")
Back to Journals » International Journal of General Medicine » Volume 14
Identification of ABCC5 Among ATP-Binding Cassette Transporter Family as a New Biomarker for Hepatocellular Carcinoma Based on Bioinformatics Analysis
Authors Qiu Y, Li H , Xie J , Qiao X , Wu J
Received 12 August 2021
Accepted for publication 29 September 2021
Published 27 October 2021 Volume 2021:14 Pages 7235—7246
DOI https://doi.org/10.2147/IJGM.S333904
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Yuting Qiu,1,* Haobo Li,2,* Jiaheng Xie,3 Xinwei Qiao,1 Jing Wu1
1Department of Gastroenterology, Beijing Friendship Hospital, Capital Medical University, National Clinical Research Centre for Digestive Diseases, Beijing, 100050, People’s Republic of China; 2Department of Pulmonary and Critical Care Medicine, China-Japan Friendship Hospital; Graduate School of Peking Union Medical College, Chinese Academy of Medical Sciences, Beijing, 100029, People’s Republic of China; 3Department of Burn and Plastic Surgery, The First Affiliated Hospital of Nanjing Medical University, Nanjing, Jiangsu, 210029, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xinwei Qiao; Jing Wu
Department of Gastroenterology, Beijing Friendship Hospital, Capital Medical University, National Clinical Research Centre for Digestive Diseases, No. 95, Yongan Road, Xicheng District, Beijing, 100050, People’s Republic of China
Tel +86-15010573490
; +86-13701134610
Fax +86-10-63926370
Email [email protected]; [email protected]
Purpose: Liver cancer is the fifth most common type of cancer worldwide, and the ATP-binding cassette (ABC) transporter family has been widely accepted as a cause of multidrug resistance. This study was conducted to explore the potential value and mechanisms of the ABC transporter gene family in the liver hepatocellular carcinoma (LIHC).
Materials and Methods: Data were collected from different public databases. UALCAN, ONCOMINE, and GEPIA were used to retrieve a selection of differently expressed and pathological stage-related genes among the ABC family. Principal component analysis (PCA) was utilized for grouping, and its prognostic value was evaluated by univariate and multivariate Cox analyses. The co-expression pattern was constructed with UALCAN, and the functional analyses were carried out with DAVID. The correlation between the biomarker and immune infiltration, genetic alteration frequency, and drug sensitivity were explored with TIMER, cBioPortal, GDSC and CTRP, respectively. Finally, tSNE algorithm was used to explore the distribution of ABCC5 expressed cells.
Results: Among the ABC transporter family members, ABCC5 was differently expressed and strongly related to the pathological stage of LIHC. PCA divided patients of LIHC into two groups, and Cox analyses demonstrated that ABCC5 was an independent risk factor of LIHC. Functional analyses indicated that the genes were enriched in the pathways of transmembrane transporter, ATPase activity, and bile secretion. ABCC5 is also associated with immune infiltration of cells like macrophages, neutrophils, and dendritic cells. The genetic alteration frequency of ABCC5 confirmed its potential value in LIHC. In addition, several drugs were explored and found to be relevant to LIHC. The t-SNE showed that expression of ABCC5 was most concentrated in macrophages, followed by hepatocytes.
Conclusion: ABCC5 may facilitate LIHC progression through different mechanisms and be a potential biomarker and target for diagnosis, prognosis, and therapy of LIHC.
Keywords: LIHC, bioinformatics analysis, ATP-binding cassette transporter family, ABCC5
Introduction
Liver cancer is the fifth most common type of cancer worldwide. According to statistics, the incidence of liver cancer accounted for 4.7% of cancers in 2018, while the number of deaths caused by liver cancer accounted for 8.2%, ranking it the fourth leading cause of cancer-related mortality worldwide.1 Liver cancer tends to occur in elderly males with a male-to-female incidence ratio of approximately 2–2.5:1. Histopathology subtyping has shown that liver hepatocellular carcinoma (LIHC) represents about 90% of primary hepatic carcinomas.2 As a result, LIHC is a serious and challenging global health problem. The major causes of LIHC include hepatitis C or B, alcoholic and non-alcoholic fatty liver disease (NAFLD), and metabolic syndrome (MS), which is a pathological cluster of metabolic disorders that may also contribute to LIHC.3,4 One of the reasons for its high mortality is the difficulty in detecting LIHC in an early stage. Currently, the treatment for LIHC consists of radical liver resection, liver transplantation, transcatheter arterial chemoembolization, radiofrequency ablation, radioembolization, and tyrosine-kinase inhibitors.4
Despite the great amount of attention for biomarkers, no effective new biomarkers for diagnosis and surveillance of LIHC have been discovered. The recommended biomarkers for clinical usage such as Alpha-Fetoprotein (AFP), Alpha-Fetoprotein Lens Culinaris Agglutinin-3 (AFP-L3), and Des-γ-carboxy prothrombin (DFP) were insufficient in sensitivity and specificity.5 Therefore, exploring and confirming new biomarkers is of great significance for early diagnosis and prognosis of LIHC.
Transporters have an essential role to play in various physiological processes. The human adenosine triphosphate (ATP)-binding cassette (ABC) transporter family is one of the largest gene families of transporters.6 The ABC transporters are a type of ATP-powered pump and include 49 ABC genes which are divided into seven subfamilies (ABCA, ABCB, ABCC, ABCD, ABCE, ABCF, and ABCG). In humans, the ABC transporter family is mainly expressed in the liver, gut, kidney, placenta, and blood-brain and blood-testis barrier. They contribute to the transport of many endogenous substances, such as peptides, metal ions, and inorganic anions across the plasma as well as intracellular membranes.7 It has been reported that various mutations in genes of the ABC family can lead to different genetic diseases. For instance, mutations in the ABCA4 gene contribute to Stargardt macular degeneration, progressive familial intrahepatic cholestasis is caused by ABCB11 deficiency, and ABCC7, which is also known as cystic fibrosis transmembrane conductance regulator, is linked to cystic fibrosis.8–10 ABC transporters are also involved in absorbing, distributing, and excreting the drugs and their metabolites,11 which is associated with drug resistance of cancer cells. For example, ABCB1 is confirmed to be involved in multidrug resistance (MDR) and it can also be named MDR.6 In addition, recent studies have discovered other functions of the ABC family in cancers. For example, ABCB6 was observed to be over-expressed in LIHC and accelerate tumor progression by inhibiting ferroptosis.12 They may be responsible for the spread and metastasis of tumor cells as well, which presents great potential for treatment purposes.13
The ABCC transporter family comprises the ABC subfamilies and consists of 13 transporters, most of which belong to a large group of multidrug resistance-associated proteins (MRPs).14 It has been reported that MRPs are related to tumor resistance to many kinds of anti-cancer drugs and their metabolites.14,15 The ABCC transporters act as ion channels for ion transport and have diverse functions, including toxin secretion and signal transduction.7
In the study, we made a comparison of the expression of all members of the ABC transporter family between LIHC and normal tissues and selected one or several potential biomarkers for LIHC. Then we explored the potential function and value of these biomarkers by several public databases. We analyzed the differences in expression of biomarkers between tumor and normal liver tissue, and further explored their prognostic value, co-expression pattern, functional pathways, correlation with immune infiltration, genetic alteration frequency, drug sensitivity, and distribution in LIHC. This study evaluates the potential of the ABC transporter family in diagnosis and surveillance of LIHC, and being targets for anti-cancer therapy in LIHC.
