")
Back to Journals » Cancer Management and Research » Volume 13
Identification of Prognostic Biomarkers Among FAM83 Family Genes in Human Ovarian Cancer Through Bioinformatic Analysis and Experimental Verification
Authors Lin S, Du J, Hao J, Luo X, Wu H, Zhang H, Zhao X, Xu L, Wang B
Received 13 July 2021
Accepted for publication 23 October 2021
Published 15 November 2021 Volume 2021:13 Pages 8611—8627
DOI https://doi.org/10.2147/CMAR.S328851
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Ahmet Emre Eşkazan
Shaochong Lin,1,2 Junpeng Du,3 Jun Hao,1 Xiaohua Luo,1 Han Wu,1 Huifang Zhang,1 Xinxin Zhao,1 Lida Xu,1 BaoJin Wang1,2
1Department of Obstetrics and Gynecology, The Third Affiliated Hospital of Zhengzhou University, Zhengzhou, 450052, People’s Republic of China; 2Henan International Joint Laboratory of Ovarian Malignant Tumor, Zhengzhou, 450052, People’s Republic of China; 3Department of Pediatric Surgery, The Third Affiliated Hospital of Zhengzhou University, Zhengzhou, 450052, People’s Republic of China
Correspondence: BaoJin Wang
Department of Obstetrics and Gynecology, The Third Affiliated Hospital of Zhengzhou University, No. 7, Kangfu Front Street, Erqi District, Zhengzhou, 450052, People’s Republic of China
Tel +86 156 3927 7692
Fax +86 371-66903480
Email [email protected]
Purpose: Family with sequence similarity 83 (FAM83) is a newly discovered oncogene family, and the members of which can affect the prognosis of patients with malignant tumors via various mechanisms. However, the functions and molecular mechanisms of FAM83 genes in ovarian cancer (OC) have not yet been investigated. This study aimed to explore the clinical significance and prognostic value of FAM83 genes in OC.
Materials and Methods: We used a series of bioinformatics databases (Oncomine, GEPIA, cBioPortal, Kaplan–Meier plotter, DAVID and TIMER) to investigate the expression status, prognostic value, genetic alteration and biological function of all eight FAM83 genes in OC. In addition, a tissue microarray cohort (TMA) comprising 99 ovarian tumor tissues and 19 normal ovarian tissues was used to validate the protein expression and clinicopathological significance of FAM83H.
Results: Several datasets demonstrated the mRNA levels of FAM83A/D/E/F/H were significantly higher in OC compared with that in normal tissue. Moreover, the upregulation of FAM83D/H has been mutually confirmed in the Oncomine and GEPIA datasets. Kaplan–Meier survival analysis indicated that the FAM83D/H upregulation could predict poor prognosis of OC patients who had shorter overall survival (OS) and progression-free survival (PFS). In addition, cBioportal analysis indicated that the genetic alterations of FAM83 genes might affect the survival outcomes of patients with OC. Furthermore, KEGG analysis suggested that FAM83D/H are involved in the progression of OC through the cell cycle signaling pathway, and they had significant co-expression relationship with cell cycle-related genes. Finally, immunohistochemistry analysis confirmed the high expression of FAM83H protein in OC tissue, suggesting that its expression is positively correlated with the FIGO stage and pathological subtype of OC.
Conclusion: This study elucidated the expression status and prognostic value of FAM83 genes in OC and identified that FAM83D/H might be potential targets for the prognostic monitoring and targeted therapy of OC.
Keywords: FAM83s, ovarian cancer, prognosis, immunohistochemistry, biomarker
Introduction
Ovarian cancer (OC) is one of the most common gynecological malignancies and the leading cause of gynecological cancer-related worldwide, accounting for approximately 185,000 deaths annually.1 Although surgical resection, chemoradiotherapy and immune targeted therapy have remarkably improved the survival of patients with OC in recent decades, the prognosis of these patients remains unsatisfactory, with 5-year overall survival (OS) rates being less than 30%.2–4 The main reason for this situation is that the pathogenesis of OC is not yet completely clear.5
Cell signaling networks rigorously regulate numerous intracellular signaling pathways to precisely control the progression of cellular activities.6 When these important signaling pathways are disrupted, abnormal cellular changes may occur, leading to tumorigenesis.7 Therefore, the search for key factors involved in the intracellular regulatory networks of OC might proffer a theoretical basis for individualized diagnosis and treatment of OC. Recent evidence suggests that Family with sequence similarity 83 (FAM83) genes are aberrantly expressed in a variety of tumors and can facilitate aspects of their malignant progression, such as metastasis and drug resistance, leading to poor prognosis.8,9 Additionally, these genes play crucial roles in several classical signaling pathways, such as the EGFR, MAPK and PI3K/AKT pathways, all of which are closely related to the occurrence and development of OC.10,11 However, the relationship between FAM83 genes and OC remains unclear. Thus, an in-depth exploration of the biological significance of FAM83 genes in OC is warranted.
FAM83 is a newly discovered oncogene family consisting of eight genes (FAM83A–H), all of which have a highly conserved structural domain of unknown function (DUF1669) at their N-terminal ends.12 The first family members to be identified and studied were FAM83A and FAM83B. Lee et al showed that FAM83A overexpression could make breast cancer cells resistant to tyrosine kinase inhibitors.13 Meanwhile, Cipriano et al found that aberrant FAM83B expression induced carcinogenesis in breast epithelial cells by activating the RAS signaling pathway.14 These studies demonstrated for the first time that members of this newly identified family play pivotal roles in cancer-related signaling pathways and can promote malignant transformation of cells when their protein function is dysregulated. Subsequently, the oncogenic effects of FAM83 genes have been increasingly demonstrated in other cancer types, including OC. For instance, the upregulation of FAM83B in ovarian cancer cells increased their proliferation and growth rates and reduced the apoptosis rate.15 Mechanistically, Zhu et al and Zhang et al verified that FAM83D overexpression could promote malignant progression of OC via the EGFR pathway or the PI3K/AKT/mTOR signaling pathway.16,17 However, the clinical value and biological significance of other FAM83 genes in OC have rarely been reported.
To our knowledge, this is the first study to comprehensively and systematically identify the expression status and prognostic significance of FAM83 genes in OC by investigating several public databases. Besides that, FAM83D and FAM83H were identified, which could be potential biomarkers closely related to the malignant clinical features of OC, by immunohistochemistry (IHC) and clinicopathological analysis. In summary, the study provides a new viewpoint on the mechanisms of OC pathogenesis and novel targets for individualized diagnosis and treatment of OC.
Materials and Methods
Oncomine
Oncomine (https://www.oncomine.org/) is a mainstream public data-mining platform that contains gene expression data of 3606 OC samples and 12,764 normal samples for cancer genetic information analysis.18 In this study, we compared the mRNA expression of the eight FAM83 genes between OC tissue samples and normal tissue samples. The parameter-filtering criteria were as follows: p-value < 0.01, fold change > 1.5, gene rank in the top 10%, and data type: mRNA.
GEPIA
Gene Expression Profiling Interactive Analysis (GEPIA, http://gepia.cancer-pku.cn/) is a powerful bioinformatics analysis platform that contains extensive gene expression data of 9734 tumor samples and 8587 normal samples from TCGA and the GTEx projects.19 It has notable advantages in revealing differences in expression of target genes between cancer and normal tissue. Hence, we compared the mRNA expression levels of FAM83 genes between 426 OC and 88 normal ovarian tissue samples using GEPIA.
cBioPortal
The cBioportal (https://www.cbioportal.org/) database allows an in-depth investigation of tumor mechanisms from the DNA level, which greatly aids cancer researchers in revealing the relationship between molecular genomic profiles and clinical prognosis.20 We thoroughly explored the genetic alterations, such as amplification, mutation and deep deletion, in the eight FAM83 genes and investigated whether these gene alterations were associated with OS in patients with OC. In addition, the co-expression genes of FAM83D/H were selected from cBioPortal database for functional enrichment analysis based on a Spearman correlation coefficient of > 0.35 or < −0.35.
Kaplan–Meier Plotter
Kaplan–Meier plotter (https://kmplot.com/analysis/) is a professional online tool that can clarify the effect of a certain gene on cancer prognosis.21 We searched for FAM83s in the Kaplan–Meier plotter database to investigate their effect on OS and PFS in patients with OC. Considering their overexpression and significant relationship with poor prognosis, FAM83D and FAM83H were selected for further analysis in different subgroups with the aim to reveal the correlation of FAM83D/H with various clinicopathological factors.
GO Enrichment Analysis and KEGG Pathway Analysis
The Database for Annotation, Visualization and Integrated Discovery (DAVID, https://david.abcc.ncifcrf.gov/) is a well-known online biological annotation tool that can identify the most significantly enriched processes from a large-scale gene or protein expression dataset.22 We uploaded the FAM83D/H co-expression data into DAVID for Gene Ontology (GO) enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses. The results indirectly revealed the biological processes and signaling pathways in which that FAM83D/H are involved.
TIMER
The Tumor Immune Estimation Resource (TIMER, https://cistrome.shinyapps.io/timer/) database is a powerful tool to explore the relationship between gene expression and tumor immune infiltration.23 We used the TIMER algorithm to assess the association of the expression of FAM83D/H with the abundance of infiltrated immune cells (CD4+ cells, CD8+ cells, macrophages, neutrophils, B cells and dendritic cells) in the OC tumor microenvironment. The TIMER 2.0 (http://timer.comp-genomics.org/) was further used to examine the effect of different copy numbers of FAM83D/H on immune infiltration in OC.
Patients and Clinical Tissue Samples
In this study, we collected 118 paraffin-embedded tissue samples from patients who underwent gynecological surgery at the Third Affiliated Hospital of Zhengzhou University from January 2016 to May 2021. Informed consent was obtained from patients and their families, and this study was approved by the Ethics Committee of the Third Affiliated Hospital of Zhengzhou University (NO.202105401) in accordance with the ethical guidelines of the Declaration of Helsinki. Tumor tissue derived from 99 patients who underwent surgical treatment for ovarian cancer, and normal ovary tissue derived from 19 patients who underwent hysterectomy for uterine fibroids. Table S1 summarizes the clinicopathological characteristics of study participants.
