")
Back to Journals » OncoTargets and Therapy » Volume 10
Hypermethylation of secreted frizzled-related proteins predicts poor prognosis in non-M3 acute myeloid leukemia
Authors Guo H, Zhang T, Wen X, Zhou JD, Ma J, An C, Zhang W, Xu Z, Lin J, Qian J
Received 8 March 2017
Accepted for publication 31 May 2017
Published 20 July 2017 Volume 2017:10 Pages 3635—3644
DOI https://doi.org/10.2147/OTT.S136502
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr William C. Cho
Hong Guo,1,2,* Ting-juan Zhang,2,3,* Xiang-mei Wen,1,2 Jing-dong Zhou,2,3 Ji-chun Ma,1,2 Cui An,4 Wei Zhang,2,3 Zi-jun Xu,1,2 Jiang Lin,1,2 Jun Qian2,3
1Laboratory Center, Affiliated People’s Hospital of Jiangsu University, 2The Key Lab of Precision Diagnosis and Treatment of Zhenjiang City, 3Department of Hematology, Affiliated People’s Hospital of Jiangsu University, Zhenjiang, 4Department of Hematology, People’s Hospital of Yu City, Dezhou, People’s Republic of China
*These authors contributed equally to this work
Objective: Secreted frizzled-related proteins (SFRPs) as Wnt signaling antagonists have been found to be dysregulated by promoter hypermethylation in several cancers including acute myeloid leukemia (AML). This study aimed to investigate the methylated status of SFRPs promoter region and its clinical relevance in Chinese non-M3 AML patients.
Methods: SFRPs methylation in 139 primary non-M3 AML patients was determined using methylation-specific real-time quantitative polymerase chain reaction.
Results: The frequency of aberrant methylation was as follows: 30.2% for SFRP1, 27.3% for SFRP2, 5.0% for SFRP4, and 1.4% for SFRP5. Hypermethylation of at least one SFRP gene occurred in 51.8% (72/139) of non-M3 AML patient samples, which was significantly higher compared to normal control (0/21) (P<0.001). Hypermethylation of SFRP1 was potentially associated with N/K-RAS mutations (P=0.043), and the frequency of SFRPs methylation was higher in patients ≥50 years compared to those <50 years, especially for SFRP2 (P<0.05). Furthermore, both whole cohort and cytogenetically normal (CN) patients with high SFRPs-methylated group showed a shorter overall survival (OS) compared to those with low group (P=0.036 and P=0.035, respectively). Moreover, Cox regression multivariate analysis revealed that SFRPs hypermethylation acts as an independent prognostic biomarker among both whole cohort (hazard ratio =1.804, P=0.026) and CN (hazard ratio =2.477, P=0.023) patients. In leukemic cell line HL60 treated with 5-aza-2'-deoxycytidine, the alteration of SFRP1/2 expression inversely correlated with change in SFRP1/2 methylation (r=-0.975, P=0.005 and r=-0.975, P=0.005, respectively). A tendency of negative correlation was observed between SFRP1 expression and its promoter methylation in AML patients (r=-0.334, P=0.038).
Conclusion: These findings suggested that hypermethylation of SFRP1/2 was a frequent event and silenced SFRP1/2 expression in AML. Moreover, hypermethylation of SFRPs promoter was an adverse risk factor for OS in Chinese non-M3 AML patients.
Keywords: SFRPs, methylation, RQ-MSP, non-M3 AML, prognosis
Introduction
Acute myeloid leukemia (AML), the most common type of leukemia in adults, is a group of heterogeneous malignant disorders characterized by clonal proliferation and differentiation arrest of myeloid progenitor cells.1,2 A multistep process involving genetic and epigenetic aberrations induces normal hematopoietic stem cells (HSCs) to lose the ability of differentiation to various mature cell lineages, to transform into leukemic stem cells, and to accumulate in the bone marrow (BM).3,4 Genetic aberrations, involving chromosomal abnormalities (translocation, addition, and deletion) and gene alterations (mutation, deletion, amplification, and translocation), have been implicated in the pathogenesis of leukemia and identified in the special subtypes of AML.5–7 During last decades, epigenetic disturbances, such as aberrant DNA promoter methylation and covalent histone modification, have also been demonstrated to play crucial roles in the process of leukemogenesis.8–11
In the past few years, numerous studies have indicated that the pathogenesis of AML involves the abnormal activation of Wnt signaling pathway, which has a key role in extensive cellular processes of hematopoietic progenitor proliferation and differentiation.12–14 Wnt signaling pathway comprises the canonical pathway with β−catenin and the non-canonical pathways involving calcium ions and planar cell polarity.15–17 Growing evidence suggests that the Wnt signaling pathway plays an important role in normal HSCs maintenance, whereas nuclear β-catenin is a key molecule that mediates transcriptional activation of specific gene expression and is thought to have the most significance in cancer development. Moreover, a number of studies have identified that dysregulation of this pathway by promoter hypermethylation leads to leukemia.18–22 Negative regulators of the Wnt signaling pathway, including the WIF1, the four mammalian homologues of Dickkopf (DKK), and the secreted frizzled-related proteins (SFRPs), are epigenetically silenced in tumor cells.23–26
SFRPs, the largest family among Wnt antagonists, suppress the Wnt signaling pathway by interacting with the frizzled receptor. Four of the five known SFRP genes (SFRP1/2/4/5) have been found to be epigenetically silenced in cancer cells.27 Hypermethylation of the above-mentioned four SFRP genes, which were associated with aberrant Wnt signaling activation, has been shown in AML.28–30 Furthermore, several studies have suggested that hypermethylation of SFRPs promoter was an adverse risk factor for survival in patients with AML;28,30,31 however, few studies have been conducted in the Chinese population.
