")
Back to Journals » International Journal of General Medicine » Volume 14
Identification of Hub Genes in Patients with Alzheimer Disease and Obstructive Sleep Apnea Syndrome Using Integrated Bioinformatics Analysis
Authors Wu L , Wang W , Tian S , Zheng H , Liu P, Wu W
Received 7 October 2021
Accepted for publication 23 November 2021
Published 9 December 2021 Volume 2021:14 Pages 9491—9502
DOI https://doi.org/10.2147/IJGM.S341078
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Lanxiang Wu,1 Wenjun Wang,2 Sheng Tian,1 Heqing Zheng,1 Pan Liu,1 Wei Wu1
1Department of Neurology, The Second Affiliated Hospital of Nanchang University, Nanchang, Jiangxi, 330006, People’s Republic of China; 2Department of Respiratory Diseases, The Second Affiliated Hospital of Nanchang University, Nanchang, Jiangxi, 330006, People’s Republic of China
Correspondence: Wei Wu
Department of Neurology, The Second Affiliated Hospital of Nanchang University, 1 Minde Road, Nanchang, Jiangxi, 330006, People’s Republic of China
Tel +86-13807038803
Fax +86-07916300507
Email [email protected]
Background: Obstructive sleep apnea syndrome (OSA) is associated with an increased risk of Alzheimer’s disease (AD). This study aimed to identify the key common genes in AD and OSA and explore molecular mechanism value in AD.
Methods: Expression profiles GSE5281 and GSE135917 were acquired from Gene Expression Omnibus (GEO) database, respectively. Weighted gene co-expression network analysis (WGCNA) and R 4.0.2 software were used for identifying differentially expressed genes (DEGs) related to AD and OSA. Function enrichment analyses using Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway, and the protein–protein interaction network (PPI) using the STRING database were subsequently performed on the shared DEGs. Finally, the hub genes were screened from the PPI network using the MCC algorithm of CytoHubba plugin.
Results: Seven modules and four modules were the most significant with AD and OSA by WGCNA, respectively. A total of 33 common genes were screened in AD and OSA by VENN. Functional enrichment analysis indicated that DEGs were mainly involved in cellular response to oxidative stress, neuroinflammation. Among these DEGs, the top 10 hub genes (high scores in cytoHubba) were selected in the PPI network, including AREG, SPP1, CXCL2, ITGAX, DUSP1, COL1A1, SCD, ACTA2, CCND2, ATF3.
Conclusion: This study presented ten target genes on the basis of common genes to AD and OSA. These candidate genes may provide a novel perspective to explore the underlying mechanism that OSA leads to an increased risk of AD at the transcriptome level.
Keywords: Alzheimer’s disease, AD, obstructive sleep apnea, weighted gene co-expression network analysis, WGCNA, hub genes
Introduction
Alzheimer’s disease (AD) ranks first among the common dementia type of the world. According to epidemiological investigation from the International Alzheimer’s disease association, about 45 million people has been suffered from AD, and the number is expected to increase to 131 million in 2050.1 Despite the widespread prevalence of AD, the potential mechanism remains elusive, and current managements are limited. It brings not only a serious medical economic burden but also social pressure. In recent years, with the rapid economic development and population aging, the prevalence of Obstructive sleep apnea syndrome (OSA) is on the rise in numerous countries. OSA is a complex, multifactorial disease affecting millions of people worldwide, and it may lead to multiple serious complications such as cardiovascular incidences, cognitive dysfunction, and metabolic syndromes. OSA characterized with intermittent hypoxemia, hypercapnia, inflammation, oxidative stress and sleep fragmentation, is a major type of sleep disorders.2 The symptoms and treatments of OSA vary according to severity. Therefore, early diagnosis and management of OSA are critical for promoting treatments and preventing complications.
AD and OSA are regarded as age-related diseases. Epidemiological investigations have indicated that the prevalence of OSA was 84% in the elderly, and 35.2% of OSA patients were accompanied with cognitive dysfunction.3 AD and OSA belong to two different systems, but more and more studies have demonstrated that these two diseases interact with each other and share some pathological mechanisms. The correlation of AD with OSA is gradually gaining research attention. Mild cognitive impairment and AD were more possible to be found in patients with OSA.4 AD is a heterogeneous disease affected by some modifiable risk factors, such as lack of exercise, hypertension, diabetes, and obesity.5 Recently, sleep disorders, including OSA, have also been recognized as an independent contributor promoted the aggressiveness of AD.6 In animal models, OSA can cause neuroinflammation and oxidative stress damage in the brain, including in key areas involved in AD pathophysiology such as hippocampus.7 Neuroimaging studies have shown that OSA patients had structural and functional changes in brain regions related to cognitive function, including frontal lobe, hippocampus, and parietal lobe.8 In a study of OSA, compared with the serum levels of AD biomarkers (Aβ40, t-tau, p-tau) in controls, the serum levels in OSA patients were significantly increased.9 Moreover, the levels of these markers in the serum were significantly negatively correlated with cognitive function.10,11 It was currently believed that OSA with deficits in cognitive performance was related to neuroinflammation and oxidative stress triggered by chronic intermittent hypoxia, and the pathological changes of brain tissue in hypoxia-sensitive areas may be the basis of cognitive dysfunction.12 Although studies have shown that there is a certain correlation between AD and OSA, the relationship between the two remains incompletely understood, especially at the genetic levels. Therefore, it is essential to analyze these two diseases using bioinformatics methods at the genetic and molecular levels.
Traditional biological research focuses on exploring the pathogenesis of diseases at the molecular level, and cannot grub the effective biological significance contained in massive data. The field of the research of life science has rapidly grown and evolved by bioinformatics analysis through advancements of sequencing technology over the past decades. WGCNA is a novel bioinformatics method, which can obtain co-expression modules with high biological significance by specific screening of genes related to traits, and is widely used in the study of comorbidities.13,14 In the present study, based on a comprehensive bioinformatics analysis, we firstly identified DEGs between AD and OSA samples, and further clarify their potential molecular mechanisms. In further studies, these candidate genes and pathways could be investigated more closely to identify new and clear gene targets and potentially provide guidance for subsequent clinical studies.
Materials and Methods
The workflow of the analysis and hub gene extraction curation pipeline was shown in Figure 1. We elaborated on each step in the following sub-sections.