Immunohistochemistry (IHC)
Tissue microarray blocks were prepared by first fixing tissue samples in 10% formaldehyde for 24 hours and then embedding in paraffin. Paraffin-embedded 4-μm-thick tissue slices were obtained and treated with 0.3% H2O2 to quench the activity of endogenous peroxidases and for antigen retrieval. The sections were incubated overnight with an anti-FAM83H antibody (Bioss, bs-16-18R) at 4°C and then with a secondary antibody at room temperature. Two senior pathologists independently analyzed the IHC staining results under a double-blind situation. The samples were scored according to the proportion of positive cells as follows: 0, 0–10%; 1, 10–29%; 2, 30–49%; 3, 50–74%; and 4, 75–100%. The staining intensity was evaluated as follows: 0, no staining; 1, weak; 2, medium; and 3, strong. The final immunostaining score (ranging from 0 to 12) was calculated by multiplying the two scores, and samples were classified into four levels based on their final scores: 1+ group, 0–3; 2+ group, 4–6; 3+ group, 7–9; and 4+ group, 10–12.
Statistical Analysis
GraphPad Prism 7.00 software was used to conduct statistical analysis. Differences between the two groups were estimated using Student’s t-test (unpaired, two-tailed). The Wilcoxon signed-rank test was used to analyze the relationships between FAM83H expression and cancer histology subtypes. OS and progression-free survival (PFS) were calculated using the Kaplan–Meier method. Spearman correlation analysis was used to measure the correlation between the expression of the two studied genes. In all analyses, p-value less than 0.05 was considered statistically significant.
Results
Expression Levels of FAM83 Members in OC
Abnormal activation and expression of oncogenes are crucial for malignant transformation of cells. A comprehensive understanding of the expression of FAM83 genes in multiple tumors is essential. As shown in Figure 1, abnormal expression of FAM83 genes, especially FAM83A, FAM83D and FAM83H, is observed in various cancers. Regarding OC, Oncomine analysis revealed that the mRNA expression of FAM83D/F/H was significantly higher in OC tissue than in normal ovarian tissue (p < 0.05) (Table 1 and Figure S1). In the Yoshihara Ovarian dataset, FAM83D mRNA expression in serous ovarian adenocarcinoma was 6.972 times higher than that in peritoneum tissue (p <0.0001). Similarly, the Lu Ovarian dataset demonstrated that FAM83D expression was significantly elevated in serous ovarian and endometrioid ovarian adenocarcinoma compared with that in normal ovarian surface epithelium (p = 6.90E−3, fold change = 1.705; p = 2.61E−2, fold change = 1.680). Moreover, the Yoshihara Ovarian dataset revealed that FAM83F mRNA expression was markedly increased by 4.497 times in serous ovarian adenocarcinoma in comparison with peritoneum tissue (3.00E−4). Significant FAM83H overexpression in serous ovarian adenocarcinoma tissue in comparison with that in ovarian surface epithelium tissue was also found in the Lu Ovarian dataset (p = 3.80E−3, fold change = 1.644).
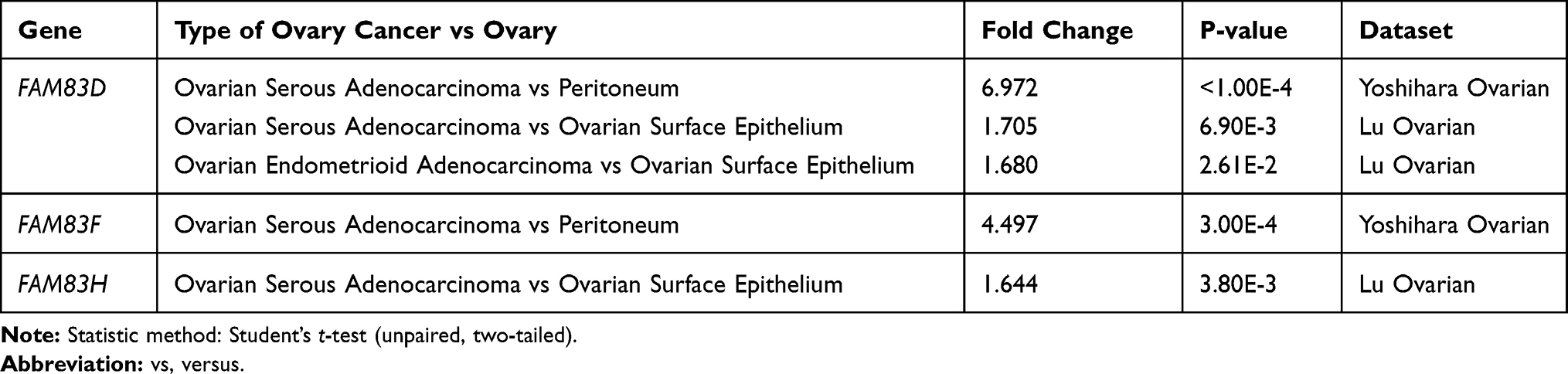 |
Table 1 FAM83s Expression in Different Type of Ovarian Cancer and Normal Ovarian Tissues (Oncomine) |
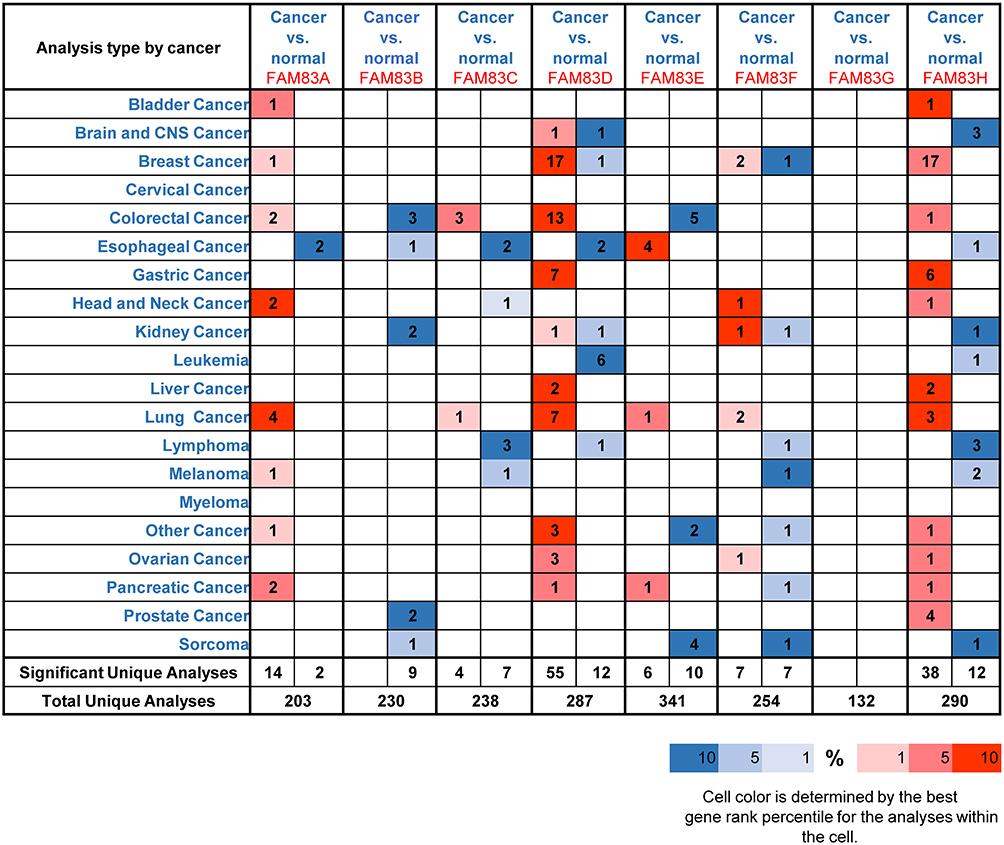 |
Figure 1 Transcriptional level of FAM83 family members in various types of cancer (ONCOMINE). Notes: The graph shows the numbers of datasets with statistically significant mRNA overexpression (red) or down-regulated expression (blue) of the target gene. The threshold was designed with following parameters: p-value <0.001, fold change >1.5 and gene rank in the Top 10%. The number on each cell represents the number of datasets that meet the filtering threshold. Cell color is determined by the best gene rank percentile for the analyses within the cell. Student’s t-test was used to identify the difference between two groups. |
Next, we used GEPIA to verify differences in the mRNA expression of FAM83 genes between OC samples (n = 426) and normal ovarian samples (n = 88). As demonstrated in the box plots in Figure 2, the expression levels of FAM83A/D/E/H were significantly increased, whereas those of FAM83B/C/F/G showed no difference in the OC samples compared with the normal samples. Taken together, the high expression of FAM83D/H in OC was confirmed in the two independent databases.