Materials and Methods
Selection of the Potential Biomarker for LIHC
On one hand, ONCOMINE (https://www.oncomine.org/resource/main.html) is a comprehensive web resource providing practical RNA expression analyses,16 and UALCAN (http://ualcan.path.uab.edu/) is a comprehensive network resource that supplies The Cancer Genome Atlas (TCGA)-based analysis.17 The transcriptional levels of each gene in the ABC transporter family are firstly compared between LIHC tumor and normal tissues with data from ONCOMINE, and the differentially expressed genes are selected with p < 0.05. Then, transcriptional levels of these genes are further compared between the two kinds of tissues with UALCAN to confirm the differentially expressed genes.
On the other hand, GEPIA (http://gepia.cancer-pku.cn/index.html) is designed by Peking University and is a database that is used to analyze the expression of RNA sequences.18 It is used to evaluate the relationship between the expression of the members in ABC transporter gene family and the LIHC patients’ pathological stage. The related genes and the correlation are determined.
Combining the results of the two steps, the genes which are both differentially expressed between LIHC tumor and normal tissues, and related to LIHC patients’ pathological stage are defined as our potential biomarkers for LIHC.
Preliminary Validation of the Potential Biomarker for LIHC
The expression of the selected potential biomarkers was compared between various kinds of cancers and the corresponding normal tissues to evaluate their value in tumors. The genes are thought to be valuable in tumors with significant differentially expression (p < 0.0001). Finally, the expression level of the eventually selected genes were compared between LIHC tumor and normal tissues once again in GEPIA database.
PCA Dimension Reduction Analysis
The expression of ABCC5 varies between individual patients of LIHC. Principal component analysis (PCA) dimension reduction was carried out to distinguish them based on the expression of ABCC5. The number of PCs is set to 2, and the clusters are presented in uMAP format. We differentiate these clusters into high-ABCC5 expression subgroup and low-ABCC5 expression subgroup.
The Prognostic Prediction Value of ABCC5 in LIHC
On the basis of high- and low-ABCC5 expression subgroups, GEPIA was applied to evaluate the prognostic prediction value of ABCC5 in LIHC. We conducted a Kaplan–Meier (KM) analysis in order to explore the difference in overall survival (OS) and disease-free survival (DFS) of patients between the two subgroups. Cox regression analyses were used to tell whether the expression level of ABCC5 is an independent risk factor for OS among LIHC patients.
Co-Expression Pattern and Functional Enrichment Analyses of ABCC5 in LIHC
Since the ABCC5 expression may also be influenced by other genes, we constructed a co-expression network with UALCAN to clarify its function and clinical value in LIHC development. DAVID (http://david.ncifcrf.gov) is a comprehensive bioinformatics tool in which the biological functions of provided genes are clarified.19 ABCC5 and other genes in the ABC transporter family were put into Gene Ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis.
Correlation Between Immune Infiltration and Expression of ABCC5 in LIHC
TIMER (https://cistrome.shinyapps.io/timer/) is a reliable and intuitive resource to systematically evaluate the infiltration and clinical effects of immune cells.20 In our study, the “SCNA module” was used to analyze the relationship between immune cell infiltration levels and the number of ABCC5 gene copies; additionally, the “Gene module” was applied to assess the correlation between ABCC5 and immune infiltration. The data from the cancer genome atlas (TCGA) were taken as a training group while the data from GSE112790 were used for validation.
Genetic Alteration Frequency of ABCC5 Across Cancers
cBioPortal (www.cbioportal.org), a bioinformatics service, can perform the visualization and analysis of multidimensional cancer genomics data.21 On the basis of TCGA, genetic alterations and co-expression of ABCC5 were obtained from cBioPortal. Three hundred and sixty LIHC samples from Firehose legacy and 348 LIHC samples from PanCancer Atlas are analyzed, respectively, to examine the genetic alteration of ABCC5 across LIHC.
Prediction of Drug Sensitivity Associated with ABCC5 in LIHC
To confirm the relationship between our gene and drug responses, GDSC (https://www.cancerrxgene.org/) and CTRP (http://portals.broadinstitute.org/ctrp/) database were used to analyze the relationship between half inhibitory concentration (IC50) data of different molecules and the gene expression levels. The criteria were set as FDR < 0.05.
Distribution of ABCC5 Expressed Cells in LIHC Tissues
LIHC tumor tissues were composed of different cells. The t-SNE algorithm was employed to explore the distribution of cells which express ABCC5 in LIHC tissues. The criteria were set as log2 [fold change (FC)] > 0.5 and FDR < 0.05.
Results
Expression of ABC Transporter Family in LIHC
Firstly, we probed the transcriptional levels of the ABC transporter family in LIHC and normal liver tissues with ONCOMINE, and twelve members of the ABC transporter family were retrieved (Figure 1A). Then the same analysis of transcriptional levels of these genes was performed with another database, UALCAN. Eight genes selected from ONCOMINE were as well differently expressed in UALCAN. In detail, ABCB2, ABCB3, ABCC4, and ABCC5 in LIHC samples were drastically elevated while the levels of ABCA6, ABCA8, ABCB4, and ABCD3 were significantly reduced in LIHC compared to normal liver tissues (p < 0.05, Figure 1B). Then, we evaluated the relationship between the expression of the ABC transporter family members and the LIHC patients’ pathological stage. A significant correlation between the level of ABCC1, ABCC5, ABCE1, ABCF2, and the pathological stage was found (p < 0.05, Figure 1C). As the tumor progressed, the level of ABCC1, ABCC5, and ABCF2 increased.