Therefore, our study aims to investigate the methylated status of SFRPs promoter and its clinical relevance by real-time quantitative methylation-specific polymerase chain reaction (RQ-MSP) in the Chinese non-M3 AML patients.
Materials and methods
Patients and samples
This study was approved by the Ethics Committee of Affiliated People’s Hospital of Jiangsu University. After obtaining informed consent, BM specimens were collected at the time of diagnosis during routine clinical assessment of 139 patients with non-M3 AML. All patients were treated at the Affiliated People’s Hospital of Jiangsu University between 2005 and 2015. The BM specimens from 21 healthy volunteers served as controls. The diagnosis and classification of de novo AML patients were established according to the French–American–British (FAB) and World Health Organization (WHO) criteria.32,33 Karyotypes were analyzed by the conventional R-banding method. Karyotype risk classification was identified as reported previously.34 The main clinical and laboratory features of the patients are summarized in Table 1. The BM mononuclear cells (BMMNCs) were separated by Ficoll–Hypaque gradient.
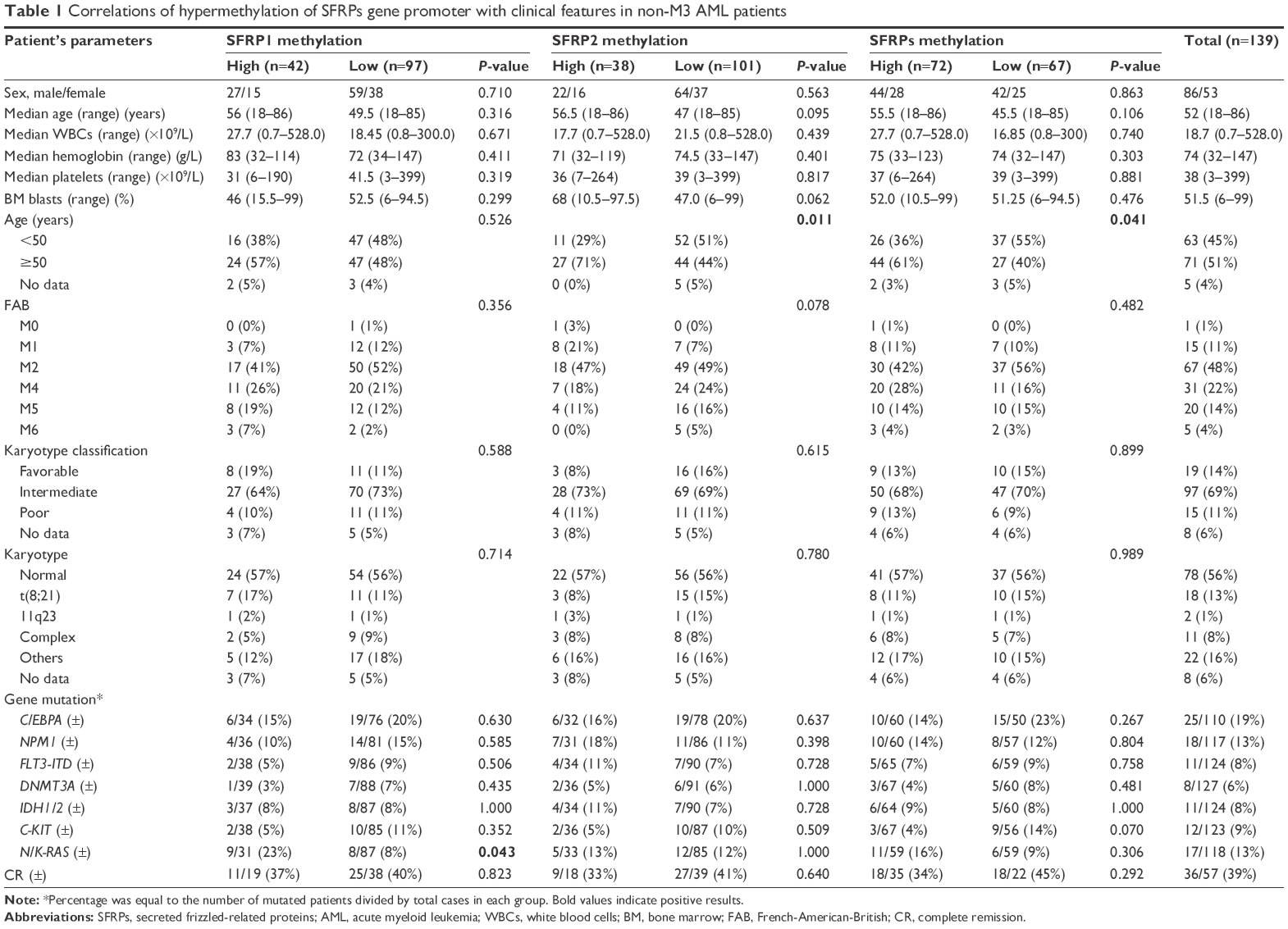 | Table 1 Correlations of hypermethylation of SFRPs gene promoter with clinical features in non-M3 AML patients |
Cell culture and 5-aza-dC treatment
Seven human AML cell lines (SHI-1, THP-1, U937, HEL, HL60, K562, and NB4) (ATCC, Manassas, VA, USA) were routinely cultured in IMDM with 10% fetal bovine serum (ExCell Bio, Shanghai, People’s Republic of China) and grown at 37°C in 5% CO2 humidified atmosphere. Moreover, HL60 cells were incubated at a final concentration of 0 μM, 0.1 μM, 1 μM, 10 μM, and 50 μM 5-aza-2′−deoxycytidine (5-aza-dC) (Sigma-Aldrich, St Louis, MO, USA) for 72 h. All cells were cultured until harvested for extracting RNA and DNA.