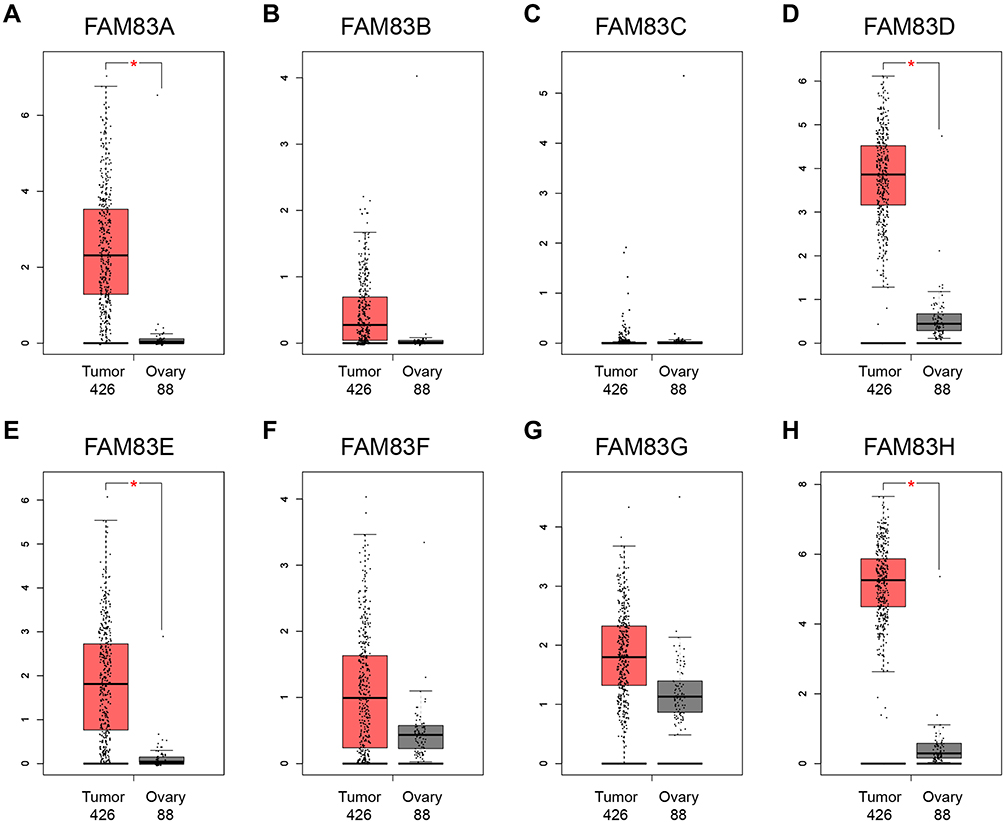 |
Figure 2 The mRNA expression levels of FAM83 family members in 426 ovarian cancer tissues and 88 normal ovary tissues (GEPIA) (A–H). Notes: *Indicates a statistically significant difference between the two groups; Student’s t-test was applied to identify the FAM83 genes expression difference between ovarian cancer and normal ovary tissue. |
Genetic Alterations of FAM83 Family Members in OC
To investigate the genetic alterations of FAM83 genes in OC, we selected 1680 ovarian serous cystadenocarcinoma cases from three datasets (606 cases from TCGA Firehose Legacy; 585 cases from TCGA PanCancer Atlas; and 489 cases from TCGA Nature 2011) for further analysis in the cBioPortal database. First, the genetic alteration rates of FAM83 genes in the three datasets were 46.83%, 39.26% and 38.87%, respectively (Figure 3A). Second, we found that all eight FAM83 genes showed different levels of genetic alterations. FAM83A/H showed the highest incidence of gene alterations among all FAM83 members (28% and 29%, respectively), while FAM83E/F/G presented fewer genetic changes. In terms of genetic variation types, the eight FAM83 genes exhibited five types of alteration, namely, amplification, mutation, deep deletion, multiple alteration and structural variation. As demonstrated in Figure 3B, amplification was the most frequent genetic alteration event in FAM83 genes, especially FAM83A/C/H. Moreover, deep deletion was found to account for a considerable proportion of alterations in FAM83E/F/G. Of note, only FAM83B and FAM83E genes showed multiple alterations and structural variations.
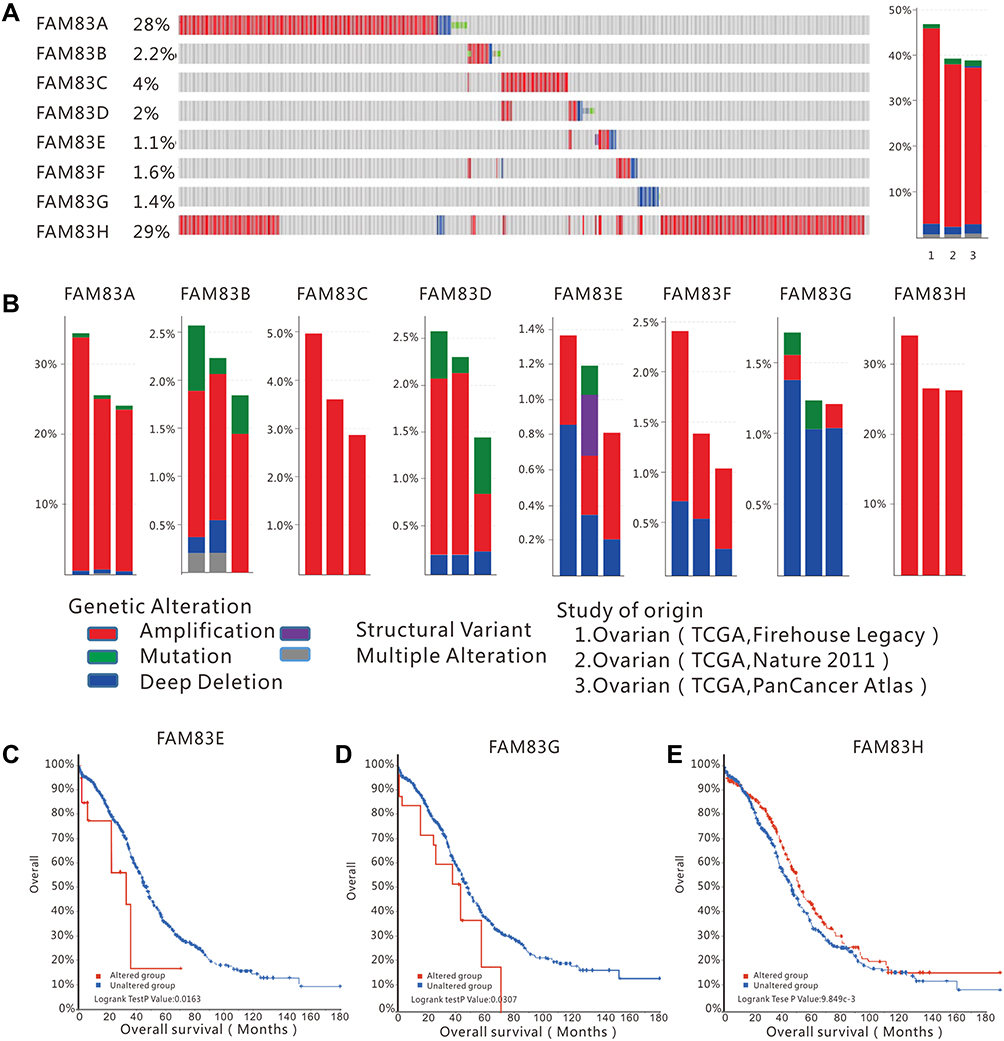 |
Figure 3 FAM83 family genes alterations in ovarian cancer (cBioportal). Notes: (A) Left: OncoPrint visual summary of alterations in FAM83s; right: summary of genetic alteration frequency in FAM83 genes. (B) Analysis of genetic alteration frequency in FAM83 family members in three datasets (TCGA Firehouse Legacy, TCGA Nature 2011 and TCGA PanCancer Atlas). The alterations included amplification, mutations, deep deletions, structural and multiple alterations. Kaplan–Meier analysis for overall survival in patients with or without FAM83 gens alterations, (C) FAM83E; (D) FAM83G and (E) FAM83H. |
Finally, the Kaplan–Meier method was applied to investigate the effect of genetic alterations of each FAM83 gene on the prognosis of patients with OC. Our results revealed that patients with alterations in FAM83E and FAM83G had shorter OS than those without any alterations (Figure 3C and D). In contrast, patients with FAM83H alterations had a favorable prognosis (Figure 3E). However, genetic alterations in the other FAM83 genes did not show a significant effect on prognosis (Figure S2). These results imply that genetic alterations of some FAM83 genes might directly affect the survival outcomes of patients with OC.
Prognostic Value of FAM83 Genes in Patients with OC
Increased expression of core oncogenes often leads to a decrease in survival time in cancer patients. In this study, we used the Kaplan–Meier method to clarify the impact of FAM83 gene expression levels on the survival of patients with OC. As shown in Figures 4 and 5, high expression of FAM83D/H were significantly correlated with poor OS and PFS, whereas high expression levels of FAM83B/E were significantly associated with favorable OS and PFS. No significant correlation was found between FAM83C/F expression and the survival time of patients with OC. Interestingly, FAM83A expression showed inconsistent associations with OS and PFS. Specifically, FAM83A upregulation had no effect on OS but was associated with shorter PFS. Survival analysis was not performed for FAM83G due to the lack of related survival data in the Kaplan–Meier plotter database.
