")
Back to Journals » OncoTargets and Therapy » Volume 12
Hypoxic non-small-cell lung cancer cell-derived exosomal miR-21 promotes resistance of normoxic cell to cisplatin
Authors Dong C, Liu X, Wang H, Li J, Dai L, Li J , Xu Z
Received 11 September 2018
Accepted for publication 31 January 2019
Published 12 March 2019 Volume 2019:12 Pages 1947—1956
DOI https://doi.org/10.2147/OTT.S186922
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr William C. Cho
Caijun Dong,1 Xingwang Liu,2 Huisheng Wang,3 Jutao Li,4 Lei Dai,5 Jun Li,6 Zhen Xu1
1Department of Cardiothoracic Surgery, Ningbo No. 2 Hospital, Ningbo, Zhejiang, People’s Republic of China; 2Sports Medicine Center, Department of Sports Medicine and Arthroscopy Surgery, Huashan Hospital, Fudan University, Shanghai, People’s Republic of China; 3Department of Orthopedics, The People’s Hospital of China Medical University, Shenyang, People’s Republic of China; 4Department of Hand and Foot Surgery I, Dalian Municipal Central Hospital Affiliated of Dalian Medical University, Liaoning, People’s Republic of China; 5Department of Thyroid Surgery, Ningbo No. 2 Hospital, Ningbo, Zhejiang, People’s Republic of China; 6Department of General Surgery II, Taihe Hospital, Shiyan, Hubei, People’s Republic of China
Purpose: To explore the effects of hypoxic non-small-cell lung cancer (NSCLC)-derived exosomes on NSCLC resistance to cisplatin.
Materials and methods: Exosomes were isolated by differential centrifugation and characterized by transmission electron microscope and Western blotting. Quantitative real-time PCR was used to measure miR-21 levels. MTT assays and colony formation assays were performed to investigate the effects of hypoxia-induced exosomes on cisplatin resistance.
Results: Hypoxic NSCLC cell-derived exosomes facilitate normoxic cell resistance to cisplatin. In addition, hypoxia enhanced the miR-21 expression in NSCLC cells and cell-derived exosomes. Interestingly, changes in miR-21 levels in the hypoxia-induced exosomes affected the sensitivity of recipient cells to cisplatin. Mechanically, exosomal miR-21 promoted cisplatin resistance by downregulating phosphatase and tensin homolog (PTEN). The expression of miR-21 in tumor cell lines and clinical NSCLC tumor samples was positively correlated with hypoxia-inducible factor-1α and negatively correlated with PTEN. Moreover, high miR-21 expression was associated with shorter median survival period in patients undergoing pharmacotherapy, but no association was observed in patients who were not under pharmacotherapy.
Conclusion: Exosomal miR-21 derived from hypoxic NSCLC cells may promote cisplatin resistance, which indicates that exosomal miR-21 might be a potential biomarker and therapeutic target to address NSCLC chemoresistance.
Keywords: non-small-cell lung cancer, exosomes, miR-21, cisplatin resistance, PTEN
Introduction
Lung cancer is among the leading causes of mortality worldwide, with >80% of lung cancer cases being non-small-cell lung cancer (NSCLC) type.1 Chemotherapy is the mainstay treatment for NSCLC, which prolongs and improves the patients’ quality of life. Cisplatin is a commonly prescribed drug for NSCLC, which has been found to provide many benefits, such as diminution of tumor size and improvement of prognosis.2 However, NSCLC cells often develop both inherent and acquired resistance to cisplatin, which leads to chemotherapy failure.3,4
Hypoxia is a condition characterized by insufficient oxygen, caused by higher proliferation rate of tumor cells than vasculature cells.5 As a typical characteristic of the tumor microenvironment, hypoxia has a profound influence on tumor procession.6,7 For instance, hypoxic environment causes changes in gene expression in tumor cells and affects tumor cell characteristics.8 Many studies have shown that hypoxia may promote tumor aggressiveness and metastasis.8–10
Exosomes are membrane-enclosed, nanoscale extracellular vesicles with a diameter smaller than 150 nm formed by bubbling inward from endosomes and shed through fusion with the cell membrane. They are produced by almost all types of cells.11 Functionally, exosomes participate in intercellular communication and transport of many functional proteins and RNAs. Several studies have highlighted the importance of exosomes in many physiological and pathological conditions.12–14 Hypoxia can alter the contents of tumor cell-derived exosomes,15 and increasing evidence suggests that exosomes can influence chemotherapy resistance among heterogeneous subpopulations of tumor cells in some types of tumors.16–18
MicroRNAs (miRNAs) are vital biomolecules in many cellular and intercellular processes. Many studies have shown that abnormal expression of miRNAs may reflect the pathological states of cells,14,19 such as tumor cell proliferation and chemotherapy resistance.20,21 MiR-21 is a highly conserved miRNA. Some studies reported that miR-21 may play a role in the resistance of tumors to many chemotherapeutic drugs.22–25 Moreover, miR-21 is found in tumor-derived exosomes and plays important roles in tumor progression.8,26–28 Several studies demonstrate that miR-21 is transferred by exosomes between tumor cells, thereby influencing cell activities.8,29
Currently, there are few reports pertaining to the role of hypoxia-induced exosomes on chemoresistance. In this study, we hypothesized that exosomes derived from hypoxic tumor cells can transfer miRNAs to normoxic cells, thereby modulating the functions of normoxic cells. Cobalt (II) chloride (CoCl2) was used to mimic hypoxia in vitro as previously described.30 Results show that hypoxic NSCLC cell-derived exosomes promoted cisplatin resistance in normoxic cells. Further experiments show that hypoxia increased NSCLC cell-derived exosomal miR-21. When the exosomal miR-21 was transferred into normoxic cells, it enhanced cisplatin resistance by decreasing phosphatase and tensin homolog (PTEN). This study provides new findings on the role of hypoxia-induced exosomal miR-21 in tumor chemotherapy resistance under hypoxic conditions. Since cisplatin resistance is a common response in NSCLC patients undergoing chemotherapy, this study provides an avenue that may be exploited to overcome NSCLC resistance to cisplatin.
Materials and methods
Cell culture and CoCl2 treatment
Cell line SK-MES-1 was purchased from the Cell Bank, Chinese Academy of Sciences. SK-MES-1 cell line was cultured in 1640 medium and A549 cell line was cultured in the F12K medium. Both cells were cultured in a cell incubator at 37°C with 5% CO2. In both media, 1% penicillin–streptomycin and 10% FBS were included. FBS was ultracentrifugated overnight to eliminate bovine exosomes. NSCLC cells at about 80% confluence were exposed to 200 μM CoCl2 medium for 72 hours under 37°C to mimic hypoxia. Cells incubated in a normal medium served as control. Following CoCl2 treatment, the supernatant was obtained for exosome isolation.