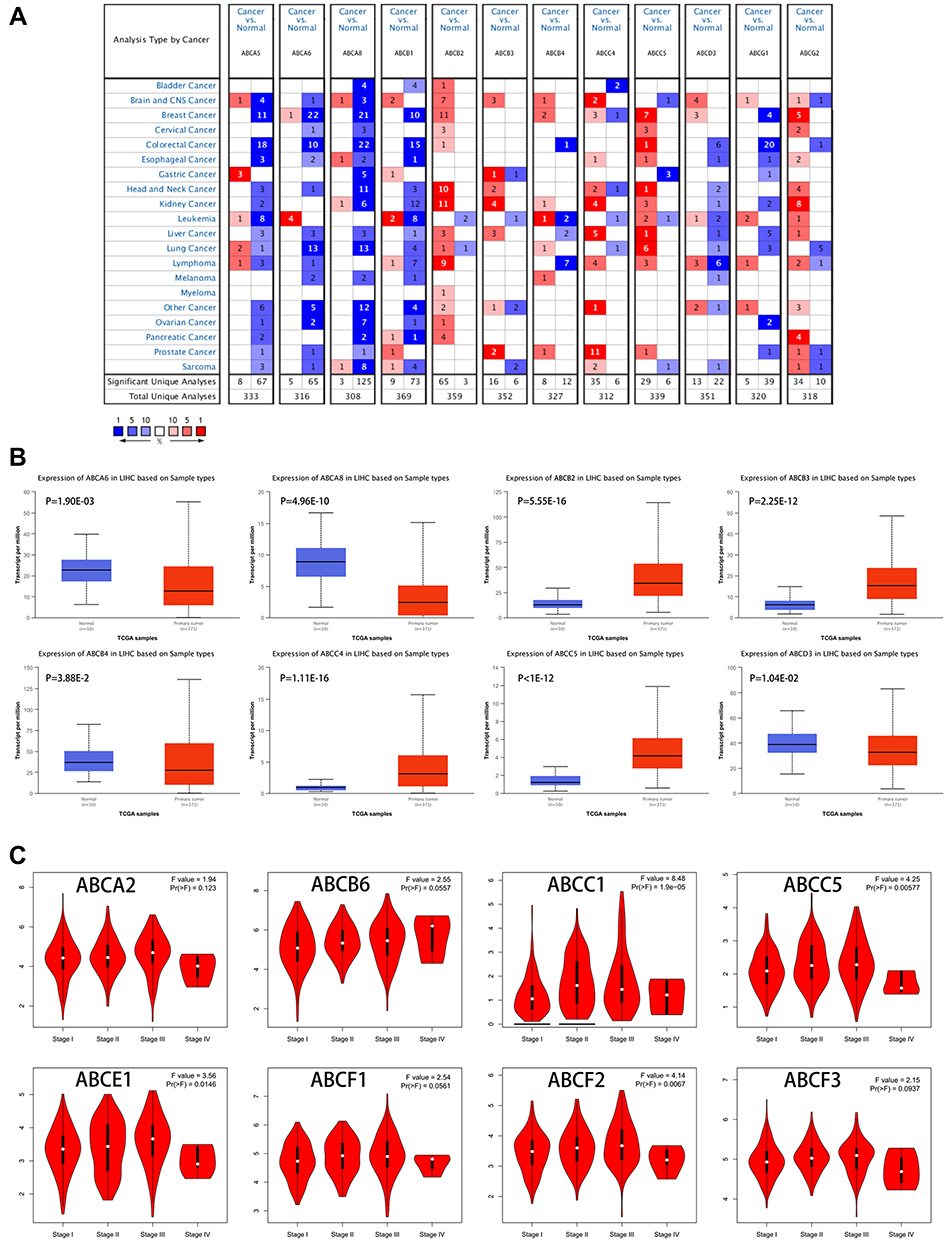 |
Figure 1 (A) mRNA levels of the ATP-binding Cassette transporter family in LIHC (ONCOMINE). The figure shows the number of datasets with statistically significant mRNA overexpression (red) or downregulated expression (blue) of the ABC transporter family. (B) Transcription of the ABC transporter family in LIHC (UALCAN). (C) Stage plot showing the correlation between the ABC transporter family expression and tumor stage in LIHC (GEPIA). |
Notably, ABCC5 showed significant differences in LIHC patients compared to healthy samples and seemed to have an important part to play in LIHC progression; therefore, we presumed that ABCC5 may be a potential disease biomarker for LIHC.
Expression of ABCC5 Across Cancers and in LIHC
UALCAN was used to analyze ABCC5 expression across various types of cancers. The results indicated that ABCC5 was significantly up-regulated in multiple cancers, including LIHC. Furthermore, compared with different kinds of cancers, there is a relatively more significant difference between the tumor and relevant controls in LIHC (Figure 2A). Then the specific analysis of ABCC5 expression in LIHC revealed that the expression of ABCC5 was exactly elevated in LIHC samples in contrast to normal liver samples (p < 0.0001, Figure 2B), and the results of “Gene Comparison” demonstrated that the expression of ABCC5 in tumor tissues was approximately 2.2 times higher than that in healthy controls (Figure 2C).
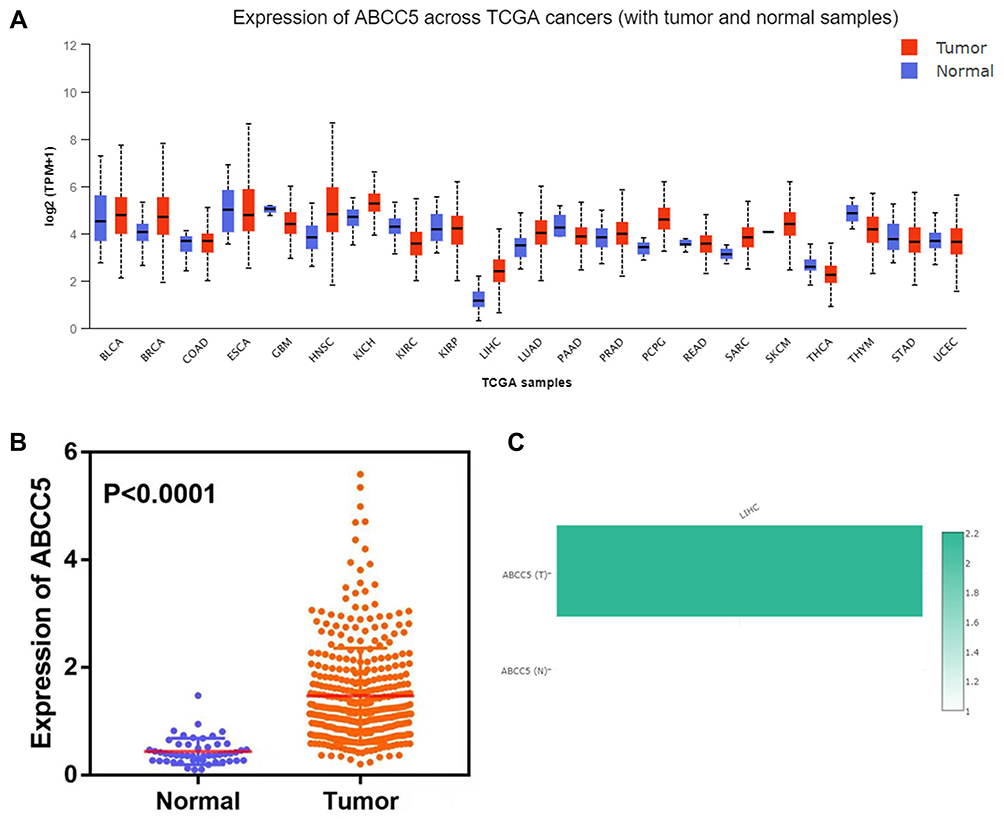 |
Figure 2 (A) Expression of ABCC5 across cancers identified by the TCGA: ABCC5 was significantly differently expressed in LIHC compared to normal tissue. (B) Analysis of 369 LIHC and 50 normal tissues showed that ABCC5 was significantly overexpressed in LIHC (P < 0.0001). (C) Expression of ABCC5 in LIHC as found in the GEPIA database: the expression of ABCC5 in LIHC was approximately 2.2 times higher than that in normal tissue. |
The Prognostic Prediction Value of ABCC5 in LIHC
The PCA divided patients of LIHC into high- and low-expression of ABCC5 subgroups (Figure 3A). Then we conducted a KM analysis in order to explore the prognostic prediction value of ABCC5 in LIHC. We found that LIHC patients in low-expression subgroup had a more prolonged OS (p = 0.000049, Figure 3B), while their DFS was not affected (p = 0.072, Figure 3C). Cox regression analyses showed that the expression level of ABCC5 was an independent risk factor for OS among LIHC patients as well (Table 1).