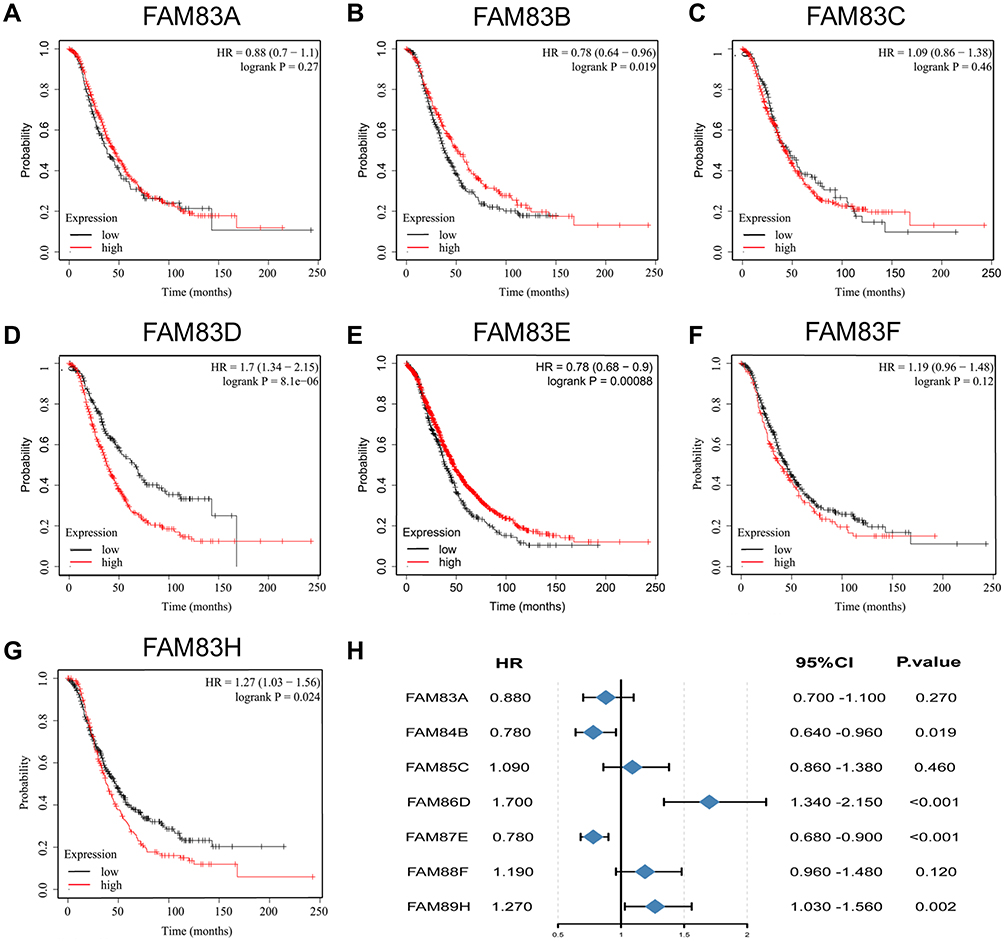 |
Figure 4 Prognostic value of FAM83s mRNA levels in patients with OC (OS in Kaplan–Meier plotter). Abbreviations: OC, ovarian cancer; OS, overall survival; HR, hazard ratio. Notes: (A–G) Prognostic significance of each FAM83 gene in OC. (H) Prognostic HRs of each FAM83 gene in OC. The red and black lines represent high and low expression, respectively. The Log rank test was used to calculate the p-value. |
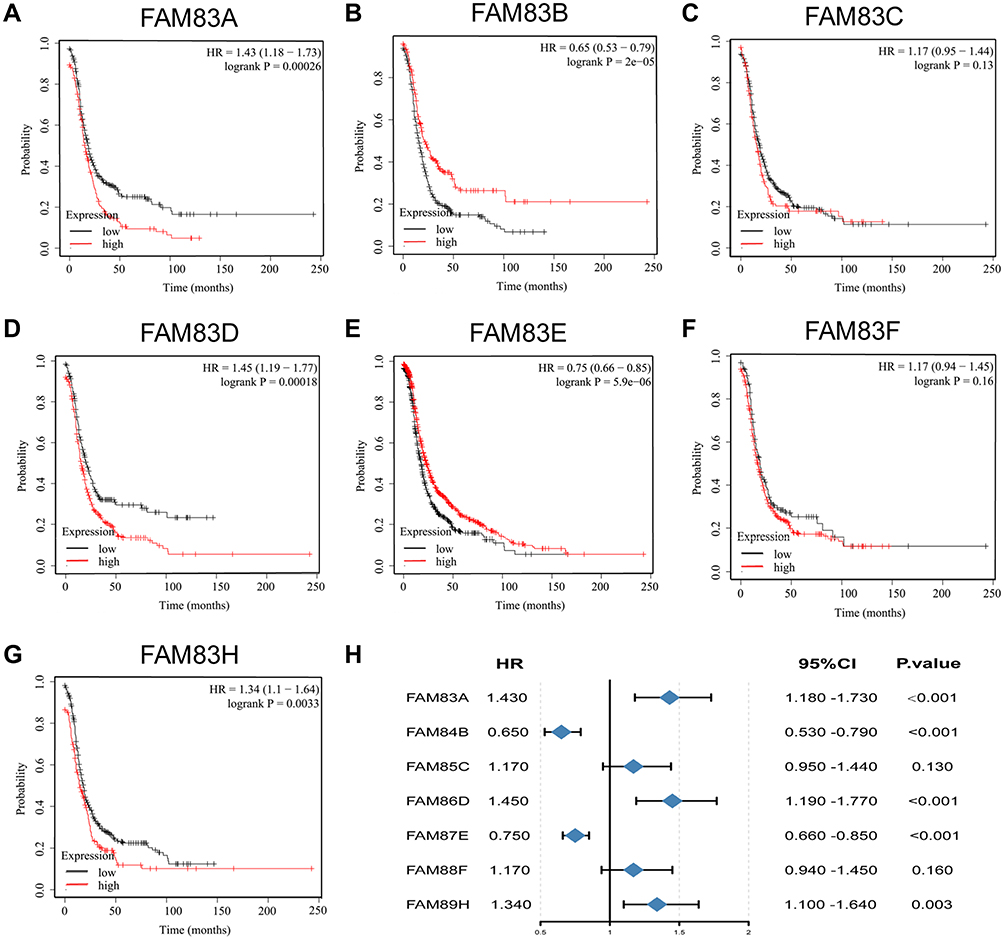 |
Figure 5 Prognostic value of FAM83s mRNA levels in patients with OC (PFS in Kaplan–Meier plotter). Abbreviations: OC, ovarian cancer; PFS, progression-free survival; HR, hazard ratio. Notes: (A–G) Prognostic significance of each FAM83 gene in OC. (H) Prognostic HRs of each FAM83 gene in OC. The red and black lines represent high and low expression, respectively. The Log rank test was used to calculate the p-value. |
Based on the overall results of Oncomine, GEPIA, cBioPortal and Kaplan–Meier plotter database analyses, FAM83D/H were selected for further analysis in diverse subgroups. We evaluated the prognostic value of FAM83D/H in different cancer histology subtypes, clinical stages, pathological grades and TP53 statuses. As shown in Table 2, increased FAM83D expression was correlated with poor OS in both the endometrioid and serous ovarian malignancy subgroups. In terms of the clinical stage, the OS of all grade patients showed a negative relationship with FAM83D expression. Additionally, FAM83D overexpression predicted short OS in the poor differentiation (Grade 3) subgroup. Similarly, the prognostic value of FAM83H was also found to be remarkably related to the above-mentioned clinical features.
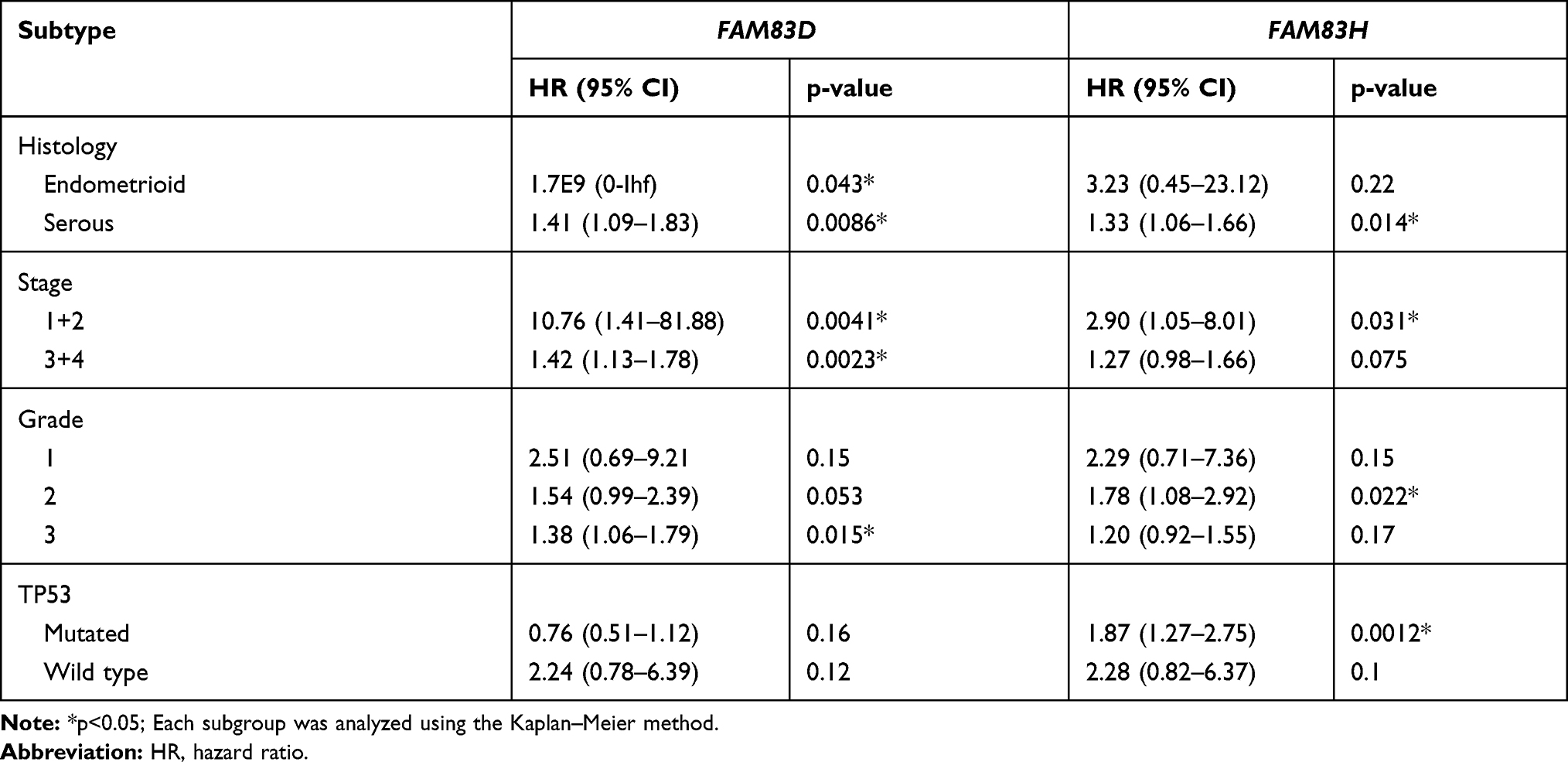 |
Table 2 The Prognostic Value of FAM83D and FAM83H in Different Subtypes |
Functional Enrichment Analysis of FAM83D and FAM83H in OC
Improved understanding of the biological functions of FAM83D/H may help to clarify their potential mechanisms in OC. To that end, we selected 353 genes and 491 genes that were closely related to FAM83D and FAM83H, respectively, according to Spearman correlation coefficients of > 0.35 or < −0.35, from the cBioPortal database. Then, we performed GO and KEGG function enrichment analyses of the associated genes in DAVID Bioinformatics Resources 6.8. FAM83D/H were found to be involved in multiple cancer-related signaling pathways and biological processes. As depicted in Figure 6A and B, genes relevant to FAM83D in OC were involved in cell cycle, DNA replication, the Fanconi anemia pathway, microRNAs in cancer, pathways in cancer, pyrimidine metabolism, the p53 signaling pathway, cell division, mitotic nuclear division, DNA repair, cell proliferation, and G1/S and G2/M transition of mitotic cell cycle. Figure 6C and D provided the significantly related pathways and biological processes of FAM83H co-expressed genes, including endocytosis, RNA transport, mRNA surveillance, cell cycle, basal transcription factors, transcription, DNA-templated, cell–cell adhesion, regulation of GTPase activity, cell division, mitotic nuclear division, protein SUMOylation, regulation of signal transduction by p53 class mediator and wound healing.