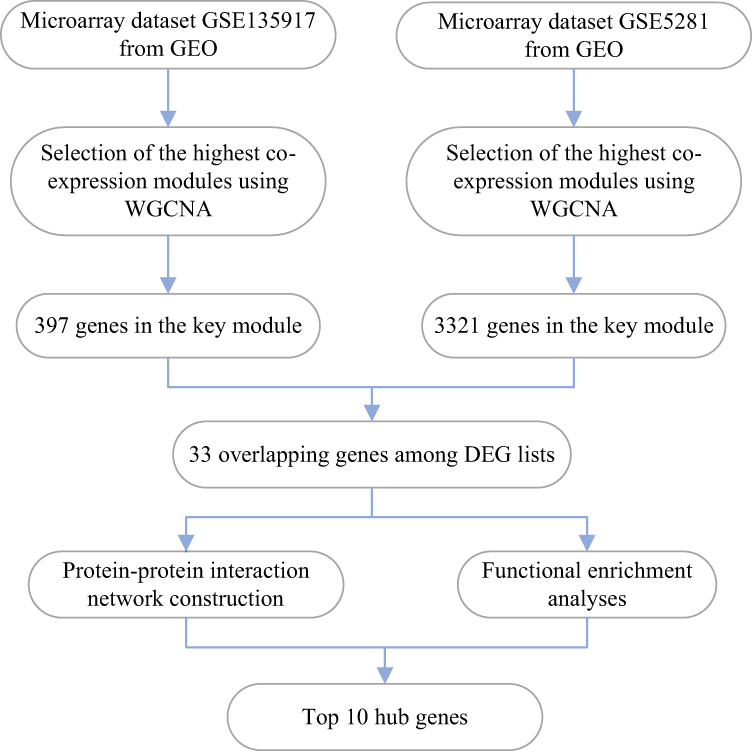 |
Figure 1 The workflow of the analysis hub gene extraction curation pipeline. Abbreviations: GEO, Gene Expression Omnibus; WGCNA, weighted gene co-expression network analysis; DEG, differentially expressed genes. |
Dataset Preparation
All gene expression data of AD and OSA were downloaded from Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/). The GSE5281 dataset was based on GPL570 platforms (HG-U133_Plus_2); Affymetrix Human Genome U133 Plus 2.0 Array), including 87 brain samples from AD patients and 74 normal aged brain samples from healthy controls. Another dataset GSE135917 including 24 subcutaneous fat samples from OSA patients and 8 subcutaneous fat samples from healthy controls, which were studied with GPL6244 platforms (HuGene-1_0-st; Affymetrix Human Gene 1.0 ST Array). R software (version 4.0.2, https://www.r-project.org/) and Bioconductor Packages were used to process the raw expression data, make expression matrix and match the probe to gene symbol. Probes were converted to the gene symbols based on a manufacturer-provided annotation file and duplicated probes sets were deleted by determining the median expression value of all its corresponding probes. Probes without corresponding gene symbols were also removed. The affy package under the R environment was used to preprocess and normalize the microarray datasets with raw data (CEL files). The parameters, RMA (for background correction) and impute (for supplemental missing value) were performed in the R software. The gene variabilities using Median Absolute Deviation (MAD) were measured and the top 5000 genes were filtered for network construction.
WGCNA Analysis
Unsigned co-expression networks were constructed using the WGCNA algorithms in R. A one-step network construction method was utilized to identify co-expression modules with the blockwise-modules function in the WGCNA package.14 Before co-expression network construction, the flash-Clust tool in the R language was used to construct sample clustering tree maps with the appropriate threshold value to detect and eliminate the outliers in the GSE5281 and GSE135917 datasets. And then, the pick soft threshold function of WGCNA was used to calculate β from 1 to 30 to select the best soft threshold. Based on the selected soft threshold, the adjacency matrix was converted to topological overlap matrix for constructing the network, and the gene dendrogram and module color were established by utilizing the degree of dissimilarity. A dynamic tree-cutting algorithm using dissimilarity matrix was applied to detect gene modules, gene sets with high topological overlap. To obtain moderately sized modules, the minimum number of genes was set at 30 and a cutline was chosen to merge modules with similar expression patterns. To identify the relationships between modules and clinic traits, we calculated the correlation between Module Eigengenes (MEs) and clinical trait and searched for the most significant associations. The MEs were calculated by the first principal component, which was considered as a representative of the expression patterns of module genes. For each module, we defined the Module Membership (MM) as correlation of gene expression profile with ME and the Gene Significance (GS) as the absolute value of the correlation between gene and clinical traits. In this study, genes with high MM in a module were assigned to the module and the module with high GS and P value <0.05 was considered to be highly related to clinical traits. In order to verify the stability of WGCNA, we used the Module Preservation function to calculate the preservation of the module. In addition, module preservation analysis was performed to calculate the Z-summary score. A Z-summary score >10 means strong evidence of module preservation, whereas a Z-summary score <2 means or 2< a Z-summary score <10 mean no evidence or weak evidence of module preservation.
KEGG and GO Enrichment Analysis
To explore Gene Ontology (GO) of DEGs, R package cluster Profiler package was used to explore the functions among genes of interest, with a cut-off criterion of adjusted p<0.05. GO annotation that contains the three sub-ontologies—biological process (BP), cellular component (CC), and molecular function (MF)—can identify the biological properties of genes and gene sets for all organisms. Kyoto Encyclopedia of Genes and Genomes (KEGG, https://www.genome.jp/kegg/) was a database for understanding high-level functions and utilities of the biological system. The GO and KEGG pathway enrichment of common genes in OSA and AD were analyzed by Cluster Profiler and DOSE package in R.15 Adjust P-value <0.05 was considered statistically significant. The results were visualized with R and Enrichr.
Protein–Protein Interaction Network Construction and Module Analysis
Genes, which owned significant correlations with clinical traits and high network connectivity, were informally referred to as intramodular hub genes of the modules. In this study, we chose the common genes in the AD and OSA related modules for further research. The STRING (Search Tool for the Retrieval of Interacting Genes) online tool, which was designed for predicting protein–protein interactions (PPI), was used to construct a PPI network of selected genes.16 And, an interaction with a score of combination > 0.15 was considered statistically significant. Next, we downloaded the interaction information and optimized the PPI network with Cytoscape software (v3.8.0) for better visualization. Minimal Common Oncology Data Elements (MCODE) was used to identify significant gene clusters and obtain cluster scores (filter criteria: degree cut-of=2; node score cut-of=0.2; k-core=2; max depth=100).
Hub Gene Selection
The plug-in CytoHubba (version 1.6) in Cytoscape was used to explore significant genes in the PPI network, and the genes with most interactions were referred as hub genes, which might play core roles in the disease pathogenesis.17 We used Maximal Clique Centrality (MCC) to calculate the hub genes.