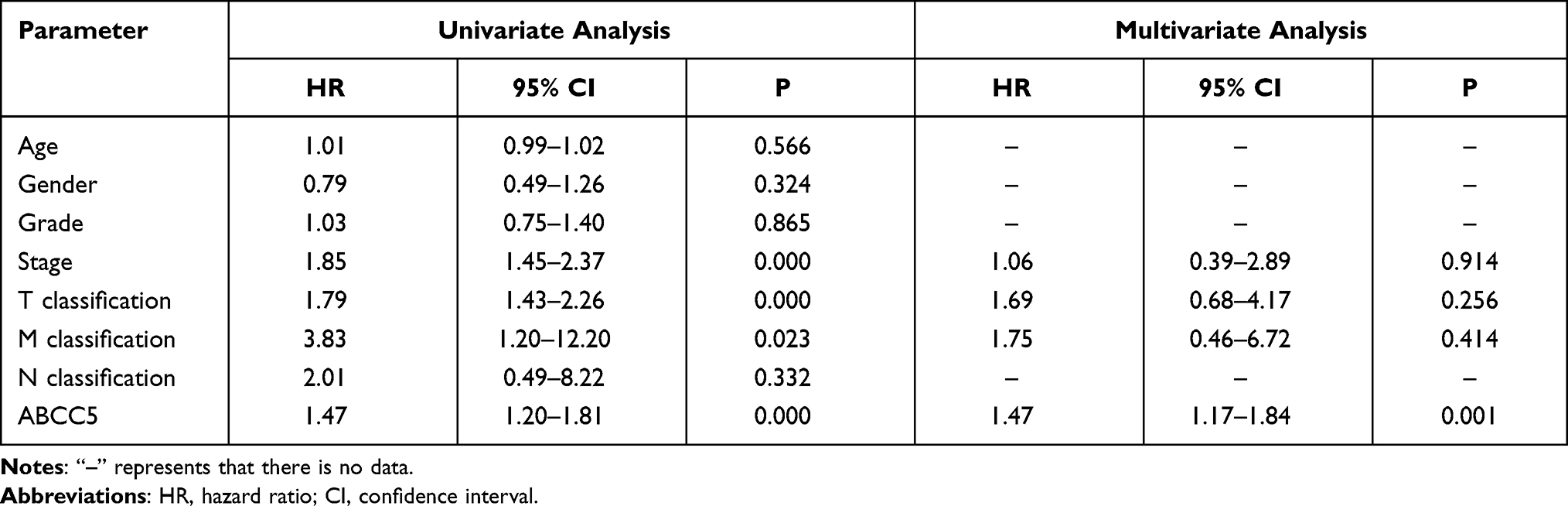 |
Table 1 Univariate and Multivariate Cox Analyses of ABCC5 Expression in LIHC |
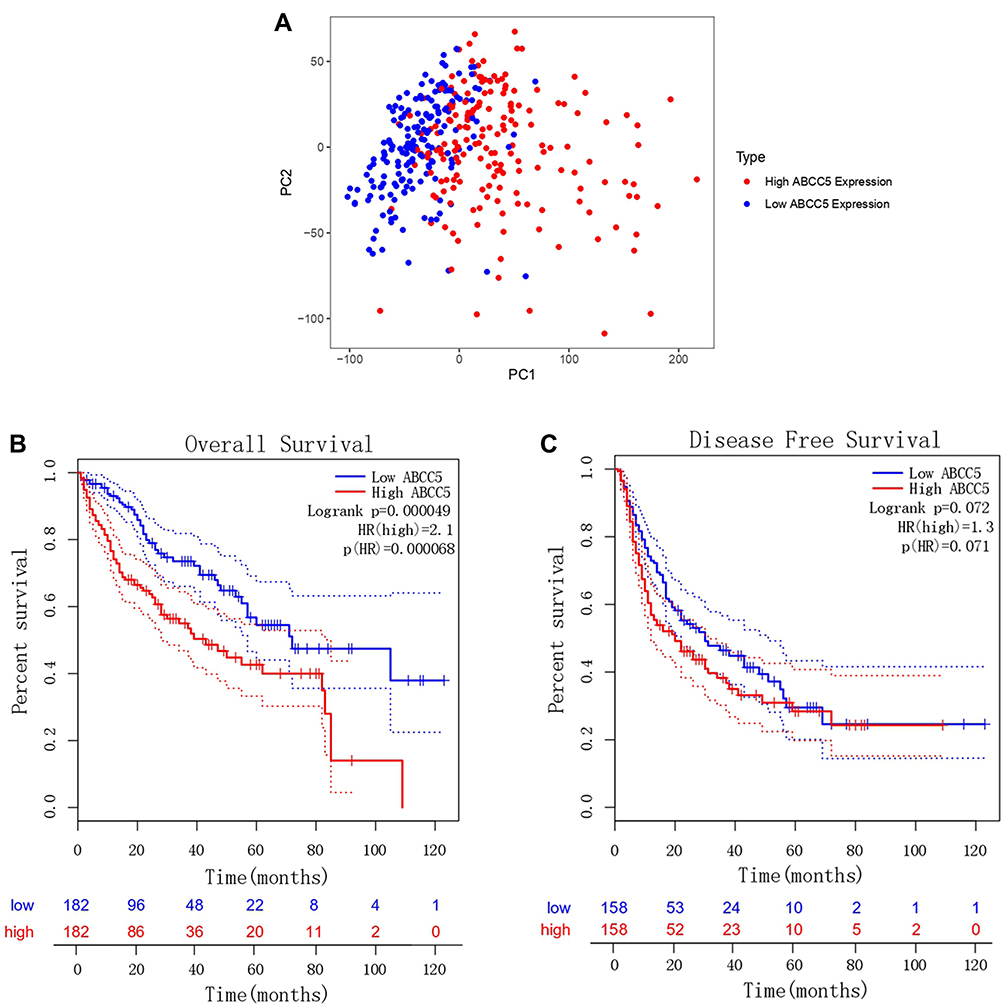 |
Figure 3 (A) Principal component analysis of ABCC5 expression in LIHC. (B) Overall survival analysis of ABCC5 expression in LIHC (GEPIA). (C) Disease-free survival analysis of ABCC5 expression in LIHC (GEPIA). |
Co-Expression Pattern of ABCC5 in LIHC
The co-expression network of ABCC5 is constructed with the application of UALCAN. The top 100 genes co-expressed with ABCC5 were displayed in heat maps, of which ABCC5, YEATS2, SLC36A1, KIAA0226, PLXNA1, VPS8, MTMR2, ACTR3, SIN3B, and ARPC2 rank the top ten (Figure 4A, Supplementary Figure 1A-C). The pattern may reveal the function of ABCC5 in the development of LIHC and can be utilized in further functional studies of ABCC5.