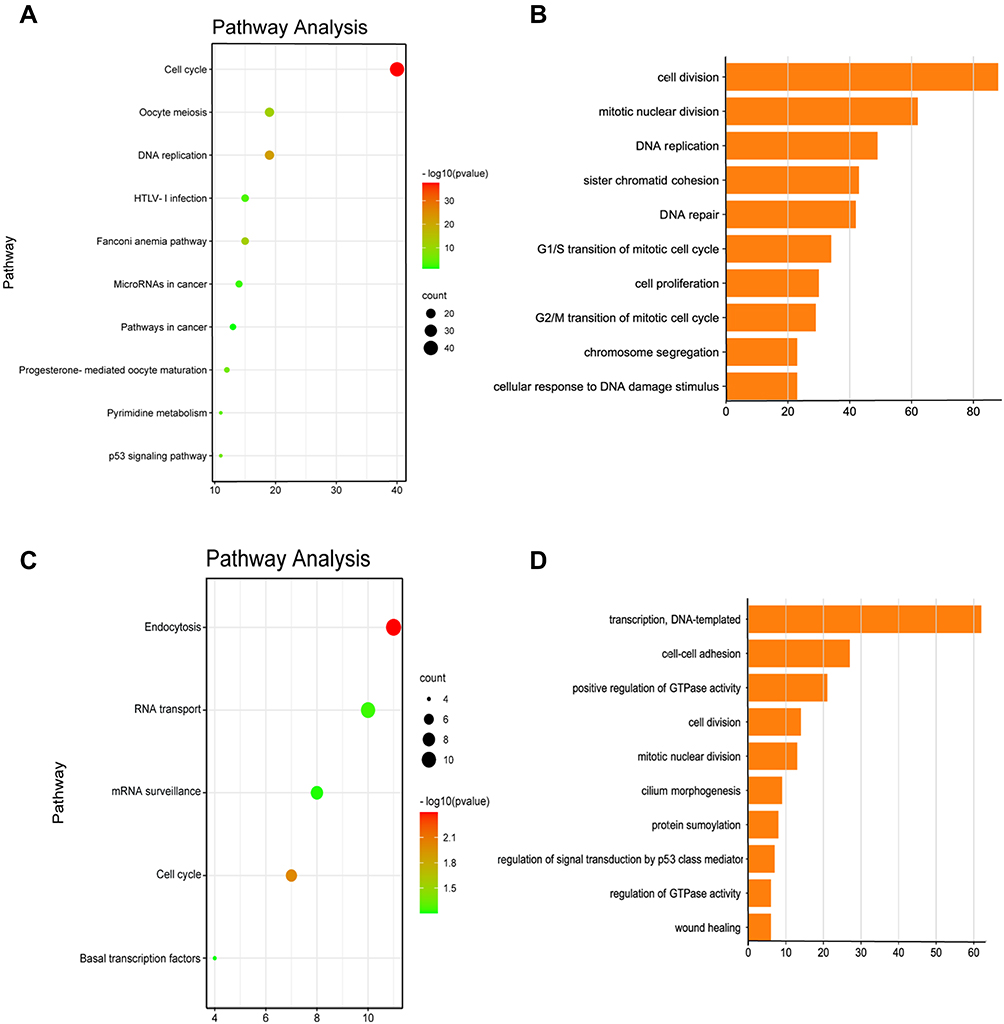 |
Figure 6 Functional enrichment analysis of FAM83D and FAM83H in ovarian cancer. Notes: (A) Significant pathway enrichment of FAM83D co-expression genes. (B) Significant biological process enrichment of FAM83D co-expressed genes. (C) Significant pathway enrichment of FAM83H co-expression genes. (D) Significant biological process enrichment of FAM83H co-expressed genes; statistic method: Spearman correlation analysis. |
Co-Expression Analysis of FAM83D and FAM83H in OC
KEGG analysis revealed that FAM83D/H are closely associated with the cell cycle signaling pathway, so we further explored their co-expression relationships with the 34 cell-cycle related genes that are pre-defined in the cBioportal database. The detailed information of the 34 genes is provided in Table S2. The co-expression analysis showed that FAM83D was positively associated with E2F8, CDK1, CCNB1, RBL1, E2F1, E2F7, E2F2, ADC25A, CDK2, CCNE1, CDKN2A, CDK4, CCNA1 and CDKN1B and negatively associated with MYC, CCND2, CDK6, RBL2 and CCND1 (Figure 7A). In addition, FAM83D was significantly and positively correlated with the genes encoding MKI67 and PCNA, two well-known tumor cell proliferation factors (Spearman correlation coefficients = 0.45 and 0.6, respectively; Figure S3). Meanwhile, FAM83H was positively associated with CCND1, MYC and JAK2 and negatively associated with CCNB1, CDK2, CDKN1B, E2F6 and CDK4 (Figure 7B).
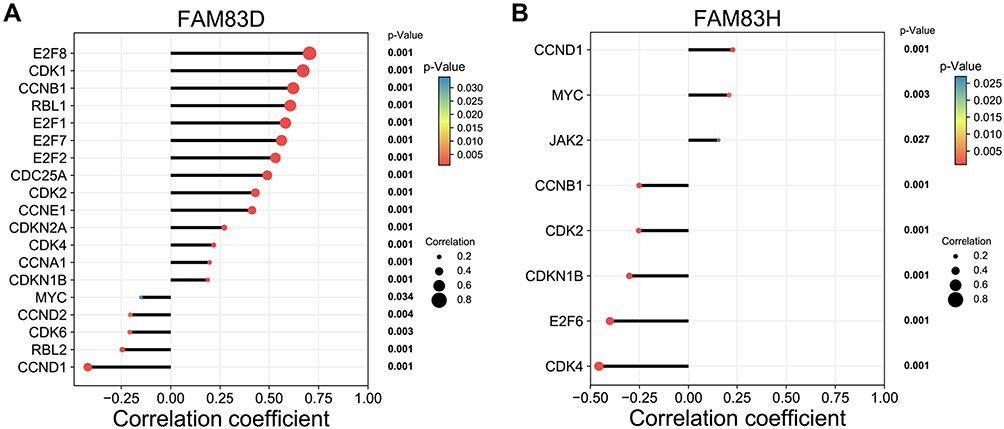 |
Figure 7 Co-expression analysis of FAM83D/H and cell cycle pathway-related genes. Notes: (A) Cell cycle-related genes significantly associated with FAM83D. (B) Cell cycle-related genes significantly associated with FAM83H. Statistic method: Spearman correlation analysis. |
FAM83D and FAM83H Related to Immune Infiltration in OC
The TIMER database was examined to estimate the association of FAM83D/H expression with immune cell infiltration. As shown in Figure 8A, FAM83D expression was negatively related to CD8+ T cell infiltration (r = −0.115, p = 1.2E−2), and positively related to CD4+ T cell infiltration (r = 0.1, p = 2.88E−2) and macrophage infiltration (r = 0.127, p = 5.24E−3). No correlation was detected between FAM83D expression and tumor cell purity or B cell, neutrophil or dendritic cell infiltration (Figure 8B. Meanwhile, FAM83H expression was positively associated with macrophage (r = −0.272, p = 1.34E−9), neutrophil (r = −0.121, p = 7.93E−3) and dendritic cell (r = −0.139, p = 2.33E−3) infiltration Figure 7B (Figure 8B). Compared with normal diploids, arm-level gain of FAM83D were negatively associated with the immune infiltration of B cells and myeloid dendritic cells, but positively associated with macrophage in OC (Figure 8C). Unlike FAM83D, the high amplification of FAM83H is positively related to the infiltration of B cells and myeloid dendritic cells, and the arm-level deletion of FAM83H is negatively related to the infiltration of CD8+ T cell (Figure 8D).