RNA isolation, reverse transcription, and real-time quantitative PCR
Total RNA was isolated from BMMNCs and AML cell lines using the Trizol reagent (Thermo Fisher Scientific, Waltham, MA, USA) and reverse-transcribed into cDNA.35 Primer sequences for SFRPs are listed in Table 2. Real-time quantitative PCR (RQ-PCR) was carried out for each sample in a final reaction volume of 20 μL, consisting of 0.4 μM of primers, 10 μL of SYBR Premix Ex Taq II, 0.4 μL of 50× ROX (TaKaRa, Tokyo, Japan), and 50 ng of cDNA. RQ-PCR was performed on StepOnePlus (Applied Biosystems, Foster City, CA, USA). PCR program was carried out at 95°C for 30 s, followed by 45 cycles at 95°C for 5 s, at T1 (in Table 2) for 30 s and 72°C for 30 s, and a fluorescence collection step at T2 (in Table 2) for 30 s, then followed by a melting program at 95°C for 15 s, 60°C for 60 s, and 95°C for 15 s. The mRNA abundance of SFRPs gene was calculated relative to the expression of the housekeeping gene (ABL).36
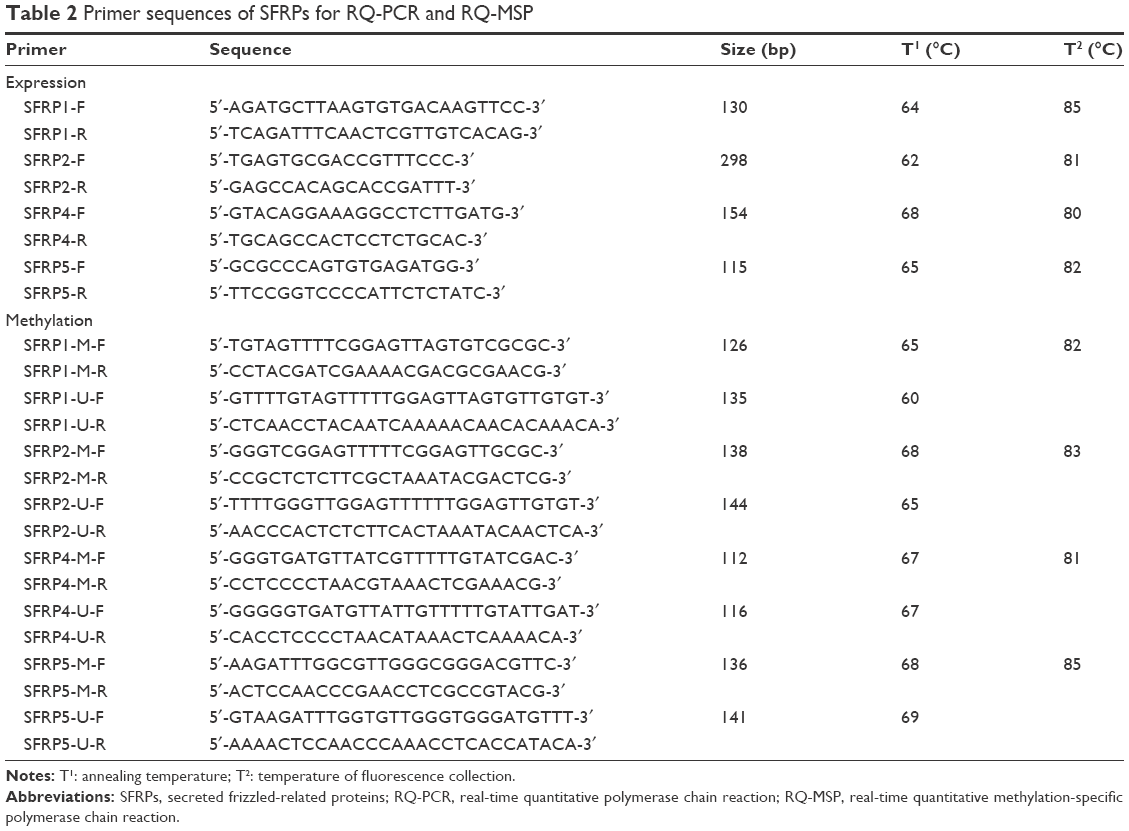 | Table 2 Primer sequences of SFRPs for RQ-PCR and RQ-MSP |
DNA isolation, bisulfite modification, and RQ-MSP
Genomic DNA was isolated using genomic DNA purification kit (Gentra, Minneapolis, MN, USA). According to the manufacturer’s instructions, 1 μg of genomic DNA was modified using the CpGenome DNA Modification Kit (Chemicon, Ternecula, Canada). The primers (Table 2) were designed using the Methylation Primer Express Software v1.0 (Applied Biosystems). RQ-MSP was carried out for each sample in a final reaction volume of 20 μL, containing 0.4 μM of primers, 10 μL of SYBR Premix Ex Taq II, 0.4 μL of 50× ROX (TaKaRa), and 50 ng of bisulfite-treated DNA. RQ-MSP was performed on StepOnePlus (Applied Biosystems). Amplification was carried out at 95°C for 30 s, followed by 40 cycles at 95°C for 5 s at T1 (in Table 2) for 30 s and 72°C for 30 s, and a fluorescence collection step at T2 (in Table 2) for 30 s to collect fluorescence data. The relative level of methylated SFRPs was calculated in relation to the reference ALU sequence using the formula: NM-SFRPs =(EM-SFRPs)ΔCT M-SFRPs (control-sample) ÷ (EALU)ΔCT ALU (control-sample).37
Gene mutation detection
NPM1, C-KIT, DNMT3A, IDH1/2, and N/K-RAS mutations were detected by high-resolution melting analysis (HRMA) as reported previously.38–40 To confirm the results of HRMA, all positive samples were detected by direct DNA sequencing, and C/EBPA and FLT3-ITD mutations were directly DNA sequenced.41,42
Statistical methods
All statistical analyses were performed using the SPSS 17.0 software package (SPSS, Chicago, IL, USA). Fisher’s exact test or Pearson’s Chi-square analysis was carried out to compare the difference of categorical variables between the patients groups, whereas Mann–Whitney U test was carried out to compare the difference of continuous variables between the two groups. Overall survival (OS) curve was plotted according to the Kaplan–Meier method and evaluated using the log-rank test. Correlation between SFRPs methylation and expression was compared by Spearman’s correlation test. A P-value <0.05 (two-tailed) was considered to be statistically significant.