Results
Identification of DEGs
According to the sample clustering results, none outlier sample in GSE135917 and GSE5281 was detected and eliminated, and then the sample dendrogram and trait heatmaps were built, respectively (Figures 2A and 3A). Merging similar modules, 7 modules and 4 modules were identified from the co-expression network of GSE5281 and GSE135917, respectively. After respectively calculating the correlations between modules and AD or OSA, we plotted the corresponding heatmaps of module-trait relationships (Figures 2B and 3B). Dendrogram of all differentially expressed genes clustered based on the measurement of dissimilarity. The color band shows the results obtained from the automatic single-block analysis (Figures 2C and 3C). In addition, the yellow module related to AD and the turquoise module related to OSA were respectively identified as the most relevant key modules (Figures 2D and 3D). By using the pick soft threshold function of WGCNA, we found the optimal soft threshold power was 1 for GSE5281 samples (Figure 2E) and 7 for GSE135917 samples (Figure 3E).
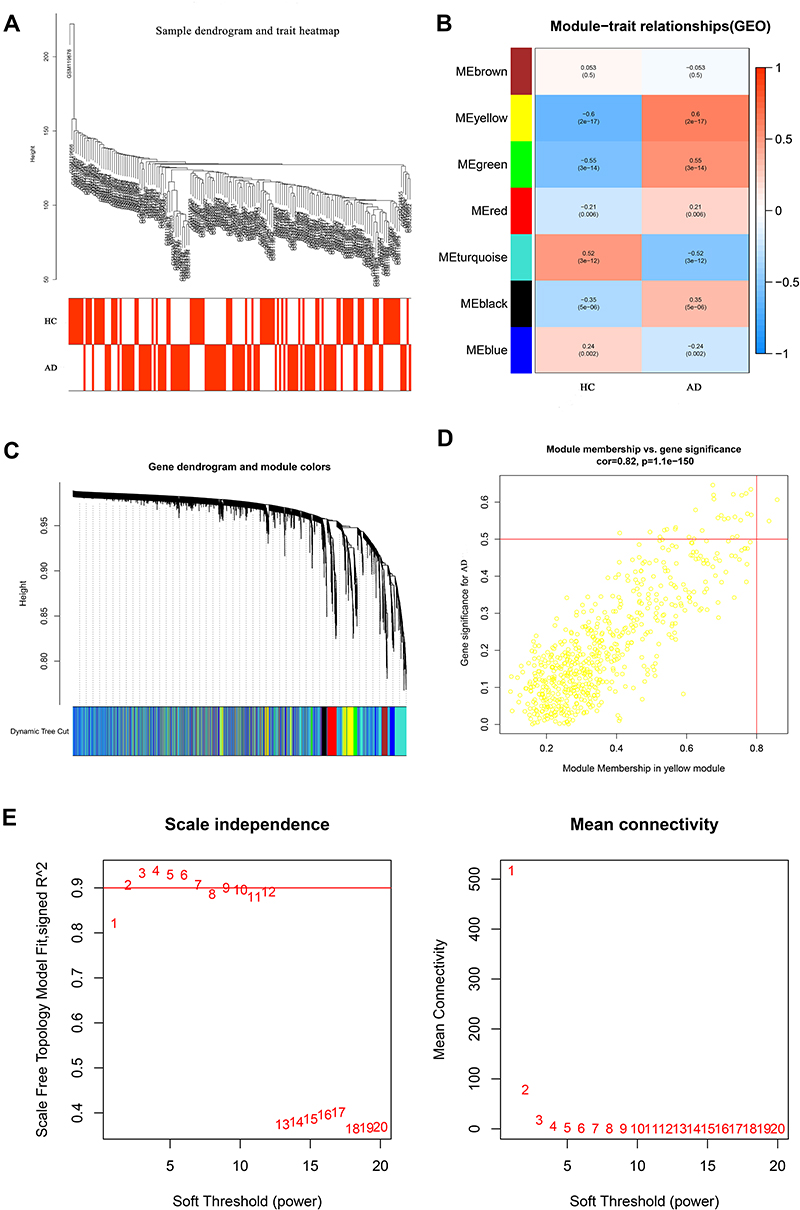 |
Figure 2 WGCNA of AD dataset (GSE5281). (A) The sample dendrogram and trait heatmaps. (B) Heatmap showed the relationships between different modules and clinical traits, each row presented a module eigengene, column to a trait. Each cell represented the correlation between module eigengenes (rows) and traits (columns), and the corresponding P value.7 modules were related to AD. (C) Dendrogram of all differentially expressed genes of AD was clustered based on the measurement of dissimilarity. The color band showed the results obtained from the automatic single-block analysis. (D) Module membership in yellow module. (E) Analysis of network topology for various soft‐thresholding powers, a soft threshold of 1 was the most suitable value to AD. The abscissa of the picture represented soft‐thresholding power. Abbreviations: WGCNA, weighted gene co-expression network analysis; HC, healthy subjects; AD, Alzheimer’s disease. |
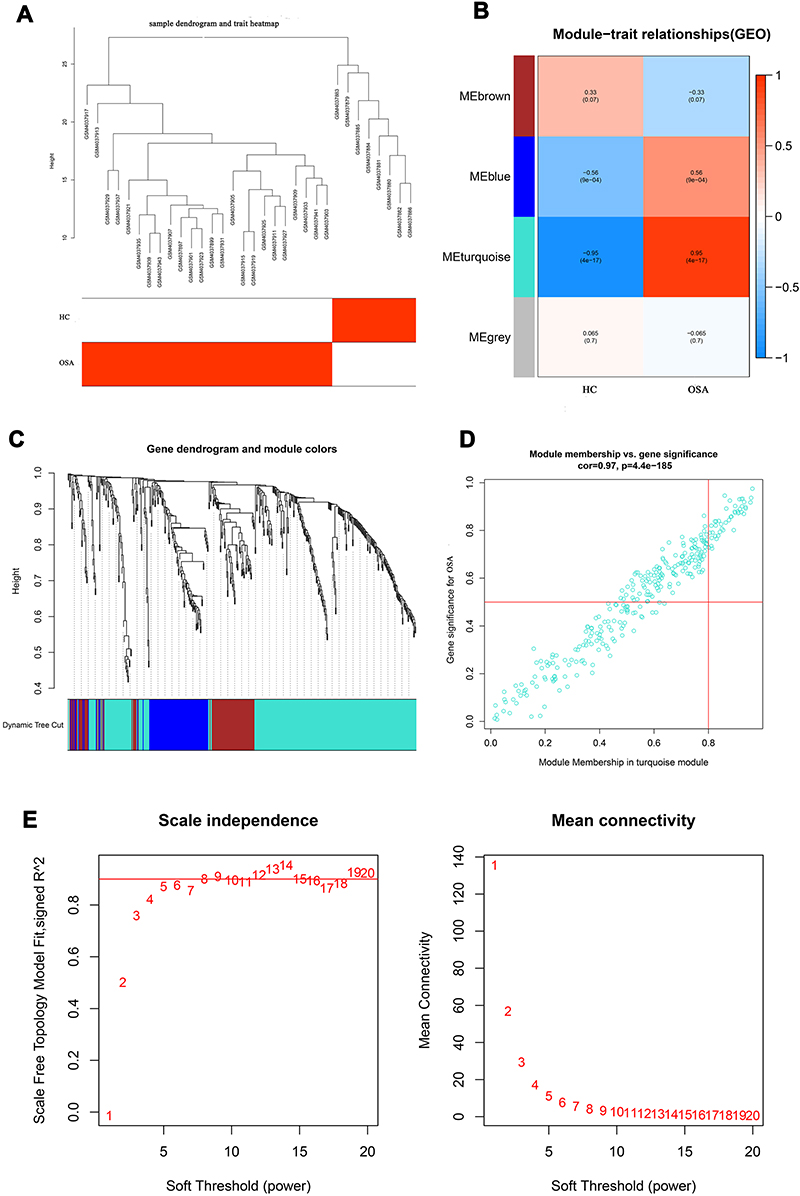 |
Figure 3 WGCNA of OSA dataset (GSE135917). (A) The sample dendrogram and trait heatmaps. (B) Heatmap shows the relationships between different modules and clinical traits, each row presents a module eigengene, column to a trait. Each cell represents the correlation between module eigengenes (rows) and traits (columns), and the corresponding P value. 4 modules are related to OSA. (C) Dendrogram of all differentially expressed genes of OSA was clustered based on the measurement of dissimilarity. The color band shows the results obtained from the automatic single-block analysis. (D) Module membership in yellow module. (E) Analysis of network topology for various soft‐thresholding powers. The abscissa of the picture represents soft‐thresholding power. A soft threshold of 7 is the most suitable value to OSA. Abbreviations: WGCNA, weighted gene co-expression network analysis; WGCNA, weighted gene co-expression network analysis; HC, healthy subjects; OSA, obstructive sleep apnea. |
