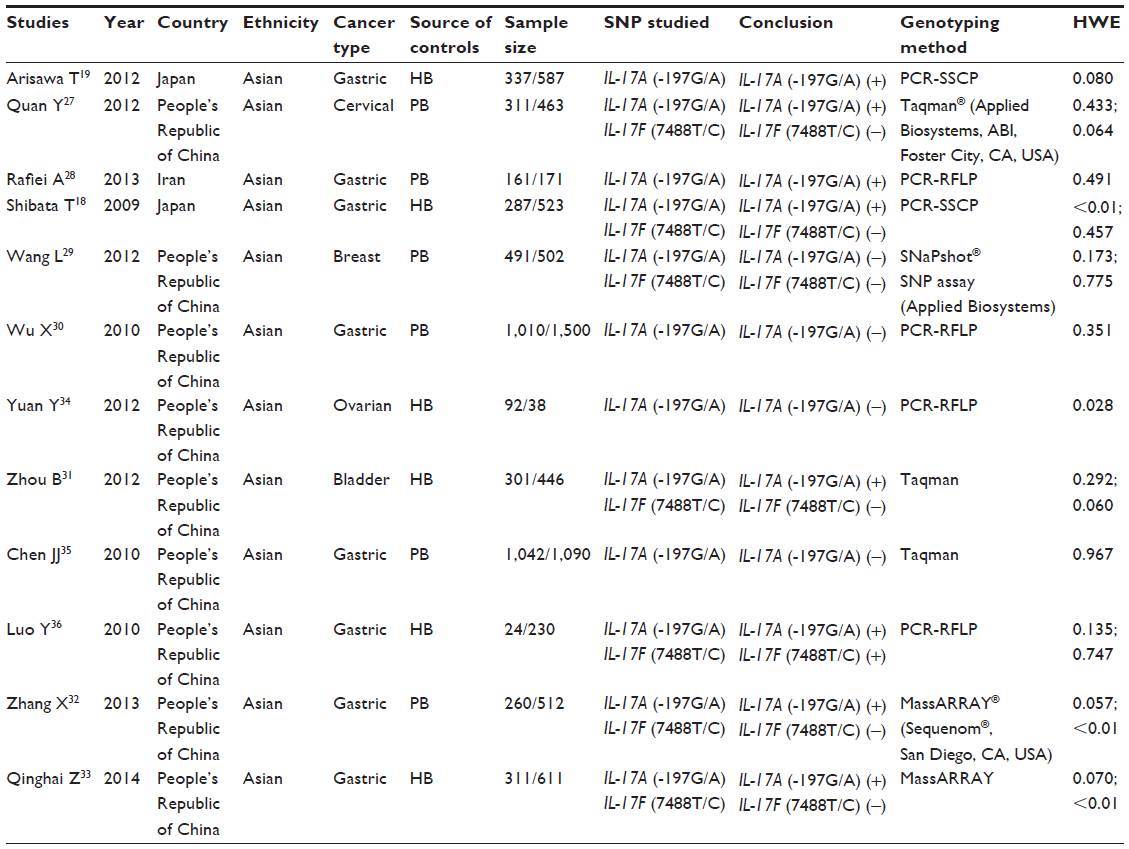 | Table 1 Characteristics of studies included in the present meta-analysis |
Quantitative data synthesis
IL-17A (-197G/A) polymorphism
The results of the meta-analysis are presented in detail in Table 2. For the IL-17A (-197G/A) polymorphism, 12 studies were combined. In the overall analysis, a significant association between IL-17A (-197G/A) polymorphism and cancer risk was found under the allele model (OR =1.23, 95% CI =1.10–1.38, P<0.01), dominant model (OR =1.22, 95% CI =1.08–1.37, P<0.01), recessive model (OR =1.50, 95% CI =1.17–1.93, P<0.01), homozygous comparison (OR =1.62, 95% CI =1.27–2.06, P<0.01), and heterozygous comparison (OR =1.11, 95% CI =1.01–1.21, P=0.03).