Exosome isolation
The exosomes in the culture media were isolated by differential centrifugation. Briefly, the supernatant was subjected to centrifugation at 2,000 g for 10 minutes to eliminate cells and centrifugation at 10,000 g for 30 minutes to remove debris. The supernatant was then ultracentrifuged at 110,000 g for 70 minutes to precipitate the exosomes. After washing the exosome pellets with PBS, they were ultracentrifuged at 110,000 g for another 70 minutes.
Transfection of oligonucleotide mimics and establishment of miR-21 knockdown (KD) cell line
To overexpress miR-21, miR-21 mimics were transfected into SK-MES-1 cells by Lipofectamine2000 (Thermo) at a final concentration of 20 nM. For miR-21 KD, oligonucleotides targeted at the miR-21 mature sequence were loaded into pSUPER-puro (Clontech), 293 T cells were used to package the virus, and stable A549 cells containing miR-21 KD were selected by puromycin.
Transmission electron microscopy
Ten microliters of exosome samples were added onto 200-mesh copper grids (Beijing Zhongjingkeyi, People’s Republic of China) for 60 seconds and then subjected to a negative staining with 2% aqueous solution of uranyl acetate for another 60 seconds at room temperature. Samples were dried by a heater for 10 minutes. Finally, the samples were imaged by an FEI Tecnai G2 spirit transmission electron microscope under a voltage of 120 kV.
Quantification of miR-21 level by quantitative real-time PCR (qRT-PCR)
The miR primer software31 was used to design the qRT-PCR primers for miR-21. U6 was used as an endogenous control. Total RNA in exosomes and cells was purified by TRIzol (Thermo Fisher Scientific, Waltham, MA, USA), and the miRNA reverse transcription was performed as described before.31 FastStart Universal SYBR Green Master ( Hoffman-La Roche Ltd., Basel, Switzerland ) was used to perform qRT-PCR and analysis was performed using the 2ΔΔCt method.
Western blot analysis
Proteins were separated by SDS-PAGE gel, and then transferred to the nitrocellulose membrane. The membrane was blocked with 5% BSA for 60 minutes and incubated with primary antibodies (1:10,000 GAPDH, Proteintech; 1:500 PTEN, Proteintech; 1:1,000 AKT, Proteintech; 1:500 hypoxia-inducible factor-1α [HIF-1α], Proteintech) in 5% BSA overnight at 4°C. This was followed by incubation with horseradish peroxidase-conjugated secondary antibody for 60 minutes at room temperature. The protein bands were visualized and analyzed.
Colony formation assay
Cells were cocultured with cell-derived exosomes and placed in a fresh 10 cm plate (and 6-well plate) containing 2 μM cisplatin for 24 hours, and then cultured in 10% FBS-DMEM without cisplatin for 2 weeks. The colonies were fixed with 4% paraformaldehyde for 5 minutes, and then stained with 0.1% crystal violet for 15 minutes. Finally, the colonies were washed three times with water. The colonies with >50 cells were counted.
Clinical data
Patients’ information were downloaded from The Cancer Genome Atlas (TCGA) lung adenocarcinoma miRNA mature strand expression RNA-seq dataset. Patients whose chemotherapeutic records could be found in the dataset were enrolled in this study, and patients whose chemotherapeutic records could not be traced were eliminated.
Statistical analysis
Data are shown as mean ± SDs. Data analysis was performed using GraphPad Prism 5.0 (GraphPad Software, La Jolla, CA, USA), and P<0.05 was considered statistically significant.
Result
Hypoxic NSCLC cell-derived exosomes reduced the sensitivity of normoxic tumor cell to cisplatin
This study used CoCl2 to mimic hypoxia in vitro. NSCLC cells treated with 200 μM CoCl2 for 48 hours showed upregulated protein level of HIF-1α compared with the control group (Figure 1A). Exosomes derived from tumor cells were initially isolated from different NSCLC cell-conditioned media. Using Western blot analysis, we confirmed the enrichment of the known exosome markers ALIX and CD6332 in the isolated NSCLC cell line-derived exosomes (Figure 1B). Transmission electron microscopy analysis was performed to examine the morphology of the purified exosomes, which showed a saucer-like shape, typical exosomes morphology, with a diameter ranging from 50 to 150 nm (Figure 1C). The role of exosomes derived from hypoxic tumor cells on the sensitivity of normoxic tumor cells to cisplatin was investigated. SK-MES-1 cells treated with hypoxic tumor cell-derived and normoxic tumor cell-derived exosomes were exposed to cisplatin (0 and 30 μM) for 24 hours, and the same treatment was performed on the A549 cells. Analysis by MTT assays indicated that treatment with hypoxic tumor cell-derived exosomes increased the normoxic tumor cell survival in response to cisplatin (Figure 1D). Normoxic tumor cells exposed to cisplatin after treatment with hypoxic tumor cell-derived exosomes formed remarkably higher number of colonies compared with those treated with normoxic tumor cell-derived exosomes (Figure 1E). These results indicate that hypoxic tumor cell-derived exosomes may reduce the sensitivity of tumor cells to cisplatin.