Results
Methylation of SFRPs in human leukemic cell lines
We analyzed the methylated status of SFRP1, SFRP2, SFRP4, and SFRP5 promoter regions in seven human AML cell lines (SHI-1, THP-1, U937, HEL, HL60, K562, and NB4) (Figure 1). SFRP1 promoter was fully methylated in five cell lines (THP-1, HEL, HL60, K562, and NB4), SFRP2 promoter was fully methylated in three cell lines (HEL, K562, and NB4) and partially methylated in three cell lines (HL60, U937, and THP-1), SFRP4 promoter was fully methylated in two lines (THP-1 and K562), and SFRP5 promoter was fully methylated in three lines (SHI-1, U937, and NB4).
 | Figure 1 Electrophoresis results of RQ-MSP products in leukemic cell lines. |
Methylation of SFRPs in AML patients
An NM-SFRP1/2/4/5 ratio equal to or above the values (determined as the mean +5 standard deviation) was set to define high SFRPs methylation in non-M3 AML samples. SFRPs methylation was found in none of the 21 (0%) controls. The frequencies of aberrant methylation among AML patients were as follows: 30.2% (42/139) for SFRP1, 27.3% (38/139) for SFRP2, 5.0% (7/139) for SFRP4, and 1.4% (2/139) for SFRP5. Hypermethylation of at least one SFRP gene occurred in 51.8% (72/139) of AML patients, and there was significant difference between controls and AML patients (P<0.001). The representative results of RQ-MSP products are shown in Figure 2.
 | Figure 2 Electrophoresis results of RQ-MSP products of SFRPs in AML patients. |
Association between SFRPs hypermethylation and clinical characteristics
There was no significant difference in the sex, blood parameters, FAB subtypes, and cytogenetic risks between the methylated status of any SFRP gene with the clinical parameters (P>0.05; Table 1). However, the SFRP1 hypermethylated patients had higher incidence of N/K-RAS mutations compared to those without SFRP1 hypermethylation (P=0.043), whereas the frequency of SFRPs methylation was higher in patients ≥50 y compared to those <50 y, especially for SFRP2 (P<0.05).
Association between SFRPs hypermethylation and prognosis
The follow-up data were obtained for 85 non-M3 AML patients. The median follow-up duration of the patients was 6 months (range: 0–79 months). There was no significant difference in the rates of complete remission after induction therapy between the patients with SFRPs methylation and those without SFRPs methylation (P=0.292; Table 1). However, the Kaplan–Meier curve showed that the OS time of high SFRPs-methylated AML was shorter than that of low SFRPs-methylated group (median: 5 months vs 7.5 months, P=0.036; Figure 3A). Furthermore, among cytogenetically normal AML (CN-AML) patients, cases with high SFRPs methylation also had significantly shorter OS than those with low methylation (median: 4 months vs 8 months, P=0.035; Figure 3B). We further performed a Cox regression multivariate analysis adjusting for prognosis-related factors (age, white blood cells, and karyotype), and revealed that SFRPs hypermethylation acts as an independent prognostic biomarker among both non-M3 AML (HR =1.804, P=0.026) and CN-AML (HR =2.477, P=0.023) patients.
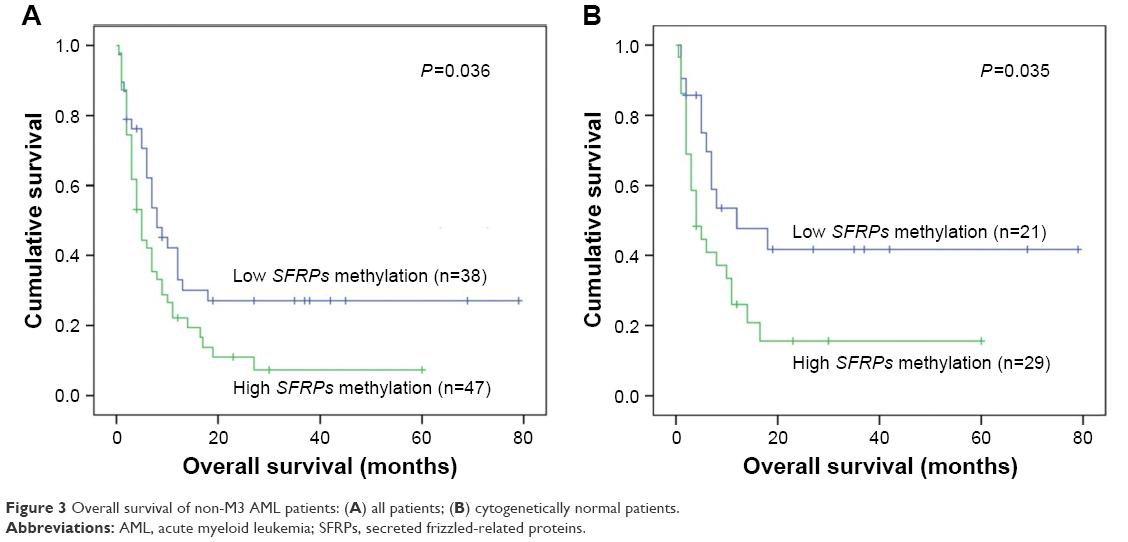 | Figure 3 Overall survival of non-M3 AML patients: (A) all patients; (B) cytogenetically normal patients. |
Effect of 5-aza-dC treatment on SFRPs methylation and expression in leukemic cell lines
To further verify the influence of methylation on SFRPs expression, the HL60 cell line was treated with 5-aza-dC for 72 h. As shown in Figure 4, SFRP1/2 expression was significantly upregulated after 5-aza-dC treatment, and the level of SFRP1/2 promoter methylation decreased. Moreover, there was an inverse correlation between SFRP1/2 hypermethylation and the expression level in the HL60 cell line (r=−0.975, P=0.005 and r=−0.975, P=0.005, respectively).