Additionally, we performed the module preservation to calculate the Z-summary scores of each module. Then key modules were screened with the following criteria: |cor| >0.5, p <0.05 and Z-summary score >10. Morever, the turquoise, yellow and green module associated with AD were identified as key modules (Figure 4A), and turquoise and blue module associated with OSA were identified as key modules (Figure 4B). A total of 33 common genes were obtained by overlapping intersecting DEGs in key modules of OSA and AD (Figure 4C).
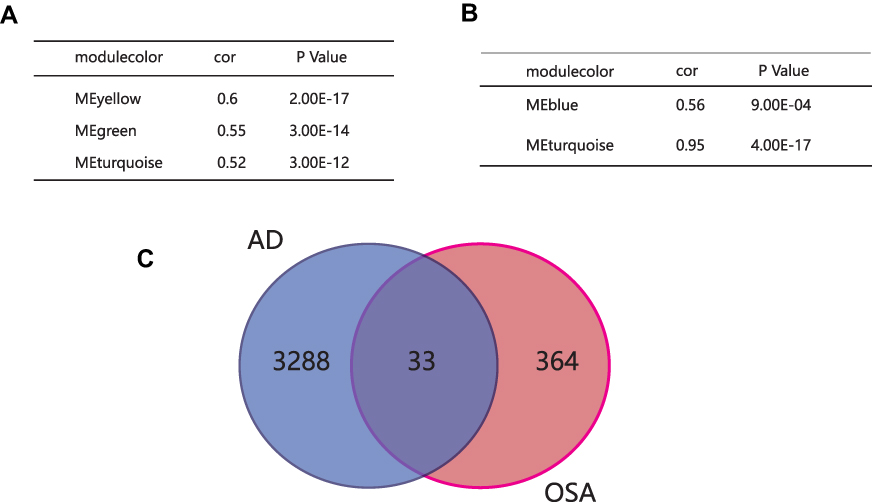 |
Figure 4 Screening for key modules and common genes. (A) Green, turquoise and yellow modules were screened in AD. (B) Blue and turquoise modules were screened in OSA. (C) 33 common genes in key modules of OSA and AD were screened. Abbreviations: OSA, obstructive sleep apnea; AD, Alzheimer’s disease; Cor, correlation coefficient. |
KEGG and GO Enrichment Analysis for Intersecting DEGs
The GO and KEGG analyses were applied to the cluster Profiler package for exploring the potential molecular functions and molecular mechanisms of DEGs. A bubble plot and bar chart of GO and KEGG were drawn (Figure 5A and B). A total of 10 BP terms were enriched, including response to oxidative stress, response to corticosteroid, leukocyte migration, positive regulation of myelination, myeloid leukocyte migration, response to steroid hormone, leukocyte chemotaxis, response to cAMP, neutrophil chemotaxis, neutrophil migration. The 10 CC terms were significantly enriched, including endoplasmic reticulum lumen, ficolin−1−rich granule membrane, tertiary granule membrane, proton−transporting V−type ATPase, V0 domain, fibrillar collagen trimer, banded collagen fibril, external side of plasma membrane, vacuolar proton−transporting V−type ATPase complex, tertiary granule, complex of collagen trimers. A total of 10 MF terms were enriched, including growth factor binding, cytokine activity, receptor ligand activity, ferroxidase activity, oxidoreductase activity, oxidizing metal ions, oxygen as acceptor, BH domain binding, death domain binding, IgG binding, IgG binding, RAGE receptor binding. The top eight KEGG pathways were most enriched, including PI3K−Akt signaling pathway, JAK−STAT signaling pathway, Focal adhesion, ECM−receptor interaction, Platelet activation, Relaxin signaling pathway, Apelin signaling pathway, Alcoholic liver disease.