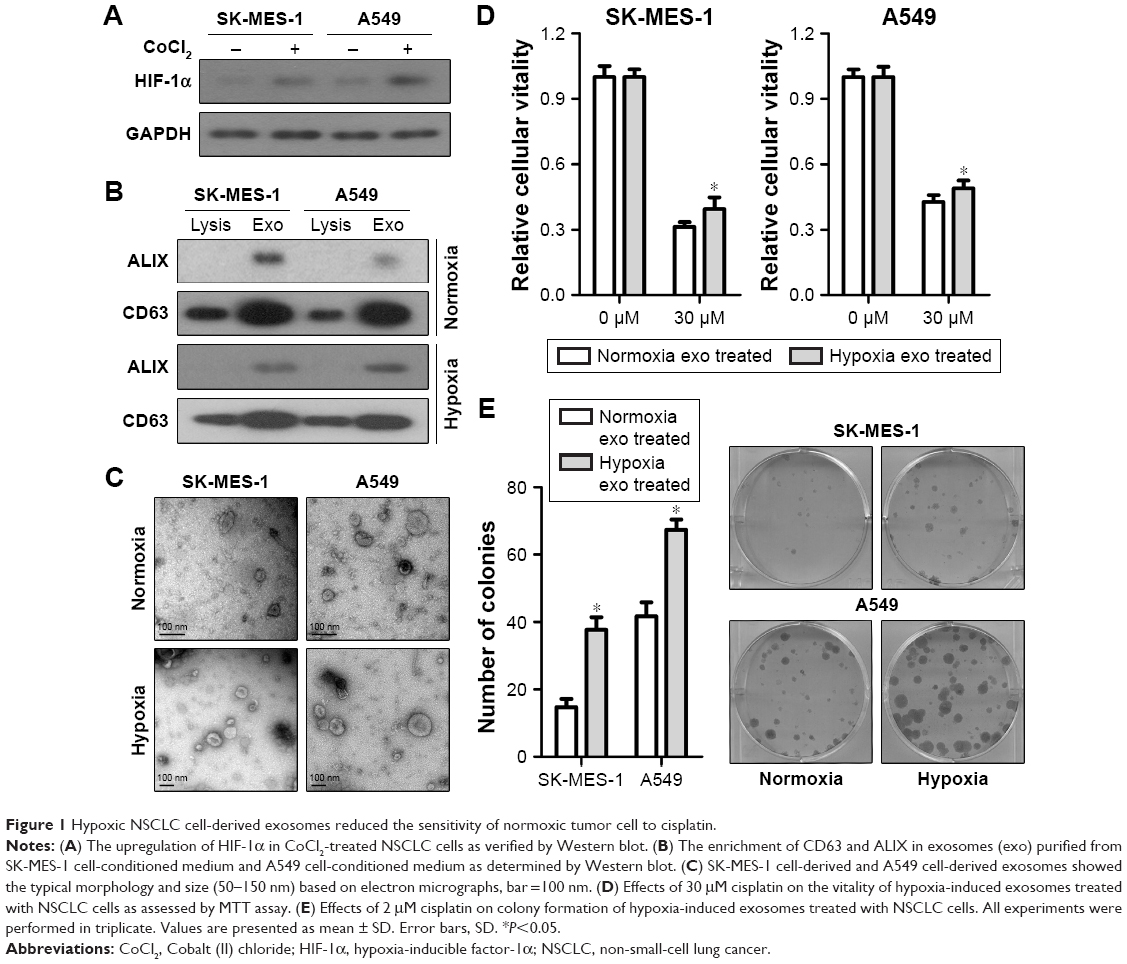 | Figure 1 Hypoxic NSCLC cell-derived exosomes reduced the sensitivity of normoxic tumor cell to cisplatin. |
Hypoxia promoted miR-21 expression in tumor cells and tumor cell-derived exosomes
Many studies have demonstrated that exosomes contain miRNAs that can be internalized by recipient cells leading to changes in cellular functions.14,33–35 For instance, miR-21, an miRNA found in exosomes, has been widely reported in a variety of tumor processes, including chemoresistance.36 HIF-1α is a hypoxia-inducible gene, and its expression has been shown to be influenced by hypoxia. Here, we found that the expression of miR-21 was positively correlated with the expression of HIF-1α in cancer cell lines (Figure 2A) and NSCLC clinical samples (Figure 2B) obtained from the common databases. Additionally, the qRT-PCR results revealed that the level of miR-21 in hypoxic tumor cells was higher than that in normoxic tumor cells (Figure 2C), as well as in exosomes (Figure 2D). A549 cells expressed more miR-21 than SK-MES-1 cells (Figure 2E). Based on these findings, we focused on miR-21. We overexpressed miR-21 in SK-MES-1 cells using miR-21 mimics and downregulated miR-21 in A549 cells using shRNA.
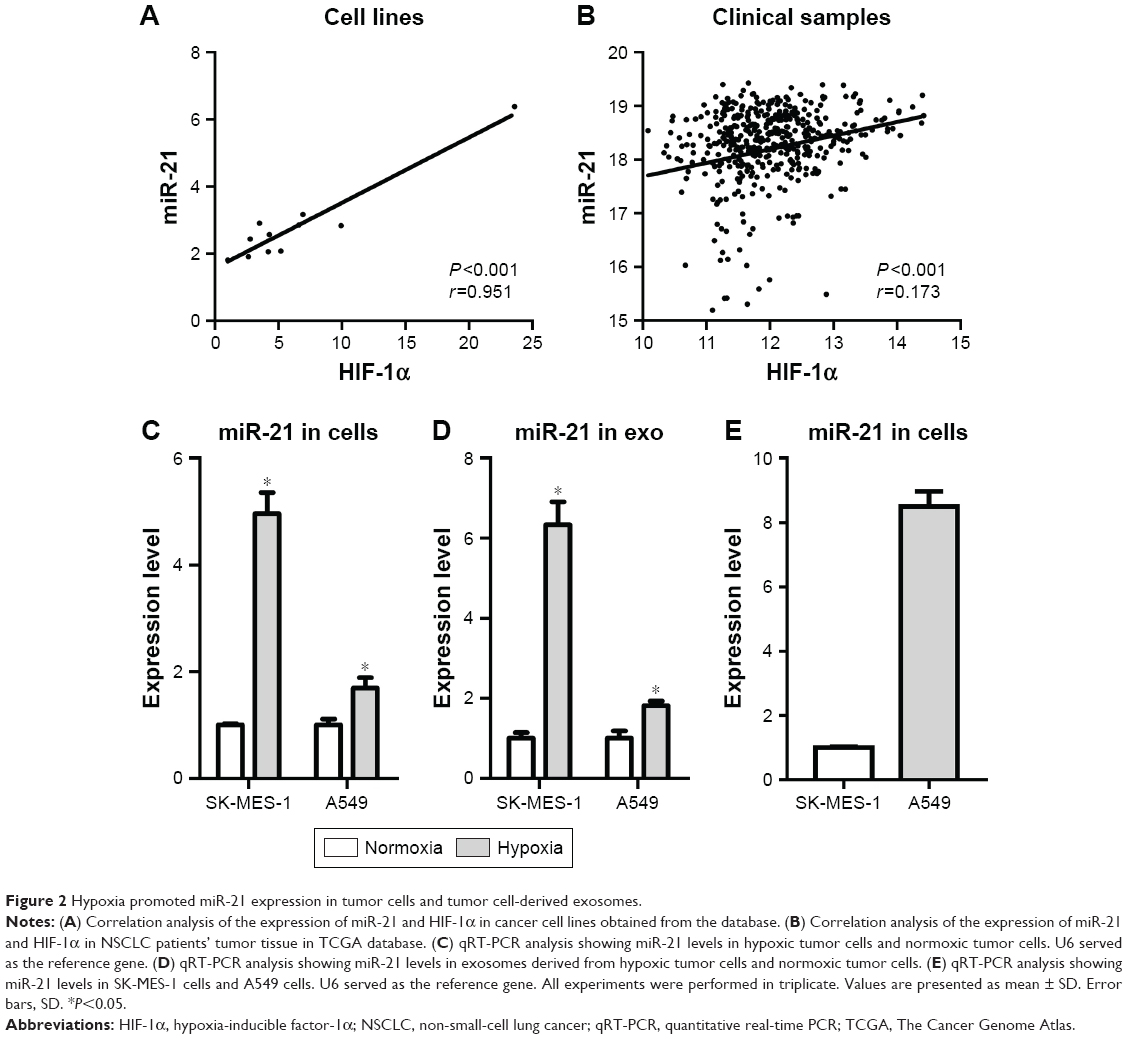 | Figure 2 Hypoxia promoted miR-21 expression in tumor cells and tumor cell-derived exosomes. |
Hypoxic tumor cell-derived exosomes transferred miR-21 to normoxic tumor cells thereby reducing sensitivity to cisplatin
To overexpress miR-21, miR-21 mimics were transfected into SK-MES-1 cells (miR-21 mimic cells). qRT-PCR analysis revealed that the transfection resulted in a 42-fold increase of miR-21 in SK-MES-1 cells (Figure 3A) and a 39-fold upregulation in exosomes (Figure 3B). Subsequently, exosomes from hypoxic miR-21 mimic cells and hypoxic control mimic cells were added into the normoxic cell culture system for 2 days. Then, we performed qRT-PCR to examine the miR-21 levels in normoxic cells. Compared with the level of miR-21 in normoxic cells treated with exosomes from hypoxic control mimic cells, there was a 15.5-fold increase in normoxic cells treated with exosomes from hypoxic miR-21 mimic cells (Figure 3C). To examine the effects of hypoxia-induced exosomal miR-21 on the sensitivity of NSCLC cells to cisplatin, normoxic cells treated with hypoxia-induced miR-21 