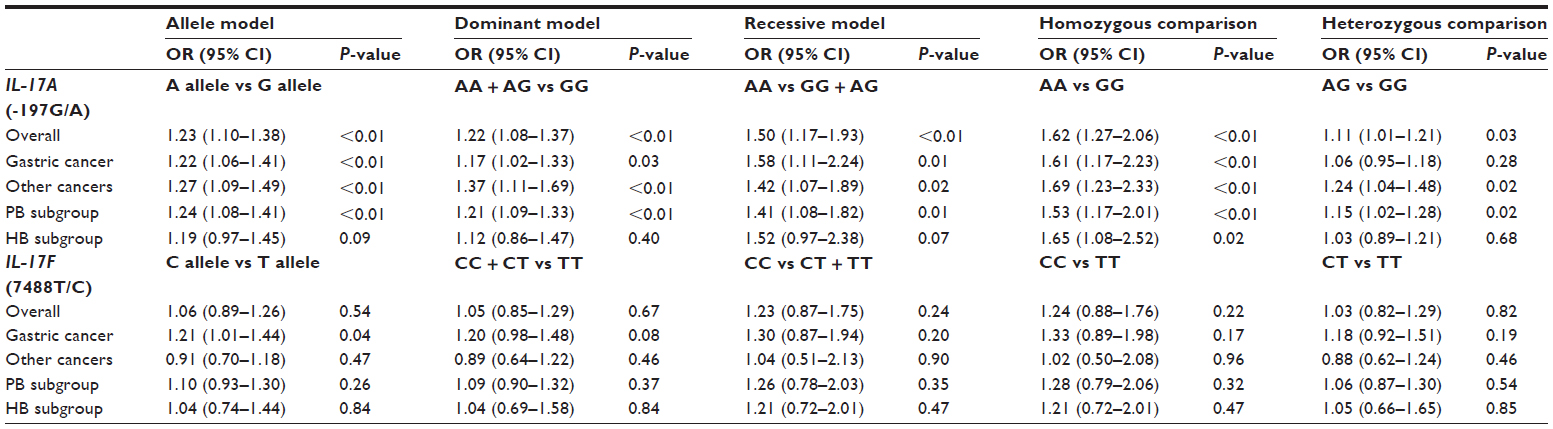 | Table 2 Meta-analysis of the IL-17A (-197G/A) polymorphism and IL-17F (7488T/C) polymorphism with risk of cancer |
Subgroup analysis, stratified by cancer type, also suggested a significant association between IL-17A (-197G/A) polymorphism and cancer risk in the gastric cancer subgroup, under the allele model (OR =1.22, 95% CI =1.06–1.41, P<0.01), dominant model (OR =1.17, 95% CI =1.02–1.33, P=0.03), recessive model (OR =1.58, 95% CI =1.11–2.24, P<0.01), and homozygous comparison (OR =1.61, 95% CI =1.17–2.23, P<0.01). Moreover, a significant association was observed in the other cancer subgroups, under any of the genetic models (P<0.05) (Table 2 and Figure 2A). Therefore, no difference was observed between the cancer types.
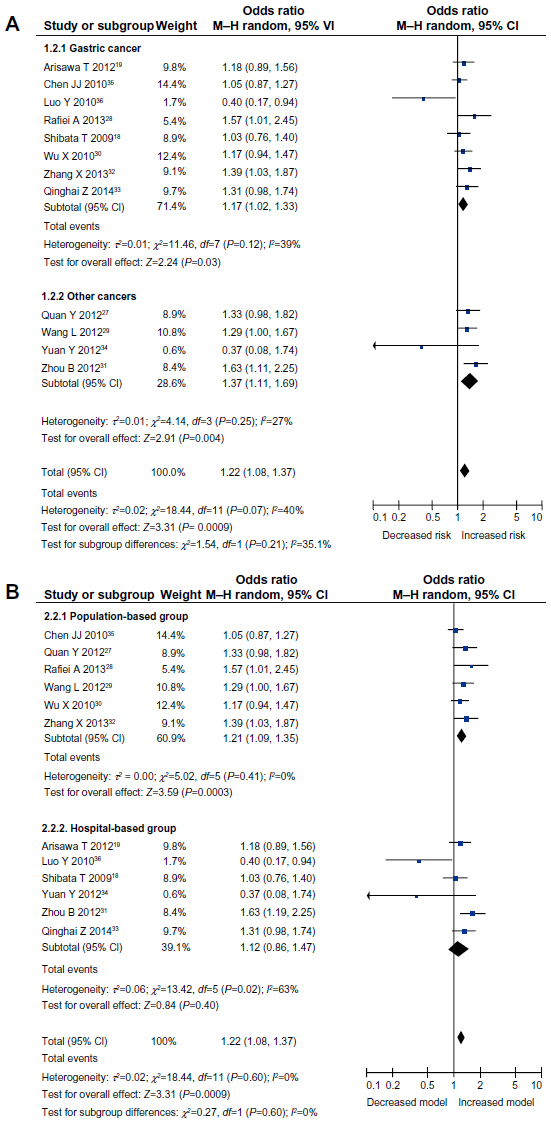 | Figure 2 Meta-analysis to evaluate the association between the IL-17A (-197G/A) polymorphism and cancer risk (dominant model: AA + AG vs GG). (A) subgroup analysis by cancer type; (B) subgroup analysis by source of control. |
Stratified analyses by the source of control also suggested that IL-17A (-197G/A) polymorphism increased the cancer risk in the population-based control subgroup, under all genetic models (allele model: OR =1.24, 95% CI =1.08–1.41, P<0.01; dominant model: OR =1.21, 95% CI =1.09–1.33, P<0.01; recessive model: OR =1.41, 95% CI =1.08–1.82, P=0.01; homozygous comparison: OR =1.53, 95% CI =1.17–2.01, P<0.01; and heterozygous model: OR =1.15, 95% CI =1.02–1.28, P=0.02). In contrast, a significant association was only found under homozygous comparison (OR =1.65, 95% CI =1.08–2.52, P=0.02) in the hospital-based control subgroup (Table 2 and Figure 2B).