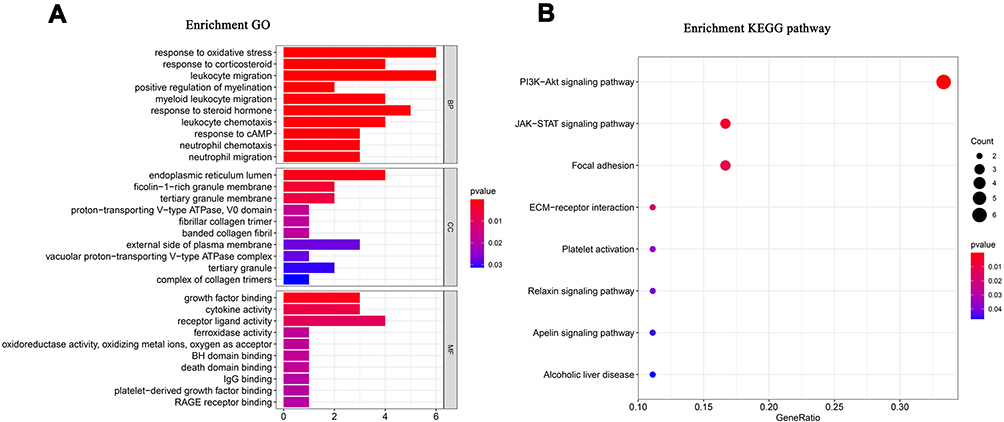 |
Figure 5 Functional enrichment and Protein–protein interaction network. (A) GO pathways enrichment analysis of AD and OSA shared genes. The length of the bar represents the number of genes, and the color change of the bar corresponds to different P values. The first 10 significantly enriched GO annotations of cellular component, biological process, molecular function. (B) Bubble plots of the first 8 significantly enriched KEGG pathways. Abbreviations: OSA, Obstructive sleep apnea; AD, Alzheimer’s Disease; GO, Gene Ontology (GO); KEGG, Kyoto Encyclopedia of Genes and Genomes. |
PPI Network Construction
The interaction network between proteins coded by DEGs, which was comprised of 32 nodes and 83 edges, was constructed by STRING and visualized by Cytoscape (Figure 6A). The MCODE plugin was used to identify gene cluster modules. We identified two modules in this network, according to the filter criteria. Cluster 1 (Figure 6C) had the highest cluster score (score: 4.80, 6 nodes and 12 edges), followed by cluster 2 (Figure 6D) (score: 4.20, 11 nodes and 21 edges).
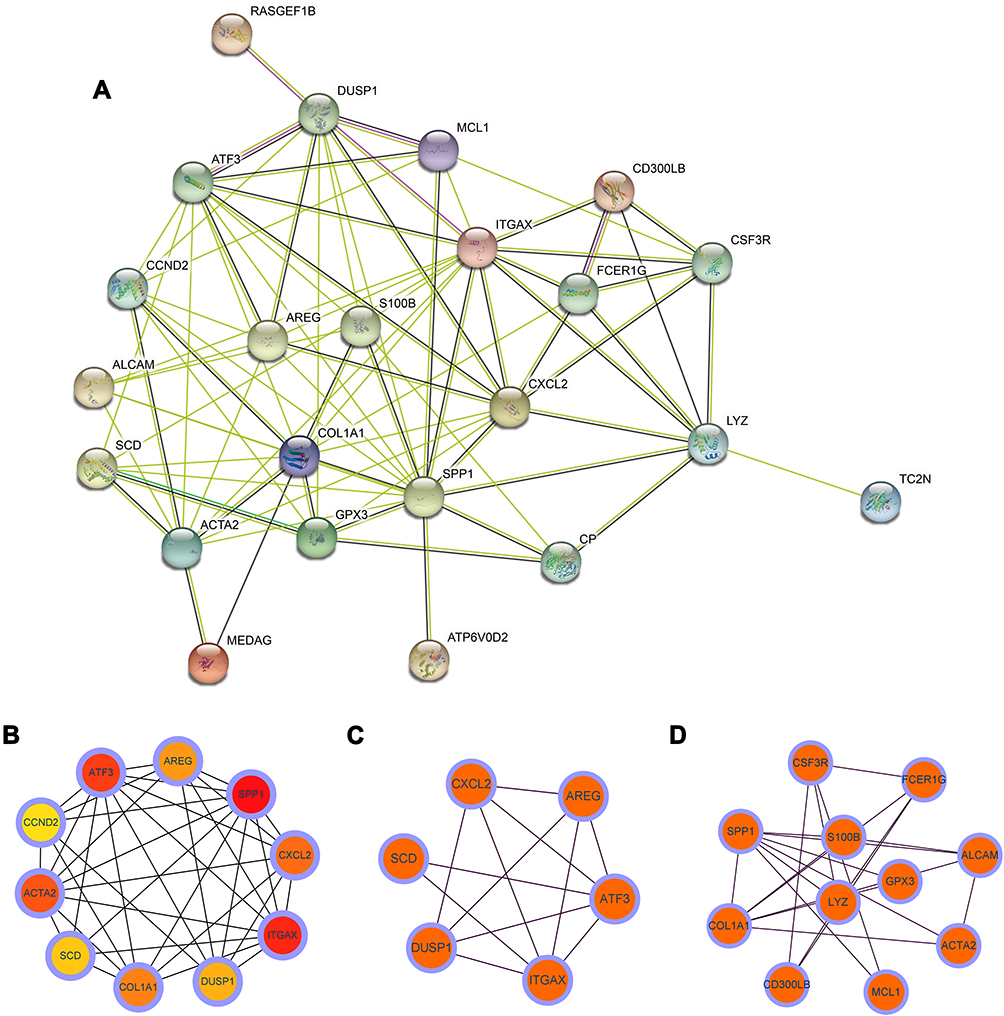 |
Figure 6 (A) Based on STRING database, protein–protein interaction networks of the DEGs in the AD and OSA were constructed. Each node represents a protein, while each edge represents one protein–protein association. (B) The hub genes were screened from the PPI network using the MCC algorithm of CytoHubba plugin. Two cluster modules were extracted by MCODE. (C) Cluster 1 had the highest cluster score (score: 4.80, 6 nodes and 12 edges), (D) followed by cluster 2 (score: 4.20, 11 nodes and 21 edges). Abbreviations: OSA, obstructive sleep apnea; AD, Alzheimer’s disease. |
Hub Gene Analysis
According to the MCC sores in the CytoHubba, the top ten highest-scored genes were selected as the hub genes (Figure 6B), including Amphiregulin (AREG), Secreted Phosphoprotein 1 (SPP1), C-X-C Motif Chemokine Ligand 2 (CXCL2), Integrin Subunit Alpha X (ITGAX), Dual Specificity Phosphatase 1 (DUSP1), Collagen Type I Alpha 1 Chain (COL1A1), Stearoyl-CoA Desaturase (SCD), Actin Alpha 2, Smooth Muscle (ACTA2), Cyclin D2 (CCND2), Activating Transcription Factor 3 (ATF3). The gene list with gene names, abbreviations, and functions calculated by CytoHubba was shown in Table 1.