mimic cell-derived and control mimic cell-derived exosomes were exposed to various concentrations of cisplatin (0, 10, 20, and 30 μM) for 24 hours. MTT assays indicated that hypoxic miR-21 mimic cell-derived exosomes led to an increase in cellular vitality in response to cisplatin (Figure 3D). When normoxic cells treated with hypoxic miR-21 mimic cell-derived exosomes were exposed to cisplatin, significantly higher number of colonies were formed compared with cells treated with control exosomes (Figure 3E). These findings demonstrate that hypoxic tumor cell-derived exosomal miR-21 can be internalized by normoxic tumor cells, thereby enhancing cisplatin resistance.
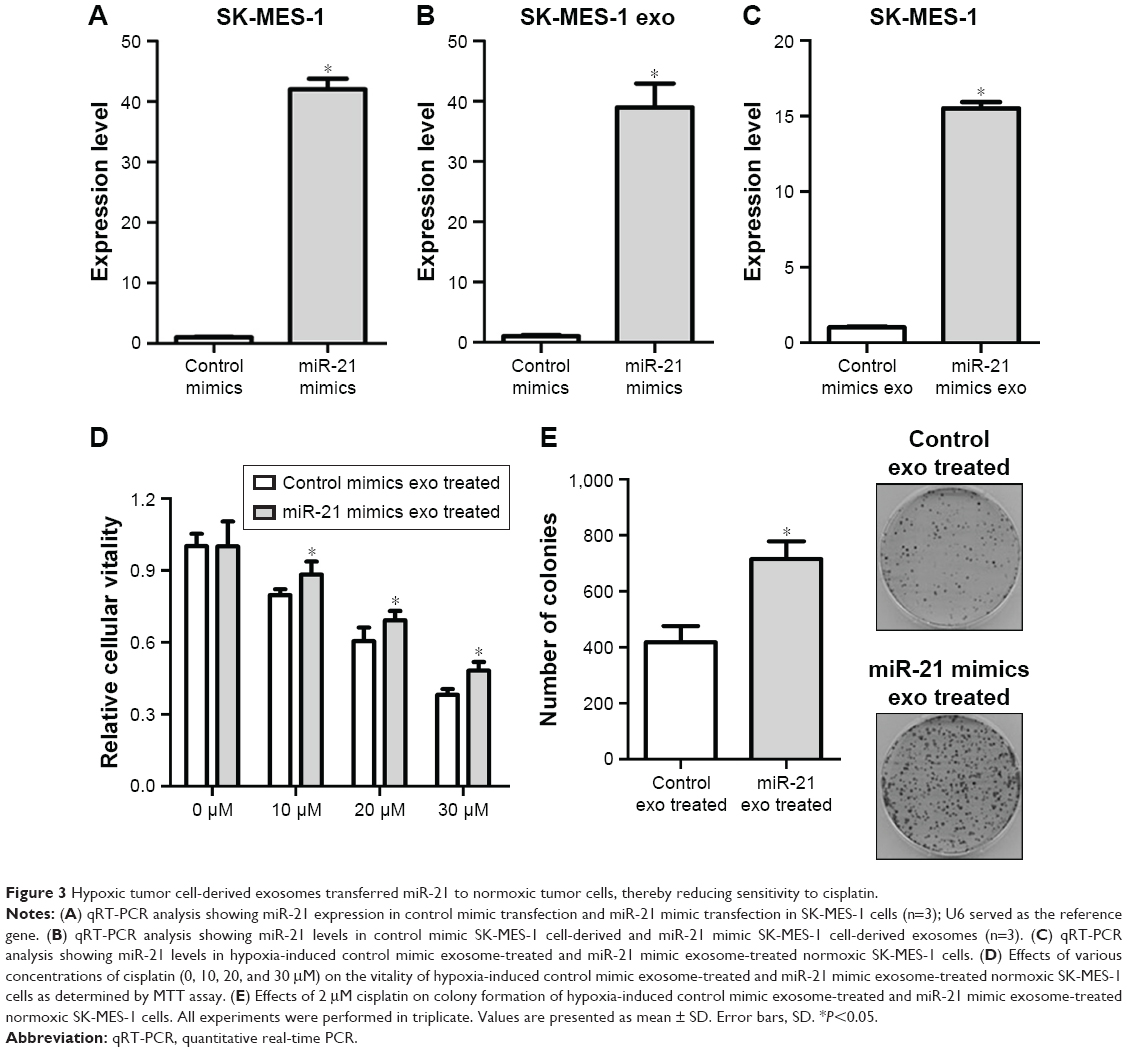 | Figure 3 Hypoxic tumor cell-derived exosomes transferred miR-21 to normoxic tumor cells, thereby reducing sensitivity to cisplatin. |
Downregulation of miR-21 in hypoxic tumor cells alleviates resistance of normoxic tumor cells to cisplatin, which is enhanced by hypoxic tumor cell-derived exosomes
ShRNA was used to knock down the miR-21 expression in A549 cells. Compared with the control cells, the expression level of miR-21 in A549 miR-21 KD cells reduced to 32% (Figure 4A), and the miR-21 level in the cell-derived exosomes was downregulated to 35% compared with the control cell-derived exosomes (Figure 4B). Moreover, the miR-21 level in normoxic cells treated with exosomes derived from hypoxic A549 miR-21 KD cells was 26% that of the normoxic cells treated with exosomes derived from hypoxic A549 control cells (Figure 4C), indicating that less miR-21 was transferred from hypoxic A549 cells into normoxic cells. We further investigated the sensitivity of normoxic cells treated with hypoxic A549 miR-21 KD cell-derived exosomes to cisplatin. The MTT assays indicated that downregulation of hypoxic A549 cell-derived exosomal miR-21 led to a decrease in normoxic cell vitality in response to cisplatin (Figure 4D). Treatment of normoxic cells exposed to cisplatin with hypoxic A549 miR-21 KD cell-derived exosomes produced a significantly lower number of colonies than normoxic cells treated with hypoxic A549 control cell-derived exosomes (Figure 4E). These findings illustrate that suppression of hypoxia-induced exosomal miR-21 may alleviate the resistance of normoxic tumor cells to cisplatin induced by hypoxic tumor cell-derived exosomes.