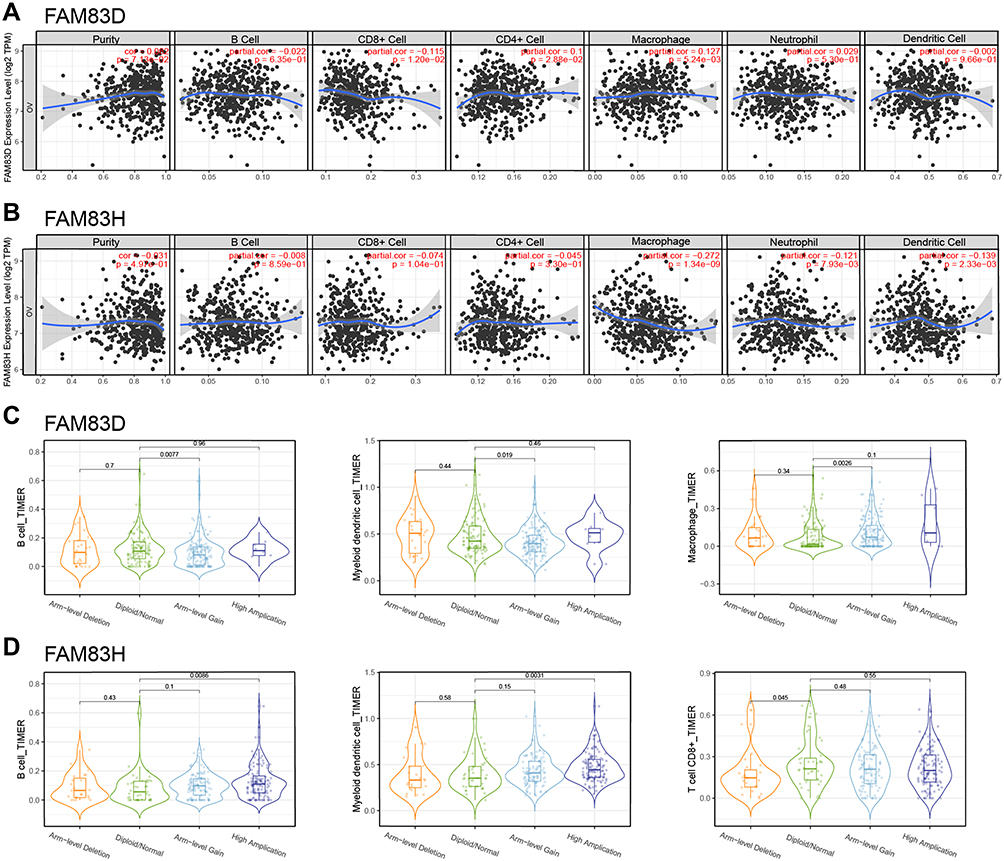 |
Figure 8 Correlation between FAM83D/H expression and immune infiltration. Notes: (A and B) Relationship between FAM83D/H expression and six immune cell infiltration including B cell, CD8+ T cell, CD4+ T cell, Macrophage, Neutrophil and Dendritic cell. (C and D) The relationship between somatic copy number alterations of FAM83D/H and immune cell infiltration; |
IHC and Clinicopathological Analysis of FAM83H in OC
There was no relevant literature yet reporting FAM83H in OC; therefore, we performed IHC analysis and clinicopathological analysis to reveal the role of FAM83H in OC. The IHC results presented in Figures 9 and S4 show that FAM83H protein was mainly localized in the cytoplasm, and its staining intensity was markedly higher in OC tissue than in normal ovarian tissue (Figure 9A, p < 0.001). Figure 9B presents four typical FAM83H protein-stained sections with scores of 1+, 2+, 3+ and 4+, demonstrating a clear trend of increasing protein expression. As shown in Figure 9C, the FIGO stage was associated with FAM83H protein expression, which was significantly upregulated in patients with stage III–IV comparison with stage I–II. There was no significant relationship between the expression of FAM83H protein and the pathological subtype and pathological T stage of OC. However, it is obvious that the expression of FAM83H was significantly higher in clear cell subtype than that in mucinous subtype.
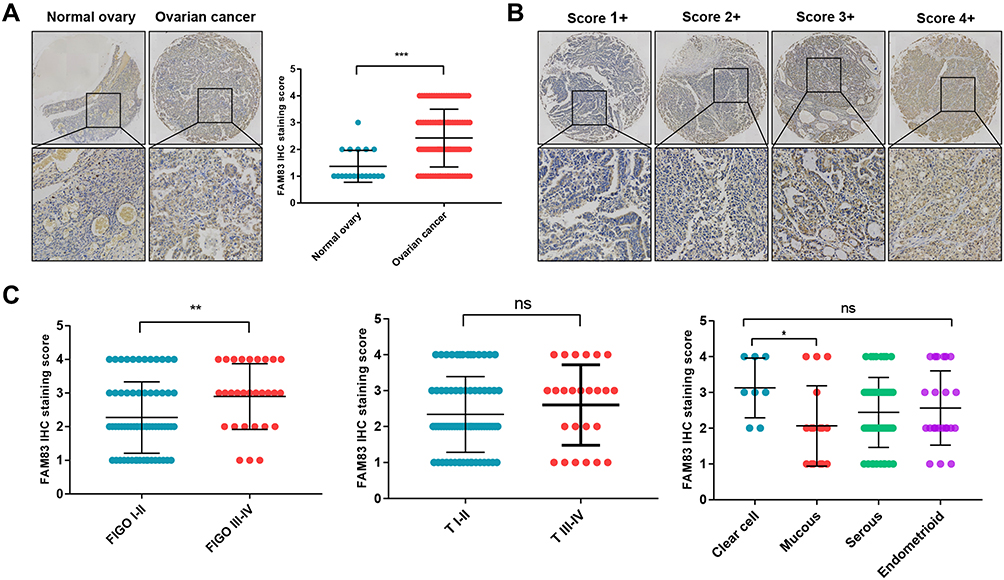 |
Figure 9 IHC analysis and clinicopathological analysis of FAM83H in OC. Abbreviations: OC, ovarian cancer; ns, not statistically significant. Notes: (A) Immunohistochemical detection of FAM83H in OC tissues. (B) Four typical immunohistochemical sections with score 1, 2, 3 and 4, respectively. (C) Association between the expression of FAM83H and different clinical features in OC; Student’s t-test (unpaired, two-tailed) or the Wilcoxon signed-rank test were used to analyze the relationships between FAM83H expression and different clinicopathological feature; *p<0.05; **p<0.01; ***p<0.001. |
Discussion
Although an increasing number of studies have demonstrated the critical regulatory role of FAM83 genes in various tumors, their prognostic value and functional mechanisms in OC have not yet been fully elucidated. In this study, we performed a systematic assessment of the biological significance of FAM83 genes in OC using both the bioinformatics data from several authoritative databases and the clinical data of patients with OC from our hospital. To our knowledge, this is the first study to reveal the expression status and prognostic significance of FAM83 genes in OC, providing a novel insight into the pathological mechanism of OC.
Apart from the conserved globular structural domain DUF1669 located at the N terminus, no other similar sequences were detected between the FAM83 genes, which partly explains the functional differences between these family members.8,12 FAM83A was first identified as a driver of tyrosine kinase inhibitor (TKI) resistance by genetic analysis and as a tumor-specific biomarker for lung cancer; it significantly accelerates the progression of lung cancer though the Wnt and Hippo Signaling pathways.13 FAM83B overexpression was found to activate the PI3K/AKT pathway and reduce cell sensitivity to inhibitors of small molecules such as PI3K, AKT and mTOR, thus promoting tumor development and chemoresistance.14 FAM83D, which encodes a mitotic spindle-associated protein, is a candidate oncogene for tumorigenesis. FAM83D knockdown has been reported to downregulate expression of the tumor suppressor gene FBXW7, which induces apoptosis and inhibits cell proliferation and clone formation in colorectal cancer cells.24 FAM83G is involved in the regulation of the BMP signaling pathway, and its protein product is a substrate for type I BMP receptor kinase.25,26 The function of FAM83H was first identified in calcium-deficient enamel hypoplasia and is closely related to the thickness, density and morphology of enamel in mice.27 However, the potential biological functions of FAM83C, FAM83E and FAM83F have not yet been reported. Given their crucial roles in tumor progression, a systematic investigation of the roles of FAM83 family genes in OC is warranted.
In this study, we first explored the similarities and differences in the transcriptome levels of FAM83 family members in different tumor types by pan-cancer analysis. The results of Oncomine analysis demonstrated that aberrant expression of FAM83 genes (especially FAM83D, FAM83F and FAM83H) was universal among various cancer types. Their upregulation and oncogenic effects have been revealed by previous researchers. For instance, Hu et al indicated that FAM83A expression was increased in non-small-cell lung cancer (NSCLC), which significantly promoted cancer cell proliferation and protected against apoptosis via the ERK and PI3K/Akt/mTOR signaling pathways.28 FAM83B expression was notably elevated in tumor tissue and cell lines of pancreatic ductal adenocarcinoma, and it increased as the advanced clinical stage and poor vital status.29 Gan et al showed that FAM83C was upregulated in NSCLC compared with normal lung tissue, and that higher FAM83C expression was remarkably associated with shorter OS and PFS in patients with NSCLC.30 FAM83D, which is currently a hot topic in cancer research, has been shown to be upregulated in hepatocellular cancer, gastric cancer, breast cancer and colorectal cancer.24,31–33 Likewise, FAM83F and FAM83H have been reported to be highly expressed in papillary thyroid cancer and pancreatic cancer, respectively.34,35 Of note, few studies have investigated the expression of FAM83E and FAM83G in tumors. In agreement with previous studies, our Oncomine analysis results suggested that FAM83D, FAM83F and FAM83H are highly expressed in OC. Notably, FAM83 genes, specifically FAM83D and FAM83H, show different or even opposite expression status in different tumor types. As the human body is a vast and complex system, the roles of FAM83 genes in tumorigenesis might differ according to the type of tissue or cancer.36
To further determine the expression of FAM83 genes in OC, we compared their mRNA expression levels between 426 OC samples and 88 normal ovarian tissue sample in GEPIA. The results suggested that FAM83A, FAM83D, FAM83E and FAM83H were significantly upregulated in OC. Notably, FAM83D upregulation has been demonstrated in former experimental studies. For example, Zhu et al showed that FAM83D was highly expressed in OC and markedly enhanced the invasion and proliferation of cancer cells.16 Moreover, Zhang et al identified FAM83D as an excellent biomarker to distinguish between invasive epithelial ovarian cancer and low malignant potential ovarian tumor owing to its aberrant expression and tendency to promote malignancy.17 However, few studies have investigated the expression of FAM83A, FAM83E and FAM83H in OC, and this is the first study to identify overexpression of these genes through big data mining. Our findings may inform future research on the pathogenesis of OC.