IL-17F (7488T/C) polymorphism
For the IL-17F (7488T/C) polymorphism, seven studies were combined. When all seven studies were pooled into the meta-analysis, there was no significant association between the IL-17F (7488T/C) polymorphism and cancer susceptibility, under all genetic models (allele model: OR =1.06, 95% CI =0.89–1.26, P=0.54; dominant model: OR =1.05, 95% CI =0.85–1.29, P=0.67; recessive model: OR =1.23, 95% CI =0.87–1.75, P=0.24; homozygous comparison: OR =1.24, 95% CI =0.88–1.76, P=0.22; and heterozygous model: OR =1.03, 95% CI =0.82–1.29, P=0.82).
In the stratified analysis by cancer type, only the allele model (OR =1.21, 95% CI =1.01–1.44, P=0.04) displayed a significant association between IL-17F (7488T/C) polymorphism and cancer risk, in the gastric cancer subgroup. No significant association was found, under any genetic models, in the other cancer subgroup (all P>0.05) (Table 2 and Figure 3A).
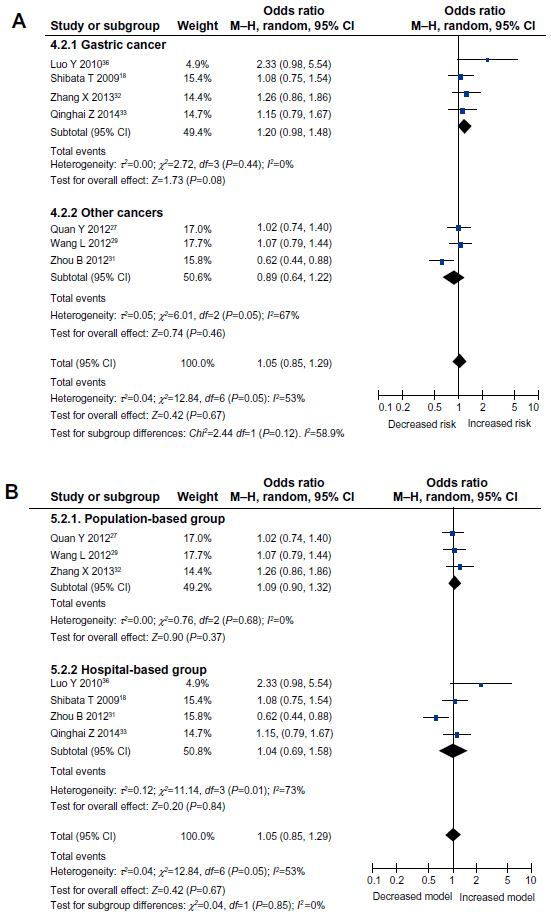 | Figure 3 Meta-analysis to evaluate the association between the IL-17F (7488T/C) polymorphism and cancer risk (dominant model: CC + CT vs TT). (A) subgroup analysis by cancer type; (B) subgroup analysis by source of control. |
Next, we performed a subgroup analysis according to the source of controls. The results showed that IL-17F (7488T/C) polymorphism was not associated with cancer risk, in either the population-base control subgroup or the hospital-based control subgroup (all P>0.05) (Table 2 and Figure 3B). Therefore, differences in the controls did not affect the association between IL-17F (7488T/C) polymorphism and cancer risk.