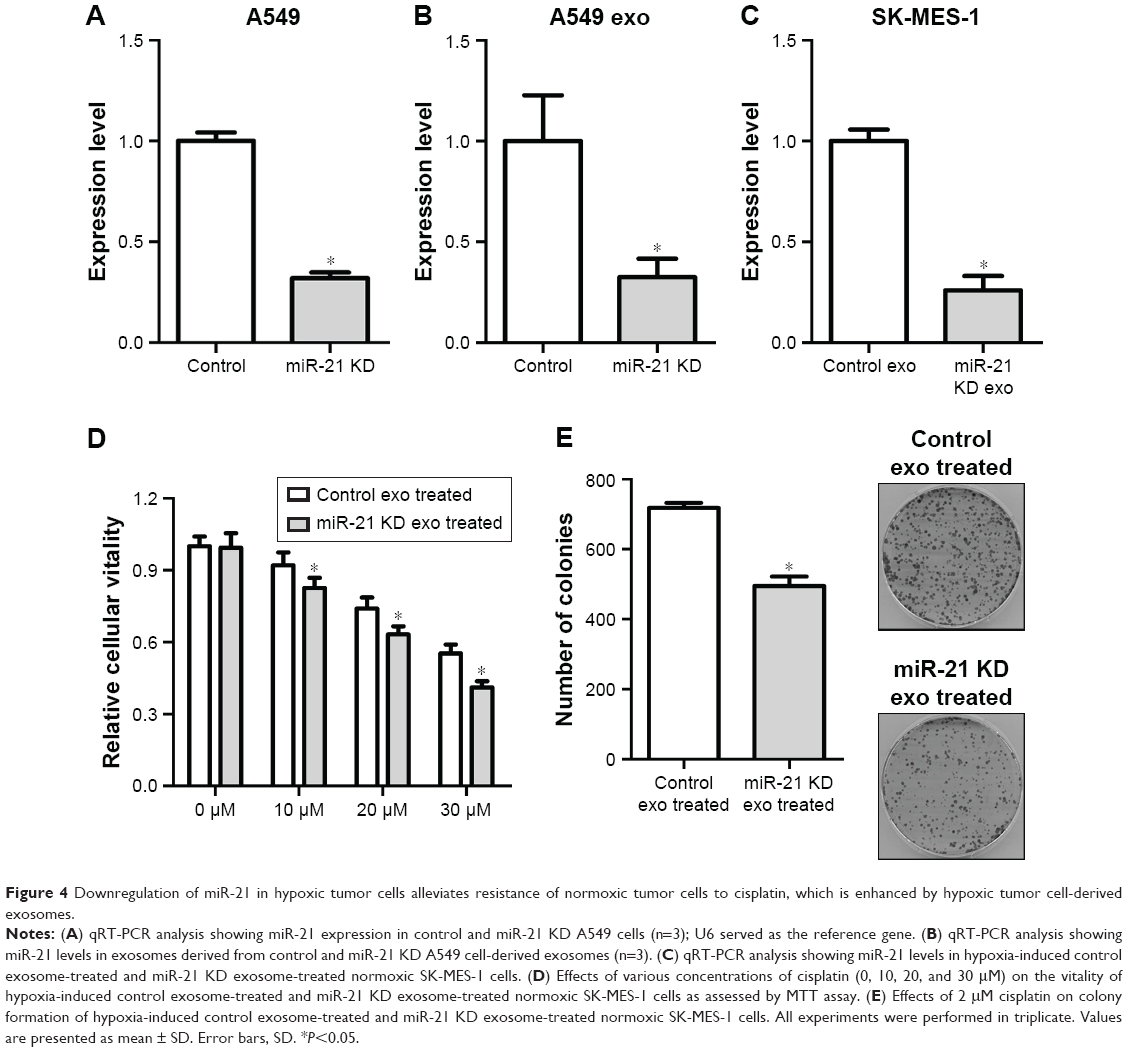 | Figure 4 Downregulation of miR-21 in hypoxic tumor cells alleviates resistance of normoxic tumor cells to cisplatin, which is enhanced by hypoxic tumor cell-derived exosomes. |
Hypoxia-induced exosomal miR-21 enhanced normoxic tumor cell resistance to cisplatin by inhibiting PTEN
Next, we explored the target gene of hypoxia-induced exosomal miR-21 that influenced the resistance of NSCLC cells to cisplatin. PTEN, tumor suppressor gene, was found to be involved in miR-21-induced resistance to anticancer drugs in human pancreatic cancers.24 Thus, we investigated whether PTEN was a target gene of hypoxia-induced exosomal miR-21 in NSCLC cells. Therefore, normoxic SK-MES-1 cells were cocultured with hypoxic miR-21 modified cell-derived exosomes and hypoxic control cell-derived exosomes for 48 hours. Compared with the groups treated with control exosomes, the expression levels of PTEN in SK-MES-1 cells were obviously downregulated in the group that was treated with exosomes derived from hypoxic miR-21 mimic cells and upregulated in the group that was treated with exosomes derived from hypoxic miR-21 KD cells (Figure 5A). Moreover, the expression level of PTEN was negatively correlated with miR-21 in NSCLC clinical samples obtained from TCGA database (Figure 5B). It has been shown that PTEN downregulation enhances the chemoresistance of tumor cells mainly through activating PI3K-AKT pathway.25,37,38 On this basis, we knock down AKT by siRNA in SK-MES-1 cells (Figure 5C). The results showed that KD of AKT restored the sensitivity of SK-MES-1 cells to cisplatin in the presence of hypoxia-induced exosomal miR-21 (Figure 5D). Collectively, these preliminary data suggested that hypoxia-induced exosomal miR-21 enhanced cisplatin resistance in normoxic tumor cells probably through the PTEN/PI3K/AKT signaling pathway.