Genetic alterations, such as CNVs and mutations, are the main drivers of gene-specific expression. To confirm the association of alterations of FAM83 genes with OC, we conducted a genetic analysis in the cBioPortal database, which revealed that the overall alteration rate in FAM83 genes ranged from 38.87% to 46.83% across three different OC datasets. The most common genetic changes in the FAM83 family were the amplification of FAM83A and FAM83H. The high amplification rate of FAM83A/H and overexpression of FAM83A/H represented amazing consistency in OC. Moreover, the Kaplan–Meier survival curve indicated that the patients with FAM83E or FAM83G alteration had longer OS than patients without any alteration, while those with FAM83H alteration had shorter OS; this suggests that alterations in these FAM83 genes directly affect the prognosis of patients with OC.
Given the prevalence of abnormalities in FAM83 expression at the transcriptome and genome levels, it is necessary to further explore the influence of abnormal expression of these genes on OC prognosis. Our Kaplan–Meier survival analysis showed that patients in the high FAM83B/E expression group had longer OS and PFS than those in the low FAM83B/E expression group, which suggests that FAM83B and FAM83E are protective factors that possibly function as tumor suppressor genes in OC. Mechanically, one study affirmed that FAM83B could inhibit the Wnt signaling pathway and thereby inhibit cisplatin resistance in OC, which further explains why patients with low FAM83B expression have a poor prognosis.15 In contrast, FAM83D and FAM83H were found to be risk factors for poor prognosis and may thus function as oncogenes. The oncogenic role of FAM83D in OC has been demonstrated by previous studies.16,17 However, the mechanism by which FAM83E and FAM83H promote or suppress the occurrence and development of OC has yet not been revealed, which warrants further investigation.
Taken together, the results of our Oncomine, GEPIA and Kaplan–Meier plotter database analyses (Table 3) indicated that the expression of FAM83D and FAM83H is significantly associated with OC prognosis. Hence, we had reason to believe that FAM83D/H may be key FAM83 family genes involved in promoting OC progression. We therefore further explored the potential mechanisms of their involvement in the pathogenesis of OC. First, KEGG pathway analysis showed that FAM83D/H were closely related to the cell cycle pathway. Next, GO enrichment analysis indicated that multiple cell cycle-related biological processes were also closely associated with FAM83D/H. Further co-expression analysis showed that many cell cycle-related genes were also significantly associated with FAM83D/H. Thus, we speculated that FAM8383D/H is involved in the malignant progression of OC through the cell cycle signaling pathway. Likewise, a strong correlation between FAM83D and the cell cycle has been demonstrated in human lung adenocarcinoma and hepatocellular carcinoma.31,37 In terms of the specific mechanism of FAM83H in the pathological progression of OC, previous studies have indicated that the oncogene MYC can enhance the proliferation of hepatocellular carcinoma cells by regulating the transcription of FAM83H.38 In addition, FAM83H expression is closely associated with β-catenin activity, and immunoprecipitation experiments showed that FAM83H can interact directly with β-catenin.39 Although few studies have reported the oncogenic function of FAM83H in OC to date, our results showed that the mRNA and protein expression of FAM83H in OC was higher than that in normal tissues and was positively associated with malignant clinical features. Our results, together with previous evidence, suggest that FAM83D and FAM83H may be promising biomarkers of OC.
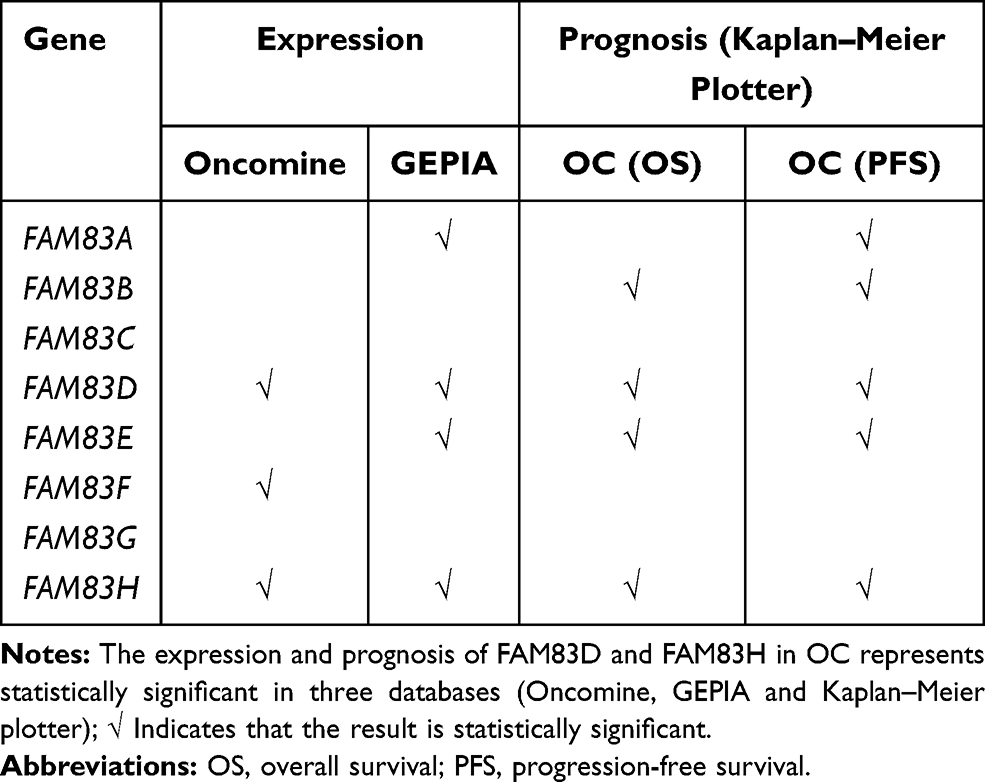 |
Table 3 Summary of Bioinformation Analysis of FAM83 Genes in Ovarian Cancer |
A comprehensive evaluation of the type, number and distribution of immune infiltrating cells in the tumor microenvironment would enable us to gain a deeper understanding of the interaction between the immune system and OC and ultimately reveal the immune escape mechanism of this cancer.40 As a special heterogeneous group, tumor-infiltrating lymphocytes (TILs) play a crucial role in tumor immunity. CD4+ T cells secrete large amounts of inflammatory cytokines, such as IFN-γ and TGF-β, and CD4+ regulatory T cells are essential in maintaining immune tolerance.41 Chen et al demonstrated that TIGIT, a well-known immune checkpoint, could enhance CD4+ regulatory T cell responses and mediate immunosuppression in a murine OC model.42 Meanwhile, the presence of CD8+ TILs was correlated with a favorable prognosis in patients with epithelial OC.43 Interestingly, our results showed that CD4+ T cells were positively correlated with FAM83D, and CD8+ T cells were negatively correlated with FAM83D. Therefore, we speculated that the increase in the CD4+/CD8+ T cell ratio might be suggestive of dysregulation of the immune status of the organism caused by high expression of FAM83D. Obviously, there was a negative association between FAM83H expression and the infiltrated abundance of six types of immune cells. Immune and inflammatory cells are vital components of the tumor microenvironment, and their reduction and deletion are known to be closely associated with the proliferation, invasion, adhesion, angiogenesis, drug resistance and other malignant processes of tumors.44 In general, we first explore the relationship between local immune status and FAM83D and FAM83H in ovarian cancer to provide a basis for immunotherapy in ovarian cancer.
In this work, we revealed close relationships between FAM83 genes and OC. However, some unavoidable limitations of our analysis must be considered. For instance, raw data were processed and analyzed in different open-access databases, which may cause inconsistent results or statistical deviations. However, the incomparable strengths of public databases are that large amounts of valuable data can be acquired in a short time and that the integrated results generated are highly credible and applicable.45 In the future, we plan to conduct in-depth investigation of the specific mechanisms of FAM83 genes in OC pathogenesis.
Conclusion
In conclusion, this study provides the first systematic insight into the aberrant expression of FAM83 family genes in OC and their differential effects on OC prognosis. Furthermore, IHC and clinicopathological analysis validated the overexpression of FAM83H in OC and revealed its significant association with the malignant progression of OC. Our results suggest that FAM83D and FAM83H might be promising prognostic biomarkers for OC and potential novel targets for OC immunotherapy.
Acknowledgments
This work was supported by the Supported by Science and Technology Tackling Project of Henan Province (No. 192102310067) and Henan International Joint Laboratory of Ovarian Malignant Tumor.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Kurnit KC, Fleming GF, Lengyel E. Updates and new options in advanced epithelial ovarian cancer treatment. Obstet Gynecol. 2021;137(1):108–121. doi:10.1097/AOG.0000000000004173
3. Shimizu A, Sawada K, Kimura T. Pathophysiological role and potential therapeutic exploitation of exosomes in ovarian cancer. Cells. 2020;9(4):814.