Sensitivity analysis
The sensitivity analysis indicated that no single study influenced the pooled OR value, either for IL-17A (-197G/A) polymorphism or for IL-17F (7488T/C) polymorphism, suggesting that the results of this meta-analysis are stable (data not shown).
Publication bias
Publication bias of the selected articles was assessed using Begg’s funnel plot and Egger’s test. The shape of the funnel plot did not show an obvious publication bias for IL-17A (-197G/A) polymorphism or IL-17F (7488T/C) polymorphism (Figure 4A and B). Similarly, no evidence of publication bias was observed by Egger’s test, for the IL-17A (-197G/A) dominant model (P=0.270) or the IL-17F (7488T/C) dominant model (P=0.250).
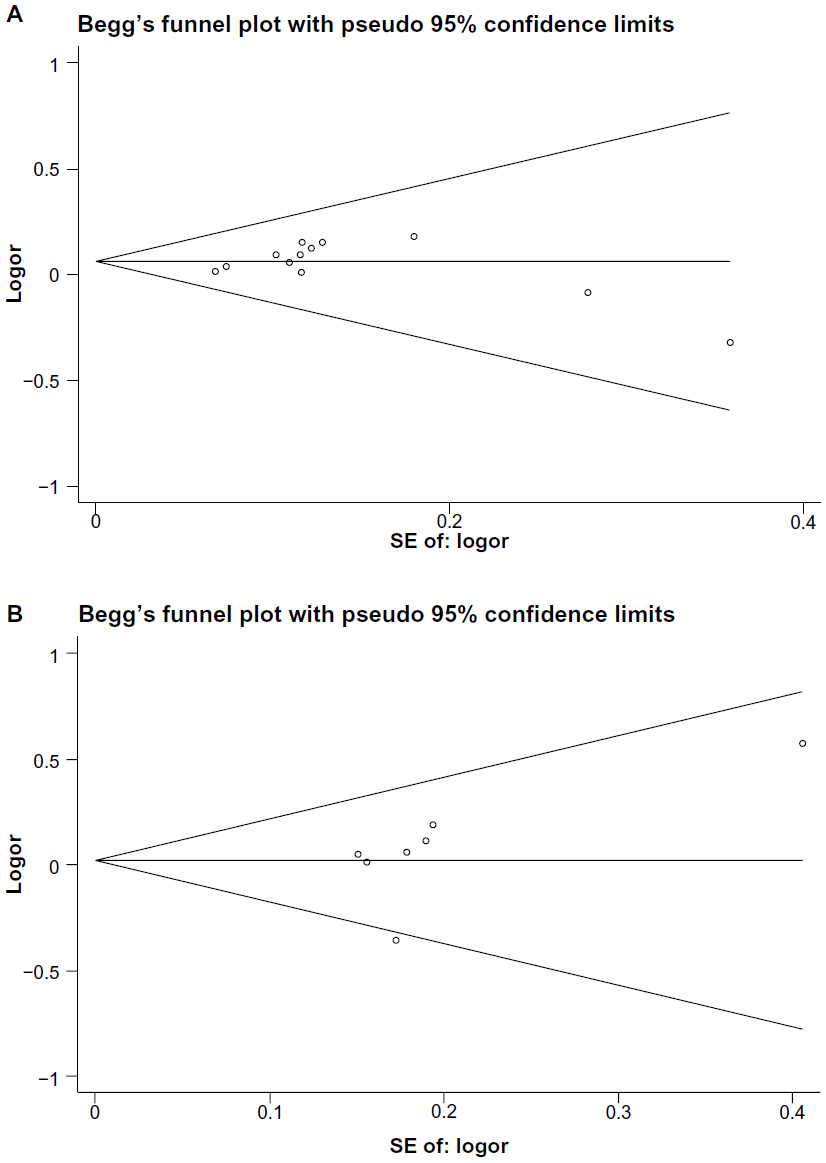 | Figure 4 Funnel plots of IL-17A (-197G/A) polymorphism and IL-17F (7488T/C) polymorphism in the dominant model. (A) IL-17A (-197G/A) polymorphism; (B) IL-17F (7488T/C) polymorphism. |
Discussion
Large-sample and unbiased, epidemiological studies of predisposition genes polymorphisms could provide insight to etiology of diseases. However, the findings are generally inconsistent, probably due to some limitation in these studies, such as small sample size, and mix of ethnicities and cancer types. Meta-analysis is considered a powerful tool for combining the contradicting results from different studies to achieve greater statistical power and obtain more reliable results than with a single study.37 In this meta-analysis, we investigated the association between IL-17A (-197G/A) (rs2275913) and IL-17F (7488T/C) (rs763780) and cancer risk. Overall, our study provided evidence that IL-17A (-197G/A) polymorphism was associated with cancer susceptibility. However, no association was detected between IL-17F (7488T/C) polymorphism and cancer susceptibility. Sensitivity analysis and publication bias further strengthened the validity of these results.