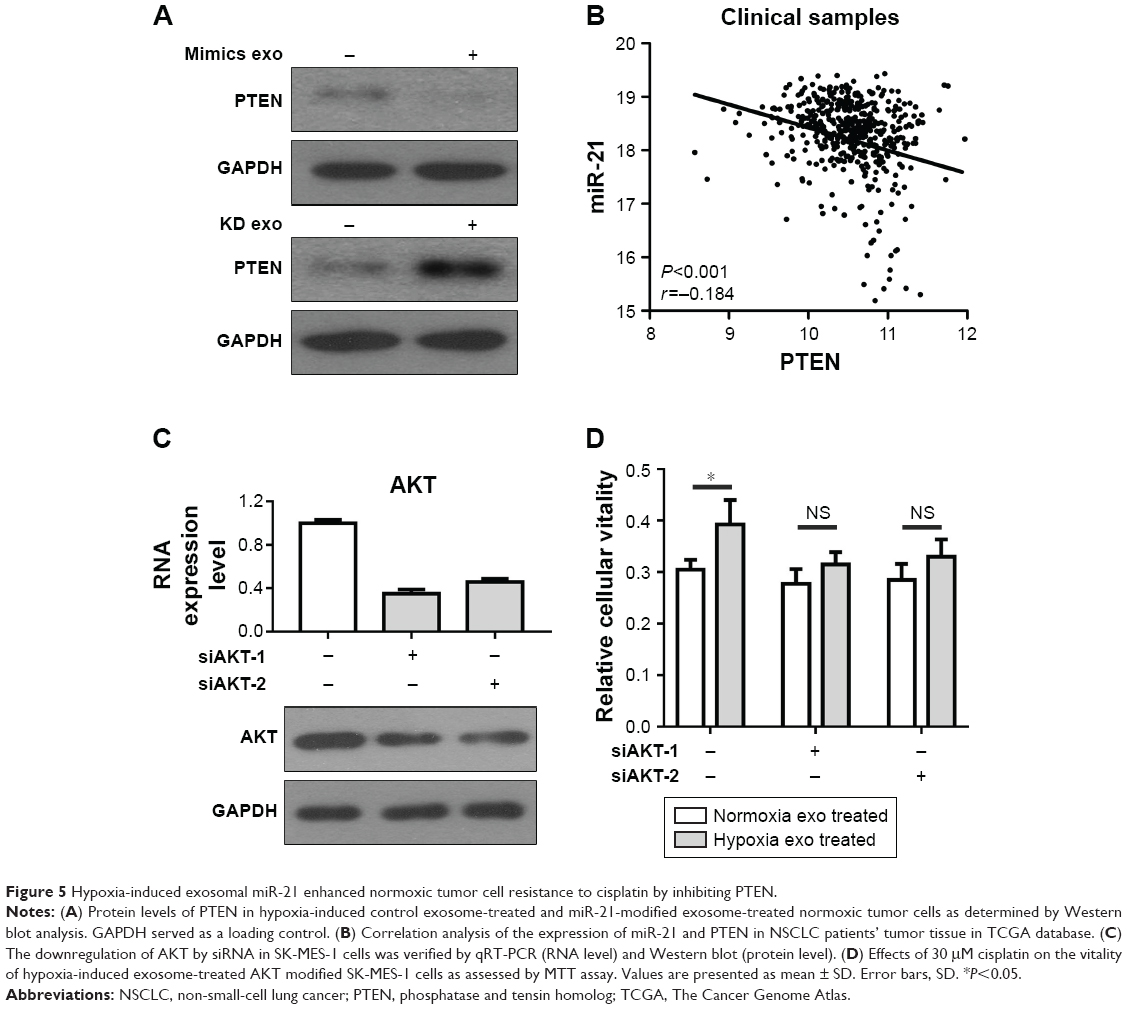 | Figure 5 Hypoxia-induced exosomal miR-21 enhanced normoxic tumor cell resistance to cisplatin by inhibiting PTEN. |
Lung cancer patients with high miR-21 expression level may have a poor prognosis while under pharmacotherapy
Finally, we investigated the clinical value of miR-21 in NSCLC pharmacotherapy. We analyzed the NSCLC dataset in TCGA. From the clinical samples obtained, we found that high miR-21 expression was associated with shorter median survival period in patients undergoing pharmacotherapy (Figure 6A), but there is no such association in patients not under pharmacotherapy (Figure 6B), which is consistent with the findings of a previous study where plasma miR-21 was correlated with the pharmacotherapy outcomes in NSCLC.39 These results indicate that the exosomal miR-21 may affect the outcome of patients under pharmacotherapy.
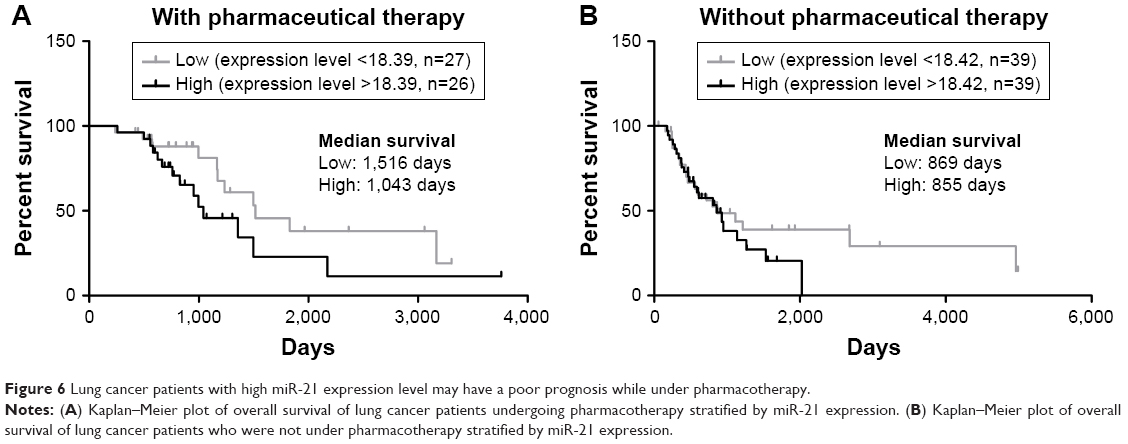 | Figure 6 Lung cancer patients with high miR-21 expression level may have a poor prognosis while under pharmacotherapy. |
Discussion
In this study, we found that hypoxic NSCLC cell-derived exosomes enhanced the resistance of normoxic NSCLC cells to cisplatin. Further investigation showed that miR-21 in exosomes derived from hypoxic NSCLC cells could be internalized by normoxic NSCLC cells to facilitate cisplatin resistance by suppressing PTEN. Clinical data also showed that hypoxia and high miR-21 expression predict poor prognosis in NSCLC patients.