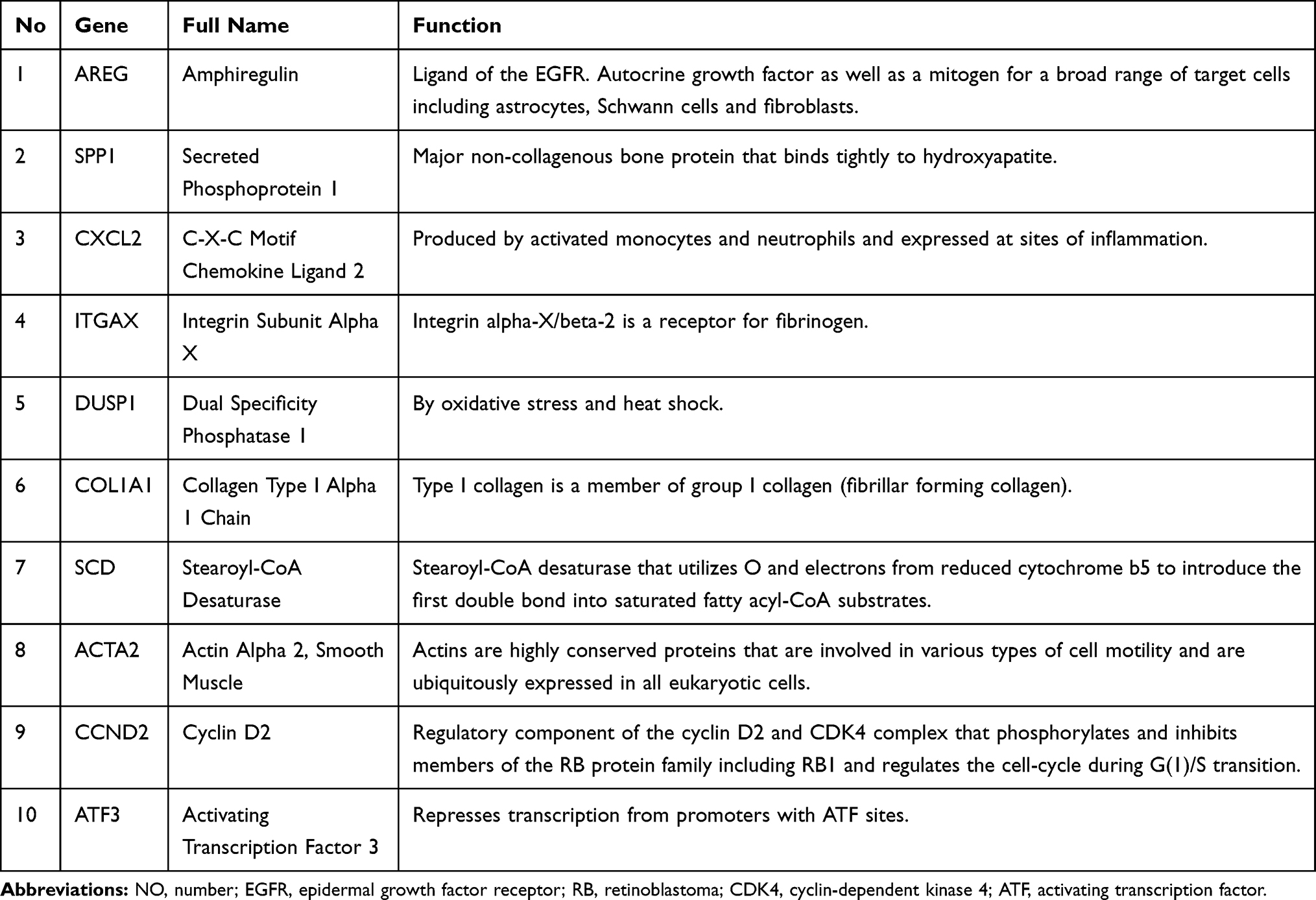 |
Table 1 Top 10 Hub Genes Ranked with Degrees |
Discussion
Alzheimer’s disease is a chronic, progressive, neurodegenerative disease. Although people are more and more aware of AD, its pathogenesis is still complicated. As previously described, Obstructive sleep apnea is a treatable sleep disorder, common in the general population, associated with excessive daytime sleepiness and neurocognitive deficits.8 Much evidence has shown that OSA can promote the occurrence and development of AD.18,19 Simultaneously, several studies also revealed that treatment of OSA in elderly patients could prevent or delay the onset of AD, mitigated the development and even reversed neuro-degenerative changes of particular brain areas in AD patients.20,21 In addition, AD patients often have compliance difficulty with the use of CPAP resulting in a bidirectional, positive feedback loop leading to worse outcomes.22 Therefore, it is imperative to find the common genes and explore the molecular mechanisms of OSA affecting AD as a means of improving the early diagnosis and treatment interventions of AD.
We analyzed AD and OSA datasets using WGCNA for the first time to identify DEGs. By bioinformatic analysis, a total of 10 hub genes were identified as shared genes in AD and OSA patients. Among all of the hub genes, Dual-specific phosphatase 1 (DUSP1), stearoyl-CoA desaturase (SCD), Secreted Phosphoprotein 1 (SPP1) and C-X-C motif chemokine ligand 2 (CXCL2) were showed a closer association with OSA affecting AD. Additionally, the functional enrichment analysis of shared genes found that AD and OSA-related genes are roughly the same in molecular function, biological process, and cell composition, indicating that the two diseases have the same mechanism in the development process. Thirdly, with significant pathway analysis, we found that AD and OSA shared some common pathways: response to oxidative stress, response to corticosteroid, positive regulation of myelination, myeloid leukocyte migration, response to steroid hormone, leukocyte chemotaxis, neutrophil chemotaxis, neutrophil migration. Consistent with a previous study, the results of the present study indicated that the Oxidative stress and neuroinflammation were the common pathways of AD and OSA.