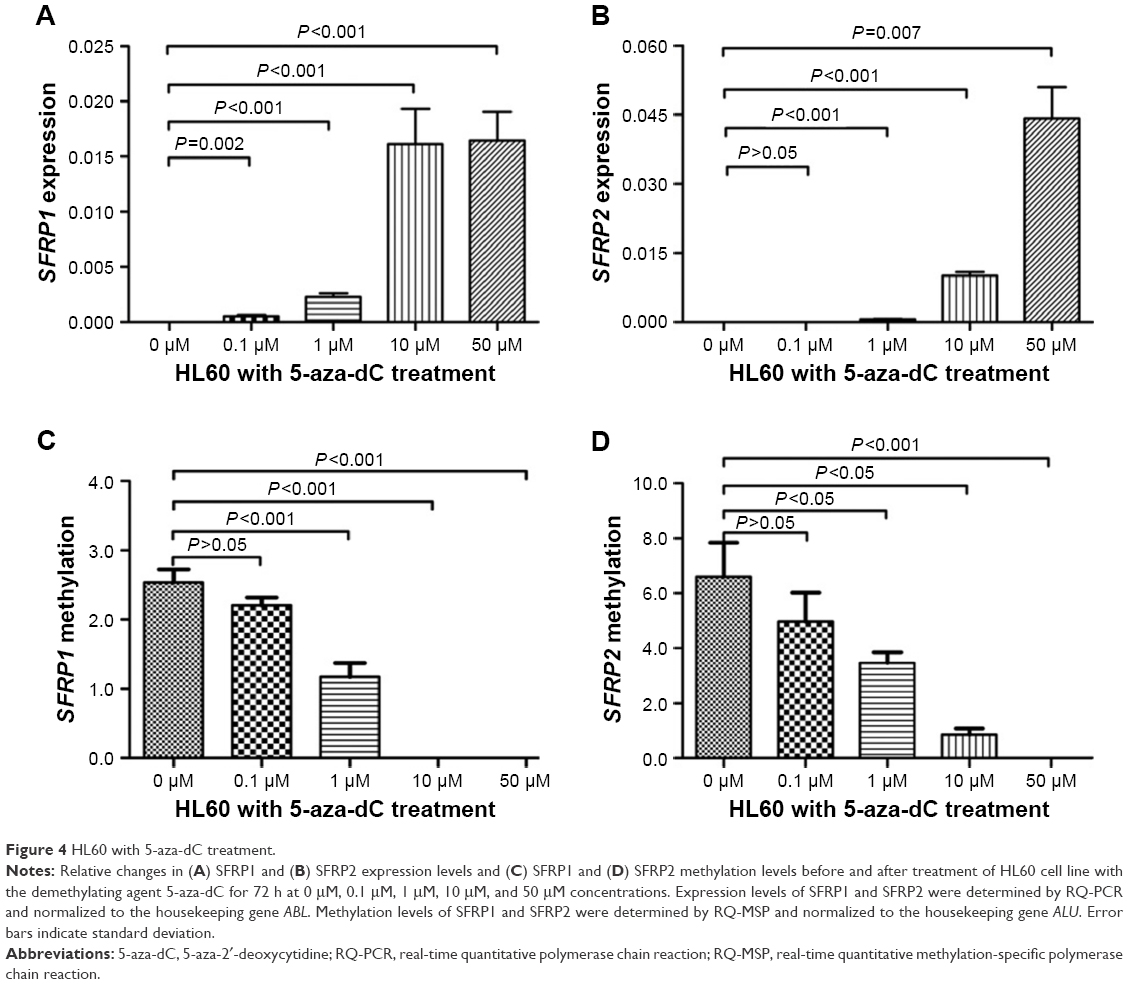 | Figure 4 HL60 with 5-aza-dC treatment. |
Correlation between expression and methylation of SFRPs in AML patients
The expression levels of SFRP1 and SFRP2 were examined in 39 non-M3 AML patients with available cDNA specimens. There was no significant correlation between SFRP2 expression and methylation (r=−0.171, P=0.297). However, a tendency of negative correlation was observed between SFRP1 expression and its promoter methylation (r=−0.334, P=0.038; Figure 5).