We first explored the effect of exosomes derived from hypoxic NSCLC cells on the sensitivity of normoxic NSCLC cells to cisplatin and found that these exosomes promoted the resistance of normoxic NSCLC cells to cisplatin. The interactions between tumor cells, the microenvironment, and exosomes are important determinants of cancer chemotherapy resistance and are associated with therapy failure.40 Hypoxia can remodel tumor microenvironments and affect tumor cell aggressiveness and metastasis.6,7,9 However, the effect of hypoxic cell-derived exosomes on chemoresistance has not been studied previously. Thus, these findings enrich our understanding on the role of hypoxia in cancer.
Hypoxia is a powerful driver of tumor progression.7,41 HIF-1α is a well-studied hypoxia-inducible gene. We found that the expression of HIF-1α and miR-21 in tumor cell lines and NSCLC clinical samples was positively correlated. Additionally, analysis of clinical data indicates that high miR-21 expression in clinical samples predicts poor prognosis in patients undergoing chemotherapy. This result is consistent with a previous study where plasma miR-21 was used as a biomarker for pharmacotherapy of NSCLC.39 Furthermore, these findings suggest that miR-21 is a hypoxia-inducible gene, and hypoxia-induced exosomal miR-21 may act as a liquid biopsy biomarker for chemoresistance.
The expression level of miR-21 is associated with chemoresistance in lung cancer patients.39 Some researchers have demonstrated that exosomal miR-21 can regulate cancer cell characteristics.8,26–28 Here, we demonstrated that hypoxia induced miR-21 expression in NSCLC cells and their exosomes. The hypoxia-induced exosomal miR-21 triggered resistance of normoxic lung cancer cells to cisplatin. This effect was abolished by miR-21 KD in hypoxic tumor cells. These findings imply that blocking the hypoxia-induced exosomal miR-21 internalization process and decreasing miR-21 in the hypoxic tumor cell and tumor cell-derived exosomes are potential therapeutic maneuvers to reduce chemoresistance in lung cancer.
We also demonstrated that exosomal miR-21 suppressed PTEN, a well-known target gene of miR-21, in recipient cells. PTEN acts via PTEN/PI3K/AKT signaling pathway to ameliorate tumor chemoresistance.25 When AKT was knocked down by siRNAs, the sensitivity of NSCLC cells to cisplatin in the presence of hypoxia-induced exosomal miR-21 was restored. This result is consistent with results from previous reports.42,43 Several types of inhibitors for PI3K and AKT exist; these inhibitors are potential candidates for combination therapy with cisplatin. However, this concept needs to be verified through further investigation.
Limitations
This study also has some limitations worth mentioning. Hypoxia can cause diverse remodeling of the tumor gene expression and the exosomal contents. In addition, there could be other exosomal molecules that may influence the chemoresistance of the normoxic cell besides miR-21. Thus, further studies should explore other hypoxia-induced exosomal molecules involved in chemoresistance. Since one miRNA may regulate multiple genes, other miR-21 target genes, such as PDCD4, involved in tumor cell chemoresistance need to be investigated. Moreover, in vitro experiments are simplified simulations of the in vivo conditions; investigating the effects of hypoxic tumor-derived exosomal miR-21 on tumor chemoresistance in vivo should be carried out in future.
Conclusion
Hypoxia triggered the expression of miR-21 in NSCLC cells and cell-derived exosomes, and exosomal miR-21 was able to induce cisplatin resistance, a characteristic of tumor cells, in normoxic tumor cells. KD of miR-21 in hypoxic tumor cells abolished this effect. This was attributed to the ability of miR-21 to decrease PTEN in recipient cells. Additionally, the expression of miR-21 in tumor cell lines and NSCLC clinical tumor samples was positively correlated with HIF-1α and negatively correlated with PTEN levels. High miR-21 expression was associated with shorter median survival period in patients undergoing pharmacotherapy. These findings provide new understanding on the roles of hypoxia-induced exosomes in tumor chemotherapy resistance. Since cisplatin resistance is a common response in NSCLC patients undergoing chemotherapy, this study shows that exosomal miR-21 can be targeted as a therapeutic strategy to overcome NSCLC chemoresistance.
Acknowledgment
This work was supported by the Postdoctoral Foundation of China (2017M621366).