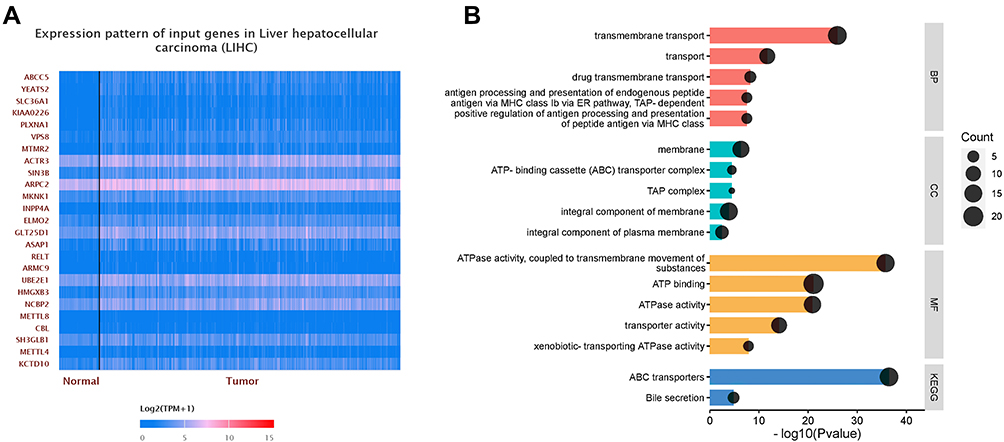 |
Figure 4 (A) The heat map of the top 25 co-expressed genes with ABCC5. (B) Functional enrichment analyses of ABCC5 and the co-expression genes in LIHC. GO enrichment analysis showed that the co-expressed genes were significantly enriched in BP, CC, and MF of LIHC. KEGG analysis showed that co-expression genes were mainly enriched in ABC transporters and bile secretion. |
GO and KEGG Enrichment Analyses of Co-Expressed Genes
We performed GO and KEGG enrichment analyses to identify the potential functions that involve genes in ABC transporter family. The genes were closely associated with transmembrane transport (especially drugs’), antigen processing and presentation in biological process (BP). While in cellular component (CC), they were majorly enriched in membrane, ABC transporter complex, integral component of membrane, integral component of plasma membrane, and TAP complex. Furthermore, in molecular function (MF), these genes were significantly linked with ATPase activity and transporter activity. KEGG analyses were also conducted, and the results showed that the ABC transporters were the most highly enriched pathway as expected, followed by bile secretion (Figure 4B). The results of functional analyses were consistent with ABCC5 as a member of ABC transporter family, and pointed to its function in transmembrane transporting, especially in drug transmembrane transporting, in LIHC progression.
Correlation Between Immune Infiltration and Expression of ABCC5 in LIHC
We conducted analyses with TIMER to evaluate the potential correlation between the expression of ABCC5 and immune infiltration in LIHC. Firstly, the “SCNA” module analysis indicated that the immune infiltration levels of some cells were related to altered copy numbers of ABCC5, such as neutrophils and macrophages (Figure 5A). Secondly, we took data from TCGA as a training group and those from GSE112790 as a validating group. ABCC5 expression appeared to have no apparent relationship with tumor purity, while it was significantly positively correlated with the infiltration levels of macrophages, neutrophils, and dendritic cells in LIHC in both training and validating groups (p < 0.05, Figure 5B). The results implied that ABCC5 may influence the progression and prognosis of LIHC in the way of regulating the level of immune cell infiltration.
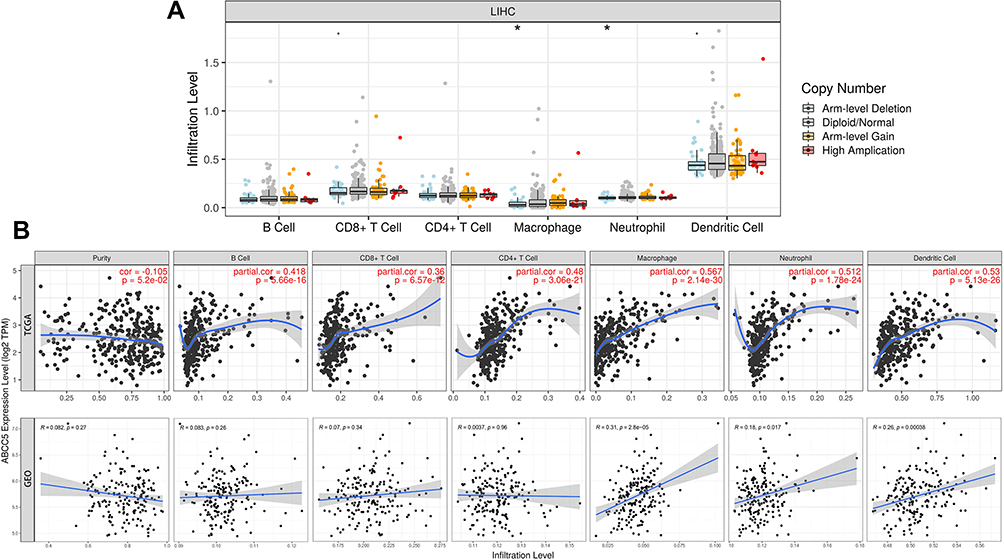 |
Figure 5 (A) Altered ABCC5 gene copy numbers was significantly correlated with the immune infiltration levels of several types of immune cell in LIHC (PanCancer Atlas) (*p < 0.05). (B) Immune infiltration linked to ABCC5 in LIHC (TCGA) as training: the expression of ABCC5 was significantly correlated with the infiltration of B cells (p = 5.66e-16), CD8 + T cells (p = 6.57e-12), CD4 + T cells (p = 3.06e-21), macrophages (p = 2.14e-30), neutrophils (p = 1.78e-24), and dendritic cells (p = 5.13e-26). Immune infiltration linked to ABCC5 in LIHC (GSE112790) as validation: the expression of ABCC5 was significantly correlated with the infiltration of macrophages (p = 2.8e-5), neutrophils (p = 0.017), and dendritic cells (p = 0.00038). |
Genetic Alteration Frequency and Drug Sensitivity of ABCC5 in LIHC
We investigated the alteration frequency of the ABCC5 gene in LIHC with data from PanCancer Atlas and Firehose legacy. The results showed that the alteration frequency of ABCC5 in LIHC was 7% and 8%, respectively (Figure 6A, Supplementary Figure 1D). There is a positive correlation between the copy number of ABCC5 and mRNA expression in LIHC tissues (Figure 6B). In addition, we found LIHC samples with deletions showed lower mRNA expression than those with diploids (Figure 6C). In the aspect of drug sensitivity, we applied GDSA and CTPR database to depict the relationships between ABCC5 expression and drug responses. The bubble heat map showed that high expression levels of ABCC5 had significant correlations with lower half inhibitory concentration (IC50) data in GDSC and CTRP database, respectively, indicating that ABCC5 exhibited drug sensitivity, including Afatinib, Erlotinib, Gefitinib and Lapatinib (Figure 6D).