IL-17 family members belong to a distinct category of cytokine and are involved in coordinating local tissue inflammation, through release of proinflammatory and neutrophil-mobilizing cytokines.20 To date, investigation into the function of the IL-17 cytokine family has been extensive, but the majority of the effort has focused on the role of two members, IL-17A and IL-17F. The IL-17A and IL-17F genes are both located at 6p12. In addition, among the IL-17 family, from the alignment of the amino acid sequences, IL-17A and IL-17F have the highest overall amino acid sequence identity and share similar functions in terms of their ability to induce chemokines, which are important in neutrophil recruitment and activation.31 IL-17A and IL-17F cytokines are both expressed by Th17 cells, which mediate chronic inflammation and cancer.9 Although the significance of IL-17A and -17F in the pathogenesis of the cancer has still remained unclear, we hypothesized that both the cytokines may affect the development of chronic inflammation followed by malignancy, coordinately or independently. Over the last several years, many studies have identified a variety of environmental, host immune status, and host genetic factors that play important roles in carcinogenesis. Moreover, Espinoza et al reported that IL-17A (-197G/A) polymorphism influences the response of the IL-17 gene promoter to factors released in response to T cell activation, thus leading to a differential IL-17 production.38 Kawaguchi et al revealed that IL-17F (7488T/C), which causes a His–to–Arg substitution at amino acid 161 (H161R) of IL-17F, lacked the ability to activate the mitogen-activated protein kinase pathway, cytokine production, and chemokine production, and blocked induction of IL-8 expression by wild type IL-17F in in vitro functional experiments.39 Therefore, we evaluated the effect of the polymorphisms of two cytokines on cancer risk. Our investigation suggests that IL-17A (-197G/A), but not IL-17F (7488T/C), contributes to cancer susceptibility.
Subgroup analyses stratified by source of controls and cancer types were also performed. For IL-17A (-197G/A) polymorphism, a significant increased cancer risk was observed in gastric cancer and other cancers, suggesting that cancer type may not affect cancer susceptibility. Moreover, stratified analysis by source of controls suggested that IL-17A (-197G/A) polymorphism was associated with increased risk of cancer in the population-based control subgroup, but not in the hospital-based control subgroup. This may have been due to some selection biases existing in the hospital-based studies because such controls may have come from a population with a related disease and may not have been representative of the general population. For IL-17F (7488T/C) polymorphism, we found little association between IL-17F (7488T/C) polymorphism and cancer risk in subgroups analysis. This result indicates that the differences in cancer types and sources of controls do not affect cancer susceptibility.
Several potential limitations of the present meta-analysis should be acknowledged. First, significant heterogeneity was observed in the overall and subgroup analyses. Although two potential sources of the heterogeneity were investigated, including the source of controls and cancer types, neither of them sufficiently explained the between-study heterogeneity. Second, the sample size is not large enough, especially for subgroup analysis. Thus, we do not have adequate power to evaluate the possible association for both polymorphisms, and the observed significant associations in some subgroup analysis may be not accurate. Third, the lack of observations concerning gene–gene and gene–environment interactions could have influenced our results. Fourth, only published studies were included in the meta-analysis, and nonsignificant or negative findings may be unpublished. Hence, some inevitable publication biases might exist in the results.