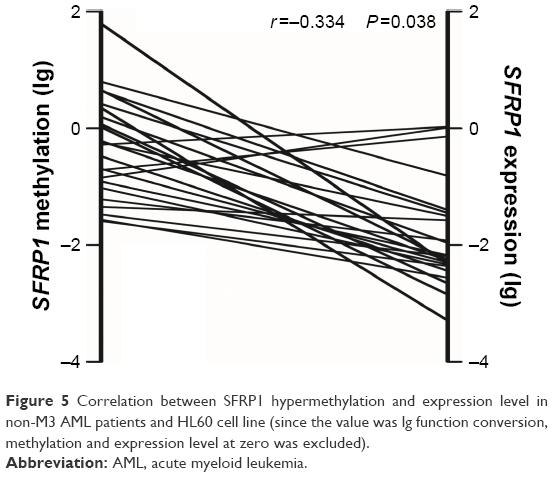 | Figure 5 Correlation between SFRP1 hypermethylation and expression level in non-M3 AML patients and HL60 cell line (since the value was lg function conversion, methylation and expression level at zero was excluded). |
Discussion
Wnt signaling pathway has a crucial role in extensive cellular processes of differentiation and proliferation of hematopoietic progenitor. The aberrant activation of this pathway contributes to leukemogenesis. To further verify the role of epigenetic dysregulation of Wnt antagonists SFRPs in leukemia, our study used quantitative methodology to explore significant methylated status of SFRPs promoter in non-M3 AML patients.
In this study, we investigated the correlations of hypermethylation of SFRPs gene promoter with clinical features in non-M3 Chinese AML patients. Previous studies have estimated the frequency of methylation events in AML patients (SFRP1: 29%–41%, SFRP2: 17%–66%, SFRP4: 0%–9%, and SFRP5: 3%–54%).28,30,31,43,44 However, Shen et al found that the frequency of promoter methylation for SFRP1/2/4/5 genes was 33.9%, 25.4%, 6.8%, and 11.9%, respectively.45 From our results, we found that the frequency of promoter methylation was quite close to the result obtained by Shen et al. It can be seen that different races may result in different outcomes. Moreover, we used RQ-MSP, which belongs to one of the semi-quantitative methodologies that provides an advantage for PCR-based assays with the use of melting curve analysis (MCA) minimizing the inclusion of false positives.46,47 RQ-MSP was able to provide an important consideration for identifying significant methylation events and eliminating very low levels of methylation that are unlikely to confer biological impact. Thus, it can be seen that variability could be caused by methodological differences. Moreover, Cheng et al found that the frequency of aberrant SFRPs promoter methylation correlated significantly with an increased age.44 Similarly, our study also proved the fact that increase in age is associated with the increased frequency of SFRPs methylation.
Furthermore, for the clinical and hematological parameters, previous studies have found the significance of gene mutations including CEBPA and N/K-RAS. Cheng et al found the association between SFRP1 hypermethylation and CEBPA mutations,44 but we did not identify any correlation between SFRP1 methylation and CEBPA mutations. Nevertheless, Takeda et al observed that methylation of SFRP2 promoter occurred more frequently in K-RAS-mutated colorectal cancers,48 yet our data showed the potential association between SFRP1 hypermethylation and N/K-RAS mutations. The discrepancies in results may also have been caused by the difference in races and number of samples Thus, further studies are required to confirm the results and identify the underlying molecular mechanism.
Further studies should analyze the association between SFRP1/2 hypermethylation and transcriptional downregulation, which could be a potential mechanism leading to the abnormal Wnt signaling pathway. In our study, we have confirmed that aberrant CpG island methylation of SFRP1/2 near the transcriptional start site is associated with gene silencing in the HL60 cell line. To further confirm the role of the epigenetic dysregulation of Wnt antagonists SFRPs in leukemia, we examined the specimens of non-M3 AML patients. Jost et al demonstrated that promoter hypermethylation of SFRP2 was associated with transcriptional downregulation in primary AML.30 However, in our study, we found that there was no significant correlation between SFRP2 expression and methylation. In contrast, there was an association between SFRP1 hypermethylation and transcriptional downregulation in non-M3 AML. Similarly, Reins et al demonstrated a significant transcriptional downregulation of SFRP1 in myelodysplastic syndrome.49 Jost et al confirmed the frequent epigenetic dysregulation of the Wnt antagonists SFRP1/2/4/5 by aberrant promoter hypermethylation in multiple myeloma.50 Thus, our data supported the hypothesis that the epigenetic dysregulation could contribute to the abnormal Wnt signaling pathway, which may be related to AML. Therefore, we need further studies to analyze in detail the functional role of SFRPs.
It is worth noting that hypermethylation of SFRPs predicts poor prognosis in non-M3 AML. Numerous studies have indicated that promoter methylation of SFRPs and other Wnt modulators carried prognostic implications for AML.28,30,31 For instance, Griffiths et al found that SFRP2/5 methylations were associated with poorer disease-free survival and OS in CN-AML.31 Jost et al reported that SFRP2 methylation was an adverse risk factor for survival in core binding factor AML.30 Although our results have not found a relationship between the hypermethylation of a single member of SFRPs and a prognosis, we confirmed that the OS time of the high SFRPs-methylated group in non-M3 AML was shorter than that of the low group. Furthermore, among CN-AML patients, cases with the high SFRPs-methylated group also had significantly shorter OS than those with low SFRPs-methylated one. Similarly, Valencia et al showed that the methylation of SFRPs and DKKs was associated with a poorer prognosis only in young adult patients with intermediate-risk cytogenetics.28 Our results were consistent with the above findings. Therefore, we indicated that the hypermethylation of SFRPs predicts poor prognosis and may be useful as a biomarker for further prognostic stratification in cytogenetically defined risk groups in non-M3 AML.
Quantitative methodology may be an important consideration for identifying significant methylation events. Thus, we used a quantitative methodology to analyze hypermethylation of SFRPs and arrived at an important conclusion that promoter hypermethylation of SFRP1/2 appears to be a common event and may have a poor impact on the prognosis in non-M3 AML.