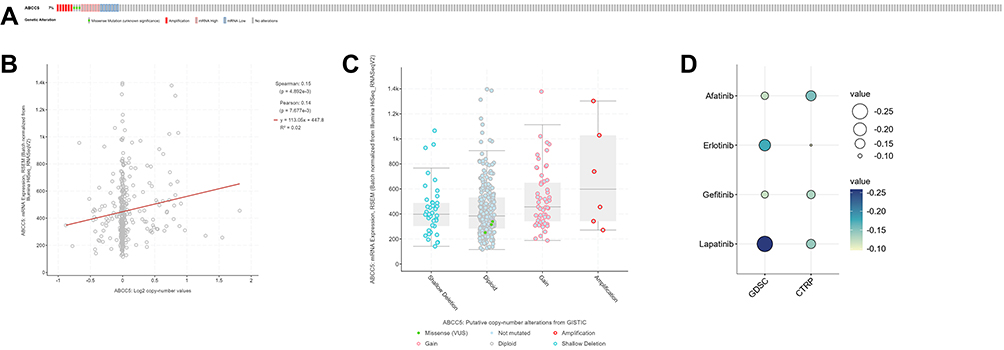 |
Figure 6 (A) ABCC5 alterations in the LIHC cohort. The different types of genetic alterations are highlighted in different colors. (B) Correlation between ABCC5 expression and the copy number of ABCC5 in LIHC. (C) LIHC samples with deletions showed lower mRNA expression than those with diploids. (D) Prediction of drug sensitivity associated with ABCC5 in LIHC, including Afatinib, Erlotinib, Gefitinib, and Lapatinib. |
Distribution of ABCC5 Expressed Cells in LIHC Tissues
Finally, we tried to explore the exact distribution of ABCC5 expression in different kinds of cells in LIHC tissues. The graphs obtained by t-SNE algorithm revealed that expression of ABCC5 was most concentrated in macrophages, followed by hepatocytes (Figure 7). The gathering expression of ABCC5 in macrophages was consistent with the result of our immune infiltration analysis, and the high expression of ABCC5 in hepatocytes hinted its potential role in LIHC.
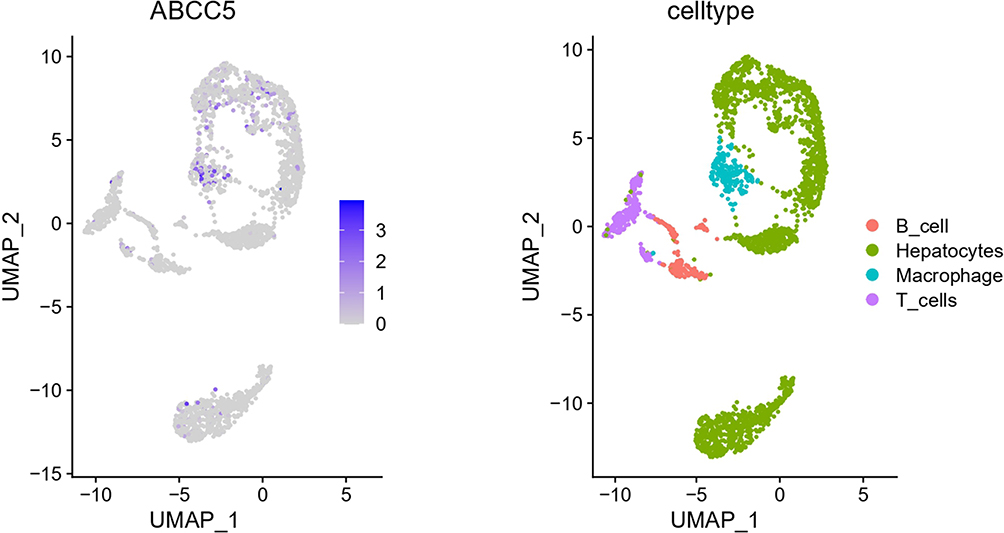 |
Figure 7 Single-cell analysis revealed the distribution of ABCC5 expressed cells in LIHC tissues. |
Discussion
It has been widely accepted that members of the ABC transporter family are related to various diseases, including cancers, and were suspected to influence proliferation, differentiation, migration, and invasion of tumor cells.13 For example, ABCG2, also known as the breast cancer resistance protein (BCRP), has functions in the development of breast cancer.22 Furthermore, this family is confirmed to be involved in multidrug resistance, which hampers the effects of anti-cancer pharmacotherapy.23 Liver cancer, of which LIHC is the most common type, is a considerable threat to public health due to its high mortality. In a study by Siegel et al, survival ranked second lowest for liver cancer (20%), just behind pancreas cancer (10%).24 Several studies have depicted the relationship between ABC transporters and LIHC. For example, Zhang et al found that low expression of ABCA8 and ABCA9 shortens the survival time of patients, while over-expression of ABCB6 in LIHC was associated with the acceleration of tumor progression by inhibiting ferroptosis.12
We researched the aberrant expression of the ABC transporter family in LIHC samples from the ONCOMINE and UALCAN databases. The differently expressed genes between LIHC and normal tissues in both databases were selected. Simultaneously, we evaluated the correlation between the expression of ABC transporter genes and the pathological stages of LIHC patients. The results showed that the expression of ABCC1, ABCC5, ABCE1, and ABCF2 increased as LIHC progressed. Combined with the statistical analyses above, ABCC5 is expressed differently in LIHC tissues and is strongly related to the pathological stage. Therefore, we selected this gene to further explore its diagnostic, prognostic, and therapeutic value as well as mechanisms in LIHC.