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel R, Ward E, Brawley O, Jemal A. Cancer statistics, 2011: the impact of eliminating socioeconomic and racial disparities on premature cancer deaths. CA Cancer J Clin. 2011;61(4):212–236. | ||
Todd RC, Lippard SJ. Inhibition of transcription by platinum antitumor compounds. Metallomics. 2009;1(4):280–291. | ||
Kostova I. Platinum complexes as anticancer agents. Recent Pat Anticancer Drug Discov. 2006;1(1):1–22. | ||
Wang D, Lippard SJ. Cellular processing of platinum anticancer drugs. Nat Rev Drug Discov. 2005;4(4):307–320. | ||
Mckeown SR. Defining normoxia, physoxia and hypoxia in tumours – implications for treatment response. Br J Radiol. 2014;87(1035):20130676. | ||
Gillies RJ, Verduzco D, Gatenby RA. Evolutionary dynamics of carcinogenesis and why targeted therapy does not work. Nat Rev Cancer. 2012;12(7):487–493. | ||
Harris AL. Hypoxia – a key regulatory factor in tumour growth. Nat Rev Cancer. 2002;2(1):38–47. | ||
Li L, Li C, Wang S, et al. Exosomes derived from hypoxic oral squamous cell carcinoma cells deliver miR-21 to normoxic cells to elicit a prometastatic phenotype. Cancer Res. 2016;76(7):1770–1780. | ||
Brahimi-Horn MC, Chiche J, Pouysségur J. Hypoxia and cancer. J Mol Med (Berl). 2007;85(12):1301–1307. | ||
Semenza GL. The hypoxic tumor microenvironment: a driving force for breast cancer progression. Biochim Biophys Acta. 2016;1863(3):382–391. | ||
Tkach M, Théry C. Communication by extracellular vesicles: where we are and where we need to go. Cell. 2016;164(6):1226–1232. | ||
Mathivanan S, Ji H, Simpson RJ. Exosomes: extracellular organelles important in intercellular communication. J Proteomics. 2010;73(10):1907–1920. | ||
Skog J, Würdinger T, van Rijn S, et al. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat Cell Biol. 2008;10(12):1470–1476. | ||
Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–659. | ||
Shao C, Yang F, Miao S, et al. Role of hypoxia-induced exosomes in tumor biology. Mol Canc. 2018;17(1):120. | ||
Chen WX, Liu XM, Lv MM, et al. Exosomes from drug-resistant breast cancer cells transmit chemoresistance by a horizontal transfer of microRNAs. PLoS One. 2014;9(4):e95240. | ||
Corcoran C, Rani S, O’Brien K, et al. Docetaxel-resistance in prostate cancer: evaluating associated phenotypic changes and potential for resistance transfer via exosomes. PLoS One. 2012;7(12):e50999. | ||
Safaei R, Larson BJ, Cheng TC, et al. Abnormal lysosomal trafficking and enhanced exosomal export of cisplatin in drug-resistant human ovarian carcinoma cells. Mol Cancer Ther. 2005;4(10):1595–1604. | ||
Kloosterman WP, Plasterk RH. The diverse functions of microRNAs in animal development and disease. Dev Cell. 2006;11(4):441–450. | ||
Katz B, Tropé CG, Reich R, Davidson B. MicroRNAs in ovarian cancer. Hum Pathol. 2015;46(9):1245–1256. | ||
Nassar FJ, Nasr R, Talhouk R. MicroRNAs as biomarkers for early breast cancer diagnosis, prognosis and therapy prediction. Pharmacol Ther. 2017;172:34–49. | ||
Au Yeung CL, Co NN, Tsuruga T, et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat Commun. 2016;7(1):11150. | ||
Feng YH, Tsao CJ. Emerging role of microRNA-21 in cancer. Biomed Rep. 2016;5(4):395–402. | ||
Wei X, Wang W, Wang L, et al. MicroRNA-21 induces 5-fluorouracil resistance in human pancreatic cancer cells by regulating PTEN and PDCD4. Canc Med. 2016;5(4):693–702. | ||
Wu YR, Qi HJ, Deng DF, Luo YY, Yang SL. MicroRNA-21 promotes cell proliferation, migration, and resistance to apoptosis through PTEN/PI3K/AKT signaling pathway in esophageal cancer. Tumour Biol. 2016;37(9):12061–12070. | ||
Sun X, Ma X, Wang J, et al. Glioma stem cells-derived exosomes promote the angiogenic ability of endothelial cells through miR-21/VEGF signal. Oncotarget. 2017;8(22):36137–36148. | ||
Xu Z, Liu X, Wang H, et al. Lung adenocarcinoma cell-derived exosomal miR-21 facilitates osteoclastogenesis. Gene. 2018;666:116–122. | ||
Zheng P, Chen L, Yuan X, et al. Exosomal transfer of tumor-associated macrophage-derived miR-21 confers cisplatin resistance in gastric cancer cells. J Exp Clin Cancer Res. 2017;36(1):53. | ||
Liu T, Chen G, Sun D, et al. Exosomes containing miR-21 transfer the characteristic of cisplatin resistance by targeting PTEN and PDCD4 in oral squamous cell carcinoma. Acta Biochim Biophys Sin (Shanghai). 2017;49(9):808–816. | ||
Xu X, Jiao X, Song N, et al. Role of miR-21 on vascular endothelial cells in the protective effect of renal delayed ischemic preconditioning. Mol Med Rep. 2017;16(3):2627–2635. | ||
Busk PK. A tool for design of primers for microRNA-specific quantitative RT-qPCR. BMC Bioinformatics. 2014;15(1):29. | ||
Thery C, Amigorena S, Raposo G. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol. 2006;30(1):3.22.1–3.22.3. | ||
Monteleone F, Taverna S, Alessandro R, Fontana S. SWATH-MS based quantitative proteomics analysis reveals that curcumin alters the metabolic enzyme profile of CML cells by affecting the activity of miR-22/IPO7/HIF-1α axis. J Exp Clin Cancer Res. 2018;37(1):170. | ||
Ostenfeld MS, Jeppesen DK, Laurberg JR, et al. Cellular disposal of miR23b by RAB27-dependent exosome release is linked to acquisition of metastatic properties. Canc Res. 2014;74(20):5758–5771. | ||
Taverna S, Giallombardo M, Pucci M, et al. Curcumin inhibits in vitro and in vivo chronic myelogenous leukemia cells growth: a possible role for exosomal disposal of miR-21. Oncotarget. 2015;6(26):21918–21933. | ||
Xu L, Huang Y, Chen D, et al. Downregulation of miR-21 increases cisplatin sensitivity of non–small-cell lung cancer. Canc Genet. 2014;207(5):214–220. | ||
Chen Q, Qin R, Fang Y, Li H. Berberine sensitizes human ovarian cancer cells to cisplatin through miR-93/PTEN/Akt signaling pathway. Cell Physiol Biochem. 2015;36(3):956–965. | ||
Zhu Y, Liu Y, Zhang C, et al. Tamoxifen-resistant breast cancer cells are resistant to DNA-damaging chemotherapy because of upregulated BARD1 and BRCA1. Nat Commun. 2018;9(1):1595. | ||
Wei J, Gao W, Zhu CJ, et al. Identification of plasma microRNA-21 as a biomarker for early detection and chemosensitivity of non-small cell lung cancer. Chin J Cancer. 2011;30(6):407–414. | ||
Yin Y, Yao S, Hu Y, et al. The immune-microenvironment confers chemoresistance of colorectal cancer through macrophage-derived IL6. Clin Cancer Res. 2017;23(23):7375–7387. | ||
Lu X, Kang Y. Hypoxia and hypoxia-inducible factors: master regulators of metastasis. Clin Canc Res. 2010;16(24):5928–5935. | ||
Holland WS, Chinn DC, Lara PN, Gandara DR, Mack PC. Effects of Akt inhibition on HGF-mediated erlotinib resistance in non-small cell lung cancer cell lines. J Cancer Res Clin Oncol. 2015;141(4):615–626. | ||
Walsh S, Gill C, O’Neill A, Fitzpatrick JM, Watson RW. Hypoxia increases normal prostate epithelial cell resistance to receptor-mediated apoptosis via AKT activation. Int J Cancer. 2009;124(8):1871–1878. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.