Conclusion
In conclusion, our meta-analysis provides evidences that the IL-17A (-197G/A) polymorphism might be associated with cancer risk in the Asian populations, while no evidence suggested a significant association between the IL-17F (7488T/C) polymorphism and cancer risk. However, large-sample studies are warranted to validate our findings, especially in some types of cancer. More studies on gene–gene and gene–environment interactions should also be considered in the future, to obtain a more comprehensive understanding of the association between IL-17 polymorphisms and cancer risk.
Acknowledgments
This work has been supported by Grants from the National Natural Science Foundation of China (No 81301835).
Disclosure
The authors report no conflicts of interest in this work.
References
Kolls JK, Lindén A. Interleukin-17 family members and inflammation. Immunity. 2004;21(4):467–476. | |
Laan M, Cui ZH, Hoshino H, et al. Neutrophil recruitment by human IL-17 via C-X-C chemokine release in the airways. J Immunol. 1999;162(4):2347–2352. | |
Schwarzenberger P, Huang W, Ye P, et al. Requirement of endogenous stem cell factor and granulocyte-colony-stimulating factor for IL-17-mediated granulopoiesis. J Immunol. 2000;164(9):4783–4789. | |
Fossiez F, Djossou O, Chomarat P, et al. T cell interleukin-17 induces stromal cells to produce proinflammatory and hematopoietic cytokines. J Exp Med. 1996;183(6):2593–2603. | |
Jovanovic DV, Di Battista JA, Martel-Pelletier J, et al. IL-17 stimulates the production and expression of proinflammatory cytokines, IL-beta and TNF-alpha, by human macrophages. J Immunol. 1998;160(7):3513–3521. | |
Awane M, Andres PG, Li DJ, Reinecker HC. NF-kappa B-inducing kinase is a common mediator of IL-17-, TNF-alpha-, and IL-1 beta-induced chemokine promoter activation in intestinal epithelial cells. J Immunol. 1999;162(9):5337–5344. | |
Benchetrit F, Ciree A, Vives V, et al. Interleukin-17 inhibits tumor cell growth by means of a T-cell-dependent mechanism. Blood. 2002; 99(6):2114–2121. | |
Murugaiyan G, Saha B. Protumor vs antitumor functions of IL-17. J Immunol. 2009;183(7):4169–4175. | |
Miyahara Y, Odunsi K, Chen W, Peng G, Matsuzaki J, Wang RF. Generation and regulation of human CD4+ IL-17-producing T cells in ovarian cancer. Proc Natl Acad Sci U S A. 2008;105(40):15505–15510. | |
Schaffar A, Huyghe AS, Bomy H, Duriez P, Ego A, Pierrat V. [Breastfeeding: opinion and knowledge of pharmacists. A study in a semi-urban territory]. Arch Pediatr. 2012;19(5):476–483. French. | |
Zhu X, Mulcahy LA, Mohammed RA, et al. IL-17 expression by breast-cancer-associated macrophages: IL-17 promotes invasiveness of breast cancer cell lines. Breast Cancer Res. 2008;10(6):R95. | |
Kuang DM, Peng C, Zhao Q, Wu Y, Chen MS, Zheng L. Activated monocytes in peritumoral stroma of hepatocellular carcinoma promote expansion of memory T helper 17 cells. Hepatology. 2010;51(1):154–164. | |
Wang B, Li L, Liao Y, et al. Mast cells expressing interleukin 17 in the muscularis propria predict a favorable prognosis in esophageal squamous cell carcinoma. Cancer Immunol Immunother. 2013;62(10):1575–1585. | |
Meng XY, Zhou CH, Ma J, Jiang C, Ji P. Expression of interleukin-17 and its clinical significance in gastric cancer patients. Med Oncol. 2012;29(5):3024–3028. | |
Radosavljevic G, Ljujic B, Jovanovic I, et al. Interleukin-17 may be a valuable serum tumor marker in patients with colorectal carcinoma. Neoplasma. 2010;57(2):135–144. | |
Kryczek I, Wei S, Szeliga W, Vatan L, Zou W. Endogenous IL-17 contributes to reduced tumor growth and metastasis. Blood. 2009;114(2):357–359. | |
Martin-Orozco N, Muranski P, Chung Y, et al. T helper 17 cells promote cytotoxic T cell activation in tumor immunity. Immunity. 2009;31(5):787–798. | |