Chronic intermittent hypoxemia increases oxidative stress by enhancing the production of relative oxygen species in the body of OSA patients, and the imbalance of oxidation/antioxidation.23 The long-term oxidative stress reaction of the blood vessel wall can cause vascular endothelial damage, especially the tiny blood vessels in the brain, leading to brain atrophy and cognitive impairment.24 Previous studies performed on the elderly found that arteriosclerosis was related to β-amyloid deposition, and it was believed that arteriosclerosis was related to the occurrence and development of AD.25 DUSP1 and SCD are thought to be involved in oxidative stress. In our research, we found that these two genes were the shared genes of OSA and AD, and we thought that the mechanism of OSA affecting AD may be related to the process of oxidative stress. DUSP1 belongs to the mitogen-activated protein kinase phosphatase family, which is closely involved in the regulation of intracellular signal transduction and is induced by growth factors, oxidative stress and continuous hypoxia. Studies have shown that the expression of DUSP1 protein in monocytes and granulocytes isolated from OSA subjects was increased significantly. DUSP1 gene expression is suppressed after continuous positive airway pressure treatment of AD.23 Studies reported that the expression of DUSP1 in the hippocampal neurons of AD patients was also increased, and overexpression of DUSP1 will decrease tau phosphorylation, and knocking down the expression of DUSP1 will increase the level of tau phosphorylation.26 Some studies believe that it can selectively regulate the production of Aβ42 in neuronal cells under hypoxic stress and can be used as a therapeutic target to prevent the progression of AD.27 SCD is a central regulator of fuel metabolism. A study focusing on the expression of SCD in brain neuron cells of AD patients found that compared with the control group, the expression of SCD in AD patients was elevated significantly, and the overexpression of SCD caused an increase in the secretion of β42 amyloid.28 At the same time, studies showed that chronic intermittent hypoxia of OSA could activate liver hypoxia-inducible factor-1, which increased the SCD expression, and the expression level of SCD was positively correlated with the severity of local hypoxia.29,30 Some previous studies have focused on the expression of DUSP1 and SCD in OSA or AD patients, respectively. We discovered for the first time that DUSP1 and SCD were shared genes in OSA and AD patients. The regulation of these two genes on oxidative stress in the body may be related to the mechanism of OSA affecting AD.
Chronic neuroinflammation has been proposed as a driving force for AD occurrence and development, which is characterized by amyloid-β deposition, neurofibrillary tangles, neuronal loss, and activation of glial cells.31 Similarly, studies have noted that both OSA patients and animal models will have peripheral and neuroinflammation. Several studies focusing on OSA and inflammation showed that the expression of multiple serum inflammatory factors in OSA patients were elevated, such as CRP, TNF-α, IL-6, IL-8, ICAM, VCAM, and the expression level was closely related to the apnea hypopnea index of OSA patients.32,33 Previous studies have found that both SPP1 and CXCL2 were related to neuroinflammation. In our research, we found that these two genes may be related to the pathogenesis of OSA affecting AD. SPP1 is a cytokine expressed by cytotoxic T cells and a marker of neuroinflammation. Increased expression of SPP1 genes has been observed in AD brains and in a transgenic mouse model of AD.34 In the context of AD, studies showed SPP1 could promote the monocyte-macrophage’ recruitment into AD brains, and their polarization towards an anti-inflammatory, highly phagocytic phenotype to facilitate Abeta clearance.26,35 CXCL2 is a pro-inflammatory factor, produced by activated monocytes and neutrophils and expressed at sites of inflammation. In a study of AD, the level of serum CXCL2 expression in B3 capillaries was significantly higher compared to healthy controls.36 Recently, study have pointed out that the serum chemokine levels of OSA patients were significantly increased, which was related to the degree of systemic inflammation.37 Together, we found that the molecular mechanism of OSA affecting AD may be related to the regulation of neuroinflammation by SPP1 and CXCL2.
This study may help clarify the pathogenesis of OSA affecting AD. Despite our systematic findings, this study is not free from limitations. First, the present study was a retrospective analysis based on existing databases and public domain information. Second, though genes shared by OSA and AD were identified, the biological function of these genes in AD has not yet been fully understood. Thus, the work to reveal the pathogenesis of these two diseases at the molecular level remains indispensable.
Conclusion
The present study was the first to our knowledge to investigate the shared genes of AD and OSA using bioinformatics methods. These key genes might help to provide a novel perspective to explore the underlying mechanism about OSA affecting AD.
Ethical Statement
The authors are accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. This study was conducted in accordance with the Declaration of Helsinki (as revised in 2013). The study was approved by the Second Affiliated Hospital of Nanchang University research committee.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant numbers: 31860293), and Natural Science Foundation of Jiangxi Province (Grant numbers: 20192BAB205047, 20151BAB205018).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Prince M, Ali GC, Guerchet M. Recent global trends in the prevalence and incidence of dementia, and survival with dementia. Alzheimers Res Ther. 2016;1(8):30.
2. JOAQUIN DURÁN SERR. Obstructive sleep apnea-hypopnea and related clinical features in a population-based sample of subjects aged 30 to 70 yr. Am J Respir Crit Care Med. 2001;3(16):685–689.
3. Senaratna CV, Perret JL, Lodge CJ. Prevalence of obstructive sleep apnea in the general population: a systematic review. Sleep Med Rev. 2017;34(34):70–81. doi:10.1016/j.smrv.2016.07.002
4. Polsek D, Gildeh N, Cash D, et al. Obstructive sleep apnoea and Alzheimer’s disease: in search of shared pathomechanisms. Neurosci Biobehav Rev. 2018;86:142–149. doi:10.1016/j.neubiorev.2017.12.004
5. Gill Livingston JHAS, Jiska Cohen-Mansfield CCSG, Eric B, et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet. 2020;2(396):413–446.
6. Dunietz GL, Chervin RD, Burke JF, et al. Obstructive sleep apnea treatment and dementia risk in older adults. Sleep. 2021;44(9):076. doi:10.1093/sleep/zsab076
7. Gozal D, Daniel JM, Dohanich GP. Behavioral and anatomical correlates of chronic episodic hypoxia during sleep in the rat. J NEUROSCI. 2001;21(7):2442–2450. doi:10.1523/JNEUROSCI.21-07-02442.2001
8. Kaminska M, Lafontaine AL, Kimoff RJ. The Interaction between obstructive sleep apnea and parkinson’s disease: possible mechanisms and implications for cognitive function. Parkinson's Dis. 2015;2015:849472. doi:10.1155/2015/849472
9. Kong W, Zheng Y, Xu W, et al. Biomarkers of Alzheimer’s disease in severe obstructive sleep apnea-hypopnea syndrome in the Chinese population. Eur Arch Otorhinolaryngol. 2021;278(3):865–872. doi:10.1007/s00405-020-05948-2
10. Wang JXZQ. Effects of somatotropic axis on cognitive dysfunction of obstructive sleep apnea. Sleep Breath. 2020;1(24):175–182.
11. Kong W, Zheng Y, Xu W, et al. Biomarkers of Alzheimer’s disease in severe obstructive sleep apnea–hypopnea syndrome in the Chinese population. Eur Arch Otorhinolaryngol. 2021;8(273):865–872.