Conclusion
Both whole cohort and CN patients with high SFRPs-methylated group showed a shorter OS compared to those with low group. The alteration of SFRP1/2 expression was inversely correlated with change of SFRP1/2 methylation in leukemic cell line HL60. These findings suggested that the hypermethylation of SFRPs promoter was an adverse risk factor for survival in non-M3 AML.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (81270630), Special Funds of “Ke Jiao Qiang Wei” Project of Jiangsu Province, 333 Project of Jiangsu Province (BRA2016131), Six Talent Peaks Project in Jiangsu Province (2015-WSN-115), China Postdoctoral Science Foundation-funded project (2016M601748), Social Development Foundation of Zhenjiang (SH2015058, SH2016045, SH2016046), Social Development Foundation of Kunshan (KS1624), and the Key Medical Talent Program of Zhenjiang City. The authors are very grateful to Dong-ming Yao (Affiliated People’s Hospital of Jiangsu University) for providing technical assistance for the study.
Disclosure
The authors report no conflicts of interest in this work.
References
Estey E, Dohner H. Acute myeloid leukaemia. Lancet. 2006;368(9550):1894–1907. | ||
Lane SW, Scadden DT, Gilliland DG. The leukemic stem cell niche: current concepts and therapeutic opportunities. Blood. 2009;114(6):1150–1157. | ||
Huntly BJP, Gilliland DG. Cancer biology: summing up cancer stem cells. Nature. 2005;435(7046):1169–1170. | ||
Jordan CT, Guzman ML, Noble M. Cancer stem cells. N Engl J Med. 2006;355(12):1253–1261. | ||
Kirwan M, Vulliamy T, Marrone A, et al. Defining the pathogenic role of telomerase mutations in myelodysplastic syndrome and acute myeloid leukemia. Hum Mutat. 2009;30(11):1567–1573. | ||
Slovak ML, Kopecky KJ, Cassileth PA, et al. Karyotypic analysis predicts outcome of preremission and postremission therapy in adult acute myeloid leukemia: a Southwest Oncology Group/Eastern Cooperative Oncology Group Study. Blood. 2000;96(13):4075–4083. | ||
Byrd JC, Mrozek K, Dodge RK, et al. Pretreatment cytogenetic abnormalities are predictive of induction success, cumulative incidence of relapse, and overall survival in adult patients with de novo acute myeloid leukemia: results from Cancer and Leukemia Group B (CALGB 8461). Blood. 2002;100(13):4325–4336. | ||
Nguyen TT, Mohrbacher AF, Tsai YC, et al. Quantitative measure of c-abl and p15 methylation in chronic myelogenous leukemia: biological implications. Blood. 2000;95(9):2990–2992. | ||
Jones PA, Baylin SB. The fundamental role of epigenetic events in cancer. Nat Rev Genet. 2002;3(6):415–428. | ||
Esteller M. Profiling aberrant DNA methylation in hematologic neoplasms: a view from the tip of the iceberg. Clin Immunol. 2003;109(1):80–88. | ||
Galm O, Herman JG, Baylin SB. The fundamental role of epigenetics in hematopoietic malignancies. Blood Rev. 2006;20(1):1–13. | ||
Kostroma II, Gritsaev SV, Sidorova ZhY, et al. Aberrant methylation of the promoter regions of the SOX7 and p15INK4b genes and Wnt signaling pathway antagonists in patients with acute myeloid leukemias. Ter Arkh. 2016;88(7):31–36. | ||
Mikryukov A, Moss T. Agonistic and antagonistic roles for TNIK and MINK in non-canonical and canonical Wnt signalling. PLoS One. 2012;7(9):e43330. | ||
Kirstetter P, Anderson K, Porse BT, Jacobsen SE, Nerlov C. Activation of the canonial Wnt pathway leads to loss of hematopoietic stem cell repopulation and multilineage differentiation block. Nat Immunol. 2006;7(10):1048–1056. | ||
Xavier CP, Melikova M, Chuman Y, Uren A, Baljinnyam B, Rubin JS. Secreted Frizzled-related protein potentiation versus inhibition of Wnt3a/β−catenin signaling. Cell Signal. 2014;26(1):94–101. | ||
Malbon CC. Frizzleds: new members of the superfamily of G-protein-coupled receptors. Front Biosci. 2004;9:1048–1058. | ||
Wang HY, Malbon CC. Wnt-Frizzled signaling to G-protein-coupled effectors. Cell Mol Life Sci. 2004;61(1):69–75. | ||
Khan NI, Bendall LJ. Role of WNT signaling in normal and malignant hematopoiesis. Histol Histopathol. 2006;21(7):761–774. | ||
Staal FJ, Clevers HC. WNT signalling and haematopoiesis: a WNT-WNT situation. Nat Rev Immunol. 2005;5(1):21–30. | ||
Reya T, Duncan AW, Ailles L, et al. A role for Wnt signalling in self-renewal of haematopoietic stem cells. Nature. 2003;423(6938):409–414. | ||
Rattis FM, Voermans C, Reya T. Wnt signaling in the stem cell niche. Curr Opin Hematol. 2004;11(2):88–94. | ||
Clevers H, Nusse R. Wnt/β-catenin signaling and disease. Cell. 2012;149(6):1192–1205. | ||
Kohno H, Amatya VJ, Takeshima Y, et al. Aberrant promoter methylation of WIF-1 and SFRP1, 2, 4 genes in mesothelioma. Oncol Rep. 2010;24(2):423–431. | ||
Guo Y, Guo W, Chen Z, Kuang G, Yang Z, Dong Z. Hypermethylation and aberrant expression of Wnt-antagonist family genes in gastric cardia adenocarcinoma. Neoplasma. 2011;58(2):110–117. | ||
Chim CS, Pang R, Fung TK, Choi CL, Liang R. Epigenetic dysregulation of Wnt signaling pathway in multiple myeloma. Leukemia. 2007;21(12):2527–2536. | ||