In the human liver, ABCC transporter proteins are differentially expressed and regulated and involved in eliminating toxic metabolites.14 In the ABCC transporter subfamily, ABCC8 gene is related to congenital hyperinsulinism of infancy.25 It has been identified that ABCC5 is a member of the MRP subfamily and is associated with resistance to several nucleosides. It is involved in cGMP secretion as well.6 A previous study has also confirmed that dramatically increased levels of ABCC5 existed in LIHC.26
The PCA divided patients of LIHC into high- and low-expression subgroups. Our KM analysis demonstrated that LIHC patients with low transcriptional levels of ABCC5 tend to have a more prolonged OS, and both Cox analyses identified ABCC5 expression as an independent risk factor of OS in LIHC. Subsequently, the construction of a co-expression pattern identified the genes co-expressed with ABCC5 for functional analyses. Both GO and KEGG analyses showed that genes were enriched in pathways related to transmembrane transport, including drug transmembrane transport. Many anti-cancer drugs require transmembrane transporters to access the cancer cells and act on intracellular targets, for which ABC transporters are responsible.27 In addition to directly affecting drug efficacy by regulating the uptake of endogenous compounds and xenobiotics, transmembrane transporters can also influence cellular metabolic processes like glycolysis and oxidative phosphorylation.28–30 Therefore, it is certain that an abnormal expression of the ABC transporter family could interfere with drug absorption and metabolism. ABCC5 was reported to be associated with the tumor response to gemcitabine-based chemoradiotherapy, which ultimately affects patients’ survival.31 Other enriched pathways included ATPase activity and bile secretion. ABC proteins contain nucleotide-binding domains (NBDs), which are ATPase subunits.32 Moreover, a defect in the bile acid secreted by the liver may also contribute to LIHC oncogenesis.33
Analyses of correlation between ABCC5 expression in LIHC and immune infiltration illustrated that the ABCC5 expression level was statistically significant related to cell infiltration of several types of immune cells, including macrophages, neutrophils, and dendritic cells (DCs). It has been reported that a few ABC transporters are involved in cellular processes such as antigen processing and immunity.15 Macrophages are equally important in LIHC. Macrophages have an essential part in chronic liver inflammation, which is a crucial step in the initiation and progression of LIHC. Tumor-associated macrophages (TAMs) are a well-known component of the tumor microenvironment, the majority of which are M2-polarized macrophages. Yao et al found that M2-polarized macrophages promoted the migration and epithelial-mesenchymal transition of LIHC cells by regulating the TLR4/STAT3 signaling pathway.34,35 In addition, neutrophils are also accumulated in and become a major component of the tumor microenvironment of LIHC. It has been demonstrated that they promote LIHC development by inducing the production of the pro-metastatic factor OSM.36 DCs are a type of antigen-presenting cells. The lack of crosstalk between DCs is one of the causes of tumor tolerance in LIHC.37 Hence, the elevation of ABCC5 expression levels in LIHC patients may suggest a potential target in immunotherapy, which could benefit the patients’ prognosis.38
The high frequency of genetic alterations in the ABCC5 gene and the significant difference in ABCC5 protein levels between tumor and normal samples both confirm the sensitivity of ABCC5 as a biomarker for LIHC at the gene and protein levels. There have been reports of numerous new biomarkers for the diagnosis and prognosis of LIHC, such as lncRNA-D16366, deoxycytidine kinase, and circ_0067934.39–41 Though all these new biomarkers lack sufficient clinical testing and validation, they may have the potential to become diagnostic, prognostic, and therapeutic targets in LIHC and provide insights into the mechanisms of initiation and progression of LIHC.
At present, cancer treatment has entered the era of precision medicine. As mentioned previously, ABCC5 may play an important role in LIHC, so it is crucial to explore drugs related to ABCC5. We identified 4 different types of drugs that were significantly sensitive to ABCC5 in LIHC. It is beneficial to improve the prognosis of LIHC patients and explore the mechanism of ABCC5.
This study has several limitations that should be addressed. Firstly, the data we applied for statistical analyses were obtained from public databases and did not reflect authentic clinical conditions. Secondly, we moderately investigated the prognostic value of ABCC5 in LIHC and more in-depth research is required to validate its clinical application value. Thirdly, prospective studies should be performed since our study has been retrospective. Fourthly, clinical tissues for validation were not included in this study and further experiments with animal and human samples are necessary to clarify the actual underlying mechanisms and relationships between ABCC5 and LIHC.
Conclusion
In the bioinformatic analysis, ABCC5 is the only member in ABC transporter family that is both differentially expressed between LIHC and normal liver tissues, and strongly related to pathological stage of LIHC. ABCC5 is confirmed as an independent risk factor for LIHC patients. It may facilitate LIHC progression through different mechanisms, including transmembrane transport and immune infiltration. ABCC5 may be a potential biomarker and target for diagnosis, prognosis, and therapy of LIHC.
Acknowledgments
This research was supported by the Digestive Medical Coordinated Development Center of Beijing Hospitals Authority under Grant No. XXZ0105; Special Scientific Research Fund for Tutor under Grant No. YYDSZX201901; Medical and Health Public Foundation of Beijing under Grant No. YWJKJJHKYJJ-B17262-067; and the Science and Technology Development Project of China State Railway Group under Grant No. N2019Z004. We thank the editors and reviewers for their comments and instruction, and thank each other for the support and encouragement in the writing process.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. European Association for the Study of the Liver. Electronic address: [email protected]. EASL clinical practice guidelines: management of hepatocellular carcinoma. J Hepatol. 2018;69(1):182–236.
3. Kulik L, El-Serag HB. Epidemiology and management of hepatocellular carcinoma. Gastroenterology. 2019;156(2):477–491.e1. doi:10.1053/j.gastro.2018.08.065
4. Uzunlulu M, Telci Caklili O, Oguz A. Association between metabolic syndrome and cancer. Ann Nutr Metab. 2016;68(3):173–179. doi:10.1159/000443743
5. Piñero F, Dirchwolf M, Pessôa MG. Biomarkers in hepatocellular carcinoma: diagnosis, prognosis and treatment response assessment. Cells. 2020;9(6):1370. doi:10.3390/cells9061370
6. Dean M, Hamon Y, Chimini G. The human ATP-binding cassette (ABC) transporter superfamily. J Lipid Res. 2001;42(7):1007–1017. doi:10.1016/S0022-2275(20)31588-1
7. Vasiliou V, Vasiliou K, Nebert DW. Human ATP-binding cassette (ABC) transporter family. Hum Genomics. 2009;3(3):281–290. doi:10.1186/1479-7364-3-3-281
8. Meng X, Clews J, Martin ER, et al. The structural basis of cystic fibrosis. Biochem Soc Trans. 2018;46(5):1093–1098. doi:10.1042/BST20180296
9. Lenis TL, Hu J, Ng SY, et al. Expression of ABCA4 in the retinal pigment epithelium and its implications for stargardt macular degeneration. Proc Natl Acad Sci U S A. 2018;115(47):E11120–E11127. doi:10.1073/pnas.1802519115
10. Li LT, Li ZD, Yang Y, et al. ABCB11 deficiency presenting as transient neonatal cholestasis: correlation with genotypes and BSEP expression. Liver Int. 2020;40(11):2788–2796. doi:10.1111/liv.14642