Shibata T, Tahara T, Hirata I, Arisawa T. Genetic polymorphism of interleukin-17A and -17F genes in gastric carcinogenesis. Hum Immunol. 2009;70(7):547–551. | |
Arisawa T, Tahara T, Tsutsumi M, Shibata T. Influence of IL17A polymorphisms on the aberrant methylation of DAPK and CDH1 in non-cancerous gastric mucosa. BMC Med Genet. 2012;13:59. | |
Kawaguchi M, Adachi M, Oda N, Kokubu F, Huang SK. IL-17 cytokine family. J Allergy Clin Immunol. 2004;114(6):1265–1273; quiz 1274. | |
Tein JY, Coxe S, Cham H. Statistical power to detect the correct number of classes in latent profile analysis. Struct Equ Modeling. 2013;20(4):640–657. | |
Thakkinstian A, McElduff P, D’Este C, Duffy D, Attia J. A method for meta-analysis of molecular association studies. Stat Med. 2005;24(9):1291–1306. | |
Higgins JP, Thompson SG. Quantifying heterogeneity in a meta-analysis. Stat Med. 2002;21(11):1539–1558. | |
Begg CB, Mazumdar M. Operating characteristics of a rank correlation test for publication bias. Biometrics. 1994;50(4):1088–1101. | |
Egger M, Davey Smith G, Schneider M, Minder C. Bias in meta-analysis detected by a simple, graphical test. BMJ. 1997;315(7109):629–634. | |
Arisawa T, Tahara T, Shiroeda H, et al. Genetic polymorphisms of IL17A and pri-microRNA-938, targeting IL17A 3′-UTR, influence susceptibility to gastric cancer. Hum Immunol. 2012;73(7):747–752. | |
Quan Y, Zhou B, Wang Y, et al. Association between IL17 polymorphisms and risk of cervical cancer in Chinese women. Clin Dev Immunol. 2012;2012:258293. | |
Rafiei A, Hosseini V, Janbabai G, et al. Polymorphism in the interleukin-17A promoter contributes to gastric cancer. World J Gastroenterol. 2013;19(34):5693–5699. | |
Wang L, Jiang Y, Zhang Y, et al. Association analysis of IL-17A and IL-17F polymorphisms in Chinese Han women with breast cancer. PLoS One. 2012;7(3):e34400. | |
Wu X, Zeng Z, Chen B, et al. Association between polymorphisms in interleukin-17A and interleukin-17F genes and risks of gastric cancer. Int J Cancer. 2010;127(1):86–92. | |
Zhou B, Zhang P, Wang Y, et al. Interleukin-17 gene polymorphisms are associated with bladder cancer in a Chinese Han population. Mol Carcinog. 2013;52(11):871–878. | |
Zhang X, Zheng L, Sun Y, Zhang X. Analysis of the association of interleukin-17 gene polymorphisms with gastric cancer risk and interaction with Helicobacter pylori infection in a Chinese population. Tumour Biol. 2014;35(2):1575–1580. | |
Qinghai Z, Yanying W, Yunfang C, Xukui Z, Xiaoqiao Z. Effect of interleukin-17A and interleukin-17F gene polymorphisms on the risk of gastric cancer in a Chinese population. Gene. 2014;537(2):328–332. | |
Yuan Y, Hu Y, Tao K, Zhang R. Association analysis of IL-17A and IL-17F gene polymorphism with epithelial ovarian cancer. Journal of Hunan Normal University (Medical Science). 2012;9(3):21–24, 33. | |
Chen JJ. Association Study of Polymorphisms in IL23R and IL17A Genes with the Susceptibility of Gastric Cancer [Master’s thesis]. Nanjing: Nanjing Medical University; 2010. | |
Luo Y. Association Analysis of Interleukin-17 Genes’ Single Nucleotide Polymorphisms (SNPs) with H. Pylori Infection Associated Gastric Disease in Chinese Population. Chongqing: Third Military Medical University; 2010. | |
Munafò MR, Flint J. Meta-analysis of genetic association studies. Trends Genet. 2004;20(9):439–444. | |
Espinoza JL, Takami A, Nakata K, et al; Japan Marrow Donor Program. A genetic variant in the IL-17 promoter is functionally associated with acute graft-versus-host disease after unrelated bone marrow transplantation. PLoS One. 2011;6(10):e26229. | |
Kawaguchi M, Takahashi D, Hizawa N, et al. IL-17F sequence variant (His161Arg) is associated with protection against asthma and antagonizes wild-type IL-17F activity. J Allergy Clin Immunol. 2006;117(4):795–801. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.