12. Lal C, Hardiman G, Kumbhare S, et al. Proteomic biomarkers of cognitive impairment in obstructive sleep apnea syndrome. Sleep Breath. 2019;23(1):251–257. doi:10.1007/s11325-018-1693-8
13. Ian M, Copple WDH, Callegaro G, et al. Characterisation of the NRF2 transcriptional network and its response to chemical insult in primary human hepatocytes: implications for prediction of drug-induced liver injury. Arch Toxicol. 2019;2(93):385–399.
14. Zhai X, Xue Q, Liu Q. Colon cancer recurrence-associated genes revealed by WGCNA co-expression network analysis. Mol Med Rep. 2017;5(16):6499–6505.
15. Guangchuang YLWYH. clusterProfiler: an R Package for Comparing Biological Themes Among Gene Clusters. OMICS. 2012;5(16):284–287.
16. Damian Szklarczyk AL, Msnt JH-C, Peer Bork LJJA. STRING v11: protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019;47(47):D607–13. doi:10.1093/nar/gky1131
17. Chin C, Shu-Hwa C, Hsin-Hung W, et al. cytoHubba: identifying hub objects and subnetworks from complex interactome. BMC Syst Biol. 2014;4(8):S11. doi:10.1186/1752-0509-8-S4-S11
18. Xu X. Caregiver-Reported Sleep Disturbances Are Associated with Behavioral and Psychological Symptoms in an Asian Elderly Cohort with Cognitive Impairment-No Dementia. J Geriatr Psychiatry Neurol. 2018;2(31):70–75.
19. Montgomery W, Amir G, Kristin K, et al. Alzheimer’s disease severity and its association with patient and caregiver quality of life in Japan: results of a community-based survey. BMC Geriatr. 2018;1(18):141. doi:10.1186/s12877-018-0831-2
20. Anne-Cécile Troussière CMCJ, Florence Richard XDPD, Florence Pasquier SB. Treatment of sleep apnoea syndrome decreases cognitive decline in patients with Alzheimer’s disease. J Neurol Neurosurg Psychiatry. 2014;12(85):1405–1408. doi:10.1136/jnnp-2013-307544
21. Kitamura T, Miyazaki S, Sulaiman HB, et al. Insomnia and obstructive sleep apnea as potential triggers of dementia: is personalized prediction and prevention of the pathological cascade applicable? EPMA J. 2020;11(3):355–365. doi:10.1007/s13167-020-00219-w
22. Elias A, Cummins T, Tyrrell R, et al. Risk of Alzheimer’s Disease in Obstructive Sleep Apnea Syndrome: amyloid-β and Tau Imaging. J Alzheimers Dis. 2018;66(2):733–741. doi:10.3233/JAD-180640
23. Hoffmann MS, Singh P, Wolk R, et al. Obstructive sleep apnea and intermittent hypoxia increase expression of dual specificity phosphatase 1. Atherosclerosis. 2013;231(2):378–383. doi:10.1016/j.atherosclerosis.2013.09.033
24. Li X, Lyu P, Ren Y, et al. Arterial stiffness and cognitive impairment. J Neurol Sci. 2017;380:1–10. doi:10.1016/j.jns.2017.06.018
25. Hughes TM, Craft S, Lopez OL. Review of ‘the potential role of arterial stiffness in the pathogenesis of Alzheimer’s disease’. Neurodegener Dis Manag. 2015;5(2):121–135. doi:10.2217/nmt.14.53
26. Wirths O, Breyhan H, Marcello A, et al. Inflammatory changes are tightly associated with neurodegeneration in the brain and spinal cord of the APP/PS1KI mouse model of Alzheimer’s disease. Neurobiol Aging. 2010;31(5):747–757. doi:10.1016/j.neurobiolaging.2008.06.011
27. Jung S, Nah J, Han J, et al. Dual-specificity phosphatase 26 (DUSP26) stimulates Aβ42 generation by promoting amyloid precursor protein axonal transport during hypoxia. J Neurochem. 2016;137(5):770–781. doi:10.1111/jnc.13597
28. Astarita G, Jung KM, Vasilevko V, et al. Elevated stearoyl-CoA desaturase in brains of patients with Alzheimer’s disease. PLoS One. 2011;6(10):e24777. doi:10.1371/journal.pone.0024777
29. Li J, Thorne LN, Punjabi NM, et al. Intermittent hypoxia induces hyperlipidemia in lean mice. Circ Res. 2005;97(7):698–706. doi:10.1161/01.RES.0000183879.60089.a9
30. Li J, Bosch-Marce M, Nanayakkara A, et al. Altered metabolic responses to intermittent hypoxia in mice with partial deficiency of hypoxia-inducible factor-1alpha. Physiol Genomics. 2006;25:450–457. doi:10.1152/physiolgenomics.00293.2005
31. Flores B, von Bernhardi R. Transforming growth factor β1 modulates amyloid β-induced glial activation through the Smad3-dependent induction of MAPK phosphatase-1. J Alzheimers Dis. 2012;32(2):417–429. doi:10.3233/JAD-2012-120721
32. Rashid Nadeem MD, Janos Molnar MD, Essam M, et al. Serum Infl ammatory Markers in Obstructive Sleep Apnea: a Meta-Analysis. J Clin Sleep Med. 2013;09(9):1003–1012. doi:10.5664/jcsm.3070
33. Arnaud C, Bochaton T, Pépin JL, et al. Obstructive sleep apnoea and cardiovascular consequences: pathophysiological mechanisms. Arch Cardiovasc Dis. 2020;113(5):350–358. doi:10.1016/j.acvd.2020.01.003
34. Sathe G, Albert M, Darrow J, et al. Quantitative proteomic analysis of the frontal cortex in Alzheimer’s disease. J Neurochem. 2021;156(6):988–1002. doi:10.1111/jnc.15116
35. Rentsendorj A, Sheyn J, Fuchs DT, et al. A novel role for osteopontin in macrophage-mediated amyloid-β clearance in Alzheimer’s models. Brain Behav Immun. 2018;67:163–180. doi:10.1016/j.bbi.2017.08.019
36. Annie G, Bryant MH. Cerebrovascular Senescence Is Associated with Tau Pathology in Alzheimer’s Disease. Front Neurol. 2020;2020(11):575953.
37. Fu C, Lu H, Wu X, et al. Chronic intermittent hypoxia decreases pulmonary clearance of (99m) Tc-labelled particulate matter in mice. Am J Transl Res. 2017;9(6):3060–3072.
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.