Kim JT, Li J, Jang ER, et al. Deregulation of Wnt/β-catenin signaling through genetic or epigenetic alterations in human neuroendocrine tumors. Carcinogenesis. 2013;34(5):953–961. | ||
Suzuki H, Watkins DN, Jair KW, et al. Epigenetic inactivation of SFRP genes allows constitutive WNT signaling in colorectal cancer. Nat Genet. 2004;36(4):417–422. | ||
Valencia A, Roman-Gomez J, Cervera J, et al. Wnt signaling pathway is epigenetically regulated by methylation of Wnt antagonists in acute myeloid leukemia. Leukemia. 2009;23(9):1658–1666. | ||
Figueroa ME, Skrabanek L, Li Y, et al. MDS and secondary AML display unique patterns and abundance of aberrant DNA methylation. Blood. 2009;114(16):3448–3458. | ||
Jost E, Schmid J, Wilop S, et al. Epigenetic inactivation of secreted Frizzled-related proteins in acute myeloid leukaemia. Br J Haematol. 2008;142(5):745–753. | ||
Griffiths EA, Gore SD, Hooker C, et al. Acute myeloid leukemia is characterized by Wnt pathway inhibitor promoter hypermethylation. Leuk Lymphoma. 2010;51(9):1711–1719. | ||
Bennett JM, Catovsky D, Daniel MT, et al. Proposed revised criteria for the classification of acute myeloid leukaemia. A report of the French-American-British Cooperative Group. Ann Intern Med. 1985;103(4):620–625. | ||
Swerdlow SH, Campo E, Harris NL, et al. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues. Lyon: IARC Press; 2008. | ||
Grimwade D, Hills RK, Moorman AV, et al. Refinement of cytogenetic classification in acute myeloid leukemia: determination of prognostic significance of rare recurring chromosomal abnormalities among 5876 younger adult patients treated in the United Kingdom Medical Research Council trials. Blood. 2010;116(3):354–365. | ||
Yao DM, Qian J, Lin J, et al. Aberrant methylation of CCAAT/enhancer binding protein zeta promoter in acute myeloid leukemia. Leuk Res. 2011;35(7):957–960. | ||
Guo H, Lin J, Wen XM, et al. Decreased SFRP2 expression is associated with intermediate and poor karyotypes in de novo acute myeloid leukemia. Int J Clin Exp Pathol. 2014;7(8):4695–4703. | ||
Qian J, Zhu ZH, Lin J, et al. Hypomethylation of PRAME promoter is associated with poor prognosis in myelodysplastic syndrome. Br J Haematol. 2011;154(1):153–155. | ||
Lin J, Yao DM, Qian J, et al. Recurrent DNMT3A R882 mutations in Chinese patients with acute myeloid leukemia and myelodysplastic syndrome. PLoS One. 2011;6(10):e26906. | ||
Lin J, Yao DM, Qian J, et al. IDH1 and IDH2 mutation analysis in Chinese patients with acute myeloid leukemia and myelodysplastic syndrome. Ann Hematol. 2012;91(4):519–525. | ||
Yang X, Qian J, Sun A, et al. RAS mutation analysis in a large cohort of Chinese patients with acute myeloid leukemia. Clin Biochem. 2013;46(7–8):579–583. | ||
Wen XM, Lin J, Yang J, et al. Double CEBPA mutations are prognostically favorable in non-M3 acute myeloid leukemia patients with wild-type NPM1 and FLT3-ITD. Int J Clin Exp Pathol. 2014;7(10):6832–6840. | ||
Kottaridis PD, Gale RE, Frew ME, et al. The presence of a FLT3 internal tandem duplication in patients with acute myeloid leukemia (AML) adds important prognostic information to cytogenetic risk group and response to the first cycle of chemotherapy: analysis of 854 patients from the United Kingdom Medical Research Council AML 10 and 12 trials. Blood. 2001;98(6):1752–1759. | ||
Ghasemi A, Rostami S, Chahardouli B, Alizad Ghandforosh N, Ghotaslou A, Nadali F. Study of SFRP1 and SFRP2 methylation status in patients with de novo acute myeloblastic leukemia. Int J Hematol Oncol Stem Cell Res. 2015;9(1):15–21. | ||
Cheng CK, Li L, Cheng SH, et al. Secreted-frizzled related protein 1 is a transcriptional repression target of the t(8;21) fusion protein in acute myeloid leukemia. Blood. 2011;118(25):6638–6648. | ||
Shen JZ, Xu CB, Fu HY, Wu DS, Zhou HR, Fan LP. Methylation of secreted frizzled related protein gene in acute leukemia patients in China. Asian Pac J Cancer Prev. 2011;12(10):2617–2621. | ||
Kristensen LS, Mikeska T, Krypuy M, Dobrovic A. Sensitive Melting Analysis after Real Time-Methylation Specific PCR (SMART-MSP): high-throughput and probe-free quantitative DNA methylation detection. Nucleic Acids Res. 2008;36(7):e42. | ||
Candiloro IL, Mikeska T, Dobrovic A. Assessing combined methylationsensitive high resolution melting and pyrosequencing for the analysis of heterogeneous DNA methylation. Epigenetics. 2011;6(4):500–507. | ||
Takeda M, Nagasaka T, Dong-Sheng S, et al. Expansion of CpG methylation in the SFRP2 promoter region during colorectal tumorigenesis. Acta Med Okayama. 2011;65(3):169–177. | ||
Reins J, Mossner M, Neumann M, et al. Transcriptional down-regulation of the Wnt antagonist SFRP1 in haematopoietic cells of patients with different risk types of MDS. Leuk Res. 2010;34(12):1610–1616. | ||
Jost E, Gezer D, Wilop S, et al. Epigenetic dysregulation of secreted Frizzled-related proteins in multiple myeloma. Cancer Lett. 2009;281(1):24–31. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.