11. Liu X. ABC family transporters. Adv Exp Med Biol. 2019;1141:13–100.
12. Zhang J, Zhang X, Li J, et al. Systematic analysis of the ABC transporter family in hepatocellular carcinoma reveals the importance of ABCB6 in regulating ferroptosis. Life Sci. 2020;257:118131. doi:10.1016/j.lfs.2020.118131
13. Pasello M, Giudice AM, Scotlandi K. The ABC subfamily A transporters: multifaceted players with incipient potentialities in cancer. Semin Cancer Biol. 2020;60:57–71. doi:10.1016/j.semcancer.2019.10.004
14. Gu X, Manautou JE. Regulation of hepatic ABCC transporters by xenobiotics and in disease states. Drug Metab Rev. 2010;42(3):482–538. doi:10.3109/03602531003654915
15. Liu X. Transporter-mediated drug-drug interactions and their significance. Adv Exp Med Biol. 2019;1141:241–291.
16. Rhodes DR, Yu J, Shanker K, et al. ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia. 2004;6(1):1–6. doi:10.1016/S1476-5586(04)80047-2
17. Chandrashekar DS, Bashel B, Balasubramanya SAH, et al. UALCAN: a portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia. 2017;19:649–658. doi:10.1016/j.neo.2017.05.002
18. Tang Z, Li C, Kang B, et al. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–W102. doi:10.1093/nar/gkx247
19. Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44–57. doi:10.1038/nprot.2008.211
20. Li T, Fan J, Wang B, et al. TIMER: a web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Res. 2017;77(21):e108–e110. doi:10.1158/0008-5472.CAN-17-0307
21. Gao J, Aksoy BA, Dogrusoz U, et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013;6(269):pl1. doi:10.1126/scisignal.2004088
22. Gil-Martins E, Barbosa DJ, Silva V, et al. Dysfunction of ABC transporters at the blood-brain barrier: role in neurological disorders. Pharmacol Ther. 2020;213:107554.
23. Chen Z, Shi T, Zhang L, et al. Mammalian drug efflux transporters of the ATP binding cassette (ABC) family in multidrug resistance: a review of the past decade. Cancer Lett. 2016;370(1):153–164. doi:10.1016/j.canlet.2015.10.010
24. Siegel RL, Miller KD, Fuchs HE, et al. Cancer statistics, 2021. CA Cancer J Clin. 2021;71(1):7–33.
25. De Franco E, Saint-Martin C, Brusgaard K, et al. Update of variants identified in the pancreatic β-cell KATP channel genes KCNJ11 and ABCC8 in individuals with congenital hyperinsulinism and diabetes. Hum Mutat. 2020;41(5):884–905. doi:10.1002/humu.23995
26. Wurmbach E, Chen YB, Khitrov G, et al. Genome-wide molecular profiles of HCV-induced dysplasia and hepatocellular carcinoma. Hepatology. 2007;45(4):938–947. doi:10.1002/hep.21622
27. Girardi E, César-Razquin A, Lindinger S, et al. A widespread role for SLC transmembrane transporters in resistance to cytotoxic drugs. Nat Chem Biol. 2020;16(4):469–478. doi:10.1038/s41589-020-0483-3
28. Jetter A, Kullak-Ublick GA. Drugs and hepatic transporters: a review. Pharmacol Res. 2020;154:104234. doi:10.1016/j.phrs.2019.04.018
29. Icard P, Shulman S, Farhat D, et al. How the warburg effect supports aggressiveness and drug resistance of cancer cells? Drug Resist Updat. 2018;38:1–11. doi:10.1016/j.drup.2018.03.001
30. Reczek CR, Birsoy K, Kong H, et al. A CRISPR screen identifies a pathway required for paraquat-induced cell death. Nat Chem Biol. 2017;13(12):1274–1279. doi:10.1038/nchembio.2499
31. Flegel WA, Srivastava K, Sissung TM, et al. Pharmacogenomics with red cells: a model to study protein variants of drug transporter genes. Vox Sang. 2021;116(2):141–154. doi:10.1111/vox.12999
32. Theodoulou FL, Kerr ID. ABC transporter research: going strong 40 years on. Biochem Soc Trans. 2015;43(5):1033–1040. doi:10.1042/BST20150139
33. Jia W, Xie G, Jia W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat Rev Gastroenterol Hepatol. 2018;15(2):111–128. doi:10.1038/nrgastro.2017.119
34. Tian Z, Hou X, Liu W, et al. Macrophages and hepatocellular carcinoma. Cell Biosci. 2019;9(1):79. doi:10.1186/s13578-019-0342-7
35. Yao -R-R, Li J-H, Zhang R, et al. M2-polarized tumor-associated macrophages facilitated migration and epithelial-mesenchymal transition of HCC cells via the TLR4/STAT3 signaling pathway. World J Surg Oncol. 2018;16(1):9. doi:10.1186/s12957-018-1312-y
36. Peng ZP, Jiang ZZ, Guo HF, et al. Glycolytic activation of monocytes regulates the accumulation and function of neutrophils in human hepatocellular carcinoma. J Hepatol. 2020;73(4):906–917. doi:10.1016/j.jhep.2020.05.004
37. Lurje I, Hammerich L, Tacke F. Dendritic cell and T cell crosstalk in liver fibrogenesis and hepatocarcinogenesis: implications for prevention and therapy of liver cancer. Int J Mol Sci. 2020;21(19):7378. doi:10.3390/ijms21197378
38. Cao J, Kong F-H, Liu X, et al. Immunotherapy with dendritic cells and cytokine-induced killer cells for hepatocellular carcinoma: a meta-analysis. World J Gastroenterol. 2019;25(27):3649–3663. doi:10.3748/wjg.v25.i27.3649
39. Chao Y, Zhou D. lncRNA-D16366 is a potential biomarker for diagnosis and prognosis of hepatocellular carcinoma. Med Sci Monit. 2019;25:6581–6586. doi:10.12659/MSM.915100
40. Song D, Wang Y, Zhu K, et al. DCK is a promising prognostic biomarker and correlated with immune infiltrates in hepatocellular carcinoma. World J Surg Oncol. 2020;18(1):176. doi:10.1186/s12957-020-01953-1
41. Zhou C, Li R, Mi W. circ_0067934: a potential biomarker and therapeutic target for hepatocellular carcinoma. Ann Clin Lab Sci. 2020;50(6):734–738.
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.