4. Rottmann M, Burges A, Mahner S, et al. Cancer of the ovary, fallopian tube, and peritoneum: a population-based comparison of the prognostic factors and outcomes. J Cancer Res Clin Oncol. 2017;143(9):1833–1844. doi:10.1007/s00432-017-2422-6
5. Salamini-Montemurri M, Lamas-Maceiras M, Barreiro-Alonso A, et al. The challenges and opportunities of LncRNAs in ovarian cancer research and clinical use. Cancers. 2020;12(4):1020. doi:10.3390/cancers12041020
6. Hongdusit A, Liechty ET, Fox JM. Optogenetic interrogation and control of cell signaling. Curr Opin Biotechnol. 2020;66:195–206. doi:10.1016/j.copbio.2020.07.007
7. Wu J, Jiang H, Luo S, et al. Caspase-mediated cleavage of C53/LZAP protein causes abnormal microtubule bundling and rupture of the nuclear envelope. Cell Res. 2013;23(5):691–704. doi:10.1038/cr.2013.36
8. Bozatzi P, Sapkota GP. The FAM83 family of proteins: from pseudo-PLDs to anchors for CK1 isoforms. Biochem Soc Trans. 2018;46(3):761–771. doi:10.1042/BST20160277
9. Snijders AM, Lee SY, Hang B, Hao W, Bissell MJ, Mao JH. FAM83 family oncogenes are broadly involved in human cancers: an integrative multi-omics approach. Mol Oncol. 2017;11(2):167–179. doi:10.1002/1878-0261.12016
10. Cipriano R, Miskimen KL, Bryson BL, Foy CR, Bartel CA, Jackson MW. Conserved oncogenic behavior of the FAM83 family regulates MAPK signaling in human cancer. Mol Cancer Res. 2014;12(8):1156–1165. doi:10.1158/1541-7786.MCR-13-0289
11. Liu C, Huang XY, Huang Y. FAM83G promotes proliferation, invasion, and metastasis by regulating PI3K/AKT signaling in hepatocellular carcinoma cells. Biochem Biophys Res Commun. 2021;567:63–71. doi:10.1016/j.bbrc.2021.05.081
12. Fulcher LJ, Bozatzi P, Tachie-Menson T, et al. The DUF1669 domain of FAM83 family proteins anchor casein kinase 1 isoforms. Sci Signal. 2018;11(531):531. doi:10.1126/scisignal.aao2341
13. Lee SY, Meier R, Furuta S, et al. FAM83A confers EGFR-TKI resistance in breast cancer cells and in mice. J Clin Invest. 2012;122(9):3211–3220. doi:10.1172/JCI60498
14. Cipriano R, Graham J, Miskimen KL, et al. FAM83B mediates EGFR- and RAS-driven oncogenic transformation. J Clin Invest. 2012;122(9):3197–3210. doi:10.1172/JCI60517
15. He S, Wang W, Wan Z, et al. FAM83B inhibits ovarian cancer cisplatin resistance through inhibiting Wnt pathway. Oncogenesis. 2021;10(1):6. doi:10.1038/s41389-020-00301-y
16. Zhu H, Diao S, Lim V, Hu L, Hu J. FAM83D inhibits autophagy and promotes proliferation and invasion of ovarian cancer cells via PI3K/AKT/mTOR pathway. Acta Biochim Biophys Sin. 2019;51(5):509–516. doi:10.1093/abbs/gmz028
17. Zhang Q, Yu S, Lok SIS, Wong AST, Jiao Y, Lee LTO. FAM83D promotes ovarian cancer progression and its potential application in diagnosis of invasive ovarian cancer. J Cell Mol Med. 2019;23(7):4569–4581. doi:10.1111/jcmm.14360
18. Rhodes DR, Kalyana-Sundaram S, Mahavisno V, et al. Oncomine 3.0: genes, pathways, and networks in a collection of 18,000 cancer gene expression profiles. Neoplasia. 2007;9(2):166–180. doi:10.1593/neo.07112
19. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–W102. doi:10.1093/nar/gkx247
20. Cao T, Pan W, Sun X, Shen H. Increased expression of TET3 predicts unfavorable prognosis in patients with ovarian cancer-a bioinformatics integrative analysis. J Ovarian Res. 2019;12(1):101. doi:10.1186/s13048-019-0575-4
21. Guo L, Fang T, Jiang Y, Liu D. Identification of immune checkpoint inhibitors and biomarkers among STAT family in stomach adenocarcinoma. Am J Transl Res. 2020;12(9):4977–4997.
22. Yin X, Zhang X, Liu Z, et al. Assessment for prognostic value of differentially expressed genes in immune microenvironment of clear cell renal cell carcinoma. Am J Transl Res. 2020;12(9):5416–5432.
23. Zhao R, Peng C, Song C, et al. BICC1 as a novel prognostic biomarker in gastric cancer correlating with immune infiltrates. Int Immunopharmacol. 2020;87:106828. doi:10.1016/j.intimp.2020.106828
24. Mu Y, Zou H, Chen B, Fan Y, Luo S. FAM83D knockdown regulates proliferation, migration and invasion of colorectal cancer through inhibiting FBXW7/Notch-1 signalling pathway. Biomed Pharmacother. 2017;90:548–554. doi:10.1016/j.biopha.2017.03.073
25. Bozatzi P, Dingwell KS, Wu KZ, et al. PAWS1 controls Wnt signalling through association with casein kinase 1alpha. EMBO Rep. 2018;19(4):e44807. doi:10.15252/embr.201744807.
26. Cummins TD, Wu KZL, Bozatzi P, et al. PAWS1 controls cytoskeletal dynamics and cell migration through association with the SH3 adaptor CD2AP. J Cell Sci. 2018;131(1):jcs202390.
27. Kweon YS, Lee KE, Ko J, Hu JC, Simmer JP, Kim JW. Effects of Fam83h overexpression on enamel and dentine formation. Arch Oral Biol. 2013;58(9):1148–1154. doi:10.1016/j.archoralbio.2013.03.001
28. Hu H, Wang F, Wang M, et al. FAM83A is amplified and promotes tumorigenicity in non-small cell lung cancer via ERK and PI3K/Akt/mTOR pathways. Int J Med Sci. 2020;17(6):807–814. doi:10.7150/ijms.33992
29. Shen CQ, Yan TT, Liu W, et al. High expression of FAM83B predicts poor prognosis in patients with pancreatic ductal adenocarcinoma and correlates with cell cycle and cell proliferation. J Cancer. 2017;8(16):3154–3165. doi:10.7150/jca.20086
30. Gan J, Li Y, Meng Q. Systematic analysis of expression profiles and prognostic significance for FAM83 family in non-small-cell lung cancer. Front Mol Biosci. 2020;7:572406. doi:10.3389/fmolb.2020.572406
31. Wang D, Han S, Peng R, et al. FAM83D activates the MEK/ERK signaling pathway and promotes cell proliferation in hepatocellular carcinoma. Biochem Biophys Res Commun. 2015;458(2):313–320. doi:10.1016/j.bbrc.2015.01.108
32. Wang F, Zhang S, Wei Y, Chen H, Jiao Z, Li Y. Upregulation of family with sequence similarity 83 member D expression enhances cell proliferation and motility via activation of Wnt/beta-catenin signaling and predicts poor prognosis in gastric cancer. Cancer Manag Res. 2019;11:6775–6791. doi:10.2147/CMAR.S203082
33. Zhai X, Yang Z, Liu X, Dong Z, Zhou D. Identification of NUF2 and FAM83D as potential biomarkers in triple-negative breast cancer. PeerJ. 2020;8:e9975. doi:10.7717/peerj.9975
34. Fuziwara CS, Saito KC, Leoni SG, Waitzberg AFL, Kimura ET. The highly expressed FAM83F protein in papillary thyroid cancer exerts a pro-oncogenic role in thyroid follicular cells. Front Endocrinol (Lausanne). 2019;10:134. doi:10.3389/fendo.2019.00134
35. Zhuang H, Zhang C, Hou B. FAM83H overexpression predicts worse prognosis and correlates with less CD8(+) T cells infiltration and Ras-PI3K-Akt-mTOR signaling pathway in pancreatic cancer. Clin Transl Oncol. 2020;22(12):2244–2252. doi:10.1007/s12094-020-02365-z
36. Kim KM, Hussein UK, Bae JS, et al. The expression patterns of FAM83H and PANX2 are associated with shorter survival of clear cell renal cell carcinoma patients. Front Oncol. 2019;9:14. doi:10.3389/fonc.2019.00014
37. Shi R, Sun J, Sun Q, et al. Upregulation of FAM83D promotes malignant phenotypes of lung adenocarcinoma by regulating cell cycle. Am J Cancer Res. 2016;6(11):2587–2598.
38. Kim KM, Park SH, Bae JS, et al. FAM83H is involved in the progression of hepatocellular carcinoma and is regulated by MYC. Sci Rep. 2017;7(1):3274. doi:10.1038/s41598-017-03639-3
39. Kim KM, Hussein UK, Park SH, et al. FAM83H is involved in stabilization of beta-catenin and progression of osteosarcomas. J Exp Clin Cancer Res. 2019;38(1):267. doi:10.1186/s13046-019-1274-0
40. An Y, Yang Q. Tumor-associated macrophage-targeted therapeutics in ovarian cancer. Int J Cancer. 2021;149(1):21–30. doi:10.1002/ijc.33408
41. La Rocca C, Carbone F, Longobardi S, Matarese G. The immunology of pregnancy: regulatory T cells control maternal immune tolerance toward the fetus. Immunol Lett. 2014;162(1 Pt A):41–48. doi:10.1016/j.imlet.2014.06.013
42. Chen F, Xu Y, Chen Y, Shan S. TIGIT enhances CD4(+) regulatory T-cell response and mediates immune suppression in a murine ovarian cancer model. Cancer Med. 2020;9(10):3584–3591. doi:10.1002/cam4.2976
43. Sato E, Olson SH, Ahn J, et al. Intraepithelial CD8+ tumor-infiltrating lymphocytes and a high CD8+/regulatory T cell ratio are associated with favorable prognosis in ovarian cancer. Proc Natl Acad Sci USA. 2005;102(51):18538–18543. doi:10.1073/pnas.0509182102
44. Harjunpaa H, Llort Asens M, Guenther C, Fagerholm SC. Cell adhesion molecules and their roles and regulation in the immune and tumor microenvironment. Front Immunol. 2019;10:1078.
45. Liu Z, Shen F, Wang H, et al. Abnormally high expression of HOXA2 as an independent factor for poor prognosis in glioma patients. Cell Cycle. 2020;19(13):1632–1640. doi:10.1080/15384101.2020.1762038
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.